VESZPRÉMI EGYETEM
*(25*,.210(= *$='$6È*78'20È1<,.$5
Állattudományi Intézet
Állatélettani és Takarmányozástani Tanszék
DOKTORI (PhD) ÉRTEKEZÉS
Készült a Veszprémi Egyetem Állattenyésztési Tudományok Doktori Iskola keretében
'RNWRUL,VNRODYH]HW MH 7DQV]pNYH]HW
DR.SZABÓ FERENC DR.HUSVÉTH FERENC
egyetemi tanár egyetemi tanár
az MTA doktora az MTA doktora
7pPDYH]HW
DR.DUBLECZ KÁROLY
egyetemi docens
AZ ÉTKEZÉSI TOJÁS ZSÍRSAVÖSSZETÉTELÉNEK ÉS OXIDATÍV STABILITÁSÁNAK BEFOLYÁSOLÁSA
TAKARMÁNYOZÁSSAL
Készítette
P
ÁLL
ÁSZLÓKESZTHELY 2003
AZ ÉTKEZÉSI TOJÁS ZSÍRSAVÖSSZETÉTELÉNEK ÉS OXIDATÍV STABILITÁSÁNAK BEFOLYÁSOLÁSA
TAKARMÁNYOZÁSSAL
Értekezés doktori (PhD) fokozat elnyerése érdekében Írta:
PÁL LÁSZLÓ
Készült a Veszprémi Egyetem Állattenyésztési Tudományok Doktori Iskola keretében
7pPDYH]HW 'U'XEOHF].iURO\HJ\HWHPLGRFHQV&6F
Elfogadásra javaslom igen / nem ……….
A jelölt a doktori szigorlaton %-ot ért el
Keszthely, 2003. január 29. ……….
Szigorlati Bizottság elnöke Az értekezést bírálóként elfogadásra javaslom
Bíráló neve: igen / nem ……….
Bíráló neve: igen / nem ……….
A jelölt az értekezés nyilvános vitáján %-ot ért el
Keszthely, 2003 ……….
Bíráló Bizottság elnöke A doktori (PhD) oNOHYpOPLQ VtWpVH«««««
………
Az EDT elnöke
TARTALOMJEGYZÉK
1. BEVEZETÉS 9
2. IRODALMI ÁTTEKINTÉS 11
2.1. A zsírsavak felépítése és funkciója 11
2.2.A többszörösen telítetlen zsírsavak 12
2.2.1. Szerkezet és csoportosítás ... 12
2.2.2. Metabolizmus és fontosabb funkciók ... 14
2.2.3. Természetes források... 16
2.2.4. A humán fogyasztás és szükséglet alakulása... 16
2.2.5. Az n-6/n-3-as zsírsavarány és az emberi egészség ... 18
2.3.Az étkezési tojás zsírsavösszetételének módosítása ... 20
2.3.1. A zsírok szerepe a tojótyúk takarmányozásában ... 20
2.3.2 A tojótyúk lipidmetabolizmusának jellegzetességei ... 21
2.3.3. Az n-3-as zsírsavak hatása a tyúk lipidmetabolizmusára ... 23
2.3.4. A tojás lipidjei és zsírsavösszetétele... 25
2.3.5. Az n-3-as zsírsavak hatása a tojástermelési paraméterekre ... 26
2.3.6. A takarmányozás hatása a tojás zsírsavösszetételére... 28
2.3.7. Az n-3-as zsírsavak és a tojás organoleptikus tulajdonságai ... 31
2.3.8. A tojótyúkok fajtája és életkora... 33
2.3.9. Az n-3-as zsírsavakban gazdag tojás élettani hatásai ... 34
2.4.Az étkezési tojás oxidatív stabilitásának befolyásolása ... 35
2.4.1. A lipidperoxidáció és az antioxidáns rendszer ... 35
2.4.2. Az E-vitamin ... 38
2.4.3. A tojás antioxidáns védelme... 40
2.4.4. A takarmány zsírkiegészítése, a tárolás és a feldolgozás hatása... 41
2.4.5. A tojás oxidatív stabilitásának fokozása természetes antioxidánsokkal... 43
3. ANYAG ÉS MÓDSZER ... 45
3.1. Kísérleti állatok és takarmányok ... 45
3.1.1. 1. kísérlet ... 45
3.1.2. 2. kísérlet ... 48
3.1.3. 3. kísérlet ... 50
3.2. Vizsgált paraméterek, mintavételi és kezelési módszerek... 53
3.2.1. 1. és 2. kísérlet... 53
3.2.2. 3. kísérlet ... 53
3.3. Analitikai módszerek ... 55
3.3.1.A takarmányminták sav- és peroxidszámának meghatározása ... 55
3.3.2. A takarmány- és a tojássárgája-minták zsírsavösszetételének meghatározása ... 55
3.3.3. A takarmány- és tojássárgája-minták A- és E-vitamin- tartalmának meghatározása ... 56
3.3.4. A tojássárgája-minták oxidatív stabilitásának meghatározása... 57
3.3.5. A vérminták egyes paramétereinek meghatározása... 57
3.4. Statisztikai módszerek ... 58
4. KÍSÉRLETI EREDMÉNYEK ... 59
4.1. 1. kísérlet ... 59
4.1.1. A kísérleti takarmánykeverékek sav- és peroxidszáma ... 59
4.1.2. A tojástermelés fontosabb mutatószámai... 59
4.1.3. A tojássárgája összlipidtartalma és zsírsavösszetétele... 61
4.2. 2. kísérlet ... 65
4.2.1. A kísérleti takarmánykeverékek sav- és peroxidszáma ... 65
4.2.2. Tojástermelési paraméterek ... 65
4.2.3. A tojássárgája összlipidtartalma és zsírsavösszetétele... 67
4.2.4.A tojássárgája E-vitamin-tartalma ... 70
4.2.5. A tojássárgája A-vitamin-tartalma... 71
4.2.6. A tojássárgája oxidatív stabilitása ... 72
4.3. 3. kísérlet ... 73
4.3.1. Tojástermelési mutatószámok ... 73
4.3.2. A vérplazma glükózkoncentrációja ... 74
4.3.3. A vérplazma trigliceridszintje ... 75
4.3.4. A vérplazma együttes VLDL- és LDL-szintje... 75
5. AZ EREDMÉNYEK ÉRTÉKELÉSE... 79
5.1. 1. kísérlet ... 79
5.2. 2. kísérlet ... 83
5.3. 3. kísérlet ... 92
6. KÖVETKEZTETÉSEK, JAVASLATOK ... 99
7. ÚJ KUTATÁSI EREDMÉNYEK ... 101
8. ÖSSZEFOGLALÁS ... 102
9. SUMMARY... 107
10. TUDOMÁNYOS KÖZLEMÉNYEK JEGYZÉKE ... 112
11. IRODALOMJEGYZÉK ... 117
KÖSZÖNETNYILVÁNÍTÁS... 142
KIVONAT
AZ ÉTKEZÉSI TOJÁS ZSÍRSAVÖSSZETÉTELÉNEK ÉS OXIDATÍV STABILITÁSÁNAK BEFOLYÁSOLÁSA TAKARMÁNYOZÁSSAL
$ V]HU] PXQNiMD VRUiQ D] Q-3-as többszörösen telítetlen zsírsavakban
JD]GDJ WRMiV HO iOOtWiViKR] NDSFVROyGy WDNDUPiQ\R]iVL YL]VJiODWRNDW
végzett. Az értekezés három kísérlet eredményeit ismerteti. A kísérletek folyamán a tojótyúkok termelési paramétereinek, a tojássárgája zsírsavösszetételének, A- és E-vitamin-tartalmának, oxidatív stabilitásának, valamint a vérplazma glükóz-, triglicerid- és lipoproteinszintjének mérésére került sor.
