DOKTORI (Ph.D.) ÉRTEKEZÉS
MARÁCZI KATALIN
PANNON EGYETEM GEORGIKON KAR
KESZTHELY
2014
PANNON EGYETEM GEORGIKON KAR
NÖVÉNYTERMESZTÉSI ÉS KERTÉSZETTUDOMÁNYI DOKTORI ISKOLA
Iskolavezető Dr. Kocsis László D.Sc.
egyetem tanár
Témavezető
Horváthné Dr. Baracsi Éva Ph.D.
egyetemi docens
Exota örökzöld lomblevelű díszcserjék új télállósági zónákba sorolása hidegtűrési és
ökofiziológiai vizsgálatok alapján
D
OKTORI(P
H.D.)
ÉRTEKEZÉSMaráczi Katalin
Keszthely
2014
Exota örökzöld lomblevelű díszcserjék új télállósági zónákba sorolása hidegtűrési és ökofiziológiai vizsgálatok alapján
Értekezés doktori (PhD) fokozat elnyerése érdekében Írta:
Maráczi Katalin
Készült a Pannon Egyetem Növénytermesztési és Kertészeti Tudományok Doktori Iskolájában
Témavezető: Horváthné Dr. Baracsi Éva Elfogadásra javaslom (igen / nem)
(aláírás)**
A jelölt a doktori szigorlaton 97,5 %-ot ért el.
Az értekezést bírálóként elfogadásra javaslom:
Bíráló neve: …... …... igen /nem
……….
(aláírás) Bíráló neve: …... …...) igen /nem
……….
(aláírás) A jelölt az értekezés nyilvános vitáján …...%-ot ért el.
Keszthely, ……….
a Bíráló Bizottság elnöke A doktori (PhD) oklevél minősítése…...
………
Az EDHT elnöke
TARTALOMJEGYZÉK
1 KIVONATOK ... 7
1.1 Magyar nyelvű kivonat ... 7
1.2 Abstact ... 9
1.3 Auszug ... 10
2 BEVEZETÉS ... 11
3 CÉLKITŰZÉS ... 12
4 IRODALMI ÁTTEKINTÉS ... 14
4.1 A fás szárú növények jelentősége ... 14
4.2 Az örökzöld növények jelentősége ... 16
4.2.1 A lomblevelű örökzöldek származása ... 18
4.2.2 A lomblevelű örökzöldek igényei ... 19
4.2.3 A lomblevelű örökzöldek felhasználási lehetőségei ... 20
4.2.4 A lomblevelű örökzöldek választékának alakulása a Prenornál ... 20
4.3 A klímaváltozás tendenciái Európában... 22
4.3.1 A klímaváltozás várható tendenciái Magyarországon ... 22
A csapadék mennyiségének a változása ... 22
A hőmérsékleti értékek változása ... 23
4.3.2 Az éghajlat változásának tendenciái Keszthelyen ... 24
A csapadék mennyiségének változása ... 24
A hőmérsékleti értékek változása ... 24
4.4 A növények télállóságának és fagytűrésének környezeti feltételei és élettani háttere 25 4.5 A fásszárú dísznövények fagytűrőképességének meghatározása télállósági zónák alapján 30 4.6 A növényekre ható stressz ... 36
4.7 A stressz hatásának mérése ökofiziológiai módszerekkel ... 38
4.7.1 A stressz hatása a fotoszintetikus pigmentek mennyiségére ... 38
4.7.2 A stressz és a klorofill fluoreszcencia kapcsolata ... 41
4.8 A lomblevél szöveti szerkezetének szerepe a környezeti feltételekhez való alkalmazkodásban ... 44
5 ANYAG ÉS MÓDSZER ... 46
5.1 A kísérletekben szereplő növényanyag bemutatása... 46
5.2 A kísérleti területek bemutatása ... 53
5.3 A piackutatás és adat felvételezés módszere ... 57
5.4 Meteorológiai megfigyelések 2007 és 2014 között ... 58
5.5 Klímakamrás vizsgálat módszere ... 61
5.6 A szabadföldi télállósági megfigyelés módszere ... 61
5.7 A fenológiai megfigyelések módszere ... 62
5.8 A klorofill-tartalom mérések módszere ... 62
5.9 A fluoreszcencia mérések módszere ... 63
5.10 A levélanatómiai vizsgálatok módszere ... 64
5.11 Statisztikai kiértékelés ... 64
6 EREDMÉNYEK ÉS ÉRTÉKELÉSÜK ... 65
6.1 A vizsgálatainkban szereplő taxonok hazai fellelhetősége és télállóságuk felvételezése Magyarország gyűjteményes kertjeiben ... 65
6.2 A vizsgálatainkban szereplő taxonok piackutatásának eredményei ... 67
6.3 A hidegtűrési és télállósági vizsgálatok eredményei ... 69
6.3.1 A klímakamrás kísérlet eredményei ... 69
6.3.2 A szabadföldi télállósági megfigyelések eredményei ... 72
6.4 A fenológiai megfigyelések eredményei két élőhely és hét év (2007-2014) átlagában 82 6.5 A roncsolásmentes ökofiziológiai vizsgálatok eredményei ... 93
6.5.1 SPAD mérések eredményei ... 93
6.5.2 A fluoreszcencia mérések eredményei ... 97
A 2009. évi mérések eredményei ... 97
A 2010. évi mérések eredményei ... 99
A 2009. évi és 2010. évi eredmények összehasonlítása ... 102
Az egyes fajok fluoreszcencia méréseinek az eredményei ... 103
6.6 A levélanatómiai vizsgálatok eredményei ... 111
6.6.1 Az Ilex aquifolium levélanatómiai vizsgálatának eredménye ... 112
6.6.2 Az Ilex cornuta levélanatómiai vizsgálatának eredménye ... 113
6.6.3 A Nandina domestica levélanatómiai vizsgálatának eredménye ... 115
6.6.4 A Viburnum cinnamonifolium levélanatómiai vizsgálatának eredménye ... 116
6.6.5 A Viburnum x pragense levélanatómiai vizsgálatának eredménye ... 117
6.6.6 A Viburnum tinus levélanatómiai vizsgálatának eredménye ... 118
7 KÖVETKEZTETÉSEK ... 120
7.1 A vizsgált fajok télállósági zónába sorolása ... 120
7.2 A SPAD-mérésekből levonható következtetések... 121
7.3 A fluoreszcencia mérésekből levonható következtetések ... 122
7.4 A levélanatómiai vizsgálatokból levonható következtetések ... 123
8 ÖSSZEFOGLALÁS ... 124
9 ÚJ TUDOMÁNYOS EREDMÉNYEK ... 129
10 THESIS POINTS ... 130
11 KÖSZÖNETNYILVÁNÍTÁS ... 131
12 IRODALOMJEGYZÉK ... 132
13 MELLÉKLETEK ... 149
1. melléklet. A lomblevelű örökzöldek fajtaválasztékának alakulása a Prenornál ... 149
2. melléklet. A főbb meteorológiai jellemzők 2007-2014 közötti telekről Keszthelyen 151 3. melléklet. A főbb meteorológiai jellemzők 2007-2014 közötti telekről Cserszegtomajon ... 152
4. melléklet. Télállósági vizsgálatokhoz tartozó bonitálási táblázat ... 153
5. melléklet. Statisztikai mutatószámok és korrelációs mérőszámok a 2010-es SPAD mérésekhez ... 154
6. melléklet. Statisztikai mutatószámok és korrelációs mérőszámok a 2013-as SPAD mérésekhez ... 155
7. melléklet. A SPAD-mérések szórásainak értéke 2010-ben ... 156
8. melléklet. A SPAD-mérések szórásainak értéke 2013-ban ... 157
9. melléklet. A PSII aktuális kvantumhatásfoka a három különböző élőhelyen a 2009-es évben ... 158
10. melléklet. A PSII aktuális kvantumhatásfoka a két különböző élőhelyen a 2010-es évben 159 11. melléklet. A kéttényezős varianciaanalízis varianciatáblái a fluoreszcencia mérésekhez ... 160
Jelmagyarázat
ΦPSII=Y=ΔF/Fm’= (Fm’ −Fs)/Fm’
Fm’a fényadaptált fotoszintetikus rendszerre jellemző maximális fluoreszcencia Fs az adott fényintenzitás hatására kiváltott fluoreszcencia egyensúlyi szintje ΔF/F m ' effektív vagy aktuális kvantumhatásfok
Fv/Fm optimális kvantumhatásfok
Az értekezésben szereplő fotókat készítették:
Horváthné Dr. Baracsi Éva: 6; 8; 9; 16-32, 34-46 Maráczi Katalin: 5; 7; 11-15; 33, 69; 71-80 Dr. Pintér Csaba: 70
1 KIVONATOK
1.1 Magyar nyelvű kivonat
Exota örökzöld lomblevelű díszcserjék új télállósági zónákba sorolása hidegtűrési és ökofiziológiai vizsgálatok alapján
A kutatómunkánkkal szeretnénk hozzájárulni a hazai lomblevelű örökzöld fásszárú dísznövénykínálat bővítéséhez. A vizsgálatainkba nagy egyedszámú állományában két élőhelyen az alábbi taxonokat vontuk be: Aucuba japonica Thunb. ’Rozzanie’, Cotoneaster franchetii Bois, Elaeagnus pungens Thunb. ’Maculata Aurea’, Ilex cornuta Lindl. and Paxt., Ligustrum sinense Lour., Ligustrum texanum Thunb., Nandina domestica Thunb., Osmanthus heterophyllus (G. Don) P.S. Green, Phillyrea angustifolia L., Photinia x fraseri Dress’Red Robin’, Prunus lusitanica L., Sarcococca hoockeriana Baill., Viburnum cinnamonifolium Redh., Viburnum tinus L. Annak érdekében, hogy ezen fajok igényeit megismerjük többféle vizsgálatot végeztünk.
