DOKTORI (PhD) ÉRTEKEZÉS
GYENES VIKTOR
KESZTHELY
2010
PANNON EGYETEM GEORGIKON KAR NÖVÉNYVÉDELMI INTÉZET
HERBOLÓGIAI ÉS NÖVÉNYVÉDŐSZER KÉMIAI OSZTÁLY
ÁLLAT- ÉS AGRÁRKÖRNYEZET-TUDOMÁNYI DOKTORI ISKOLA
ISKOLAVEZETŐ: D
DRR.. hhaabbiill.. AANNDDAA AANNGGÉÉLLAA Egyetemi tanár
TÉMAVEZETŐ: D
DRR.. hhaabbiill.. BBÉÉRREESS IIMMRREE Professor emeritus
A PONGYOLA PITYPANG
(TARAXACUM OFFICINALETARAXACUM OFFICINALETARAXACUM OFFICINALETARAXACUM OFFICINALE G. H. WEBER EX WIGGERS)
JELENTŐSÉGE, BIOLÓGIÁJA ÉS INTEGRÁLT GYOMSZABÁLYOZÁSA
KÉSZÍTETTE: GYENES VIKTOR
KESZTHELY
2010
A PONGYOLA PITYPANG
(TARAXACUM OFFICINALETARAXACUM OFFICINALETARAXACUM OFFICINALETARAXACUM OFFICINALE G. H. WEBER EX WIGGERS)
JELENTŐSÉGE, BIOLÓGIÁJA ÉS INTEGRÁLT GYOMSZABÁLYOZÁSA ÉRTEKEZÉS DOKTORI (PhD) FOKOZAT ELNYERÉSE ÉRDEKÉBEN
ÍRTA: GGYEYENNEESS VVIIKKTTOORR
KÉSZÜLT A PANNON EGYETEM
ÁLLAT- ÉS AGRÁRKÖRNYEZET-TUDOMÁNYI DOKTORI ISKOLA KERETÉBEN
TÉMAVEZETŐ:DDRR..BBÉÉRREESS IIMRMREE
ELFOGADÁSRA JAVASLOM (IGEN / NEM). ...
ALÁÍRÁS
A JELÖLT A DOKTORI SZIGORLATON …...%-OT ÉRT EL.
KESZTHELY,2007. OKTÓBER 24. ...
A SZIGORLATI BIZOTTSÁG ELNÖKE
AZ ÉRTEKEZÉST BÍRÁLÓKÉNT ELFOGADÁSRA JAVASLOM:
BÍRÁLÓ NEVE:………..…… IGEN / NEM
...
ALÁÍRÁS
BÍRÁLÓ NEVE:………..…… IGEN / NEM
...
ALÁÍRÁS
A JELÖLT AZ ÉRTEKEZÉS NYILVÁNOS VITÁJÁN ……%-OT ÉRT EL. KESZTHELY,2010.………..
...
A BÍRÁLÓ BIZOTTSÁG ELNÖKE
A DOKTORI (PhD) OKLEVÉL MINŐSÍTÉSE:………..
...
AZ EDT ELNÖKE
TARTALOMJEGYZÉK
I KIVONATOK--- 1 -
I.1 MAGYAR NYELVŰ KIVONAT--- -1-
I.2 ANGOL NYELVŰ KIVONAT – ABSTRACT--- -2-
I.3 SPANYOL NYELVŰ KIVONAT – EXTRACTO--- -2-
II BEVEZETÉS--- 3 -
III IRODALMI ÁTTEKINTÉS--- 6 -
III.1 EREDETE, ELNEVEZÉSE--- -6-
III.2 RENDSZERTANI BESOROLÁSA, JELLEMZÉSE, VÁLTOZATAI--- -6-
III.2.1 A Taraxacum nemzetség rendszertana--- 6 -
III.2.2 Morfológiája--- - 10 -
III.2.3 Más fajokkal való rokonsága, változékonysága--- - 12 -
III.3 GAZDASÁGI JELENTŐSÉGE---12-
III.3.1 Kártétele--- - 12 -
III.3.2 Haszna--- - 13 -
III.4 FÖLDRAJZI ELTERJEDÉSE---14-
III.5 ELŐFORDULÁSI HELY---14-
III.5.1 Klimatikus igénye--- - 14 -
III.5.2 Talaj igényei--- - 15 -
III.5.3 Előfordulása növénytársulásokban--- - 15 -
III.6 NÖVEKEDÉSE ÉS FEJLŐDÉSE---15-
III.6.1 Morfológiája--- - 15 -
III.6.2 Fiziológiája--- - 16 -
III.6.3 Fenológiája--- - 18 -
III.7 SZAPORODÁSA---19-
III.7.1 Virágzás--- - 19 -
III.7.2 Kaszatprodukció és terjesztése--- - 19 -
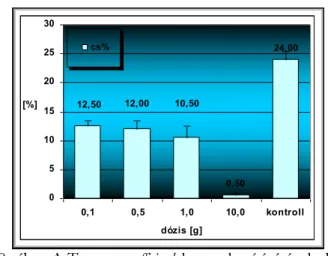
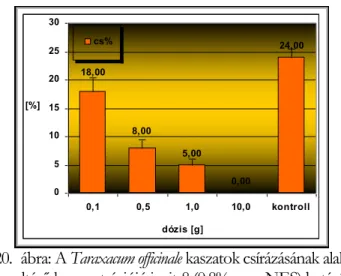
III.7.3 Kaszatok életképessége és csírázása--- - 20 -
III.7.4 Vegetatív szaporodása--- - 21 -
III.8 POPULÁCIÓ DINAMIKA---22-
III.9 INTEGRÁLT GYOMSZABÁLYOZÁSÁNAK MÓDSZEREI---22-
III.9.1 Kémiai módszerek--- - 22 -
III.9.2 Mechanikai és agrotechnikai módszerek--- - 24 -
III.9.3 Fizikai módszerek--- - 25 -
III.9.4 Biológiai módszerek – növényevők, betegségek, paraziták--- - 26 -
IV ANYAG ÉS MÓDSZER--- - 29 -
IV.1 LABORATÓRIUMI VIZSGÁLATOK---29-
IV.1.1 Csírázásbiológiai vizsgálatok--- - 29 -
IV.1.1.1 Magnyugalom (primer dormancia) vizsgálata--- 29 -
IV.1.1.2 Éves csírázási ritmus vizsgálata--- 29 -
IV.1.1.3 Magkórtani vizsgálatok--- 29 -
IV.1.1.4 Fény szerepe a kaszatok csírázásában--- 30 -
IV.1.1.5 NaCl (sóstressz) vizsgálata a kaszatok csírázásában--- 30 -
IV.1.1.6 Ozmotikus potenciál (szárazságstressz) vizsgálata a kaszatok csírázásában--- 30 -
IV.1.1.7 Allelopátiás vizsgálatok--- 31 -
IV.1.1.8 Hőmérséklet hatása a kaszatok csírázására--- 31 -
IV.1.1.9 Növekedési hormonok hatása a kaszatok csírázásában--- 31 -
IV.1.2 Regenerálódás vizsgálat--- - 32 -
IV.1.2.1 Regenerálódó képesség éves ritmusának vizsgálata--- 32 -
IV.1.2.2 A Taraxacum officinale gyökerek regenerálódó képessége--- 32 -
IV.1.3 Tápelem felvétel vizsgálat--- - 32 -
IV.1.3.1 NPK koncentráció tartalom éves ritmusának vizsgálata--- 32 -
IV.1.4 Genetikai vizsgálatok--- - 32 -
IV.1.4.1 A Keszthely térségi pongyola pitypang populáció polimorfizmus vizsgálata--- 32 -
IV.2 ÜVEGHÁZI TENYÉSZEDÉNYES VIZSGÁLATOK---34-
IV.2.1 Csírázásbiológiai vizsgálatok--- - 34 -
IV.2.1.1 Talajmélység hatása a kaszatok csírázására--- 34 -
IV.2.2 Növekedés analízis--- - 35 -
IV.2.2.1 Növekedés analízis vizsgálat--- 35 -
IV.2.3 Kompetíciós képesség vizsgálata--- - 35 -
IV.2.3.1 Kompetíciós képesség vizsgálata lucerna és angolperje állományban--- 35 -
IV.2.4 Regenerálódás vizsgálat--- - 36 -
IV.2.4.1 Sekély kapálás (1 cm) hatása a Taraxacum officinale regenerálódó képességére--- 36 -
IV.2.5 Herbicidérzékenységi vizsgálatok--- - 37 -
IV.2.5.1 Preemergens kezelések--- 37 -
IV.2.5.2 Posztemergens kezelések--- 37 -
IV.3 SZABADFÖLDI VIZSGÁLATOK---37-
IV.3.1 Herbicid érzékenységi vizsgálatok--- - 37 -