Az eredmények alapján a tojótyúkok takarmányának az n-3-as zsírsavakban gazdag lenolajjal vagy csukamájolajjal 2%-EDQ W|UWpQ
kiegészítése három hét etetés után a táplálkozásélettani ajánlásoknak
PHJIHOHO Q-6/n-3-as zsírsavarányt eredményezett a tojássárgájában. Az n-6-os linolsavban gazdag tökmagolaj 2 és 4%-os kiegészítési szinten a tojástermelési paramétereket nem rontotta, de a tojássárgája
]VtUVDY|VV]HWpWHOpWNHGYH] WOHQLUiQ\EDPyGRVtWRWWD
A 4% csukamájolajat tartalmazó takarmányhoz adott 30 és 60 NE/kg E-vitamin kiegészítések nagyságukkal arányosan fokozták a tojás TBARS értékekkel jellemzett oxidatív stabilitását. A tojássárgája A-vitamin- tartalma, az E-vitamin-tartalommal ellentétben, nem volt összefüggésben a tojássárgája oxidatív stabilitásával.
Az eredmények szerint a tojótyúkok takarmányának n-6-os zsírsavakban gazdag olajkiegészítése befolyásolhatja a vérglükózszint
LQ]XOLQUDpVJOXNDJRQUDDGRWWYiODV]UHDNFLyMiQDNMHOOHP] LW$WDNDUPiQ\
n-3-DV ]VtUVDYDNEDQ JD]GDJ RODMNLHJpV]tWpVpQHN LO\HQ MHOOHJ KDWiVDL
nincsenek.
ABSTRACT
NUTRITIONAL MANIPULATION OF THE FATTY ACID COMPOSITION AND OXIDATIVE STABILITY OF TABLE EGG
The author performed nutritional investigations related to the production of table eggs rich in n-3 polyunsaturated fatty acids. The thesis demonstrates three experiments. The results show that the
supplementation of diets of laying hens with 2% linseed or cod liver oil rich in n-3 fatty acids after three weeks feeding period resulted in an optimal ratio of n-6 to n-3 fatty acids of egg yolk. Pumpkin seed oil rich in n-6 type linoleic acid at the inclusion level of 2 and 4% did not reduce the performance parameters of hens but, led to an unfavourably fatty acid composition of egg yolk. Oxidative stability of egg yolk expressed in TBARS values was improved proportionally by dietary supplementation of vitamin E at the level of 30 and 60 IU/kg, when diets contained 4% cod liver oil. In contrast to vitamin E, vitamin A content of egg yolk was not related to the oxidative stability of egg yolk. Supplementation of diets with oils rich in n-6 fatty acids may alter responsiveness of plasma glucose level to insulin and glucagon. Feeding diets supplemented with oils high in n-3 fatty acids do not have similar effects.
AUSZUG
UNTERSUCHUNGEN ZUR BEEINFLUSSUNG DES FETTSÄUREMUSTERS UND DER OXIDATIONSSTABILITÄT VON SCHALENEI MITTELS FÜTTERUNG
Der Autor führte Fütterungsversuche in Zusammenhang mit der Produktion von Schaleneiern reich an mehrfach ungesättigten Fettsäuren des Typs n-3 durch. Die These demonstriert drei Versuche. Die Ergebnisse zeigen, dass eine Supplementierung von Leinöl oder Dorschöl reich an n-3 Fettsäuren in der Höhe von 2% zum Legehennenfutter führte zu einem optimalen Verhältnis von n-6 und n-3 Fettsäuren des Eidotters.
Bei einer Zulage von 2 und 4% Kürbiskernöl reich an Linolsäure (Typ n- 6) zur Ration wurden die Legeleistungsmerkmale nicht verringert, aber ein ungünstiges Fettsäuremuster des Dotters wurde ermittelt. Die Oxidationsstabilität des Dotters anhand der TBARS-Werte war proportional zur Zulage von 30 und 60 IE/kg Vitamin E in der Dorschölgruppe verbessert. Im Gegensatz zum Vitamin E-Gehalt lag für Vitamin A-Gehalt keine Beziehung zur Oxidationsstabilität des Dotters vor. Die Supplementierung von Ölen reich an n-6 Fettsäuren zum Futter kann die Reaktion des Glukosespiegels im Blutplasma gegen Insulin und Glukagon beeinflussen. Die Zulage von Ölen reich an n-3 Fettsäuren zur Ration hat keinen ähnlichen Einfluss zur Folge.
1. BEVEZETÉS
A modern ipari társadalmak kialakulásával a táplálkozási szokások
MHOHQW VHQ iWDODNXOWDN D G|QW HQ PH] JD]GDViJL WHUPHOpVW IRO\WDWy
közösségek étkezési szokásaihoz képest. Kevesebb rostban dús zöldséget, gyümölcsöt, gabonafélét, viszont több cukrot fogyasztunk. A mozgásszegény életmódot folytatóknál gyakran a szükségesnél nagyobb
D] HQHUJLDEHYLWHO DPHO\ HOV VRUEDQ D ]VtURNEyO V]iUPD]LN $
magyarországi halálozás csaknem háromnegyed részét a táplálkozással és
pOHWPyGGDO |VV]HIJJ V]tY- és érrendszeri, valamint rosszindulatú daganatos betegségek okozzák (Zajkás, 1999).