Meghatároztuk klímakamrás vizsgálatok és szabadföldi viszonyok között a fagytűrésüket és ezek alapján a klímazónába sorolásukat pontosítottuk. Hazánk szubmediterrán régiójára vonatkozóan a fenológiai jellemzőiket meghatároztuk, valamint eltérő élőhelyeken ökofiziológiai vizsgálatokat végeztünk. A télállósági, ill. fenológiai megfigyeléseket 2007 és 2014 között folytattuk. A Cotoneaster franchetii-t a 6-os télállósági zónába soroltuk.
Hazánkban elsőként használtunk hordozható ökofiziológiai mérésekre alkalmas készülékeket fásszárú dísznövények állapotfelmérésére természetes élőhelyükön. Ezeknek a vizsgálatoknak az elvégzése arra irányult, hogy a kísérleti lomblevelű örökzöld fajokról megállapítsuk, hogy miként reagálnak az eltérő élőhelyekre.
A levél klorofill tartalma az egyik legfontosabb paraméter, amely a növény fiziológiai állapotával van kapcsolatban. A lombozat klorofill koncentrációjának az ismerete pontos becslést adhat a növény vitalitásáról. Kísérleteinkben a hordozható SPAD-502-es műszert alkalmaztunk a klorofill-tartalom mérésre. A mértékegység nélküli SPAD-index a tenyészidőszak előrehaladtával, a levelek öregedésével nőtt, továbbá a SPAD értékek szórásadataiból a lomblevelű örökzöldek esetében a nagyobb mértékű levélváltásra következtethetünk, valamint megállapítottuk, hogy a színes levelű fajták esetében az őszi SPAD-érték változás összefüggésben van a domborzati viszonyokból eredő besugárzás
mértékével.
A klorofill fluoreszcencia vizsgálat gyorsan és pontosan számszerűsíti a növényi stressz-toleranciát. Ennek mérésére vizsgálatainkban a PAM-2000-es fluorométert használtuk.
Bebizonyosodott, hogy a Cotoneaster franchetii és a Prunus lusitanica kevésbé érzékeny a különböző élőhelyekre, valamint az Ilex cornuta és az Osmanthus heterophyllus konténeres növényként jól alkalmazható.
1.2 Abstact
Classification of foreign broadleaf evergreen shrubs to new winter hardiness zones based on their cold tolerance and ecophysiological examinations
The investigations were made between 2007 and 2014 included the following taxa:
Aucuba japonica Thunb. ’Rozzanie’, Cotoneaster franchetii Bois, Elaeagnus pungens Thunb.
’Maculata Aurea’, Ilex cornuta Lindl. and Paxt., Ligustrum sinense Lour., Ligustrum texanum Thunb., Nandina domestica Thunb., Osmanthus heterophyllus (G. Don) P.S. Green, Phillyrea angustifolia L., Photinia x fraseri Dress’Red Robin’, Prunus lusitanica L., Sarcococca hoockeriana Baill., Viburnum cinnamonifolium Redh., Viburnum tinus L.. Their frost tolerance was determined by freezing chamber tests and in two different field conditions.
According to these, the classification of their USDA hardiness zone was refined and their phenological characteristics in the sub-Mediterranean region of Hungary were determined.
Non-destructive ecophysiological examinations were carried out to see how the broadleaf evergreen species respond to different habitats. The knowledge of foliage chlorophyll concentration can give an accurate estimation of the vitality of plants, which was found by using an instrument called SPAD-502 to this study. The chlorophyll fluorescence quantifies the plant stress tolerance quickly and accurately, and the PAM-2000 fluorometer was used to this examination.
1.3 Auszug
Neuklassifikation der Winterhärtezonen ausländischer Laubimmergrünen auf der Grundlage von Untersuchungen ihrer Kältetoleranz und ökophysiologischen
Eigenschaften
In den Untersuchungen wurden die folgende Taxen zwischen 2007 und 2014 eingeschlossen: Aucuba japonica Thunb. ’Rozzanie’, Cotoneaster franchetii Bois, Elaeagnus pungens Thunb. ’Maculata Aurea’, Ilex cornuta Lindl. and Paxt., Ligustrum sinense Lour., Ligustrum texanum Thunb., Nandina domestica Thunb., Osmanthus heterophyllus (G. Don) P.S. Green, Phillyrea angustifolia L., Photinia x fraseri Dress’Red Robin’, Prunus lusitanica L., Sarcococca hoockeriana Baill., Viburnum cinnamonifolium Redh., Viburnum tinus L.. Ihre Frosttoleranz wurde in der Klimakammer und auf zwei verschiedenen Versuchsgeländen überprüft und aufgrund dieses Versuches wurden die Frosthärtezonen verfeinert. Im Submediterranen Raum von Ungarn wurden die phänologischen Merkmale der Pflanzen festgestellt. Die non-destruktive ökophysiologische Untersuchungen der Laubimmergrünen sollte feststellen, wie sie auf verschiedene Lebensräume reagieren. Das Wissen über die Chlorophyllkonzentration des Laubes kann uns eine zuverlässige Schätzung der Vitalität der Pflanzen geben. Für diese Messungen wurde das Messgerät SPAD-502 benutzt. Der Chlorophyllfluoreszenz –Test qualifiziert schnell und präzise die Stresstoleranz von Pflanzen.
Zu diesem Test wurde das PAM-2000 Flourometer benutzt.
2 BEVEZETÉS
A hazai kertkultúra a rendszerváltás óta sokat változott köszönhetően annak, hogy megnyíltak a határok az áruk és az egyének számára, valamint a gazdasági átalakulás során megjelentek a külföldön már rég jelenlévő értékesítési csatornák. Lassan eltűnnek a régi korszak kiskertjei a bungalóval, s előtérbe kerülnek az igényes és színvonalas parkosítások, mind a magánkertekben, mind pedig a közterületeken. Az a hibás szemlélet, hogy a dísznövény luxuscikk, szerencsére már idejét múlt. Sokkal inkább az életminőség-javításának egy fontos eszköze azáltal, hogy az egészséges környezet fenntartásában is szerepet játszik. A zöldfelület megköti a szén-dioxidot, oxigént bocsát ki, árnyékot ad, a fásszárúakkal beültetett területeken kedvezőbb a mikroklíma. A dísznövények fontos szerepet játszanak továbbá a víz körforgásában és megtartásában és mindezek által lakható és egészséges környezetet biztosítanak az emberek számára.
A megváltozott fogyasztói szokások az elmúlt évtizedben a melegebb területekről származó fajok iránti igény növekedését eredményezték. A kereskedelmi forgalomban megjelenő lomblevelű örökzöld díszcserje fajok előtérbe kerülése szemmel látható. Hazánk természetes flórája földtörténeti okok miatt szegény örökzöld lomblevelű növényekben, a választékot csupán a borostyán (Hedera) és néhány törpecserje {babérboroszlán (Daphne laureola), csodabogyók (Ruscus)} alkotják. A kertjeinkben meghonosodott egzóták, mint például a közönséges mahónia (Mahonia aquifolium) Észak-Amerikában, az örökzöld madárbirsek (Cotoneaster) és az örökzöld borbolyák (Berberis) pedig csaknem kivétel nélkül Kelet-Ázsiában, főleg Kínában és Japánban őshonosak. Napjainkban a kereskedelmi forgalomban megjelenő lomblevelű örökzöld fajok a magyarországinál valamivel melegebb és főképp enyhébb telű térség növényei. Ezért a szélesebb körben való alkalmazásuk előtt szükséges megvizsgálni őket úgy fenológiai, mint ökofiziológiai szempontból annak érdekében, hogy díszítő értékük teljes mértékben kiteljesedhessena hazai klímában is. Az új fajok megjelenése a díszfaiskolai termesztés számára is új feladatokat ad a termesztés oldaláról, de ugyanakkor új lehetőségeket is jelent a termékpalettájuk bővítésére.
Így a többi kertészeti ágazathoz hasonlóan, a díszfaiskolai termesztésnek is fel kell készülni a klíma várható módosulásaira. Kutatásainkban a fentiek figyelembevételével olyan melegigényes örökzöld lomblevelű díszcserjéket vontunk be, amelyek az elkövetkező években hazánk egyes területein egyre inkább az érdeklődés előterébe, ill. alkalmazásra kerülhetnek.