IV.3.1.1 Posztemergens kezelések--- 37 -
IV.4 EREDMÉNYEK STATISZTIKAI ELEMZÉSE---38-
V EREDMÉNYEK ÉS KÖVETKEZTETÉSEK--- - 39 -
V.1 LABORATÓRIUMI VIZSGÁLATOK---39-
V.1.1 Csírázásbiológiai vizsgálatok--- - 39 -
V.1.1.1 Magnyugalom (primer dormancia) vizsgálata--- 39 -
V.1.1.2 Éves csírázási ritmus vizsgálata--- 39 -
V.1.1.3 Magkórtani vizsgálatok--- 41 -
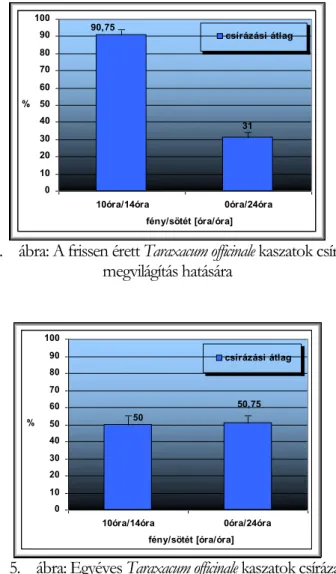
V.1.1.4 Fény szerepe a kaszatok csírázásában--- 42 -
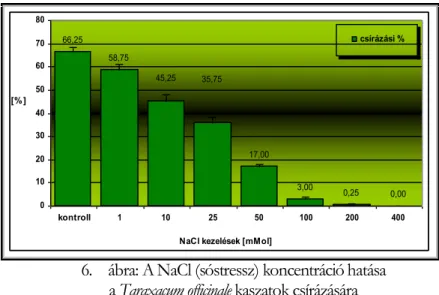
V.1.1.5 A NaCl (sóstressz) vizsgálata a kaszatok csírázásában--- 44 -
V.1.1.6 Az ozmotikus potenciál (szárazságstressz) vizsgálata a kaszatok csírázásában--- 44 -
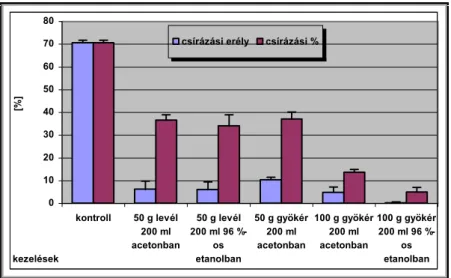
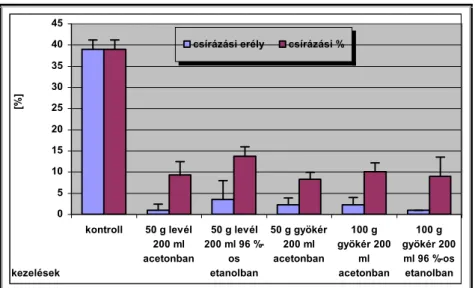
V.1.1.7 Allelopátiás vizsgálatok--- 45 -
V.1.1.8 Hőmérséklet hatása a kaszatok csírázására--- 53 -
V.1.1.9 Növekedési hormonok hatása a kaszatok csírázásában--- 55 -
V.1.2 Regenerálódás vizsgálatok--- - 57 -
V.1.2.1 Regenerálódó képesség éves ritmusának vizsgálata--- 57 -
V.1.2.2 A Taraxacum officinale gyökerek regenerálódó képessége--- 59 -
V.1.3 Tápelem felvétel vizsgálat--- - 69 -
V.1.3.1 NPK koncentráció tartalom éves ritmusának vizsgálata--- 69 -
V.1.4 Genetikai vizsgálatok--- - 71 -
V.1.4.1 A hazai pongyola pitypang populáció polimorfizmus vizsgálata--- 71 -
V.2 ÜVEGHÁZI TENYÉSZEDÉNYES VIZSGÁLATOK---73-
V.2.1 Csírázásbiológiai vizsgálatok--- - 73 -
V.2.1.1 A talajmélység hatása a kaszatok csírázásra--- 73 -
V.2.2 Növekedés analízis--- - 75 -
V.2.2.1 Őszi kelésű pongyola pitypang növekedés analízis vizsgálata--- 75 -
V.2.3 Kompetíció--- - 80 -
V.2.3.1 Kompetíciós képesség vizsgálata lucerna és angolperje állományban--- 80 -
V.2.4 Regenerálódás vizsgálat--- - 87 -
V.2.4.1 Sekély kapálás (1 cm) hatása a Taraxacum officinale regenerálódó képességére--- 87 -
V.2.5 Herbicidérzékenységi vizsgálatok--- - 89 -
V.2.5.1 Preemergens kezelések--- 89 -
V.2.5.2 Posztemergens kezelések--- 89 -
V.3 SZABADFÖLDI VIZSGÁLATOK---91-
V.3.1 Herbicidérzékenységi vizsgálatok--- - 91 -
V.3.1.1 Posztemergens kezelések--- 91 -
VI ÖSSZEFOGLALÁS--- - 94 -
VII IRODALOMJEGYZÉK--- - 98 -
VIII KÖSZÖNETNYILVÁNÍTÁS--- 111 -
IX AZ ÉRTEKEZÉS TÉZISEI--- 112 -
IX.1 MAGYAR NYELVŰ TÉZISPONTOK--- -112-
IX.2 ANGOL NYELVŰ TÉZISPONTOK--- -113-
I KIVONATOK
I.1 MAGYAR NYELVŰ KIVONAT
A disszertáció célja, hogy a Taraxacum officinale hazai populációjával kapcsolatos eddigi ismereteinket bővítsük, további értékes és hasznos adatokat nyerjünk, melyek ismeretében könnyebben tudjuk kialakítani az integrált gyomszabályozás minden elemét.
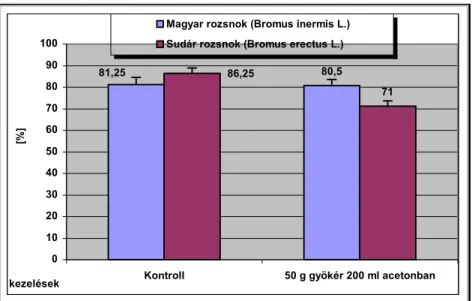
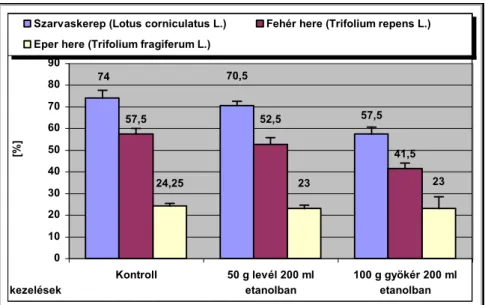
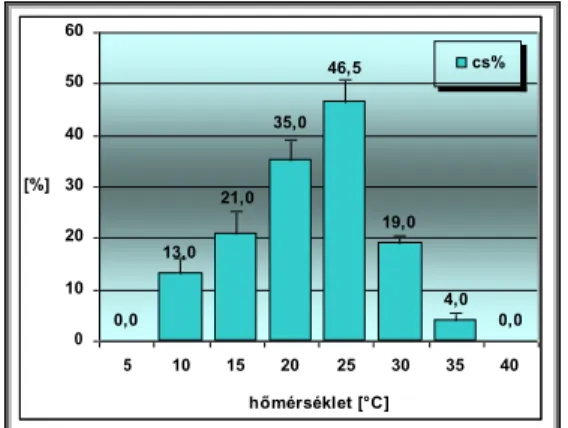
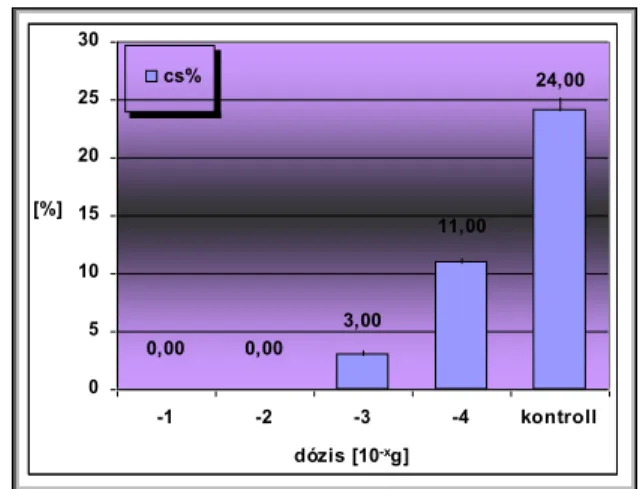
Csírázásbiológiai vizsgálataink során megállapítottuk, hogy a frissen érett kaszatoknak nincs magnyugalma. A kaszatok egyenletesen és igen magas %-ban csíráznak, azonban 2 év eltelte után csírázóképességük jelentősen csökken. A fénynek csírázás növelő hatását bizonyítottuk. A NaCl (sóstressz) koncentráció emelkedésével csökken a kaszatok csírázóképessége, majd végül megszűnik. Az ozmotikus potenciál (szárazságstressz) növekedésével szintén csökken a kaszatok csírázóképessége, majd megszűnik. A kaszatok 5°C és az alatti valamint 40°C és annál magasabb hőmérsékleten már nem csíráznak. Növekedési hormonok (naftil-ecetsav, indolil-vajsav) kis mennyiségben növelik a kaszatok csírázóképességét, nagy mennyiségben pedig csökkentik azt. A Taraxacum officinale gyökerének és hajtásának acetonos és etanolos kivonata jelentős mértékben csökkenti számos fűfaj (Lolium perenne, Arrhenatherum elatius, Dactylis glomerata, Poa pratensis, Festuca rubra, Bromus inermis, Bromus erectus) és pillangós (Lotus corniculatus, Trifolium repens, Trifolium fragiferum) magvainak csírázását. Magkórtani vizsgálataink során számos gomba fajt és nemzetséget figyeltünk meg a kaszatok felületén. A kaszatok tömeges csírázása és kelése a talajfelszínről történik, 5 cm-nél mélyebb rétegekből már nem kelnek ki.
A pongyola pitypang vegetatív szaporodását vizsgálva megállapítottuk, hogy a gyökérnyakhoz közelebbi szegmenseket erőteljesebb hajtásképzés jellemzi, mint amelyek távolabb helyezkednek el. Intenzív regenerálódási periódusát késő ősszel és télen figyeltük meg. A helytelenül elvégzett gyomszabályozás eredményeképpen 14 nap múlva már jelentős hajtástömeget képez. A feldarabolt szegmensek hosszának van jelentős szerepe a hajtásképzésben, átmérőjének nincs.
A pongyola pitypang tápelem vizsgálata során a nitrogén, foszfor és kálium koncentrációjának változását vizsgáltuk egy éven keresztül a gyökerekben és hajtásokban.