Az utóbbi 10-15 év kutatásai alapján az elfogyasztott összes zsír mennyiségén, a zsírok telített és telítetlen zsírsavainak arányán túl az n-6- os és n-3-as többszörösen telítetlen zsírsavak arányát is FpOV]HU
figyelembe venni az egészséges táplálkozás érdekében. A fejlett országok többségében az n-3-DV ]VtUVDYDNQDN D] DMiQORWWQiO NLVHEE PpUWpN IRJ\DV]WiVD D MHOOHP] ( ]VtUVDYFVRSRUW NHGYH] KDWiViW PXWDWWiN NL
többek között egyes keringési, tumoros illetve gyulladással járó megbetegedések esetén (Wiseman, 1997; Simopoulos, 1999; Lewis és mtsai, 2000). Az n-3-as zsírsavak azonban a lipidperoxidációra rendkívül hajlamosak, antioxidáns kiegészítés nélküli fokozott felvételük a szervezet antioxidáns rendszerét terheli (Simopoulos, 1991). Számos
EHWHJVpJ KiWWHUpEHQ IHOWpWHOH]KHW HQ D] DQWLR[LGiQV UHQGV]HU HJ\HQV~O\iW pULQW R[LGDWtYVWUHVV]iOODSRWK~]yGLNPHJ(VWHUEDXHU/DSHQQDpV
mtsai, 1998).
Az n-3-as többszörösen telítetlen zsírsavak leggazdagabb természetes forrásai D]VtURV´ WHQJHUL KDODN 6RN RUV]iJEDQQHKH]HEE HOpUKHW VpJN PDJDViUXNpVDIRJ\DV]WyLV]RNiVRNQHPWHV]LNOHKHW YpKRJ\D]pWHOHN NHGYH] EE Q-6/n-3-DV ]VtUVDYDUiQ\iKR] KR]]iMiUXOMDQDN /HKHW VpJ YDQ
azonban szélesebb körben fogyasztott élelmiszerek, mint például a tojás vagy a baromfihús zsírsavösszetételének takarmányozás segítségével
W|UWpQ PyGRVtWiViUD +DUJLV pV 9DQ (OVZ\N $ V]HUYH]HW
antioxidáns rendszerével kapcsolatban álló egyes elemek és vitaminok koncentrációja (pl. szelén, E-vitamin) az n-3-DV ]VtUVDYDNNDO HJ\LGHM OHJ Q|YHOKHW D WRMiVEDQ tJ\ D] HJpV]VpJPHJyYy IXQNFLRQiOLV pOHOPLV]HUUp
válhat (Bárdos és mtsai, 1999). A témával gazdag nemzetközi irodalom foglalkozik, s hazánkban is számos kísérletet végeztek e területen (Kovács és mtsai, 1998, 2003; Csuka és Baumgartner, 1999; Husvéth és
PWVDL $ NOI|OG|Q PiU SLDFL IRUJDORPEDQ OpY Q-3-as” (más néven ,,omega-3-as”) tojás hazánkban még nem kapható. Kutatómunkánk
D EHOI|OG|Q HOpUKHW Q-3-DV ]VtUVDYIRUUiVRNUD pSO LVPHUHWDQyagot
NtYiQMD E YtWHQL KR]]iMiUXOYD H WHUPpN PLQpO VLNHUHVHEE KD]DL HO iOOtWiViKR]
9L]VJiODWDLQNVRUiQDN|YHWNH] FpONLW ]pVHNHWIRJDOPD]WXNPHJ
1. kísérlet
• $ 0DJ\DURUV]iJRQ LV HOpUKHW OHKHWVpJHV Q-3-as zsírsavforrások közül a lenolaj és a csukamájolaj két kiegészítési szintjének (2 és 4%) összehasonlítása a tojástermelési paraméterek alakulása és a tojás zsírsavösszetételének javítása szempontjából.
• A tojótyúkok takarmányozásában eddig még nem használt, n-6-os zsírsavakban gazdag tökmagolaj alkalmazhatóságának értékelése, tekintettel a tojástermelésre és a tojás zsírsavösszetételére.
2. kísérlet
• $ WDNDUPiQ\KR] DGRWW NO|QE|] V]LQW (-vitamin kiegészítések hatásának vizsgálata az n-3-as zsírsavakkal dúsított tojás zsírsavösszetételére, A- és E-vitamin-tartalmára, illetve oxidatív stabilitására
• A tojás A-vitamin-tartalma és oxidatív stabilitása közti összefüggés megállapítása.
3. kísérlet
• 9L]VJiOQL NtYiQWXN KRJ\ D] HOWpU Q-6/n-3-as zsírsavarányú takarmányok mennyiben befolyásolják a vérplazma glükóz- és trigliceridkoncentrációjának, valamint nagyon alacsony (VLDL) és
DODFVRQ\/'/V U VpJ OLpoprotein-szintjének változását inzulin- és
JOXNDJRQNH]HOpVWN|YHW HQ
2. IRODALMI ÁTTEKINTÉS
2.1.A ZSÍRSAVAK FELÉPÍTÉSE ÉS FUNKCIÓJA
$ ELRNpPLD QHYH]pNWDQiEDQ D OLSLG IRJDORP iOODWL YDJ\ Q|YpQ\L HUHGHW YiOWR]DWRVV]HUNH]HW DSROiURVROGyV]HUHNEHQ(pl. kloroform, éter) oldódó szerves vegyületeket jelöl (Stryer, 1988). A lipidek csoportjába tartoznak
D] HJ\V]HU ]VtURN YDJ\ JOLFHULGHN DPHO\HN KRVV]DEE V]pQOiQF~ WHOtWHWW
vagy telítetlen zsírsavaknak glicerinnel alkotott észterei. Az összetett foszfo-, gliko- pV V]ILQJROLSLGHN V]WHURLGRN pV NDURWLQRLGRN LV D] pO
szervezetek lipo anyagaihoz sorolhatók. Az olaj kifejezés a
V]REDK PpUVpNOHWHQ IRO\pNRQ\ ]VtURN PHJMHO|OpVpUH V]ROJiO iP NpPLDL
értelemben különválasztásuk nem indokolt. A takarmányozástanban