3 CÉLKITŰZÉS
Kutatómunkánkkal hozzá szeretnénk járulni a hazai lomblevelű örökzöld fásszárú dísznövénykínálat bővítéséhez, amelynek érdekében a kísérletben szereplő fajokra vonatkozóan – Aucuba japonica Thunb. ’Rozzanie’, Cotoneaster franchetii Bois, Elaeagnus pungens Thunb. ’Maculata Aurea’, Ilex cornuta Lindl. and Paxt., Ligustrum sinense Lour., Ligustrum texanum Thunb., Nandina domestica Thunb., Osmanthus heterophyllus (G. Don) P.S. Green, Phillyrea angustifolia L., Photinia x fraseri Dress’Red Robin’, Prunus lusitanica L., Sarcococca hoockeriana Baill., Viburnum cinnamonifolium Redh., Viburnum tinus L. – a következők vizsgálatát tűztük ki célul:
1. A hazai gyűjteményes kertek ún. tartamkísérleteiben való előfordulásuk és télállóságuk felmérését, valamint volumenük meghatározását a hazai dísznövény forgalmazásban.
2. Fagytűrésük meghatározását klímakamrás vizsgálatok és szabadföldi viszonyok között.
3. Klímazónába sorolásuk pontosítását hazánk éghajlatára vonatkozóan.
4. Fenológiai jellemzőik meghatározását hazánk szubmediterrán régiójában.
5. Ökofiziológiai vizsgálatok végzését (SPAD-502, PAM-2000) eltérő élőhelyeken.
6. Hat kiválasztott faj levélanatómiai jellemzőinek meghatározását.
1. táblázat: Az egyes vizsgálatokban szereplő taxonok áttekintő táblázata
klímakamrás vizsgálat szabadföldi télállóság vizsgálat fenológiai megfigyelés SPAD mérés klorofill fluoreszcencia mérés levélanatómiai vizsgálat
A. japonica ’Rozzanie’ igen igen igen igen igen
C. franchetii igen igen igen igen igen
E. pungens ’Maculata
Aurea’ igen igen igen igen igen
I. cornuta igen igen igen igen igen
L. sinense igen igen igen igen
klímakamrás vizsgálat szabadföldi télállóság vizsgálat fenológiai megfigyelés SPAD mérés klorofill fluoreszcencia mérés levélanatómiai vizsgálat
L. texanum igen igen igen igen igen
N. domestica igen igen igen igen igen
O. heterophyllus igen igen igen igen
Ph. angustifolia igen igen igen igen igen
Ph. fraseri x ’Red Robin’ igen igen igen igen igen
P. lusitanica igen igen igen igen igen
S. hoockeriana igen igen igen igen igen
V. cinnamonifolium igen igen igen igen igen igen
V. tinus igen igen igen igen igen
V. x pragense igen
I. aqufolium igen
4 IRODALMI ÁTTEKINTÉS
4.1 A fás szárú növények jelentősége
Ha azt szeretnénk, hogy a kertünk egy éven át díszítsen, akkor virágokat ültetünk bele, de ha azt szeretnénk, hogy életünk végéig legyen miben gyönyörködnünk, akkor cserjéket és fákat telepítünk. Ezek a fás szárú növények folyamatosan nőnek, évről évre változnak, a látvány soha nem lesz ugyanaz. Ezen növények folyamatos fejlődésével a kert súlypontjai is megváltoznak, más lesz a hangsúlyos. Azzal, hogy megjelennek a levelek, a virágok és a termések, amelyek magasabban helyezkednek el, mint az ágyás növényei, szó szerint felsőbb dimenzióba lép a kertünk. Hála a fák és cserjék fás részei karakterességének, a kert föld feletti része még akkor is látványos, ha az évelők már nem díszítenek. Ezáltal a kertnek bizonyos fajta állandóságot kölcsönöznek évszakról évszakra, évről évre. Az elegáns ágak, a különleges törzsmintázat a lomb nélküli cserjéket és fákat is látványossá teszi még a legzordabb teleken is. E tulajdonságaik miatt a fás szárú növények a kertek uralkodó elemeivé válnak, fejlődésükkel a táj kialakításában nagymértékű szerepet játszanak (Hillier és Kelly, 1997).
A díszfák és díszcserjék esztétikai hatása elsősorban formai, melynek a téralakításban van jelentősége, virágzáskor és az őszi lombszíneződésnél – színes lombú fajták esetén az egész vegetáció idején – érvényesül színhatásuk, mely a zöldfelület karakterisztikus képének kialakításában nélkülözhetetlen. Környezetük mikroklímájának kialakításában szerepük elsődleges: szabályozzák a hőmérsékletet és a páratartalmat, csökkentik a levegőszennyeződés, a zajártalom és a szél erősségének mértékét. Az összefüggő díszfa- és díszcserjeállományok mellett a szoliter fák és cserjék is igen fontos esztétikai elemek. A fás szárú dísznövények a zöldfelület hosszú életű növényanyagai. Funkcióképességük a kiültetésük után hosszú idő alatt (fajtól függően 3-20 év) alakul ki. Díszértéküket évtizedeken át megtartják, eszmei értékük ezalatt növekszik (Nagy, 1980).
A parkokban a növények alkalmazásánál fontos azok esztétikai értéke, amely jó tervezéssel kiemelhető, viszont a rosszul összeválogatott növények vegyes érzelmeket válthatnak ki a szemlélőből. Példaként álljon itt, hogy az oszlopos formájú növények (legismertebb a Populus nigra ´Italica´) felkiáltójelként jelennek meg, a figyelem felhívására is alkalmasak. Az oszlopos és a piramis formájú örökzöldek (Chamaecyparis, Taxus, Thuja) ünnepélyes érzetet kelt a kertben. Jó társítással a világító fehér törzsű Betula-k vertikális elemei a térnek. A kis koronás fák a keskeny utcák fásításának fontos elemei (Kiácz és Szendrői, 1980).
A növények a velük beültetett területen csökkentik a talajt érő besugárzást, növelik a levegő páratartalmát, valamint a növénnyel beültetett és a be nem ültetett terület között légmozgás alakul ki, megindul a légcsere.
A fák, cserjék árnyékában a léghőmérséklet akár 2 °C-al is alacsonyabb lehet, mint a tőle pár méterre lévő járdán. Közérzetünket javítja már a kis légmozgás is. Erre is alkalmasak a városi parkok zöldfelületei, mivel az átforrósodott házfalakról, úttestről felfele induló levegő ezek hűvösebb levegőjével kicserélődik, ami helyi légmozgást eredményez. Lakótelepeinken gyakran alakulnak ki huzatos helyek, szélcsatornák, aminek káros hatását magas növények telepítésével mérsékelhetjük. Közlekedési útvonalak szélvédelmét jól szolgálják a fasorok, facsoportok, a szélvédő erdősávok (Kiácz és Szendrői, 1980).
A cserjék fontos, nélkülözhetetlen elemei a parkoknak és a fasoroknak. Egyrészt jelentős asszimiláló felülettel rendelkeznek, másrészt a kert meghatározó elemei. Az útmenti fasorok mellett a sövénycserjék külön védelmi funkciót töltenek be azáltal, hogy a gépjárművek kipufogó zónáiban nyútanak mechanikai visszatartó hatást. A sövénycserjék elsősorban a gyermekeket védik, mivel ők a kipufogó zónához közeli levegőt lélegzik be (Radó, 2001).
Jól kialakított fa és cserjesávval a zajhatás csökkenthető. Gyorsforgalmú utak mentén a kisodródó autók felfogásával a baleset súlyosságát jelentősen csökkenti a mellé vékony vesszejű cserjéből telepített cserjesáv (Kiácz és Szendrői, 1980).
Gelbrich (1974) egytől húszig terjedő skálán értékelte a különféle zöldfelületeket, azok nyári és téli zöldtömege, a mikroklímára történő hatása, a fűst szűrése, a zajvédelme és az optikai elválasztás alapján. Leggyengébb eredményt (0,7) ért el a télen is zöld pázsit, amelynek még a fenntartási költsége is magas. 4,9-es értéket kapott a vegetációs időben nagy zöldtömeget adó, kevés fenntartást igénylő cserjecsoport. 8,9 értéket ért el a park, ahol a gyep és a cserjék, fák koronaszintje elválik egymástól. Legjobbnak értékelte (17,7) a több szintes biológiailag aktív fás cserjés felületet, ahol a cserjék, a lombos és fenyőfélék intenzíven növekszenek, főleg nyáron jelentős zöldtömeget nevelnek. Ez meghaladta a fenyőerdő (11,2) és a lomberdő (13,2) értékét.
Radó (2001) szerint nagyon fontos szerepet töltenek be a fák a környezetünk védelmében. Ezt alátámasztandóan kiemeli, hogy 1 lombköbméter átlagosan 4 m³ asszimiláló felületnek felel meg, valamint ugyanekkora lombtömeg 4500 g szennyező anyagot képes kiszűrni a levegőből egy vegetációs időszak alatt. Számítások szerint 1 lombköbméter asszimilációs felület 47 l vizet párologtat el egy vegetációs időszakban, ami egy 50 éves fa esetében 4,2 m³-t jelent.