Ősszel kelő pongyola pitypang növekedés analízise során vizsgáltuk a növény levélterületét, levélterület indexét (LAI), specifikus levélterületét (SLA), a hajtás friss-, és száraztömeg növekedésének ütemét, a hajtás abszolút növekedési sebesség indexét (AGR), a nettó asszimilációs ráta indexét (NAR) valamint a termésnövekedés sebesség indexét (CGR).
A pongyola pitypang és angolperje közötti kompetíció során megállapítottuk, hogy a gyomnövény mért paramétereiben (gyökér-, és hajtáshossz) jóval alulmaradt a kontrollhoz képest, mint az angolperje ugyanazon tulajdonságait tekintve. Angolperje erősebb kompetítora a pitypangnak, mint fordított esetben.
A pongyola pitypang és lucerna kompetíció eredményeiből megállapítottuk, hogy a gyökér és hajtás hossz, friss-, és száraztömeg változása mindkét tesztnövény estében azonos mértékű volt. A két növény egymásnak egyenlő mértékben kompetítorai.
Az alkalmazott PCR-RFLP módszerrel, a vizsgált fragmentumokon a használt restrikciós enzimekkel nem tudtunk nagy számban polimorfizmusokat kimutatni.
Herbicidérzékenységi vizsgálataink eredményei alapján megállapítottuk, hogy az EPTC, metribuzin és linuron hatóanyagok kiváló hatással vannak a kelő kaszatok ellen. Közepes hatásfokkal bírnak az S- metolaklór és pendimetalin hatóanyagok.
A magról kelő pongyola pitypang ellen kiválóan alkalmazható a 2,4-D aminsó, fluroxipír, glifozát, glufozinát-ammónium valamint klórszulfuron+flupírszulfuron-metil hatóanyag kombináció. Jó eredménnyel használható a tribenuron-metil + tifenszulfuron-metil.
Évelő pongyola pitypang növények ellen kiváló eredménnyel használható a 2,4-D aminsó, fluroxipír, glifozát valamint a glufozinát-ammónium hatóanyag bármelyike.
I.2 ANGOL NYELVŰ KIVONAT – ABSTRACT
IMPORTANCE, BIOLOGY AND INTERGRATED WEED CONTROL OF COMMON DANDELION
(TTTTARAXACUM ARAXACUM ARAXACUM ARAXACUM OFFICINALEOFFICINALEOFFICINALEOFFICINALE G.H.WEBER EX WIGGERS)
We determined that seeds after mature can germinate at a high rate. They have no primary dormancy.
The germination % decreases significantly after 2 years. We demonstrated the increasing effect of light on the germination. The higher is the concentration of sodium chloride (NaCl) the lower is the germination % of seeds. We have the same results of the osmotic potential. Seeds do not germinate below 5°C and over 40°C.
The acetone and ethanol extract of root and shoot of Taraxacum officinale decreases significantly the germination % of several seeds of grass (Lolium perenne, Arrhenatherum elatius, Dactylis glomerata, Poa pratensis, Festuca rubra, Bromus inermis, Bromus erectus) and Fabaceae species (Lotus corniculatus, Trifolium repens, Trifolium fragiferum). On basis of seed pathology examinations we found out several species and genus of fungus on the seeds. Seeds of Taraxacum officinale germinate and emerge from the soil surface. More than 5 cm deep seeds do not germinate even emerge. The regenerative capacity of roots was the highest in autumn and winter and the least was in late spring and summer months. We have examined the change of nitrogen, phosphorus and potassium nutrient concentration in the roots and shoots of common dandelion during a year. We examined leaf area index (LAI), specific leaf area (SLA), absolute growth rate (AGR), net assimilation rate (NAR) and the crop growth rate (CGR). Perennial ryegrass is more competitive than common dandelion. Alfalfa has the same competitive capacity as common dandelion. We determined that EPTC, metribuzin, linuron are the most efficacious against common dandelion at preemergent technology. The 2,4-D aminsalt, fluroxypyr, glyphosate, gluphosinate-ammonium and chlorsulfuron-methyl+thifensulfuron-methyl were the most effective against Taraxacum officinale seedlings. 2,4-D aminsalt, fluroxypyr, glyphosate, gluphosinate- ammonium were the most effective against perennial common dandelion.
I.3 SPANYOL NYELVŰ KIVONAT – EXTRACTO
IMPORTANCIA, BIOLOGÍA Y CONTROL INTEGRADO DE MALHIERBA DE DIENTE DE LEÓN
(TTTTARAXACUM ARAXACUM ARAXACUM ARAXACUM OFFICINALEOFFICINALEOFFICINALEOFFICINALE G.H.WEBER EX WIGGERS)
Hemos determinado que las semillas después de la matura germinan en proporción muy alta. Las semillas no tienen dormancia primaria. La germinación decrece significativamente después de dos años. La germinación % de semillas decrece en concentración grande de cloruro de sodio y potencial de ósmosis. Las semillas no germinan debajo de 5°C y sobre de 40°C. El extracto de acetona y de alcohol de raíz y de brótano de Taraxacum officinale disminuye la germinación de diferentes cespedes (Lolium perenne, Arrhenatherum elatius, Dactylis glomerata, Poa pratensis, Festuca rubra, Bromus inermis, Bromus erectus) y especies de Fabaceae (Lotus corniculatus, Trifolium repens, Trifolium fragiferum). Hemos detectado muchos hongos de diferentes familias sobre las semillas. Major parte de la germinación ocurre entre 0-3 cm de capas del suelo. Más profundo de 5 cm ya no hay germinación. La capacidad regenerativa de las raices fue alta en meses de otoño e invierno. Fue baja en meses de primavera y verano. Examinamos el cambio de tres elementos nutritivos (nitrógeno, fósforo, potasio) de las raices y brotanos de diente de león durante un año. Hemos examinado el índice de área foliar (IAF), área foliar especifíca (AFE), tasa de crecimiento absoluto (TCA), tasa neta de asimilación (TNA) y tasa de crecimiento cultivo (TCC). Césped inglés es más competitivo que diente de león. Alfalfa tiene la misma habilidad competitiva que la hierba. EPTC, metribuzina y linuron fueron más efectivo contra la malhierba en aplicación de pre-emergente. 2,4-D sal amina, fluroxipir, glifosato, glufosinato-amónico, clorsulfuron- metil+tifensulfuron-metil fueron lo mejor contra la plántula de Taraxacum officinale. 2,4-D sal amina, fluroxipir, glifosato y glufosinato-amónico fueron los más efectivos contra la hierba perennial de diente de león.
II BEVEZETÉS
Gyomok okozta károsításról, bántalomról beszélünk, ha egyes növényfajok vagy fajok egy csoportja zavarja az ember termelőtevékenységét, befolyásolja egészségét vagy pihenését (FRYER 1979). A „gyom”
fogalma nem tudományos fogalom, hanem köznyelvi kifejezés, és a növényeknek széles körét értik alatta (WELLS 1978). Nemcsak azokat a növényeket tekintik gyomoknak, amelyek zavarják a mezőgazdasági termelést, hanem azon fajokat is, amelyek csúnyák, szúrósak, tüskések, stb.
A gyomnövények tudományos definícióját tekintve számos próbálkozás történt, azonban több nehézség is felmerült.
A felmerülő nehézségek közül az első a szubjektív álláspont, amely alapján a vegetáció egyes fajait értékeljük. Ez megnehezíti az objektív, minden körülmények között érvényes meghatározást. Az emberek előítéleteik alapján választják ki a vegetáció tagjai közül azokat, amelyeket gyomnak neveznek. A fogalom minden esetben haszontalanságot, nem kívánatos, negatív jelleget takar.
A definíciót megnehezítő második körülmény a közelmúltból ered. Az emberiség, létszámának rohamos emelkedése miatt, igyekszik minél több földterületet művelés alá vonni és így egyre több növénnyel kerül
„konfliktusba”, vagyis egyre több növény kerül a gyomnövények listájára (WELLS 1978).
A gyomnövényeket termőhelyi előfordulásuk szerint is csoportosítjuk. Így beszélünk szántóföldi, kertészeti, ruderális és erdészeti, továbbá vízi gyomokról, vagyis minden növény lehet gyomnövény attól függően, hogy az ember annak tekinti-e vagy sem (WELLS 1978). A gyomnövények definíciójának kérdésével többen is foglalkoztak (RADEMACHER 1948;HARLAN.DEWET 1965;KING 1966;NUMATA 1976;EGGERS- NIEMANN 1980; HOLZNER-NUMATA 1982). A gyom fogalmát tekintve szubjektív és ökológiai meghatározások különböztethetők meg. Hazai szakirodalomban megtalálható szubjektív gyomdefiníciók a következők:
PETHE FERENC (1805): „Dudva alatt most minden olyan palántát értünk, mely az önként termesztett palánta között, a neki szabott helyen magában vadon terem, legyen bár az a leghasznosabb palánta magában”.
BALÁS ÁRPÁD (1876): „Gyom, gaz, dudva alatt olyan növényeket értünk, melyek művelt növényeink között akaratunk ellenére megjelenvén, azokból tért és tápanyagot elfognak és rendszeres növekedésüket gátolják”.
CSERHÁTI SÁNDOR (1899): „Gyomnak mondunk minden olyan növényt, mely akaratunk ellenére fordul elő ott, ahol valamely növényt termelünk. A gyom károsítja a termést, mert csökkenti a hozamot és esetleg rontja a minőséget”.
WÁGNER JÁNOS (1908): Azok a növények, melyek bárhol a mi akaratunk ellenére tenyésznek, melyeket öntudatosan nem vetettünk és amelyek az elvetett növények kárára vannak, legalább annyiban, hogy azt a helyet foglalják el, amelyet egy szándékosan ültetett és jelen céljainkra szolgáló növény elfoglalhatott volna, ha máskülönben nem is mérgesek, nem is ártalmasak: gyomoknak hívjuk”.