KDV]QiOW Q\HUV]VtU IRJDORP D WDNDUPiQ\EyO pOHOPLV]HUE O H[WUDNFLyYDO NLQ\HUKHW |VV]HVOLSLGHWWDNDUMD
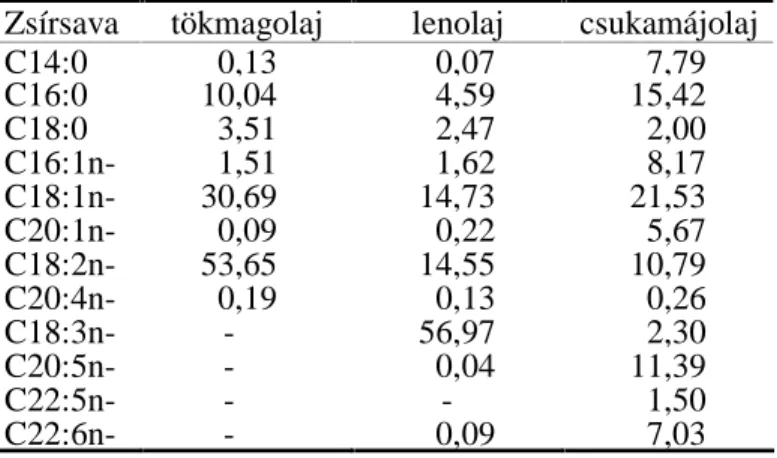
Néhány zsírsav szerkezeti felépítését az 1. ábra szemlélteti. A
]VtUVDYDN W|EEVpJH V]LQWp]LVNMHOOHJpE O DGyGyDQ SiURV V]pQDWRPV]iP~
a szénhidrogén lánchoz két végén metil illetve karboxil csoport
NDSFVROyGLN $] iOODWL V]HUYH]HWHW IHOpStW OHJJ\DNRULEE ]VtUVDYDN -22 szénatomot tartalmaznak (Gurr, 1996). A telített zsírsavak szénláncában csak egyszeres kötések találhatók, míg a telítetlen zsírsavak egy vagy
W|EE NHWW V N|WpVW LV WDUWDOPD]QDN tJ\ HJ\V]HUHVHQ YDJ\ W|EEV]|U|VHQ WHOtWHWOHQHN$NHWW VN|WpVHNV]iPiQpVHOKHO\H]NHGpVpQW~ODNHWW VN|WpV
geometriai konfigurációja (cisz és transz helyzet) is meghatározó
MHOHQW VpJ 6WU\HU&LV]NHWW VN|WpVHVHWpQDNpWKLGURJpQDWRPD
molekula azonos oldalán helyezkedik el, míg transz konfigurációban a molekula ellentétes oldalán található. A geometriai izoméria a zsírsavak
IRUPiMiW pV IL]LNDL MHOOHP] LW LV EHIRO\iVROMD D FLV] ]VtUVDYDN OiQFD
meghajlik, a transz zsírsavaké a telített zsírsavakéhoz hasonlóan egyenes.
$ V]pQOiQFRW IHOpStW V]pQDWRPRN Q|YHNY V]iPR]iVD D NDUER[LOFVRSRUW
irányából a metilcsoport felé halad. A zsírsavak jelölésében a szénatomok
V]iPDXWiQNHWW VSRQWWDOHOYiODV]WYDiOODNHWW VN|WpVHNV]iPD$NHWW V
kötések száma után álló n-x (vagy ω-x) kifejezések azt mutatják, hogy az
XWROVy NHWW V N|WpV D OiQFYpJL PHWLOFVRSRUWWyO [ V]pQDWRPQ\L WiYROViJUD
helyezkedik el.
Az állati és emberi szervezetben megtalálható zsírsavak strukW~UiOLV DONRWyL D VHMWPHPEUiQRN NHWW V UpWHJ DODSV]HUNH]HWpW
létrehozó foszfolipideknek (Gurr és Harwood, 1991). Meghatározzák a
PHPEUiQ IL]LNDL MHOOHP] LW D OLSLG-fehérje interakciókat, így hatással lehetnek a transzportfolyamatokra, a hormonok és neurotranszmitterek receptoraira (Nicolas és mtsai, 1991). Prekurzorául szolgálnak sok biológiailag aktív molekulának (pl. eikozanoidok), befolyásolják bizonyos enzimfehérjék génjének expresszióját (Clarke, 2000). A triglicerideket alkotó zsírsavak feladata els VRUEDQ D] HQHUJLDUDNWiUR]iV
$ KLGURJpQEHQ JD]GDJ ]VtUVDYDN QDJ\PpUWpN UHGXNiOWViJD
következtében a zsírok energiatartalma közel kétszerese a glikogén energiatartalmának. Mindezeken túl a trigliceridek hidrofób jellegüknél fogva majdnem vízmentes, dehidratált formában raktározhatók, ellentétben a poláros, hidratált fehérjékkel és szénhidrátokkal (Stryer, 1988). A dehidratált zsír 1 g-ja így körülbelül hatszor annyi energiát képes tárolni, mint 1 g hidratált glikogén, ami megmagyarázza a zsírnak a glikogénQHO V]HPEHQL HYRO~FLyV HO Q\pW pV HQHUJLDUDNWiUNpQW YDOy
elterjedését.
1. ábra. Néhány zsírsav szerkezeti felépítése (Gurr, 1996) 2.2. A TÖBBSZÖRÖSEN TELÍTETLEN ZSÍRSAVAK
2.2.1. Szerkezet és csoportosítás
A természetben leggyakoribb többszörösen telítetlen zsírsavak (polyunsaturated fatty acids, PUFA) 2-NHWW VN|WpVWWDUWDOPD]QDN*XUU pV +DUZRRG 1DJ\REE KiQ\DGEDQ D PHPEUiQRNDW IHOpStW
foszfolipidekben vannak jelen. A növényekkel ellentétben az állati és az
COOH
COOH
COOH
COOH CH3
CH3
CH3
CH3
sztearinsav C18:0
olajsav C18:1n-9
linolsav C18:2n-6
-linolénsav C18:3n-3
1
1
1
1 18
18
18
18
10 9
10
13 12 9
9 10 16 15 13 12
HPEHUL V]HUYH]HW QHP NpSHV NHWW V N|WpVHN NLDODNtWiViUD D ]VtUVDYOiQF Q- 3-as és n-6-os pozíciójában (Bezard és mtsai, 1994). E zsírsavak nélkülözhetetlen, esszenciális jellegét két amerikai kutató, Burr és Burr (1929) fedezte fel patkányokkal folytatott kísérleteik során. A kialakult
KLiQ\WQHWHN PHJV]QWHWKHW N LOOHWYH PHJHO ]KHW N YROWDN EL]RQ\RV
speciális zsírsavak adagolásával, amelyek közül a linolsav (C18:2n-6) volt a leghatékonyabb. A zsírsavcsoportot elnevezték F-vitaminnak, bár ma már az esszenciális zsírsavak elnevezés az általános. Az n-3-as családba tartozó zsírsavak nélkülözhetetlen struktúrális szerepét bizonyos szövetekben (agy, retina, ivarszervek) Neuringer és mtsai (1988) írták le.