Az összefüggő városi zöldfelületek páratartalma 5-7 %-al meghaladja a beépített, vagy burkolt területen mért értéket. Ezt javítja még, hogy a fák és a cserjék lombjára lehulló csapadék egy része elpárolog anélkül, hogy leérne a talajra. Ezen túlmenően a zöldfelület nem termel port, az ide kerülő légszennyezés jelentős részét megkötik a fás szárú növények (Kiácz és Szendrői, 1980).
4.2 Az örökzöld növények jelentősége
Örökzöldek ültetésével az a célunk, hogy a kertünk télen is díszítsen. Ez volt már gróf Ambrózy-Migazzi István álma is, létrehozni az örökké zöldellő „sempervirens” kertet. Ő egy széles látókörű, sokat utazó ember volt, aki látta a Riviérát és megismerkedett egy-egy mediterrán régió, illetve speciális mikroklímájú térség (például Anglia, Algéria, a Genfi-tó) gazdag növényvilágával is. Hazatérve nem tudott belenyugodni otthonának csaknem féléves korpárságába, ezért már 1892-ben Malonyán „délszaki növényekből” egy gyűjteményes parkot létesített, amely ma az örökzöld fás növények leggazdagabb genetikai állományforrása Közép-Európában (Internet 1.). Amikor az első világháború következményeként malonyai birtokát elvesztette és kénytelen volt az ősi tanai birtokára (Vas megye - Tanakajd ) költözni, elhatározta, hogy a malonyai főpróba után itt a megyében valósítja meg az igazi mesterművet.
Megvásárolta a Kám község határában lévő, akkor „Jelihálásnak” nevezett területet, mely akkor főleg legeltetéssel hasznosított akácos, csarabos, nyíres állapotában létezett. A munkát 1922-ben kezdte el. A változó domborzati viszonyok változatos éghajlatbeli kitettséget, fény-, hő- és páraviszonyokat, egyszóval, mikroklímát nyújtanak és nagyon sokféle növénytársulást hordoznak, illetve hozzájuk illő új növénytársulások kialakítását teszik lehetővé. Ambrózy a munkáknál mindenkor a változatos domborzatot, a talajt és mikroklímát vette figyelembe az odatelepíthető növényközösségek megtervezéséhez. Bőséges növényanyag állt rendelkezésre Tanán, emellett faiskolát és szaporító telepet létesített a behozatott mag és más növényanyag megnevelésére. Ilyen módon a fenyők, tuják, tiszafák, a magyal, a babérmeggy, a borbolyák, kecskerágók, madárbirsek, buxusok tömegeit ültette el, és az így kialakított párás-árnyas környezetbe a rododendronok több tucat fajtáját telepítette be. Emellett a nárciszok több mint 10 változatát, a páfrányok, a sáfrányok, liliomok, összefoglalóan a hagymás, hagymagumós és gyöktörzses növények sok-sok fajtáját ültette széjjel a kert különböző részeibe. Tíz év munkájával a háta mögött már szép, 5 hektárnyi betelepített kerttel, de még messze a megvalósítástól, 1933-ban elragadta a halál (Fábián és Sipos, 1999). Ma az arborétum és az
arborétumot kiszolgáló egységek 106,6 ha védett területet tesznek ki.
Hazánk flórája szegény mind tűlevelű, mind pedig lomblevelű örökzöldekben. Ormos (1967) szerint mivel a télen zöld lomblevelű növények a mi éghajlatunkon különlegességnek számítanak, ezért a felhasználásuk is ennek megfelelően történjen: a kertünk kiemelkedő pontjaira kerüljenek és olyan helyre, hogy az ablakokból télen is gyönyörködhessünk bennük.
Amíg a fenyőfélékből egy átlagos méretű kertbe csupán néhány példány helyezhető el a méretük és a hozzájuk kapcsolódó rossz hangulat (temető) miatt, addig a lomblevelűekből tetszés szerint válogathatunk és ültethetünk anélkül, hogy hátrányos túlzsúfoltság következne be vagy a környező tájtól idegenül hatna (Debreczy és Csapody, 1971).
A növény külső megjelenésében kifejezésre jut belső tulajdonsága, mert származási helyének ökológiai viszonyait tükrözi vissza. Ezért a kertbe olyan növénycsoportokat kell kialakítani, amelyek ökológiailag azonos értékűek és megfelelő társítással az esztétikai hatás fokozására alkalmasak (Lange, 1928).
A mediterrán tájat lomblevelű örökzöldek és az aromás illatú örökzöld félcserjék tömeges előfordulása jellemzi. A fenyőfák koronája lazán szétálló (atlaszcédrus) vagy ernyőszerűen ellapuló (pínea fenyő, libanoni cédrus). Itt nem kell tartani a hótöréstől, hónyomástól. E fenyők nagy része (pl. a píneafenyő) nálunk már nem télálló. Helyette az idős korban ugyancsak ellapuló feketefenyőt vagy más merev, hosszú tűjű fajokat (sárgafenyő, Jeffrey fenyő) ültethetjük. Jellemző még a határozott függőleges tagolást adó oszlopciprus (Cupressus sempervirens ’Stricta’, ősi kultúrforma!). Nálunk, ha nem elég meleg a kertünk, a nyugati tuja hasonló megjelenésű oszlopváltozataival (Thuja occidentalis ’Columna’,
’Malonyana’, ’Smaragd’) vagy magasra növő pikkelylevelű oszlopborókákkal (Juniperus virginiana ’Pseudocupressus’ ) érhetünk el hasonló hatást (Schmidt 2003).
A lombos fák közül jól érvényesülnek az ernyős koronájúak (júdásfa, papíreperfa, mézesfa), a régi kultúrvidéket jelző gömbkoronák (pl. a csillogó lombú gömbmeggy), valamint a velük szép kontrasztot adó oszlopkoronák. Nagyobb kertben jó hátteret adnak a nagy levelű, vagy fénylő lombú fák (platán, császárfa, csertölgy). A hatást illatos félcserjékkel (levendula), nyáron virágzó cserjékkel (barátcserje, nyári orgona), télálló pálmaliliomokkal, valamint néhány különleges igényű, déli származású díszfa, díszcserjével (gránátalma, füge) tehetjük teljesebbé. A mediterrán kert a száraz, forró táj illúzióját kelti (Schmidt 2003).
A botanikai terminológia szerint örökzöld az a növény, amelyik egy évnél tovább (tehát az új levelek kihajtása után is) megtartja a lombját. Az új hajtásokkal a régi lombot leváltó növényeknek a hivatalos neve a télizöld, azok pedig, amelyeknek a puha levelei a tél közepére többnyire elfagynak a félig örökzöld növények. A gyakorlat viszont örökzöldnek
nevez minden olyan növényt, aminek a levele zölden telel át (Schmidt, 2003).
4.2.1 A lomblevelű örökzöldek származása
Hazánk flórája szegény lomblevelű örökzöldekben. Ennek csak részben oka az éghajlati adottság. A jégkorszak előtt Közép-Európa növényzete a jelenlegihez hasonló éghajlati viszonyok között sokkal fajgazdagabb volt, amiben lomblevelű örökzöldek is jócskán előfordultak. A több hullámban bekövetkező eljegesedés azonban megtizedelte a gazdag jégkorszak előtti flórát. Pusztító hatását fokozta, hogy a lehűlések elől délre
„menekülő”, majd az eljegesedési hullámot követő felmelegedés miatt ismét északra húzódó növények vándorlását akadályozták a kelet-nyugati irányú hegyláncok, az Alpok, a Kárpátok, a Balkán és a Pireneusok (Schmidt, 2003). A hideg elsősorban a fás szárú növényeket pusztította, mivel azok jobban ki voltak téve a fagyhatásnak, mint az évelők és az egynyáriak (Józsa, 1993).
A Földnek vannak részei, amelyek nagyon gazdagok örökzöldekben. Legtöbbjük a trópusokon, a szubtrópusi területeken és a mediterránban él. Az első két terület számunkra érdektelen, mivel az ott élő növények nem viselik el a hazai teleket. A mediterrán területen a nyár száraz és meleg, télen sok a csapadék, de nagy hidegek nem fordulnak elő. Ilyen típusú terület a Földközi-tenger térsége, valamint Kalifornia nagy része. Ezeken a területeken nagyon sok ún. keménylombú örökzöld növény él. A mediterrán terület északi részén, ahol nagy hidegek is előfordulnak, számtalan örökzöld él, amelyeket eredményesen nevelhetünk a mérsékelt égövben is (pl. Pyracantha, Cistus). Számunkra a legfontosabbak a mérsékelt égövben előforduló örökzöldek (Józsa, 1993). Észak-Amerikában a Sziklás-hegység észak- déli vonulatának köszönhetően a jégkorszak előtti flóra jórészt fennmaradt. A hegység vonulata melletti hatalmas prérin a növényzet akadálytalanul vándorolhatott délre, majd ismét északra a több évezredenként váltakozó hideg és meleg elől. Kelet-Ázsiában pedig, a Himalájától délre gyakorlatilag nem volt jégkorszak, a lehűlést fékezték a Csendes-óceán meleg áramlatai, a jégárakat és a fagyos szeleket pedig felfogták az irdatlan hegylánc égbe nyúló csúcsai. Észak-Amerika és Kelet-Ázsia növényzete hasonló éghajlat mellett ma sokkal több fajból áll, mint az európai növényzet. Éppen ezért nem csoda, hogy a kertjeinkben meghonosodott exota fák és cserjék többsége ebből a két géncentrum valamelyikéből származik. Ez egyaránt vonatkozik a lombhullatókra, valamint a tű- és lomblevelű örökzöldekre. A lomblevelű örökzöldek közül a közönséges mahónia (Mahonia aquifolium) észak-amerikai, az örökzöld madárbirsek (Cotoneaster) és az örökzöld borbolyák (Berberis)
pedig csaknem kivétel nélkül Kelet-Ázsiában, főleg Kínában és Japánban honosak (Schmidt, 2003).