UJVÁROSI MIKLÓS (1957): „Általános értelemben azokat a növényeket, amelyek az ősi természetes növényzetben nem fordulnak elő csak kultúrterületeken, vagy az ősi vegetáció tagjai, de kultúrterületeken alkalmazkodásuk következtében teret hódítottak, gyomnak szoktuk nevezni”. „Szántóföldeken gyomnak nevezünk minden növényt, melyet nem vetettünk, hasznot nem hoz és jelenléte káros azzal, hogy a vetett növény elől elfoglalja a helyet vagy felhasználja a talaj tápanyag- és vízkészletét. Ez a meghatározás érvényes a kerti és szőlőkultúrákra is”.
UBRIZSY GÁBOR (1962): „A gyomnövény fogalmának legtágabb értelmezése szerint minden olyan növény, mely mezőgazdasági művelés alatt álló területeken szándékos vetés nélkül lép fel, jelenlétével a műveleti teret és fényt foglal el, felhasználja a talaj tápanyag- és vízkészletét, hasznot nem hoz: gyomnövény”.
HUNYADI KÁROLY (1974): „Gyomnövénynek nevezünk bármely fejlődési stádiumban lévő olyan növényt vagy növényi részt (tarack, rizóma, hagyma, hagymagumó, stb.), amely ott fordul elő, ahol nem kívánatos”.
Az ökológusok a gyomokat a bolygatáshoz (talajművelés stb.) legjobban alkalmazkodó növényeknek tekintik. Ökológiai megközelítés szerint „a gyomnövények a másodlagos szukcesszió pionír fajai, ahol a szántóföld egy speciális terület” (BUNTING 1960). HOLZNER (1978) a pionír tulajdonságot az emberi tevékenységhez kapcsolja: „A gyomok az emberi termesztési tevékenységhez legjobban alkalmazkodó növények és azt jelentősen befolyásolják”.
Szerte a világon foglalkoznak növénytermesztéssel és a gyomok jelenléte, valamint az általuk okozott károk mindig problémát jelentenek.
A termesztendő növényre, két egymástól jól elkülöníthető tényező hat:
· abiotikus: időjárás (szélkár, belvíz, árvíz, jégkár, stb.)
· biotikus: ide sorolandók a kórokozók (vírusok, gombák, baktériumok; melyek által okozott kár mértéke 12%), az állati kártevők (nematódák, rovarok, atkák, rágcsálók, vadak, stb; az általuk okozott károk mértéke eléri a 14%-ot) és a gyomok (magról kelő egy-, és kétszikűek, az évelő egy-, és kétszikű valamint az élősködő gyomfajok tartoznak ide. Kártételük mértéke eléri a 9%-t)
Földünkön megközelítőleg 200000 növényfaj él, és mintegy 6700 gyomnövény (3,35%) befolyásolja a mezőgazdasági termelést. 200 azon fajok száma, amelyek világviszonylatban gondot okoznak és fontos gyomnövénynek tekinthetők (HOLM ET AL.1977). Ez mindössze csak 0,1%-a a fajok számának.
A felhasznált peszticideknek megközelítőleg 50-60%-t a herbicidek képviseltetik, ez is mutatja, hogy a kártételük nem elhanyagolható. A gyomok kártétele (közvetett és közvetlen) a következőkben jelentkezik:
· talaj hőmérsékletének csökkentése
· talaj tápanyagkészletének csökkentése
· talaj vízkészletének kimerítése
· termelési költségek növelése
· termés minőség és mennyiség rontása
· állati kártevők tápnövényei
· növényi kórokozók gazdanövényei
· élősködő gyomfajok
A rendszeres herbicid használat, a vetésforgó hiánya, a talajművelési rendszerek megváltozása, a nagymértékű műtrágyaadagok, valamint a nagy növényszám hatására a gyomösszetétel és gyomvegetáció jelentős változásokon ment keresztül az 50-es évektől kezdődően.
Vannak olyan fajok, melyek jobban elterjedtek, míg mások egyre háttérbe szorultak. A nem elhanyagolható költségek mellett jelentkezhetnek a túlzott vegyszerezésből eredő káros hatások és az egyes rezisztens fajok elszaporodásából eredő újabb vegyszerezési problémák stb. is. Ezenfelül a világproblémává kiszélesedett környezetvédelem is azt követeli, hogy a vegyszerezés mértékét a legszükségesebbre korlátozzuk.
Mai tudásunk szerint a gyomvegetáció ismeretében a rendelkezésre álló agrotechnikai lehetőségek maximális kihasználása a szükséges kémiai gyomirtással együtt adja meg azt a gyommentességet, ami a nagy termések eléréséhez szükséges. Ehhez pedig a gyomnövények ismeretén túl a művelt területek gyomösszetételének, az irtandó gyomok ökológiájának és egyes kultúrák ökológiai viszonyainak ismerete szükséges (UJVÁROSI 1973).
Munkánkban célul tűztük ki, hogy a Taraxacum officinale gyomnövényről szóló biológiai ismereteinket gyarapítsuk, további hasznos adatokat kapjunk. A kitűzött cél érdekében a következő vizsgálatok végeztük:
· Csírázásbiológiai vizsgálatok
· Regenerálódás vizsgálatok
· Tápelemtartalom vizsgálat
· Genetikai vizsgálatok
· Növekedés analízis vizsgálatok
· Kompetíciós képesség vizsgálatok
· Herbicidérzékenységi vizsgálatok
III IRODALMI ÁTTEKINTÉS
III.1 EREDETE, ELNEVEZÉSE
A gyomnövény első tudományos osztályba sorolását (klasszifikálása) 1753-ban Lineaus alkotta meg. A növényt Leontodon taraxacum néven jegyezte (JAEGER 1955). WIGGERS (1746-1811) írta le a Taraxacum nemzetséget, Georg Heinrich Weber pedig megalkotta 1780-ban a jelenlegi osztályba sorolását (BRITTON AND BROWN 1970).
A Taraxacum név eredete bizonytalan, azonban HOLM ET AL.(1997), JENNISKENS (1984) és MITICH
(1989) számos lehetséges forrást említenek munkáikban. A Taraxacum név arab eredetű, mely a „tarachakum”
(vadcseresznye), „tarakhshaqun” (vad cikória), „tharachschakuh”, „talkh chakok” vagy „tarashqun” (keserű gyógynövény) szavakból ered (DWYER 1977;JENNISKENS 1984;MITICH 1989). Egy másik magyarázat szerint görög eredetű szó, mely a „taraxis” (szembetegség), „tarassen” vagy „tarasos” (rendetlenség), „trogimon”
(ehető) és „akeomai” vagy „”akos” (orvosol, meggyógyít) szavakból ered (POWELL 1972;JENNISKENS 1984;
MITICH 1989). Az Officinale jelentése gyógyító, gyógyhatású vagy orvosság előállítására képes (SCHMIDT 1979).
A pitypang közönséges elnevezése egy szójáték eredménye, mely egy 13-ik századi Wales-i szókapcsolat
„Dant y Llew” (jelentése: „oroszlánfog”) alapján alakult ki (HEDRICH 1972). Elnevezése az éretlen kaszat alakjának (LOVELL AND ROWAN 1991), a levelek egyenetlen alakjának (JACKSON 1982), a fészekvirágzatban megjelenő sárga virágainak (ANGIER 1980) vagy az erős fehér karógyökerének (kihúzva egy agyagtalajból olyan, mintha egy oroszlán fogát húznánk ki) (DWYER 1977) köszönhető. A francia elnevezése „pissenlit”
(ágyba vizelés) a növény részeinek vizelethajtó hatásából ered (LOVEL AND ROWAN 1991).
III.2 RENDSZERTANI BESOROLÁSA, JELLEMZÉSE, VÁLTOZATAI
III.2.1 A TaraxacumTaraxacumTaraxacumTaraxacum nemzetség rendszertana
A pitypang vagy gyermekláncfű (Taraxacum) fajok a kétszikű (Dicotyledonopsida) zárvatermő (Angiospermatophyta) növények őszirózsa-alkatúak (Asteridae=Synandrae) alosztályába, az asztervirágúak (Asterales) renden belül a fészekvirágzatúak (Asteraceae=Compositae) családjának nyelvesvirágúak (Cichorioideae) alcsaládjába tartoznak. A pitypang nemzetség (Taraxacum) rendkívül formagazdag. A Taraxacum fajok nagy része apomiktikus poliploid (2n=24, 32, 40, 48) esetenként aneuploidok (2n=25, 26, 27) (TUTIN ET AL.1976), de vannak normális szaporodású és hibridogén eredetű taxonok is. Az apomiktikus fajoknál az idegen megporzás teljesen hiányzik és ezért a populáción belüli változékonyság alig vagy egyáltalán nem mutatható ki. Az apomiktikus taxon (apomikta) apomixissel, vagyis nem szexuális úton szaporodik. A mag ilyenkor álivaros (pszeudoszexuális) folyamat eredményeként jön létre, amikor az embrió kizárólag anyai szövetekből áll, a keletkezett utód genetikailag a női szülővel azonos. A Taraxacum fajoknál az apomixis partenogenezis formájában valósul meg, amikor még az érintetlen petesejtből fejlődik az embrió. Érthető tehát, hogy a pitypang fajoknál a változatosság kicsi, a populációk szinte teljesen egyforma egyedekből állnak. A rendszertani csoportok igen kevés bélyegben különböznek, de ez a különbség állandó, mert nincs lehetőség semmiféle kereszteződésre. A sok kis különbségű populációk így sok kisfajt képviselnek. Az eddig leírt Taraxacum kisfajok száma már több száz. Bonyolítja a pitypang-rendszertant az is, hogy a Taraxacum nemzetségben egy fajon belül találhatóak apomiktikus és szexuális egyedek (CZIMBER ÉS MAGYAR 2005).