A többszörösen telítetlen zsírsavak közül jelenleg a linolsavat és az - linolénsavat (C18:3n-3) tartjuk esszenciálisnak abban az értelemben, hogy a takarmányban vagy táplálékban jelen kell lenniük bizonyos mennyiségben. A linolsavból kiindulva ugyanis a n-6-RV D] - linolénsavból kiindulva pedig az n-3-as zsírsavcsaládok további tagjai
NpS] GKHWQHN HJ\PiVW N|YHW GHV]DWXUiFLyV pV OiQFKRVV]DEEtWiVL
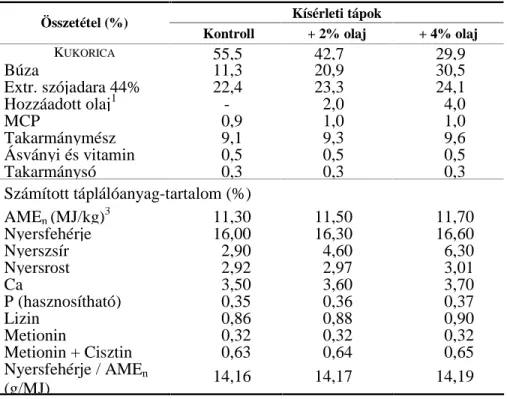
reakciók során, amelyeket a 2. ábra szemléltet (Sprecher, 1981).
2. ábra. A többszörösen telítetlen zsírsavak metabolizmusa
n-9 n-6 n-3
olajsav 18:1n-9
linolsav 18:2n-6
α-linolénsav 18:3n-3
18:2n-9 γ-linolénsav
18:3n-6
oktadekatetraénsav 18:4n-3 20:2n-9 dihomo-γ-linolénsav
20:3n-6
eikozatetraénsav 20:4n-3
20:3n-9 arachidonsav
20:4n-6
eikozapentaénsav 20:5n-3 dokozatetraénsav
22:4n-6
dokozapentaénsav 22:5n-3 dokozahexaénsav
22:6n-3
∆6-deszaturáció
∆5-deszaturáció
∆4-deszaturáció vagy retrokonverzió elongáció(+C2)
elongáció (+C2) vagy retrokonverzió Zsírsavcsalád
Reakció
dokozapentaénsav 22:5n-6
(Gurr, 1999a; Bezard és mtsai, 1994 alapján)
A 18 szénatomnál hosszabb zsírsavak ezért inkább feltételesen esszenciálisak, mert az átalakítási folyamatok zavarai vagy optimálisnál kisebb hatékonysága esetén ezek is a táplálék nélkülözhetetlen alkotóivá
OpSKHWQHN HO *XUU D $ V]HUYH]HWEHQ EHW|OW|WW QpKiQ\ IXQNFLy
kifejezetten az n-6-os vagy n-3-DVFVDOiGPHJKDWiUR]RWWWDJMDLKR]N|W GLN
(funkcionális esszencialitás), továbbá a két zsírsavcsoport közti átalakulás nem lehetséges (Sprecher, 1981). A PUFA harmadik családja az olajsavból származtatható n-9-es zsírsavak csoportja, amelyek azonban az olajsav de novo V]LQWp]LVH PLDWW QHP HVV]HQFLiOLV MHOOHJ HN %H]DUG pV
mtsai, 1994).
2.2.2. Metabolizmus és fontosabb funkciók
A hosszú szénláncú PUFA bioszintézisének lépéseit ugyanaz az enzimrendszer végzi mind a három zsírsavcsalád esetében (2. ábra). A linolsav és az -OLQROpQVDY HJ\PiVVDO NRPSHWHQV V]XEV]WUiWMDL D] HOV
deszaturációt katalizáló -deszaturáz enzimnek (Gurr, 1999a). Az egyes állatfajokbDQ D] -linolénsavnak dokozahexaénsavvá (docosahexaenoic acid '+$ YDOy iWDODNtWiVD NO|QE|] KDWpNRQ\ViJJDO PHJ\ YpJEH
(Bezard és mtsai, 1994). A rágcsálókban (pl. egerek) az átalakítás
KDWiVIRND QDJ\RQ My iP HJ\HV K~VHY N N|]WN SpOGiXO D PDFVNiN QHP
képesek az -OLQROpQVDYEyOW|UWpQ '+$-HO iOOtWiVUD3DZORVN\pVPWVDL %iU D] HPEHUEHQ P N|GLN D] iWDODNtWy HQ]LPUHQGV]HU KDWiVIRND PHJOHKHW VHQDODFVRQ\IHOQ WWHNEHQ NE-ra becsült (Emken és mtsai,
FVHFVHP NEHQ SHGLJ FVXSiQ -os (Salem és mtsai, 1996). Az eikozapentaénsavat (eicosapentaenoic acid, EPA) és a DHA-t szervezetükben nagy koncentrációban tartalmazó halak sem képesek e
KRVV]~ OiQF~ ]VtUVDYDNDW D] HPEHUQpO KDWpNRQ\DEEDQ HO iOOtWDQL .\OH
2001). A halak egy részének táplálékul szolgáló fotoszintetizáló mikroalgák zsírsavátalakító képessége azonban kiváló, lipidjeikben nagy arányban felhalmozva az EPA-t és a DHA-t (Kyle, 2001). E zsírsavak természetesen a táplálékláncból adódóan a ragadozó halak szervezetében is feldúsulhatnak. Az egyes szövetek n-6-os és n-3-as hosszú szénláncú
]VtUVDYDNDW V]LQWHWL]iOy NpSHVVpJH V]LQWpQ HOWpU $ V]HUYHN N|]O NLHPHOHQG D PiM QDJ\IRN~ DNWLYLWiVD H WHNLQWHWEHQ DPL YDOyV]tQ OHJDMyYDONLVHEE HO iOOtWiVLNDSDFLWiV~DJ\-14%), szív (8%) és vesék (12%) ellátásában is segít (Brenner, 1971). A szintézist befolyásoló takarmányozási-táplálkozási faktorok közül maguk a zsírsavak a legfontosabbak. A linolsavnak az -linolénsavhoz viszonyított
túl nagy mennyisége gátolhatja az EPA és a DHA kialakulását, az - linolénsav ugyanígy hatással lehet a linolsav származékainak
NpS] GpVpUH$WiSOiOpNNDOIHOYHWWQ-6-os és n-3-as zsírsavak aránya ezért
NULWLNXV MHOHQW VpJ $] HJ\PiVVDO NRPSHWHQV OLQROVDY pV -linolénsav serkenti a deszaturációs enzimek aktivitását. A hosszabb szénláncú
DUDFKLGRQVDY(3$pV'+$HU VHQJiWROMiNDOLQROVDY -deszaturációját, kevésbé a dihomo- -linolénsav -deszaturációját és nem befolyásolják
D] -linolénsav -deszaturációját (Bezard és mtsai, 1994). A - deszaturáz affinitása a szubsztrátokhoz az -linolénsav>linolsav>olajsav irányban csökken, bár esszenciális zsírsavhiányban az n-9-es átalakítási út dominánssá válhat.