4.2.2 A lomblevelű örökzöldek igényei
A lomblevelű örökzöldek általában a magyarországinál valamivel melegebb és főképp enyhébb telű térségek növényei. Míg a valódi mediterrán éghajlatot a hosszú, száraz és forró nyár, rövid és csapadékos tél jellemzi, amikor a hőmérséklet többnyire nem süllyed fagypont alá (Turcsányi és Siller, 2005), addig a forró, csapadékszegény nyár, a rövid tél, a tavaszi és őszi esőzések a szubmediterrán éghajlat jellemzői (Borhidi, 2007). Az ilyen helyekről származó növényeknek az egyik legfontosabb életfeltétele a szabályos periódusosság, azaz hátrányos számukra a „túl hideg” vagy a „túl száraz”, esetleg „túl hideg és túl száraz” tél, vagy a „túl nedves” és a „túl hűvös” nyár (Illés, 1981).
Az asszimilálásra mindig kész lombozat a hosszú vegetációs időszak és a télen is gyakori fagymentes időszakok kihasználásához alkalmazkodott. Nagy részük a lombhullató erdők cserjeszintében él, ahol a fák télen is védik őket az éghajlat szeszélyeitől, a talajt borító avartakaró pedig megakadályozza a gyökérzóna átfagyását.
Nyáron a lomblevelű örökzöldek a lombhullató cserjékhez nagyjából hasonló igényeket támasztanak. A tüskés és tövises fajok általában közepesen szárazságtűrők, a nagylevelű és tövistelen fajok viszont pára- és vízigényesek. Nagy részük félárnyékban érzi jól magát, de napra is telepíthetőek (Schmidt, 2003).
A téli időszak nehezebb a lomblevelű örökzöldek számára. Egy részük a nagy hidegre is érzékeny, de ennél is veszélyesebb számukra a téli napsütés és az ezzel járó nagy hőingadozás, ami élettani szárazságot okoz. A széles, örökzöld levelek, ha rájuk süt a nap, még fagyos időben is „kiengednek”, asszimilálni kezdenek, majd a rákövetkező derült éjszakán annál könnyebben elfagynak. Még nagyobb baj, hogy a napsütés párologtatásra kényszeríti a növényt egy olyan időszakban, amikor a gyökérzet vízutánpótlását a fagyos talaj korlátozza. Gyakoribb ezért, hogy télen az örökzöldek lombja nem lefagy, hanem szabályszerűen leszárad, „leég”. Ha a talajt fehér hó borítja, az erős fényvisszaverődés tovább fokozza a kárt (Schmidt, 2003).
4.2.3 A lomblevelű örökzöldek felhasználási lehetőségei
A nálunk jelenleg telepített lomblevelű örökzöldek kivétel nélkül cserjetermetűek.
Felhasználási területeik ezért általában azonosak a többi díszcserjéével. Van azonban az örökzöldek alkalmazásának néhány ökológiai és esztétikai sajátossága, ami elsősorban az említett növénycsoport természetes előfordulási körülményeiből fakad.
Szoliter cserjének azok a taxonok a legalkalmasabbak, amelyek szép színük mellett különleges formájukkal, levelükkel vagy egyedi virágaikkal is kitűnnek, sajnos azonban a lomblevelű örökzöldek érzékenyek a nagy hidegre és a téli napsütésre, ami széllel párosulva végzetes is lehet számukra. Éppen ezért a kert félárnyékos-árnyékos részeire telepítsük, lehetőség szerint nagy fák alá. Így nem süt rájuk télen a nap, a fák alatt a lehulló lomb is védelmet nyújt. A megoldás hátránya, hogy így az örökzöldeket elrejtjük, holott nagy díszértékük miatt a kert szembetűnő (és többnyire napsütötte) részeire kívánkoznának.
Ha szoliter örökzöld bokrok közé örökzöld talajtakaró cserjéket ültetünk, akkor a talaj téli védelmét élő növényzettel oldjuk meg ahelyett, hogy szalmával, avarral, tőzeggel vagy lombbal takarnánk. Még hatásosabb, ha a szoliter növények helyett kisebb-nagyobb örökzöld foltot telepítünk, ekkor már állományklímáról beszélhetünk (Schmidt, 2003).
4.2.4 A lomblevelű örökzöldek választékának alakulása a Prenornál
Közép-Európa egyik legmeghatározóbb faiskolájának kínálatát elemezve közelebb kerülhetünk a lomblevelű örökzöld taxonok fajtaválasztékának alakulásához hazánkban.
Szombathelyen lévő kertészetek, faiskolák egy részéből alapította Szombathely Város Tanácsa 1949-ben Szombathelyi Kertészeti Vállalatot, ahol csak a későbbi bővítésekkel alakul ki a faiskolai termesztés. 1994-ben a cég felveszi a Prenor Kft nevet.
A lomblevelű örökzöldek választékának alakulását a cég fellelhető árjegyzékei alapján állította össze a szerző. Ezek az árjegyzékek A/5 méretben készültek. Eleinte külön fejezetben szerepeltek az egyes növénycsoportok, így a lomblevelű örökzöldek is. 1980-tól a növények mellett rövidebb-hosszabb leírás van, 1985-től a lomblevelű növények abc sorrendben követik egymást. Az 1975/76-os árjegyzékben vannak színes képek, majd 1990-től minden árjegyzék tartalmaz színes mellékletet.
Az eltelt 90 évben a faiskolában 135 lomblevelű örökzöld taxont forgalmaztak. A kínálat változását tükröző fajokat az 1. mellékletben mutatjuk be.
1953-ban a Vállalathoz csatolják az Ungváry faiskolát, amely már 1924-ben 17 féle örökzöldet (5 Buxust, 2-2 Cotoneastert, Euonymust, az azóta sem előforduló Sarcococca ruscifolia és a Yucca filamentosa) kínált. A faiskola a kezdeti időszakban nagyrész magcsemetéről nevelt kész növényeket. Az első nyomtatott árjegyzék 1967 ősz/68 tavaszra jelenik meg, 18 örökzölddel. Az örökzöldek nagy része magról szaporított, csak néhány könnyen gyökeresedő örökzöld (Buxus sempervirens) van a kínálatban.
A fajtaválaszték folyamatosan változik. Az új fajták megjelenése kiszorította a régi fajokat. A Lonicera nitida-t (1980/81-ben utoljára), a Pyracantha coccinea-t (1980/81), a Prunus laurocerasus-t (1990/91). Rövid ideig szerepelt a kínálatban a Buxus sempervirens
’Fertődi’ és a Cotoneaster salicifolius ’Piros fűzér’.
Folytonos változás figyelhető meg a Prunus laurocerasusnál. Ahogy a divat változott, úgy kerültek be új fajták, és kerültek ki a régiek. Jelenleg a vörösesen fakadó fajták a keresettek, mint például a ’Cippora’, ’Etna’ és a ’Kleopátra’.
Berberis julianae és a Viburnum rhytidophyllum az állandóságot képviseli, folytonosan kínálják ezeket 1966-tól. Vannak jó fajták, amelyeket a bekerülésüktől folyamatosan árusítanak, pl. a Buxus sempervirens ’Suffruticosa’, a Cotoneaster dammeri
’Skogholm’, vagy a Pyracantha ’Orange Glow’.
4.3 A klímaváltozás tendenciái Európában
Az IPCC (Intergovernmental Panel on Climate Change, 2007) Negyedik Értékelő Jelentése 2007-ben megállapította, hogy a Föld éghajlati rendszere globális és regionális szinten is megváltozott az iparosodás kezdete óta, s új, a korábbinál erősebb bizonyítékok utalnak arra, hogy az elmúlt ötven év során megfigyelt felmelegedés döntő része az emberi tevékenységeknek tulajdonítható. Az IPCC Jelentés szerint a klímaváltozás során nő az egyes szélsőséges időjárási események száma és intenzitása. Minél nagyobb mértékű és minél gyorsabb ütemű az éghajlat változása, annál nehezebb az ahhoz való alkalmazkodás.
1901-2005 között Európa legnagyobb részén a hőmérséklet növekedése volt tapasztalható, az éves középhőmérséklet 0,9 °C-kal emelkedett az egész kontinensen (Kjellström, 2004; Alcamo et al., 2007). Ugyanakkor az 1977-2001-es időszakra a vonatkozó trend +0,4 °C/évtized, ami jóval magasabb, mint az átlagos trend (Jones és Moberg, 2003). Az elmúlt 25 évben a trendek magasabbak a közép- és északkelet-európai, valamint a hegyvidéki régiókban (Tank, 2004), a hőmérsékletemelkedés jelentősebb télen, mint nyáron (Wallerstein et al., 2004; Jones és Moberg, 2003). A hőhullámok növekedése miatt a hőmérsékleti változékonyság növekedése is megfigyelhető (Tank és Können, 2003). A csapadék mennyiségének a változása jelentősen eltérő régiónként és évszakonként (Christensen és Christensen, 2007).