A rendszertani besorolás fontos bélyege a külső fészekpikkely-levelek állása, melyről csak a kifejlett, élő egyedeknél bizonyosodhatunk meg (TUTIN ET AL.1976). A következő csoportokat vagy kisfajokat (30) írja le:
1. Taraxacum glaciale Huet ex Hand.-Mazz. csoport: Külső fészekpikkely levelei keskenyebbek a belsőknél, külső felületük fekete, belső részük hamvas, felállóak és tölcsér alakúak. Elterjedt az Appenini-hegységben és Görögország hegyeiben.
2. Taraxacum bessarabicum (Hornem.) Hand.-Mazz. csoport: Külső fészekpikkely levelei 7-8 mm nagyok, lándzsásak, sötétzöld színűek, felállóak vagy elfekvőek. Sós talajokat kedveli. Közép- és Kelet- Európában terjedt el.
3. Taraxacum serotinum csoport: Külső fészekpikkely levelei keskenyek, hamvaszöld színű vagy halványbarna szegélyű, vastag, puha, felálló, többé-kevésbé szőrözött, néha tölcséres. Száraz helyeken növekszik. Dél-, Közép- és Kelet Európában. Két faj, mely Európában elterjedt a Taraxacum pyropappum Boiss. & Reuter. és a Taraxacum serotinum (Waldst. & Kit.).
4. Taraxacum phymatocarpum csoport: Külső fészekpikkely levelei 0-6 mm nagyok, tojásdad alakúak, szürkés-zöld vagy egészen fekete színűek halvány szegéllyel, elterülőek, néha tölcsér alakúak.
Európában és az Alpokban terjedt el. Fajai a T. phymatocarpum J. Vahl, mely csak Grönlandon és Alaszkában fordul elő. Európai fajai a T. arcticum (Trautv.) Dahlst., T. dovrense (Dahlst.) Dahlst., T.
handelii J. Murri, és a T. reichenbachii Huter ex Dahlst.
5. Taraxacum glabrum csoport: Külső fészekpikkely levelei tojásdad-lándzsa alakúak, sötét színűek.
Észak-Nyugat Oroszország területein fordul elő. A következő fajok tartoznak ebbe a csoportba: T.
glabrum DC., T. nivale Lange ex Kihlman és T. turiense Orlova in Bobrov & Tzvelev.
6. Taraxacum pacherii csoport: Külső fészekpikkely levelei 6 mm nagyok, alakjuk tojásdadtól lándzsa alakúig bármi lehet, fekete, sötétzöld vagy olívazöld színűek többé-kevésbé halvány szegéllyel, elterülők vagy felállók. Kelet Alpok, Dél-Spanyolország hegyei valamint Korzika területein honosak.
Idetartozó fajai a T. litardieri Van Soest, a T. nevadense H. Lindb. és a T. pacheri Schultz Bip.
7. Taraxacum schroeteranum Hand. – Mazz. csoport: Külső fészekpikkely levelei tojásdad, kihegyesedő alakúak, szürkészöld színűek, gyakran vöröses halvány szegély nélküli, elterülők. Az Alpokban és Közép-Spanyolországban terjedtek el az idetartozó fajok.
8. Taraxacum bihynicum csoport: A külső fészekpikkely levelei széles, tojásdad, kihegyesedő alakú, félig vagy teljesen hártyaszerűen borított, vörös vagy lila, többé-kevésbé elterülők, gyakran pókhálószerűen erezett, tölcséres. Dél –Európa területeit hódították meg. 13 európai fajról számoltak be, ezek közül a 2 legfontosabb, mely nagy jelentőséggel bír területhódítása révén, a következő: T. megalorhizon (Forskal) Hand. – Mazz. és T. minimum (Brigant ex Guss.) N. Terrac.
9. Taraxacum obovatum csoport: Külső fészekpikkely levelei tojásdad, tojásdad-lándzsás alakúak, hamvaszöld színűek többé-kevésbé halvány szegéllyel és felállóak. Dél-Nyugat-Európa területeit népesíti be, valamint keletre egészen Szicíliáig. Idetartozó 2 faj, mely Európa-szerte elterjedt a T.
leucospermum Jordan és a T. obovatum (Willd.) DC.
10. Taraxacum glaucanthum csoport: Külső fészekpikkely levelei akár 0,8 cm hosszúak is lehetnek, tojásdad alakúak, hamvas, feltűnő szegéllyel, felállóak vagy elterülők, tölcséres. Sós talajokon fordulnak elő a következő fajok: T. glaucanthum (Ledeb.) DC., a T. hybernum Steven, a T. klokovii Litv. és a T. kok- saghyz Rodin.
11. Taraxacum ceratophorum csoport: Külső fészekpikkely levelei 7-12 mm nagyok, tojásdadtól a lándzsás alakig bármilyen lehet, zöld színűek és felállóak. Észak-Európa és az Alpok tájain fordulnak elő a fajok. 13 fajt sorolnak ebbe a csoportba, melyek közül 2 terjedt el széles körben, a T. brachyceras Dahlst. és a T. tornense T. C. E. Fries.
12. Taraxacum spectabile csoport: Külső fészekpikkely levelei tojásdad-lándzsás alakúak, néha lilás színűek egy keskeny, halvány szegéllyel, elterülők. Nedves termőhelyeken fordulnak elő, leginkább Észak és Nyugat-Európa területein. 38 fajt sorolnak ebbe a csoportba, melyek közül a következők terjedtek el széles körben: T. eximium Dahlst., T. faeroense (Dahlst.) Dahlst., T. reclinatum M. P. Christiansen és T.
spectabile Dahlst.
13. Taraxacum praestans csoport: Külső fészekpikkely levelei 6-12 mm nagyok, tojásdad-lándzsás alakúak, keskeny halvány szegéllyel, gyakran hamvas a belső felületük, felállóak vagy elterülők, szőrösek, Nedves helyek kedvelt gyomnövénye. Észak és Nyugat-Európában terjedtek el az idetartozó fajok.
42 fajt sorolnak ide, azonban a következők a jelentősebbek: T. euryphyllum (Dahlst.) M. P.
Christiansen, T. lainzii Van Soest, T. landmarkii Dahlst., T. maculigerum H. Lindb., T. naevosiforme Dahlst., T. naevosum Dahlst., T. praestans H. Lindb., T. purpuridens Dahlst., T. sagittifolium H. Lindb. és a T. stictophyllum Dahlst.
14. Taraxacum unguilobum csoport: Külső fészekpikkely levelei halványzöld színűek, végük rózsaszínes.
Nedves helyeken fordulnak elő, Észak-Nyugat-Európában. 2 fajt írtak le ebben a csoportban, a T.
fulvicarpum Dahlst. és a T. unguilobum Dahlst.
15. Taraxacum croceum csoport: Külső fészekpikkely levelei 7-12 mm hosszúak, lándzsás alakúak, sötétzöldek, néha hamvaszöld a belső felületük, felállóak vagy elterülőek, szőrözöttek. Észak-Európa területein terjedtek szét. 96 fajt sorolnak ebbe a csoportba, melyek nagy része Izland és Norvégia területein jelentős. A következő fajok a jelentősebbek közülük: T. ceratolobum Dahlst., T. craspedotum Dahlst., T. croceum Dahlst., T. cymbifolium H. Lindb., T. pycnostictum M. P. Christiansen, T. repletum (Dahlst.) Dahlst. és T. rhodolepis Dahlst.
16. Taraxacum adamii csoport: Külső fészekpikkely levelei 7-10 mm nagyok, lándzsás, sötétzöld, néha hamvas, szőrözött vagy fogazott, mereven felállóak. Nedves helyek gyomnövényei Észak és Nyugat-Európa egyes területein. 15 fajt sorolnak ide, melyek közül a következők a jelentősebbek, széles körben elterjedtebbek: T. adamii Claire, T. litorale Raunk. és T. nordstedtii Dahlst.
17. Taraxacum palustre csoport: Külső fészekpikkely levelei akár 7 mm nagyok is lehetnek, tojásdad, többé-kevésbé szélesen kicsúcsosodó, gyakran ibolya vagy lila színűek. Nedves területek benépesített lakói. Európa nagyobb részén elterjedt, kivétel a dél-nyugati része, valamint orosz és a sziget országok egyes területei. 56 fajt sorolnak ide, azonban a következők a jelentős mértékben elterjedt fajok: T. anglicum Dahlst., T. austrinum G. Hagl., T. balticiforme Dahlst., T. balticum Dahlst., T. crocodes Dahlst., T. decolorans Dahlst., T. divulsifolium Van Soest, T. frisicum Van Soest, T. germanicum Van Soest, T. heleonastes G. Hagl., T. hollandicum Van Soest, T. illyricum Dahlst., T. limnanthes G. Hagl., T. lividum (Waldst. & Kit.) Peterm., T. neoaellenii Van Soest, T. palustre (Lyons) Symons, T. scaturiginosum G.
Hagl., T. suecicum G. Hagl., T. turgosum (Schultz Bip.) Van Soest, T. udum Jordan és T. vindobonense Van Soest.
18. Taraxacum apenninum csoport: Külső fészekpikkely levelei 8 mm nagyok is lehetnek, alakjuk lándzsás- tojásdad, világos-, vagy sötétzöld színűek, ritkán lilásak, felállóak vagy elterülők. Közép és Dél- Európa hegyeiben találhatóak a fajok. 23 fajt írtak le ebben a csoportban, melyek közül a szélesebb körben elterjedt fajok a következők: T. apenninum (Ten.) Ten., T. carinthiacum Van Soest, T helveticum Van Soest, T. mattmarkense Van Soest, T. panalpinum Van Soest, T. parsennense Van Soest, T. petiolulatum Van Soest, T. saasense Van Soest, T. venustum Dahlst. és T. vetteri Van Soest.