A linolsavból és az -OLQROpQVDYEyONpS] G KRVV]DEEOiQF~]VtUVDYDN
nélkülözhetetlenek a membránok szerkezetének és funkciójának fenntartásában, másrészt prekurzorai a biológiailag rendkívül aktív eikozanoidoknak. A PUFA arányának csökkenése a membránon belül megváltoztatja a membránnak vízzel és egyéb kisebb molekulákkal szembeni átjárhatóságot (Sardesai, 1992). A telített zsírsavaknál nagyobb
KHO\HW LJpQ\O PR]JpNRQ\DEE W|EEV]|U|VHQ WHOtWHWOHQ YDJ\ HOiJD]y ]VtUVDYDN NHGYH] HN D PHPEUiQRN ~Q ÄIpOIRO\pNRQ\´ MHOOHJpQHN
fenntartásában.
A 20 szénatomot tartalmazó PUFA-ból szintetizálódó eikozanoidok (görög eicosa=20) már nagyon alacsony koncentrációban (10-9 g/g szövet)
IL]LROyJLDL KDWiVVDO UHQGHONH] YHJ\OHWHN DPHO\HN D V]HUYH]HWEHQ ]DMOy J\XOODGiVRV UHDNFLyN VHMWV]LQW NRRUGLQiWRUDL 1HHGOHPDQ pV PWVDL $] HLNR]DQRLGRN NpS] GpVH VRUiQ NLLQGXOy YHJ\ületként szolgálhat a dihomo- -linolénsav, az arachidonsav és az EPA. A
FLNORR[LJHQi] HQ]LP N|]UHP N|GpVpYHO SURV]WDJODQGLQRN WURPER[iQRN
prosztaciklinek jönnek létre. A lipoxigenáz reakcióút termékei a
OHXNRWULpQHN OLSR[LQRN pV NO|QE|] KLGUR[L-zsírsavak. A kiinduló vegyületként szolgáló n-6-os arachidonsav és az n-3-as EPA egymáshoz
YLV]RQ\tWRWW PHQQ\LVpJH G|QW MHOHQW VpJ PHUW D] XWyEEL ]VtUVDYEyO NpS] G -as sorozatú prosztaglandinok, tromboxánok, prosztaciklinek és 5-ös sorozatú leukotriének enyhébb lefolyású gyulladásos reakciót váltanak ki (Simopoulos, 1991). A szervezet homeosztázisának
V]HPSRQWMiEyOH]HNDYHJ\OHWHNEL]RQ\RVHVHWHNEHQHO Q\|VHEEHNPLQW D] DUDFKLGRQVDYEyO NpS] G HLNR]DQRLGRN $ GLKRPR- -linolénsavból kialakuló 1-es sorozatú prosztaglandinok (pl. 6-keto-PGF1, PGE1) és 3-as
OHXNRWULpQHN HU VHQ DQWL-LQIODPPDWRULNXV D]D] J\XOODGiVJiWOy MHOOHJ HN
(Gurr, 1999a).
2.2.3. Természetes források
Az n-6-os linolsav legfontosabb forrásai a növényi olajok, például a napraforgó-, kukorica-, szója-, repce- és pórsáfrányolaj (Kinsella, 1991).
$] iOODWL HUHGHW WHUPpNHN iOWDOiQRVViJEDQ MyYDO NLVHEE PHQQ\LVpJEHQ
tartalmazzák. Közülük kiemelkedik az anyatej, a máj és a tojás linolsavtartalma. A hosszú szénláncú arachidonsavban gazdag élelmiszernek számítanak a máj, a vese, a tojás és a húsok. A növényi kloroplasztiszmembránok zsírsavainak több mint a felét teszi ki a majdnem kizárólag galaktozil-GLDFLOJOLFHURORN DONRWyMDNpQW HO IRUGXOy - linolénsav (Gurr, 1999a). Egyes algákban (pl. Aphanizomenon flos- aquae D PDJDVDEEUHQG Q|YpQ\HNKH] KDVRQOyDQ D] -linolénsav a domináns többszörösen telítetlen zsírsav (Drapeau és mtsai, 2001). A növényi olajok egy része, mint például a lenolaj, szójaolaj, repceolaj és dióolaj szintén nagy koncentrációban tartalmazza (Kinsella, 1991). Az
iOODWL HUHGHW pOHOPLV]HUHNEHQ D OLQROVDYQiO iOWDOiEDQ NLVHEE PHQQ\LVpJEHQ IRUGXO HO $] HJ\HV KDORODMRN pV D EiUiQ\K~V MHOHQW JD]GDJDEE -linolénsav-forrást. Érdekes módon a lovak depózsírja 17%- ban tartalmazza ezt a zsírsavat (Gurr, 1999a).
Az EPA-t és a DHA-W E VpJHVHQ WDUWDOPD]]D D IRWRV]LQWHWL]iOy
mikroalgák többsége (Drapeau és mtsai, 2001). A hosszú szénláncú n-3- as zsírsavak ugyancsak gazdag forrásai az ún. „zsíros” halak, pl. a makréla, a hering és a lazac (Kyle, 2001) $ GUiJD KDOOLV]WHW PHOO ] DNYDNXOW~UiNEDQ V]iUD]I|OGL HUHGHW WDNDUPiQ\RQ QHYHOW KDODN
EPA+DHA-tartalma jóval elmaradhat a tengeri planktonokat vagy hallisztet fogyasztó halak zsírsavtartalmától (Kyle, 2001). A legtöbb hal húsában az EPA és a dokozapentaénsav (docosapentaenoic acid, DPA) mennyisége elmarad a DHA mennyisége mögött (Ackman és mtsai, 2001).
2.2.4. A humán fogyasztás és szükséglet alakulása
A történelem korai szakaszában az emberiség nagyobb gyümölcs és leveles zöldség fogyasztása miatt az n-6-os és n-3-as zsírsavakat közel azonos mennyiségben vette fel táplálékával (Eaton és Konner, 1985). Az elmúlt száz évben a nyugati típusú társadalmakban az n-6-os zsírsavak
IHOYpWHOH MHOHQW VHQ PHJHPHONHGHWW HOV VRUEDQ D] Q-6-os zsírsavakban gazdag növényi olajok használatának elterjedéséne miatt. Ma az ún.