4.3.1 A klímaváltozás várható tendenciái Magyarországon
A csapadék mennyiségének a változása
Az elmúlt 100 évben Magyarországon jelentősen csökkent a csapadék mennyisége és úgy tűnik, hogy ez a tendencia a 21. században is folytatódik (Gálos et al., 2007). Christensen et al. (2007) szerint a nyár hazánkban melegebb és szárazabb lesz a globális trendekhez viszonyítva. Az éghajlat-változási forgatókönyvek szerint a száraz évek és a száraz nyarak valószínűsége a 21. század első felében nem lesz nagyobb az 1951-2000-es időszakhoz képest, csak az intenzitásuk növekszik. 2051-2100 között a száraz időszakok száma szignifikánsan megnövekszik, az A1B és A2 forgatókönyvek szerint minden második nyár száraz lesz. Az egymást követő száraz események száma is több lesz, mint a 21. század első felében (Gálos et al., 2007).
A csapadék mennyiségének éves a változása nem szignifikáns, de jelentős és ellentétes
trendek várhatóak a különböző évszakokban. A téli csapadék mennyiségében növekedés várható, míg a nyáron csökkenés (Bartholy és Gelybó, 2007). Az előrejelzett csapadékcsökkenés mértéke nyáron 24-33% (A2), ill. 10-20% (B2), míg a téli csapadéknövekedés mértéke 23-37% (A2), ill. 20-27% (B2) (Bartholy et al., 2007). A 21.
század végére az eddig legszárazabb évszakunk, a tél csapadékban gazdag, és az eddig legcsapadékosabb nyár pedig a legszárazabb évszak lesz (Bartholy és Gelybó, 2007).
A szélsőséges időjárási események intenzitása és gyakorisága várhatóan növekedni fog hazánkban (Bartholy és Gelybó, 2007; Révész, 2008). A legfontosabb extrémitások közé tartozik a tavaszi fagy, az aszály, a hőhullámok és viharok, mint például a szélvihar, a jégeső és heves esőzések (Ladányi et al., 2009).
A hőmérsékleti értékek változása
Az éghajlat-változási forgatókönyvek szerint 2100-ra a Kárpát-medencében a felmelegedés várható mértéke az A2 és a B2 szcenáriók esetében mindegyik évszakban meghaladja a 2,5 °C-ot. A legnagyobb melegedés mindkét szcenárió esetében nyáron várható.
A hőmérséklet maximálisan várható növekedése 4,9-5,3 °C (A2), ill. 4,0-4,4 °C (B2). A minimum hőmérséklet várható növekedése pedig 4,2-4,8 °C (A2), ill. 3,5-4,0 °C (B2) (Bartholy et al., 2007).
Németh és Kalmár (2006) megállapította, hogy az OMSZ állomáshálózatának mérései szerint az utóbbi 10-15 évben (a növekvő hőmérsékleti tendencia mellett) szignifikánsan nagyobb az áprilisi fagyok gyakorisága és erőssége. Az utolsó tavaszi fagyok átlagos határnapja is egyre későbbre tolódik. Mindez azt jelenti, hogy a késő tavaszi fagyok gazdasági kockázata egyre jelentősebb.
Ha a téli hónapokat külön – külön megvizsgáljuk, akkor arra az érdekes következtetésre jutunk, hogy az évek során decemberben inkább hőmérséklet-csökkenés tapasztalható. Januárban a trend igen közel áll az átlaghoz, az emelkedés csekély. Csak februártól mutatható ki szignifikánsan a hőmérséklet-emelkedésre utaló tendencia (Németh és Kalmár 2006).
Az elmúlt 50 évben a minimum hőmérsékletek időbeli alakulásában március elején és végén határozott negatív hőmérsékleti anomália figyelhető meg. Előfordult, hogy március elején -25 °C alatti hőmérsékletet mértek az országban. Az elmúlt 50 évben a Kárpát-medence térségében a tavaszi időszakban április közepén és végén fordult elő két fagyhullám. Ekkor a
fagy előfordulási valószínűsége meghaladja a 10%-ot (Soltész et al., 2006).
4.3.2 Az éghajlat változásának tendenciái Keszthelyen
A csapadék mennyiségének változása
Kocsis (2008) a keszthelyi csapadékadatok elemzése során megállapította, hogy az éves csapadékösszegek esetében az 1871 és 2000 közötti időszakra vonatkozóan nem mutatható ki szignifikánsan a lineárisan csökkenő tendencia, sem az éves adatok változékonyságában bekövetkező módosulás, bár az adatok megoszlása arra utal, hogy a vizsgált időszakban az átlagnál alacsonyabb csapadékösszegű évek voltak túlsúlyban. Mind a mozgóátlagok sora, mind a klímanormálok azt jelzik, hogy a XX. század második felében a csapadék mennyisége csökkent. A klímanormálok képzésének elve alapján tíz évvel csúsztatott 30 éves periódusok lineáris tendenciái közül az 1881-1910 közötti időszak szignifikáns csapadékemelkedést mutatott. Az adatok évszakos bontását tekintve csak a tavasz esetében mutatható ki statisztikailag igazolhatóan a lineáris csapadékcsökkenés. Az évszakos adatok változékonysága egyik évszak esetében sem módosult szignifikánsan. A csapadék éves járásában, az őszi hónapokban jelentkező másodmaximum eltűnni látszik. Október hónap csapadékösszege szignifikáns csökkenést mutat 1871-2000 közötti időszakban. Ezt a 2006-ig kibővített lineáris trendelemzések is megerősítik. A csapadékmentes periódusok számának vizsgálata során megállapíthatjuk, hogy legalább egy 15 napos, vagy kettő 10-14 napos csapadékmentes periódussal kell szembesülniük gazdálkodóinknak egy-egy tenyészidőszak során. Következtetésként megállapította, hogy a globális klímaváltozás egyik következményeként nyilvántartott csapadékcsökkenés Keszthely esetében évszakos szinten érvényesül, bár az éves és havi adatokban is fellelhetőek a módosulás nyomai. A tavaszi csapadékbevétel csökkenése igen kritikusan érintheti a mezőgazdasági termelést, hiszen mind a tavaszi vetésű, mind az őszi vetésű gabonák esetében terméskiesést okozhat a csapadékszegény időjárás.
A hőmérsékleti értékek változása
Kocsis (2008) szerint az évi középhőmérsékletek adatsorában szignifikánsan kimutatható a felmelegedés (0,49 °C/100 év) 1901-2000 között, de a változékonyság módosulása nem. A 2006-ig kibővített adatsor ennél még erősebb felmelegedést jelez (0,58
°C/100 év). A felmelegedés tényét a klímanormálok alakulása is alátámasztja. A
klímanormálok képzésének elve alapján tíz évvel csúsztatott 30 éves periódusok lineáris tendenciái közül az utolsó, 1971-2000-es periódus szignifikáns melegedést jelez (0,3 °C/10 év). Az évszakok közül a nyár esetében szignifikáns hőmérsékletemelkedés tapasztalható (0,61 °C/100 év) 1901-2000. közötti időszakban, míg a többi évszak esetében nem tudott statisztikailag igazolható változást kimutatni. A 2006-ig kibővített adatsor tendenciája az előzőeket megerősíti, sőt az itt tapasztalható emelkedés intenzívebb (0,8 °C/100 év). A nyári és az őszi középhőmérsékletek változékonyságában csökkenést tapasztalt. A havi adatok esetében nem mutatható ki szignifikáns változás.
4.4 A növények télállóságának és fagytűrésének környezeti feltételei és élettani háttere
A növényeknek is folyamatosan reagálniuk kell az őket körülvevő környezet változására. A mérsékelt égövön a fás szárú növények életben maradásának előfeltétele a hőmérséklet szezonális változásaihoz való alkalmazkodás.
A hőmérséklet csökkenésére a növény úgy reagál, ahogyan azt a Le Chatelier-elv (legkisebb kényszer elve) kimondja, vagyis az egyensúlyban lévő rendszer a külső megzavarásra úgy válaszol, hogy a zavarás hatásait minimálisra csökkenti, illetve kompenzálja. Tehát a hidegben kevesebb a metabolikus energia, a víz és a tápanyag felvétele korlátozott, kevesebb bioszintézis zajlik le, az asszimiláció lecsökkent és a növekedés megállt. Minél gyakoribb és hosszabb ideig áll fenn az alacsony hőmérséklet, annál komolyabbak a következményei a növény számára (Larcher, 2003).
Különbséget szokás tenni a hidegérzékenység és a fagytűrés között is. A mérsékelt égövi növényeinkre a fagytűrés kifejezését szokás alkalmazni, míg a trópusokról származóaknál inkább a hidegérzékenységet. A trópusi növények a 0-10 ºC-os hőmérsékleten már letális károsodást szenvednek el (Larcher és Bodner, 1980).