19. Taraxacum nigricans csoport: Külső fészekpikkely levelei lándzsásak, sötét színűek, halvány szegély nélkül, felállóak vagy elterülők. Alpok, Kárpát-medence és Bulgári hegyvidékén elterjedt fajok tartoznak ide. 32 fajt sorolnak ebbe a csoportba, melyek közül a fontosabbak a következők: T.
aestivum Van Soest, T. cordatifolium Van Soest, T. nigricans (Kit.) Reichenb., T. perfissum Van Soest, T.
reophilum Van Soest, T. rhaeticum Van Soest. és T. rufocarpum Van Soest.
20. Taraxacum fontanum csoport: Külső fészekpikkely levelei 6-12 mm hosszúak, lándzsás alakúak, zöld színűek, felállók vagy hátrahajlók, általában csupaszak. Alpok, Kárpátok nyugati része valamint Korzika egyes területein terjedtek el. 18 fajt sorolnak ebbe a csoportba, melyek közül a következők a jelentősebbek: T. croceicarpum Van Soest, T. fontanicola Van Soest, T. fontanosquameum Van Soest, T.
fontanum Hand. – Mazz., T. magnopyramidophorum Van Soest. és T. pohlii Van Soest.
21. Taraxacum cucullatum csoport: Külső fészekpikkely levelei tojásdad-lándzsás alakúak, sötétzöldek, gyakran lilák, többé-kevésbé halvány szegéllyel, lágyan elterülők. Alpok tájain terjedt el. 10 faj tartozik ide: T. aureocucullatum Van Soest, T. concucullatum A. J. Richards, T. cucllatiforme Van Soest, T. cucllatum Dahlst., T. fontaniforme Van Soest, T. inclusum Walo Koch, T. luteocucullatum Walo Koch & Van Soest, T. oreophilum G. Hagl., T. sulger-bueelii Van Soest és T. tiroliense Dahlst.
22. Taraxacum dissectum (Ledeb.) Ledeb.: Külső fészekpikkely levelei keskeny tojásdad alakúak, zöldek, barnás erezettel, elterülők. Alpok, Pireneusok és Spanyolország területei.
23. Taraxacum obliquum csoport: Külső fészekpikkely levelei sötétzöldek, halvány szegéllyel, elterülők, tölcsér alakúak. Észak-Európa területein gyakori gyomfaj csoport. 2 fajt írtak le, a T. obliquum (Fries) Dahlst. és T. platyglossum Raunk.
24. Taraxacum erythrospermum csoport: Külső fészekpikkely leveleik 6 mm hosszúak is lehetnek, gyakran hamvaszöld és lilás színűek, halvány szegéllyel, elterülők vagy felállóak, ritkán visszahajlóak, általában tölcsér alakúak. Száraz területek gyomfajai egész Európában. 68 fajt jellemeztek ebben a csoportban, melyek közül a következők terjedtek el széles körben: T. austriacum Van Soest, T. badium Van Soest, T. brachyglossum (Dahlst.) Dahlst., T. braunblanquetii Van Soest, T. commixtum G. Hagl., T. decipiens Raunk., T. disseminatum G. Hagl., T. dunense Van Soest, T. glaucinum Dahlst., T. gotlandicum (Dahlst.) Dahlst., T. gracillimum Van Soest, T. isophyllum G. Hagl., T. lacistophyllum (Dahlst.) Dahlst., T. laetiforme Dahlst., T. laetum (Dahlst.) Dahlst., T. limbatum Dahlst., T. marginatum (Dahlst.) Dahlst., T. montesignum Van Soest, T. obscurans (Dahlst. ex H. Lindb. fil.), T. pindicola (Bald.) Hand. – Mazz., T. plumbeum Dahlst., T. polyschistum Dahlst., T. proximum (Dahlst.) Dahlst., T. rubicundum (Dahlst.) Dahlst., T saphycraspedum Saarson & G. Hagl., T. scanicum Dahlst., T. silesiacum Dahlst. es G. Hagl., T. taeniatum G.
Hagl. ex Holmgren, T. tenuilobum (Dahlst.) Dahlst. és T. xerophilum Marklund.
25. Taraxacum simile csoport: Külső fészekpikkely levelei zöldek, halvány szegéllyel, felállóak vagy elfekvők. Száraz területek növényei Észak-, Nyugat- és Közép-Európa tájain. 26 fajt sorolnak ebbe a csoportba, melyek közül a következők a jelentősek: T. canulum G. Hagl., T. degelii Dahlst., T. dissimile Dahlst., T. falcatum Brenner, T. isthmicola H. Lindb. fil., T. microlobum Marklund, T. placidum A. J.
Richards, T. proximiforme Van Soest, T. pseudocastaneum Van Soest, T. pseudolacistophyllum Van Soest, T.
purpureomarginatum Van Soest, T. schizophyllum Dahlst., T. simile Raunk., T. subdissimile Dahlst., T.
tanyolobum Dahlst. és T. tortilobum Florström.
26. Taraxacum fulvum csoport: Külső fészekpikkely levelei 6 mm nagyok is lehetnek, zöld színűek, gyakran halvány szegéllyel, hátrahajlók vagy elterülők. Száraz területek gyomnövényei szerte Észak-, Nyugat- és Közép-Európában. 13 fajt sorolnak ide, melyek közül szélesebb körben a következők terjedtek el: T. fulviforme Dahlst., T. fulvum Raunk., T. glauciniforme Dahlst. és T. oxoniense Dahlst.
27. Taraxacum gasparrinii csoport: A 26-os csoporthoz hasonló tulajdonságokkal rendelkezik. Száraz területeken terjedtek el nagyobb mértékben. Dél-Európa egyes részein. 11 fajt írtak le ebben a csoportban, melyek a következők: T. austriense Van Soest, T. dorchocarpum Van Soest, T. epirense Van Soest, T. gasparrinii Tineo ex Lojac., T. lambinonii Van Soest, T. nanum Van Soest, T. roseocarpum Van Soest, T. stenospermum Sennen ex Van Soest, T. thracicum Van Soest, T. vinosum Van Soest és T.
xanthiense Van Soest.
28. Taraxacum hoppeanum csoport: Hegyvidékek száraz területein terjedtek el, leginkább Közép és Dél- Európában. 8 fajt sorolnak ide: T. amborum G. Hagl., T. aquilonare Hand.-Mazz., T. capricum Van Soest, T. caramanicae Lojac., T. duriense Van Soest, T. hoppeanum Griseb., T. pieninicum Pawl. és T.
poliochlorum Dahlst.
29. Taraxacum crassipes csoport: Rét, legelő és tűlevelű erdei tisztások elterjedt fajai találhatók a csoportban. 21 fajt sorolnak ide, melyek közül a következők jelentősebbek: T. cochleatum Dahlst. &
H. Lindb. fil., T. crassipes H. Lindb. fil., T. galeatum Dahlst., T. kuusamoense H. Lindb. fil., T. polyxanthum Dahlst és T. rubrolineatum H. Lindb. fil.
30. Taraxacum officinale csoport: Bolygatott területeken, réteken, legelőkön és esős területeken terjedtek el egész Európán. Számos faj (több száz) tartozik ebbe a csoportba, melyek közül a következők terjedtek el szélesebb körben: T. acutangulum Marklund, T. aequilobum Dahlst., T. alatum H. Lindb. fil., T. ancistrolobum Dahlst., T. angustisquameum Dahlst., T. arrhenii Palmgren, T. atrovirens Dahlst., T.
aurosulum H. Lindb. fil., T. biformatum H. Lindb. fil., T. borgvallii Dahlst. ex G. Hagl., T. bracteatum Dahlst., T. caloschistum Dahlst., T. canoviride H. Lindbn. fil., T. caudatulum Dahlst., T. christiansenii G.
Hagl., T. copidophyllum Dahlst., T. cordatum Palmgren, T. crispifolium H. Lindb. fil., T. croceiflorum Dahlst., T. cyanolepis Dahlst., T. dahlstedtii H. Lindb. fil., T. dilaceratum M. P. Christiansen, T. dilatatum H. Lindb.
fil., T. duplidens H. Lindb. fil., T. ekmanii Dahlst., T. expallidiforme Dahlst., T. explicatum G. Hagl., T.
fasciatum Dahlst., T. florstroemii Marklund, T. geminatum G. Hagl., T. gibberum Marklund, T. haematicum G. Hagl., T. haematopus H. Lindb. fil., T. hamatiforme Dahlst., T. hamatum Raunk., T. hastatum Marklund, T. huelphersianum Dahlst., T. insigne E. L. Ekman ex Wiinst. & K. Jessen, T. involucratum Dahlst., T. kjellmanii Dahlst., T. laciniosifrons Wiinst., T. laciniosum Dahlst., T. laeticolor Dahlst., T leptodon Marklund, T. linguatum Dahlst. ex M. P. Christiansen & Wiinst, T. lingulatum Marklund, T.
longisquameum H. Lindb. fil., T. lucidum Dahlst., T lunare M. P. Christiansen, T. macranthum Dahlst., T.
maculatum Jordan, T. marklundii Palmgren, T. melanthoides Dahlst., T mimulum Dahlst., T. mucronatum H.
Lindb. fil., T. multilobum H. Lindb. fil., T. obliquilobum Dahlst., T. oblongatum Dahlst., T. occidentale Dahlst., T. OFFICINALE WEBER, T. pachylobum Dahlst., T. pallescens Dahlst., T. pallidulum H. Lindb. fil., T. pannucium Dahlst., T. pannulatiforme Dahlst., T. pannulatum Dahlst., T. parvuliceps H. Lindb. fil., T.
pectinatiforme H. Lindb. fil., T. piceatum Dahlst., T. polychroum E. L. Ekman ex Th. Lange, T. polyodon Dahlst., T. praecox Dahlst. ex Puolanne, T. privum Dahlst., T. pycnolobum Dahlst., T. raunkiaeri Wiinst., T. recurvum Dahlst., T. remotijugum H. Lindb. fil., T. retroflexum H. Lindb. fil., T. rhodopodum Dahlst. ex M. P. Christiansen & Wiinst, T. rubrisquameum M. P. Christiansen, T. sagittipotens Dahlst. & R. Ohlsén ex G. Hagl., T. scotiniforme Dahlst. ex G. Hagl., T. scotinum Dahlst., T. sellandii Dahlst., T. semiglobosum H.