„nyugati étrendben” az n-6-os és n-3-as zsírsavak aránya 20:1-30:1 Nagy- Britanniában, és 15:1-20:1 Kanadában (Scientific Review Committee, 1990). Az EPA és DHA becsült átlagos felvétele 100 mg/nap Ausztráliában és az USA-ban, ami az alacsony, 19 g/nap illetve 20 g/nap halfogyasztással magyarázható (Sinclair és mtsai, 1994; Gibney, 1997;
American Dietetic Association, 1995). Gibney (1997) becslése szerint az USA tíz államában az n-3-as zsírsavfogyasztás naponta 680 mg. Ollis és mtsainak (1999) vizsgálata szerint Ausztráliában az n-6-os zsírsavak felvétele 9,9 g/nap, az n-3-DV]VtUVDYDNpJQDSDPLE OPJQDSD]
-linolénsavon kívüli zsírsavak mennyisége. Egy Nagy-Britanniában végzett felmérés (Gregory és mtsai, 1990) szerint az n-6-os és n-3-as zsírsavak napi fogyasztása eléri az összes felvett energia 5 illetve 0,7%-át.
Európa egyéb országaiban nemzeti felmérés az n-6-os és n-3-as zsírsavak fogyasztásáról ezidáig nem készült.
A PUFA ajánlott napi bevitelét a napi energiafelvétel %-ában, vagy
DEV]RO~W PpUWpNHJ\VpJEHQ DGMiN PHJ D NO|QE|] DMiQOások. A tudományos szervezetek n-6-os és n-3-as zsírsavakra vonatkozó javaslatait az 1. táblázat IRJODOMD |VV]H $] HVHWHQNpQW IHOW Q NO|QEVpJHNHW QHPFVDN D NO|QE|] PHJKDWiUR]iVL PyGV]HUHN RNR]WiN KDQHPD]RSWLPiOLVWiSOiONR]iVIRJDOPiQDNHOWpU pUWHlmezése is. Az SCF (Scientific Committee for Food, 1993) és a FAO/WHO (1998) szerint az EPA és a DHA nem esszenciális zsírsav, a szervezet saját bioszintézise
NpSHV D] HO iOOtWiVXNUD $ W|EEL DMiQOiV YLV]RQW D ELRV]LQWp]LV J\HQJH
hatásfokát, a két zsírsav VSHFLiOLV ELROyJLDL IXQNFLyLW LV V]HP HO WW WDUWYD
javaslatokat ad a napi bevitelre. A Department of Health UK (1994) és az
6&) DMiQOiVDLEDQ D] HVV]HQFLiOLV ]VtUVDYKLiQ\ PHJHO ]pVpW WDUWMD V]HP HO WW PtJ D PiVLN NpW V]HUYH]HW ILJ\HOHPEH YHV]L KRJ\ a PUFA
PLQLPiOLV V]NVpJOHWHQ IHOOL EL]RQ\RV PHQQ\LVpJH NHGYH] KDWiVRNNDO
bír. A nemzetközi szervezeteken kívül számos kutatócsoport próbálta meghatározni az optimális PUFA-felvételt. A biológiai folyamatok jobb hatékonysága érdekében az energiabevitel 6-8%-a lenne ajánlatos a linolsavból, ami az ember esetében napi 7-10 g elfogyasztását jelenti.
Bjerve és mtsai (1989) 860-990 mg/nap LNA és 350-400 mg/nap EPA és DHA felvételét tartja optimálisnak. Barlow és mtsai (1990) naponta 3 g összes n-3-as zsírsav IRJ\DV]WiViW MDYDVROMiN DPLE O J OHQQH D]
EPA+DHA mennyisége. Az optimális n-6/n-3-as zsírsavarány 6:1 (British Nutrition Foundation, BNF, 1992), 5:1 (Nationale Raad voor Voeding) és 4:1-+HDOWKDQG:HOIDUH&DQDGDOHQQHDNO|QE|] DMiQOiVRN
szerint.
1. táblázat. Nemzetközi szervezetek javaslatai az n-6-os és n-3-as PUFA felvételére (az összes felvett energia %-ában)
n-6 PUFA n-3 PUFA LNA EPA + DHA
Dep.Health (1994)1 > 1,0 >2,0 >0,2 1,5 g/hét
BNF (1992)2 3-10 0,5-2,5 1,0 0,5
SCF (1993)3 2,0 0,5 - -
FAO/WHO (1998)4 4-10 0,4-2,0 - -
1Department of Health (UK); 2BNF: British Nutrition Foundation; 3Scientific Committee for Food; 4)RRG DQG $JULFXOWXUH 2UJDQL]DWLRQ:RUOG +HDOWK 2UJDQL]DWLRQ /1$ - linolénsav; EPA+DHA: eikozapentaénsav+dokozahexaénsav
2.2.5. Az n-6/n-3-as zsírsavarány és az emberi egészség
A XX. század második felében kialakult „lipid hipotézis” - amely a vér koles]WHULQV]LQWMpW HPHO WHOtWHWW ]VtUVDYDNEDQ OiWWD D V]tYNRV]RU~pU-
PHJEHWHJHGpVHN HJ\LN I RNiW– D 38)$ NHGYH] pOHWWDQL KDWiVDL IHOp
irányította a figyelmet (Gurr, 1999a). Kezdetben az n-6-os család tagjainak fontossága volt hangsúlyosabb, mivel a linolsavIHOWpWHOH]KHW HQ
szabályozó szerepet játszik a vérplazma LDL-szintjének szabályozásában
+D\HV (J\ D] HJ\pQUH MHOOHP] V]NVpJOHWL NV]|EV]LQW DODWWL
linolsavfelvétel esetén a linolsavfogyasztás növelése csökkentheti az LDL-koleszterin koncentrációját a vérplazmában. A XX. század végére
HOV VRUEDQ D OLQROVDY H]HQ HPOtWHWW NHGYH] KDWiVD PLDWW D] Q-6-os zsírsavakban gazdag olajok fogyasztása megemelkedett a nyugati társadalmakban, túlságosan tág n-6/n-3-as arányt kialakítva. Okuyama és mtsai (1996) szerint ez összefüggésben van bizonyos rákfajták, allergiás megbetegedések, trombózis és idegrendszeri zavarok gyakoriságának fokozódásával. E káros folyamatok az n-6-RV]VtUVDYDNEyONpS] G HOWpU
hatású eikozanoidokra és az n-3-DV -linolénsav-DHA átalakítási útvonal
KiWWpUEHV]RUXOiViUDYH]HWKHW NYLVV]D