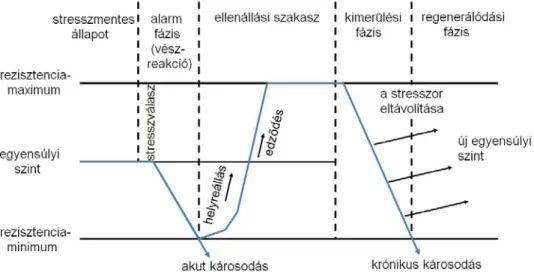
A fagytűrés a növényekben fokozatosan alakul ki és a tél vége felé az időjárás változásával fokozatosan szűnik meg (Pisek és Schiessl, 1947). Weiser (1970) szerint a mérsékelt égövben honos fás szárú növények a szezonális hideghez három lépcsős folyamat során adaptálódnak. Az első lépcső a fotoperiódusra hat erőteljesen. A fás szárú növények a rövid nappalok beköszöntével megszűntetik a növekedést, ami a hideg akklimatizáció első lépése, valamint sok szerves anyag raktározódik el. Ekkor a sejtek a 0 ºC-ot túlélik, de még nem teljesen edzettek. A második lépcsőben az alacsony hőmérséklet idézi elő a hideghez, különösen a 0 ºC alatti hőmérséklethez való alkalmazkodást metabolikus és strukturális
változásokkal. Néhány erdei faj az alacsony hőmérséklethez (-30 és -50 ºC) való kitettség által éri el a hidegtűrésnek a maximális szintjét. Ezt szokták a harmadik lépcsőként definiálni (Weiser, 1970). Tehát az alacsony hőmérséklet és a rövidülő fotoperiódus a két fő faktora a fás szárú növények hideghez való akklimatizációjának. A környezeti tényezők sorrendje lényeges: a rövid napok megelőzik az alacsony hőmérsékletet, hogy az akklimatizáció maximuma megmutatkozhasson (Irving és Lanphear, 1967; Bervaes et al., 1978). Továbbá a hőmérséklet módosíthatja a fotoperiódus hatását a növekedésre és a fejlődésre (Junttila és Kaurin, 1985; Westergaard és Eriksen, 1997), valamint a rendelkezésre álló víz mennyisége, az ásványi tápanyagok és a növény kora is befolyásolhatja az alkalmazkodást (Rollins et al., 1962; Chen et al., 1977; Pellett és Carter, 1981; McNamara és Pellett, 2000). Végül a növénynek belső ritmusa van, amely segít az alkalmazkodásban, de ez független a környezeti tényezőktől (Howell és Weiser, 1970; McKenzie et al., 1974; Siminovitch 1982). A fagytűrés a növények dinamikusan változó tulajdonsága, melyet genetikai és összetett környezeti hatások befolyásolnak (Proebsting, 1970; Szalay, 2001).
A télállóság és a fagytűrés nem összekeverendő fogalom, hanem el kell különíteni őket egymástól. A fagytűrés azt jelenti, hogy a növényeinket az alacsony hőmérséklet nem károsítja, vagyis alkalmasak a növények a fagypont alatti hőmérséklet túlélésére, míg a téltűrés egy komplex fogalom, az alacsony hőmérséklettel szembeni tolerancián kívül magában foglalja a tél egyéb károsító tényezőit is, amelyek közvetve hatnak. A téltűrés egyik jellemzője, hogy az adott faj mennyire viseli el a mélynyugalom alatti lehűlést, valamint a téli hőmérséklet-ingadozást. Hazánkban azoknál a fajtáknál elfogadható a téltűrés, amelyeknél a mélynyugalom hosszú és az alatt elviseli a -20 – -25 ºC közötti lehűlést is (Soltész, 2003).
Schmidt (1979) szerint a téltűrésre kihat a nyári szárazság is.
A mélynyugalomba való átmenet belsőleg szabályozott (Tromp, 2005). Ezt az állapotot egy hosszabb felkészülési időszak előzi meg, amely során tartalék tápanyagok halmozódnak fel, így fokozva a fagy-, illetve télállóságot. A mélynyugalom alatt a virágrügyek semmilyen körülmények között nem hajtanak ki. Az anyagcsere-folyamatok a szövetekben minimálisra csökkennek. A virágrügyek ebben az időszakban a legfagytűrőbbek.
A mélynyugalom akkor ér véget, ha a növény egy bizonyos hidegmennyiséget megkapott. A lombhullató fák rügyeinek endogén nyugalma a hideg hatására fokozatosan megszűnik, de a rügyek az alacsony hőmérséklet miatt továbbra is nyugalomra kényszerülnek. A nyugalom e formáját kényszernyugalomnak nevezzük (Pethő 1998).
Az alacsony hőmérséklet közvetlen károsító hatása a szövetekben lévő víz megfagyásából adódik. Amint a hőmérséklet 0 °C alá süllyed, megindul a sejten kívüli
(extracelluláris) jégképződés. A jég képződése miatt ozmotikus potenciálkülönbség alakul ki a sejten kívüli és a sejten belüli víz között. E potenciálkülönbség miatt a víz a sejtből kiáramlik az intercelluláris térbe, ami a sejt dehidratációjához vezet. Tehát a fagy hatására a sejtekben vízhiány alakul ki (Hoffmann, 2011). Az extracelluláris jégképződés következményeként, amely a sejteket fokozatosan dehidratálja, a sejtnedv koncentrálódik, a plazma zsugorodik, és a térfogatváltozás roncsolja a membránokat (Pethő, 1998). A hideg hatására a membránok folyadékkristályos állapotból szilárd gél halmazállapotúvá válnak, és a fehérjéik kiszakadhatnak, ezáltal megváltozik a struktúrájuk, csökken a membrán-transzport szelektivitása és a membránhoz kötött enzimek aktiválási energiája megnő. (Lyons, 1973;
Levitt, 1980; Larcher, 1985). Az intracelluláris jégképződés a protoplazmát mechanikailag károsítja, ezért ez a sejt számára végzetes (Moor, 1964). A sejten belüli jégképzéshez szükség van egy kristályosodási gócra, ahol elkezdődhet a jégkristály kialakulása. Ezek a leggyakrabban az ún. INA-baktériumok (Ice-Nucleation-Active baktériumok), tehát jégmagképző baktériumok, mint az Erwinia és a Pseudomonas fajai, de lehetnek más biológiai struktúrák, molekulák, vagy szerves és szervetlen hulladékok (Pedryc, 2008).
A jég gyorsan terjed az edénynyalábokban és a homogén szövetekben, de a haladását lassítja a nem folytonos tér, mint pl. a levegő vagy az elfásodott sejtfal (Larcher, 2003).
Ha a felmelegedés gyors, akkor az intercelluláris víz elpárolog és a plazma koagulálódhat, ami szintén végzetes a sejt számára (Pethő, 1998).
A növényi sejtek és szövetek hideghez való alkalmazkodása széleskörű változásokon megy keresztül, ami lehetővé teszi a sejt számára az alacsony hőmérséklet és a fagystressz túlélését. Ennek során csökken a szövetek víztartalma, az ozmotikus potenciál süllyed, és olyan vélelmezett krioprotektív vegyületek halmozódnak fel, mint oldható szénhidrátok és fehérjék (Levitt, 1980; Guy, 1990). A plazmamembránok biokémiai átalakulása növeli a membránok folyékonyságát, a foszfolipidek koncentráltságát és a zsírsavak telítetlenségét (Yoshida és Uemura, 1990). A sejtek strukturális változása a citoplazma méretének növekedésével és a vakuólumok nagyságának csökkenésével jár (Pomeroy és Siminovitch, 1971; Wisniewski és Ashworth, 1986). A lombhullató fák kérgének a sejtjeiben a kloroplaszt aggregálódik ahelyett, hogy a sejtben egységesen eloszlana (Kuroda és Sagisaka, 1993).
A szélsőségesen alacsony hőmérséklet nyomán fellépő fagykár mértékét nemcsak a fagy erőssége, illetve tartama határozza meg, hanem az is, hogy a növény milyen állapotban (mélynyugalom, kényszernyugalom) van. Sokszor abszolút értékben kisebb méretű fagy is okozhat jelentősebb kártételt, ha a növény már ún. fagyérzékeny periódusba (kényszernyugalom) lépett (Soltész et al., 2006).
Az egyik leggyakoribb téli fagykárosodás a farész és a bél barnulása (Larcher, 2003).
A bél elhal, a farész megsötétedik, a kambium és a háncsrész viszont tovább él. Ezt követően a növény tovább él és növekszik, az újonnan képződött farész biztosítja a tápanyag áramlását.
A legérzékenyebb szervek és szövetek közé tartozik a gyökér, a kambiális zóna, a klorotikus és az öregedő levelek, a nyitott rügyek, valamint a magban az embrió (Sakai és Larcher, 1987). A téli napégés, felhőtlen napokon, a törzs délnyugati oldalán, a kéreg gyors hőmérséklet-változása miatt alakul ki. A fagyléc, a törzs hosszanti felrepedése nagyon hideg időben következik be, amikor a belső részek magasabb víztartalma megfagyva szétrepeszti a törzset. A hajtásrügyek fagytűrése a kambiuméhoz hasonló vagy annál jobb, a virágrügyek érzékenyebbek. A tél kezdetén bekövetkező lehűlések nagyobb mértékben károsítják a hajtásrügyet, mivel akklimatizálódásuk később fejeződik be, mint a virágrügyeké (Childers, 1983).