Lindb. fil., T. eptentrionale Dahlst., T. speciosum Raunk., T. spilophyllum Dahlst., T. stenoschistum Dahlst., T.
stereodes E. L. Ekman, T. subcanescens Marklund ex Puolanne, T. subcyanolepis M. P. Christiansen, T.
subintegrum Dahlst., T. sublaciniosum Dahlst. & H. Lindb. fil., T. sublaeticolor Dahlst., T. subraticola G.
Hagl., T. subundulatum Dahlst., T. sundbergii Dahlst., T. tenebricans (Dahlst.) Dahlst., T. triangulare H.
Lindb. fil., T. trilobatum Palmgren, T. undulatiforme Dahlst., T. undulatum H. Lindb. fil., T. vastisectum Marklund ex puolanne és T. xanthosigma H. Lindb. fil.
Hazai viszonyok között 6 faj terjedt el (CZIMBER ÉS MAGYAR 2005):
1. Taraxacum serotinum (W. et K.) Poir. – kései pitypang 2. Taraxacum bessarabicum (Horn.) hand.-Mazz. – sziki pitypang 3. Taraxacum erythrospermum Andriz. – szarvacskás pitypang 4. Taraxacum obliquum (Fr.) Dahlst. – vöröses pitypang 5. Taraxacum officinale Weber ex Wiggers – pongyola pitypang 6. Taraxacum palustre (Lyons) Symons – lápi pitypang
III.2.2 Morfológiája
Hosszú, vastag függőleges gyöktörzse, karóalakú, húsos elágazó főgyökérben folytatódik, amelyből számos oldalgyökér ered. Az oldalgyökerek két sorban helyezkednek el a gyökér hossza mentén lefelé spirálisan, az órajárással megegyező irányban egyenletesen szétoszolva. A gyökér 2-3 cm átmérőjű és akár 1-2 m hosszúra is megnőhet (VON HOFSTEN 1954;SOLBRIG 1971). Gyöktörzse vörösbarna, harántráncos és rajta az előző év elhalt ripacsai találhatók. Az idősebb és erőteljesebb példányok gyöktörzse többfejű, mivel a főrügy elpusztulása után az új leveles hajtások oldalrügyből fejlődnek. Az elágazó főgyökér szintén vastag és ezen is találhatók haránt irányú ráncok. A ráncok a szövetek összehúzódásaiból adódnak és az a szerepük, hogy a tőleveles hajtásrendszer mindig közvetlenül a föld felszínén maradjon. A gyöktörzs szétdarabolva új növénnyé egészül ki, mert rajta rügyek vannak vagy képződhetnek.
UJVÁROSI (1973) az ilyen növényeket szaporodásra képes gyökerűeknek nevezi és a hemikriptofitonok harmadik csoportjába (H3) sorolja. Az oldalgyökerek feladatuk szerint lehetnek táplálékfelvevők vagy rögzítők. Ezek szöveti szerkezete is eltérő (CZIMBER ÉS MAGYAR 2005).
A pongyola pitypang föld feletti része jól fejlett, tömött levélrózsás, rövid szártagú hajtás. Szára 1-2,5 cm hosszú, különösen rövid internóduszokkal közvetlen a talajfelszínen vagy alatta (GIER AND BURRESS 1942;
HOLM ET AL.1997). A növény reproduktív életszakaszában a rövid szártagú hajtásból tőkocsány emelkedik ki. A tőkocsány hosszú, 5-50 cm, üreges, csupasz és átmérője az alapi résztől a csúcsi rész felé egyre csökken, úgymond elvékonyodik (GIER AND BURRESS 1942;GLEASON 1963;HOLM ET AL.1997), melynek végén található a fészekvirágzat.
A levélrózsa állandóan fejlődik, még télen sincs teljesen nyugalomban. A rügyeknek nincs rügypikkelye, ún. csupasz rügyek fejlődnek, és a levélkezdeményeket a nagy mennyiségben képződő soksejtű szőrök borítják. A levelek alakja nagyon változékony, mely lehet szárnyasan karéjozott, szárnyalt (WELSH 1974), karéjos vagy karéj nélküli, fogazott vagy erőteljesen bemetszett, csipkézett, fűrészes (HOLM ET AL.1997). A levélkaréjok többé-kevésbé háromszög alakúak (TUTIN ET AL.1976). Levelek 5-40 cm nagyok és 0,7-15 cm szélesek, a levél alapi részénél pedig szárnyasan kiszélesedő (GLEASON 1963;HOLM ET AL.1997;WELSH
1974). A levelek morfológiája, tagoltsága és a levélrózsa alakulása környezeti hatásoktól függ. Száraz, kopár helyen például a levélrózsa a talajhoz simul és a levéllemez erősen tagolt. Magas fű vagy kultúrnövények között levélrózsája laza, levelei felemelkedők és finoman tagoltak. A levelek felületén fedőszőrök és mirigyszőrök találhatók. A levél gerince üreges (WELSH 1974), amit tejedények fognak körül, a levélgerinc központi ürege a levélnyélben is végighúzódik. A levélér színe halványsárga, zöld, mélyvörös, barna színárnyalatú lehet (COLLINS 2000). A tőkocsány csöves, amit a szállítónyalábok szilárdítanak. A szállítónyalábok tejedényeket tartalmaznak (CZIMBER ÉS MAGYAR 2005,WELSH 1974).
A 2-5 cm átmérőjű fészekvirágzat (GIER AND BURRESS 1942;GLEASON 1963;HOLM ET AL.1997) belül üreges, mely hosszabb vagy rövidebb, vége felé elvékonyodó virágzati tőkocsányon ül. A gombszerűen kiszélesedő virágzati tengely felületén a nyelves virágok helyezkednek el. A virágok száma egy fészken belül akár 250 db is lehet. A fészekpikkely levelek keskenyek, zöldes-barnás színűek és két körben rendeződnek (HOLM ET AL.1997). A külső kör rendszerint hátrahajlik, a belső kör tagjai viszont felegyenesedők, és a ki nem nyílt virágzatot védik. Ezek csak akkor terülnek szét, mikor a sárga virágok a napfény hatására kinyílnak.
Délután a fészekpikkely levelek újra felhajlanak és az egész virágzat ismét összeborul. Ez a naponkénti nyitódás és csukódás mindaddig tart, amíg a megtermékenyítés végbe nem megy. Esős vagy hűvös időben a fészekvirágzat nem nyílik fel (CZIMBER ÉS MAGYAR 2005).
A virág kétivarú, kocsánya hiányzik, magháza alsóállású. A magház felső végén öt sziromlevélből alakult, zártcsövű, de csúcsán nyitott és hosszú, nyelvszerű pártát találunk (HOLM ET AL.1997). A pártacső színtelen, a nyelvszerű lemez (nyelves párta) élénksárga. A pártacsövet kívülről szőrkoszorú (bóbita) fogja körül, mely a csészelevelek módosulása. A pártacsőből magasan kiemelkedik a kétágú bibe, amelyet öt portokból összenőtt portokcső vesz körül. A rövid porzószálak a pártacső falára ránőttek. A ki nem fejlődött virágban a portokcső felső csúcsán zárt, és benne a bibe két ága szorosan egymáshoz simul (CZIMBER ÉS MAGYAR
2005).
A 30 μm-es pollenből (GIER AND BURRESS 1942) képződő kaszattermése szürkésbarna, zöldesbarna (olíva), visszás tojásdad alakú, hosszúkás, kissé összenyomott, durva felületű, görbe termés, amelyen hosszanti barázdák (5-8 db) futnak végig. Felső harmadában tüskés és a csúcsi részén a magház felső végéből alakult, hosszú, megnyúlt, hengerszerű terméscsőr folytatódik. A terméscsőr a végén bóbitát visel, mely 3-4 mm hosszú lehet, többé-kevésbé fehér színű (GLEASON 1963;HOLM ET AL.1997). Meleg időben a fészekpikkely levelek belső köre visszahajlik és a terméses fészek szétterül. A bóbitás terméseket a szél fújja el, de ha ez nem történik meg, a higroszkópos bóbita este összecsukódik, a fészekpikkely levelek a terméses fészket beborítják.
Nedves időben a virágzathoz hasonlóan a termések a fészekpikkely levelektől körülzárva maradnak (CZIMBER ÉS MAGYAR 2005).
III.2.3 Más fajokkal való rokonsága, változékonysága
A Taraxacum officinale rendkívüli hasonlóságot mutat más, illetőleg egyéb Taraxacum fajokhoz, melyeket a következőkben foglalunk össze.
A vörös magvú pitypang (Taraxacum laevigatum (Willd)) DC. nagyon hasonló a Taraxacum officinale növényhez, azonban ez a faj karcsúbb és leveleinek szeldeltsége (karéj) jóval erőteljesebb (GLEASON 1963). A levél karéjok általában keskenyebbek, az utolsó levélkaréj ritkán nagyobb, mint az oldalsók. A kaszatjai színe az érés során a világos vöröstől a vöröses-liláig változik (GLEASON 1963).
A Taraxacum officinale-t sokkal nehezebb megkülönböztetni a T. ceratophorum (Ledeb.) fajtól DC., mely a hegyvidéki területek közönséges pitypang faja. Ez a faj kevésbé robosztus, levél karéjai szélesebbek, kevesebb karéjokkal. A fészekvirágzata kisebb, mint a T. officinale fajé (GLEASON 1963). Levelei 8-20 cm nagyok, lándzsásak, karéjozott, sötétzöld színű, ritkán vékony és szőrözött. Karéjok nagyok. A levél bemetszése lehet még fogazott, levélnyele szárnyalt. Tőkocsánya 10-25 cm hosszú, erős és vaskos, valamint szőrrel borított.