A fagytűrést a faj illetve a fajta örökletes tulajdonságai alapvetően meghatározzák. A fagytűrő-képesség alakulását a télre való felkészülés, a fagy jelentkezésének időpontja, a hőmérséklet-változás üteme, a fás szárú növény anatómiai sajátosságai és a talaj is befolyásolják (Porpáczy, 1964). A növények különböző részeinek nem egyforma a fagytűrése, sőt az egyes szerveken belül eltérés lehet a különböző szövettájak fagytűrő-képességében is.
Emellett a fagytűrés nem statikus, hanem állandóan változik. Egy növény, illetve egy növényi rész fagytűréséről tehát csak hosszabb ideig történő, folyamatos vizsgálattal kaphatunk teljes képet (Szalai, 1994).
A felmelegedés is ugyanolyan fontos szerepet játszik a téltűrésben, mint maga a hideg, meghatározó a növények túlélésében. Ha a felmelegedés hirtelen következik be, nincs elegendő idő arra, hogy a plazmából elvont víz visszaáramoljon, a plazmakolloidok nem tudnak rehidratálódni és a növény élettani szárazságtól elpusztul (Soltész, 2003; Szalai, 1994;
Garancsy, 1999).
Az egyes sejtfunkciók eltérő módon érzékenyek a hőmérsékletre (Kisljuk, 1964). Az első detektálható hatás az alacsony hőmérsékletre, az a protoplazma áramlásának a megszűnése, amelyet közvetlenül szabályoz a légzési folyamatok energia-ellátottsága és a magas energia-tartalmú foszfátok elérhetősége. Csökken a fotoszintézis, amely már korai stádiuma gázcsere-mérő műszerekkel és klorofill-fluorométerrel mérhető, a mérés eredménye figyelmeztető lehet (Mc William és Ferrar, 1974).
A hideghatás szempontjából a fotoszintetikus apparátus a növény egyik legérzékenyebb pontja. Hidegstressz hatására a leveleken klorotikus foltok jelennek meg,
melynek oka, hogy alacsony hőmérsékleten gátolt a klorofill molekulák bioszintézise, az etioplasztok és a kloroplasztiszok fejlődése (Berry és Björkman, 1980; Yoshida et al., 1996;
Böddi et al., 1997).
A hidegérzékenység a genetikai tényezőkön túl függ a fejlettségi állapottól és az anyagcsere aktivitási szintjétől is. A hőmérséklet csökkenése elsősorban a membránokat károsítja. A fagy hatására megszűnik a membránok szemipermeabilitása, elvesztik az aktív iontranszportáló képességüket, a membránbeli foszfolipidek degradálódnak, fázisátmenet történik és megváltozik a membránfehérjék eloszlása laterális mozgásuk következtében. A legnagyobb veszélyt a sejten belüli jégképződés, a jégkristályok kialakulása jelenti. A fagy ezenkívül erős dehidratációt okoz, az így a részben extracellulálissá váló víz viselkedése nagyobb jelentőségű lehet, mint maga az alacsony hőmérséklet. A fagypont alatti hőmérsékletet a növények a fagy kikerülésével vagy eltűrésével vészelik át. A kikerülésre példa a túlhűlés, amikor oldott anyagok akkumulálódnak a sejtben és a citoplazma fagyáspontját lecsökkentik (Leskó, 2005).
A hidegstressz az anyagcsere valamennyi működését befolyásolja, hiszen minden biokémiai folyamatnak a sebessége függ a hőmérséklettől, minden enzim meghatározott hőmérsékleti optimummal rendelkezik. Mivel az egyes anyagcsere folyamatok eltérő mértékben reagálnak a hideghatásra, a sejt metabolikus egyensúlya felborul. Egyes esetekben az alacsony hőmérséklet hatása csak a hidegstressz alatt jelentkezik, a stressz elmúltával az életműködés helyreáll. Más esetekben a stressz tartós károkat okoz, így a csökkent működés a hideghatás elmúltával is kimutatható. Bizonyos esetekben pedig a hidegstressz látható jelei alacsony hőmérsékleten nem, csak azt követően, a normál hőmérsékleten jelentkeznek (post- chilling tünetek) (Janda, 2007).
A fagytűrő képesség komplex biokémiai és biofizikai folyamatok eredménye. Az alacsony hőmérsékleten a metabolikus egyensúly felborulása oxidatív stresszt okoz, így a fagytűrés kialakulásában fontos szerep jut az antioxidáns védekező mechanizmusok indukciójának is. A fagystressz közvetve szárazságstresszel jár együtt (Janda, 2007).
A fiziológiai szárazság több okból jöhet létre. Először is a hideg talajok fiziológiailag szárazak (Pethő, 1998). A talaj lehűlésével csökken a gyökér tápanyag- és vízfelvétele, ezért a lehűlt talajból a növény nem tudja fedezni az elpárologtatott vízmennyiséget, még akkor sem, ha egyébként ott rendelkezésre áll felvehető víz, ezért a növény lankad. Fagyott talajból a növény vízfelvétele teljesen megszűnik (Pethő, 1998). A légmozgás csak tovább csökkenti az amúgy is csökkent víztartalmat, hiszen jelentősen fokozza a vízleadást. A fás szárú növényeknél az ágakat borító pararéteg mérsékli a vízleadást, de a kellően be nem érett, fiatal
ágak, amelyek vékony pararéteggel fedettek, sokat szenvednek, károsodnak.
4.5 A fásszárú dísznövények fagytűrőképességének meghatározása télállósági zónák alapján
A hőmérséklet limitálja a növények termeszthetőségét. Ez az a tényező, amely meghatározza egy faj sikerét egy új élőhelyen, az az edzettsége és a fagykárral szembeni ellenállóképessége, hogy elkerülje a sérülést vagy az elpusztulást az alacsony hőmérsékleten.
A legtöbb faj az éves ciklus során bizonyos mértékű edzettséget mutat az alacsony hőmérséklettel szemben (Larcher és Bauer, 1981). Egyes fajoknál azonban ezek a változások nagyon kicsik és a fagytűrő képességük csak néhány fok, pl. a Kárminpiros délmirtusz (Metrosideros carminea) letális hőmérséklete nyáron -2 °C, télen pedig -5 °C (Warrington és Stanley, 1987). Más fajok, amelyek az amerikai és az eurázsiai kontinens északi felén őshonosak, a fagytűrésük, ill. a fagyérzékenységük közti változás jelentős és számottevő fagytűrés is kialakulhat, pl. sok lombhullató és tűlevelű faj képes elviselni a -30 °C-ot is meghaladó hideget is (Sakai, 1982). Bár ezek a fajok jelentős fagytűréssel rendelkeznek, amelyet tavasszal hirtelen elveszítenek és emiatt hasonló vagy kisebb fagytűrő-képességgel rendelkeznek majd a nyáron, mint az enyhe fagytűrő képességű fajok, éppen ezért a nem- szezonális fagyot, késő tavasszal vagy kora ősszel rosszul tűrik (Larsen, 1978; Menzies és Holden, 1981).
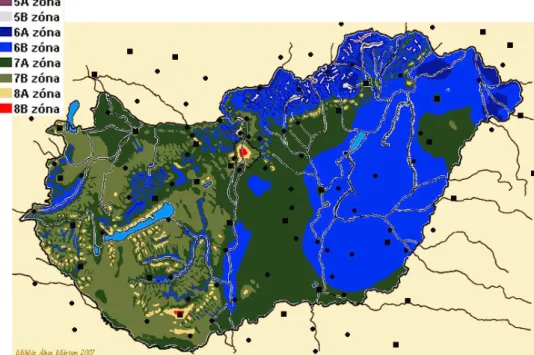
A fásszárú dísznövények téli hidegtűrő képességének megismerésében segítségünkre van a télállósági zónatérkép, az ún. USDA-zónák rendszere, amelyet az 1900-as évek elején az USA-ban dolgoztak ki. Európában először az 1960-as években készítettek télállósági zónatérképet (1. ábra), amely éveken keresztül és több helyszínen történő megfigyelésen alapszik.
1. ábra. Európa USDA zóna térképe (forrás: Internet 2)
Ez hasznos lehet annak eldöntésére, hogy egy dísznövény számára egy adott élőhely megfelelő-e. Az osztályozási rendszer korlátozott, mert az egyes zónák ~6 °C-os tartományt ölelnek fel, amely meglehetősen tág és a zónába sorolás hosszú időt (éveket) igényel (Stanley és Warrington, 1988).
Hazánk a télállósági zóna szerinti besorolás alapján (2. ábra) a 6-os és 7-es zónában, vagyis -12,3 °C és -23,3 °C között helyezkedik el. A legújabb besorolások szerint Pécs-, Budapest- és az egész Balaton környéke a 8a (-12,2 °C) zónába, míg Pécs és a főváros a 8b (- 9,3 °C) zónába tartozik.