Fészekvirágzata 35-50 mm átmérőjű. Fészektányér 15-20 x 15-20 mm. Külső fészekpikkely-levelek 7-12 mm nagyok, felállóak, tojásdadtól a lándzsás alakig bármely lehet, zöld színű halvány szegéllyel. Nyelves virágok keskenyek, sárga színűek vörös, lila vagy barna sávval. Kaszattermése barnás színű. A termés 3,5-4 mm nagy, csőre 7-11 mm hosszú, karcsú alakú. Apomiktikus faj (TUTIN ET AL.1976).
A lápi vagy mocsári pitypang (Taraxacum palustre (Lyons) Symons) szintén nagyon hasonló a pongyola pitypanghoz. Eredetileg T. turfosum néven jegyezték le (BRUNTON 1989), majd később kapta a T. palustre nevet (OLDHAM ET AL.1992). Levelei felállóak, keskenyek és fűrészes, fészekpikkelyei nagyon sötétek és szélesek (BRUNTON 1989;OLDHAM ET AL.1992).
További fajok, melyek hasonló megjelenésűek a T. officinale fajhoz a következők: T. lyratum (Lebed.) DC.
(CODY 2000), T. latilobum és T. laurentianum.
Morfológiailag hasonlóságot mutat még számos, a fészkesek (Asteraceae) családjába tartozó fajokhoz is.
Ilyen faj a Chondrilla juncea L., melynek tőlevélrózsája hasonló a pongyola pitypangéhoz, azonban szára elágazó, mely egyenes szélű leveleket és mélyen ülő fészekvirágzatot hordoz, benne 11 virággal (GLEASON
1963). Szürkés-zöld kétéves, évelő növény. Szára 50-100 cm, számtalan elágazással melyek csupaszak, ritkán durván szőrözöttek, különösen a magasabban lévő elágazások. Levelei szintén csupaszak vagy néhány durva szőrrel borított.
A Crepis L. nemzetségbe tartozó fajoknak fészekvirágzatuk sárga, nyelves virágokból áll. A levelei nem oly mértékben karéjozottak, mint a Taraxacum officinale gyomnövény esetében (GLEASON 1963). A Prenanthes L. nemzetség tagjainak hasonló levelei vannak, melyek összetéveszthetők a T. officinale leveleivel, azonban e fajoknak a tőkocsánya pikkelyezett, a nyelves virágok mellett pedig csöves virágok is vannak. A Tussilago farfara L. (martilapu) szintén nagyon hasonló a pongyola pitypanghoz, azonban ennél a fajnál a virágzatképzés és a virágzás megelőzi a levélképződést, nem úgy, mint a T. officinale faj esetében, ahol előbb levelet fejleszt, majd jóval utána a fészekvirágzatot (GLEASON 1963). Világossárga virágjaik alapján némileg összetéveszthetők a pongyola pitypang növénnyel a Hypochoeris L. nemzetség tagjai valamint a Leontodon autumnalis L (AARSEN
1981).
III.3 GAZDASÁGI JELENTŐSÉGE
III.3.1 Kártétele
A Taraxacum officinale WEB. évelő (H3-hemikriptofita), kétszikű gyomnövény, mely igen jól alkalmazkodott rétekhez, legelőkhöz, gyepekhez, kaszálókhoz, út menti területekhez, gyümölcs ültetvényekhez, valamint alkalmanként bolygatott területekhez (HOLM ET AL.1997). Problémás gyomfaj golf pályákon, parkokban, kertekben, atlétikai pályákon, mezőgazdasági kultúrákban, veteményes kertekben, kertészeti kultúrákban úgy, mint szamócában (WITTY AND BING 1985;RIDDLE ET AL.1991;HOLM ET AL. 1997). Esztétikai problémát is jelent gyepekben, ugyanis virágzásával valamint termésérlelésével megbontja a gyepek egyenletességét és sűrűségét (RIDDLE ET AL.1991).
Nyugat-Kanadában egyre növekvő problémakén tartják számon az egyéves kalászos kultúrákban, az olajos magvú vetésekben (DERKSEN AND THOMAS 1997), valamint kukorica, szója és őszibúza állományokban, ahol a 6 legfontosabb gyomnövény közé sorolták (FRICK AND THOMAS 1992). Az USA-ban a kukoricatermést hatalmas mértékekben veti vissza a pongyola pitypang, különösen akkor, ha a megelőző kultúra szintén kukorica volt, és nem szója (HARTWIG 1990). Pakisztán őszi búza kultúráinak egyik domináns gyomfaja (AHMAD 1993). További jelentősége a pillangós takarmánynövényekben, mint például lucerna és lóhere merül fel (LOWELL AND ROWAN 1991;HAMILL 1997).
Nálunk a legnagyobb kárt a csapadékos vidékeken okozza pillangós kultúrákban (UJVÁROSI 1973).
Gyepekben és füves területeken való megtelepedése és erőteljes fejlődése függ a füvek magasságától, valamint a kaszálások gyakoriságától. Minél magasabb a fű, a gyomfaj egyedszáma annál jobban csökken, köszönhető részben a fű okozta árnyékolásnak (MOLGAARD 1977).
Továbbá előfordul még nemzeti parkokban (TYSER AND WORLEY 1992) és borsmenta földeken az USA-ban (CARDA ET AL.1992). Gazdanövénye a gyapotmagfúró bogárnak (HAYNES AND SMITH 1992), káposztalepkének és hernyójának (DUSSOURD AND DENNO 1994), a zöld őszibarack levéltetűnek (KAAKEH AND HIGMIRE 1991), a Pseudomonas viridiflava baktériumnak, mely baktériumos tarkaságot és gumórothadást okoz hagymán (GITAITIS ET AL.1998), valamint számos vírusnak (BRCÁK 1979;MOUNTAIN ET AL.1983).
Gyógynövényként is alkalmazzák, azonban túlzott fogyasztása gátolhatja a máj funkcióját, inaktívvá teheti, valamint számos kellemetlen, nem kívánatos tünetet válthat ki (JACKSON 1982). Virágpora allergén hatású (HELBLING AND WUETRICH 1987), allergiás és fotoallergiás bőrkiütéseket okozhat (MARK ET AL. 1999).
III.3.2 Haszna
A Taraxacum officinale növényt évszázadok óta alkalmazzák orvoslási célból számos betegség kezelésére (CULPEPER 1826;POWELL 1972) úgy, mint a májfunkció javítására, a koleszterin és vérnyomás alacsony szinten tartására (MATTERN 1994), túlsúlyos betegek testsúly csökkentésére, epehólyag betegségek kezelésére (DALBY 1999). Vizelethajtó hatása is ismeretes (RÁCZ –KOTILLA ET AL.1974). Levelét összekeverve más növényi vegyületekkel gyógyhatású készítményként használható májra, vesére, bőrre valamint rákos eredetű megbetegedésekre is (NEAMTU ET AL.1992). Jótékony hatása van vesekőre is (GRASES ET AL.1994).
Tápértéke magas, köszönhető ez a nyomokban megtalálható esszenciális olajoknak, alacsony mennyiségben fellelhető tanninnak, mely javít a növény minőségén és emészthetőségén (palatabilitás) (FALKOWSKI ET AL.1990). A növény a fehér herében is megtalálható fehérjét tartalmaz (BOCKHOLT ET AL. 1995) valamint értékes táplálék a benne lévő zsír és szénhidráttartalom miatt (SPATZ AND BAUMGARTNER
1990). A legtöbb háziállat azonban mégsem fogyasztja szívesen a növény keserű íze miatt (FALKOWSKI ET AL.1990). Talajjavításra is felhasználták több-kevesebb sikerrel, mégis hatékonysága alulmaradt más egyéb gyomokkal szemben (LI 1996).
Allelopatikus hatással is rendelkezik (JARVIS 1989;FALKOWSKI ET AL.1990;ALVAREZ ET AL.1998). Zöld salátaként is felhasználják. Csak Torontóban 155 tonna levelet (értékét tekintve 595000 kanadai dollár) értékesítettek a piacokon 1988-89-ben. Ezzel a programmal akarták beindítani a Taraxacum officinale termesztését és gyökerének a piacokon való értékesítését (LETCHAMO AND GOSSELIN 1995). A növény kivonatait sajtgyártásban is alkalmazták, köszönhetően a benne lévő tejnedv alvadó képességének valamint proteolitikus tulajdonságának (AKUZAWA AND YOKOYAMA 1988). A növény különböző részeit felhasználják levesekhez, desszertekhez, italokhoz, beleértve teák, borok, sörök valamint kávépótlóként (leszárított gyökerét darálják és pörkölik) (GAIL 1994; DALBY 1999;KHAN 2001). Virágzata méhek számára ideális nektárgyűjtő hely, melyet a méhészek előszeretettel használnak ki (GAIL 1994;DALBY 1999).
Környezeti szennyezés bioindikátora, ugyanis széles körben elterjedt növényről lévén szó, képes a leveleiben felhalmozni a fémeket, mint As, Br, Cd, Co, Cu, Cr, Hg, Mn, Pb, Sb, Se és Zn (KULEFF AND
DJINGOVA 1984;DJINGOVA ET AL.1986;SIMON ET AL.1996;SAMKAEVA ET AL.2001), továbbá Fe és Ni elemeket is (KRÓLAK 2001, 2003). A szennyezés mértékének növekedése a kaszatok hosszának és tömegének a csökkenését okozza, azonban a kaszatok száma növekszik a gyomnövény kedvezőtlen feltételekhez való kiváló alkalmazkodása révén (SAVINOV 1998).