Az n-3-DV ]VtUVDYDN LV HO V]|U D V]tYNRV]RU~pU-megbetegedésekkel
NDSFVRODWEDQ NHUOWHN HO WpUEH 6LQFODLU eV]DN-Amerikában
YpJ]HWW IHOPpUpVHL D]W VXJDOOWiN KRJ\ D KDOIRJ\DV]WiV PHJHO ] V]HUHSet játszhat a szívkoszorúér-betegségek kialakulásában. Számos epidemiológiai tanulmány támasztotta alá ezután ezt a nézetet. Bang és
'\HUEHUJ D FDUGLRYDVFXODULV EHWHJVpJHNE O HUHG HOKDOiOR]iVRN HOWpU J\DNRULViJiW PXWDWWD NL JU|QODQGL pV GiQ HV]NLPóknál, a különbségeket a halfogyasztásra visszavezetve. A Kromhout és mtsai
(1985) által végzett ún. Zutphen-felmérés szerint az átlagban napi 42 g halat fogyasztó szívinfarktuson átesett páciensek négy év után már
MHOHQW VHQMREEW~OpOpVLHVpO\HNNHOUHQGHlkeztek, mint halat nem fogyasztó
WiUVDLN $ KDOIRJ\DV]WiV H NHGYH] KDWiVD P|J|WW G|QW HQ D KRVV]~
szénláncú n-3-as EPA-nak és DHA-nak a hatása áll. A csukamájolajban található EPA és DHA legalább napi 4 g-nyi mennyisége fenntartotta a vér triglicerid konFHQWUiFLyMiWDQRUPiOV]LQWWHOUHQGHONH] HJ\pQHNEHQD K\SHUWULJO\FHULGDHPLiEDQ V]HQYHG NpW SHGLJ V]LJQLILNiQVDQ FV|NNHQWHWWH
(Sanders és mtsai, 1981). A plazma trigliceridszint-csökkenésének hátterében a májból kiáramló VLDL mennyiségének csökkenése áll, a VLDL-szintézis gátlása által (Hayes, 2001). A koleszterinszintet szabályozó linolsavval ellentétben az n-3-as zsírsavak nem fejtenek ki
HJ\pUWHOP KDWiVW D YpU NROHV]WHULQV]LQWMpUH *XUU D (OHJHQG
mennyiségben való fogyasztásuk azonban csökkentheti a trombózis veszélyét, mert a vérlemezkék membránjába épülve megnövelik a vérzési
LG WJiWROMiNDYpUOHPH]NpNDJJUHJiFLyMiWYRQ6KDFN\
Az n-3-as zsírsavak szerepe számos egyéb területen is ismertté vált. A DHA nélkülözhetetlen az egészséges idegi (Xu és mtsai, 1996) és látásfunkciók (Holub, 2001) kialakításában és fenntartásában, különösen a magzati élet utolsó harmadában és a korai neonatális szakaszban. Az újszülött gyenge DHA-szintetizáló képességét az anyatej nagy DHA- tartalma ellensúlyozza (Holub, 2001). Az EPA és a DHA a vörös- és fehérvérsejtekben az arachidonsav metabolitjait visszaszorítva kevésbé g\XOODGiVNHOW KDWiV~ HLNR]DQRLGRN V]LQWp]LVpW VHJtWL HO *XUU D
Ezt kihasználva a rheumatoid arthritis aktív (inflammatiós) fázisának
V~O\RVViJD LV FV|NNHQWKHW YROW QpKiQ\ HVHWEHQ %ULWLVK 1XWULWLRQ
Foundation, 1992).
Az n-3-as PUFA-QDN MHOHQW V szerepet tulajdonítanak az
LQ]XOLQUH]LV]WHQFLD pV D] HOKt]iV PHJHO ]pVpEHQ LV ,Q]XOLQUH]LV]WHQFLD
esetén a vérben fiziológiás koncentrációban található inzulin nem képes
PHJIHOHO HQ NLIHMWHQL KDWiVDLW 6DOWLHO $] Q-3-as zsírsavak a hipotézis szerint közvetlenül javíthatják a vázizomsejtek inzulinérzékenységét, illetve az elhízás gátlása által közvetett módon akadályozhatják az inzulinrezisztencia kialakulását (Storlien és mtsai, 1996). A nagy zsírtartalmú, telített, egyszeresen telítetlen vagy n-6-os PUFA-ban gazdag takarmány inzulinrezisztenciát alakított ki a kísérleti patkányokban. Az alacsony n-6/n-3-as zsírsavarányt biztosító, halolajat tartalmazó takarmány viszont fenntartotta a szabályos inzulinfunkciót (Storlien és mtsai, 1991). Baur és mtsai (1998) által közétett adatok
szerint az inzulinrezisztencia mértéke negatív korrelációban áll a vázizmok foszfolipidjeinek DHA-tartalmával emberekben is. A zsírsavak
HPHONHG WHOtWHWOHQVpJL IRNiYDO Q|YHNY PHPEUiQIOXLGLWiV KDWiViUD D
glükóztranszporter-4 W|EE LG W W|OW D VHMWPHPEUiQKR] NDSFVROyGYD HO VHJtWYH D JONy] IHOYpWHOpW D] L]RP- és a zsírsejtekben (Hansen és
PWVDL $ VHMWHQ EHOO tJ\ PHJQ|YHNY JONy]-6-foszfát-szint serkenti az izomsejtek glikogénszintézisét, ami az izomsejtek glükózfelvéWHOpQHN I KDMWyHUHMH $ 38)$ N|]O HOV VRUEDQ D] Q-3-as család hosszú szénláncú tagjai eltolhatják a zsírsavak anyagcseréjét a zsírsavoxidáció irányába. A említett zsírsavak egyszerre serkentik a
]VtUVDYR[LGiFLyW pV D K WHUPHOpVW PLN|]EHQ JiWROMiN D ]VtUsavak szintézisét (Clarke, 2000). A sejtek PPAR (peroxisome proliferator activated receptor HOHPHLKH] N|W GQHN DPHO\HN V]iPRV JpQ PHJIHOHO
szakaszával reakcióba lépve befolyásolják a DNS-transzkripció mértékét,
D NHOHWNH] IHKpUMH PHQQ\LVpJpW 6HVVOHU ps Ntambi, 1998). A zsírsavoxidáció növelését a zsírsavoxidációt szabályozó enzimfehérjék (pl. karnitin-palmitoiltranszferáz) fokozottabb szintézisével érik el (Mascaro és mtsai, 1998), ugyanakkor serkentik a termogén peroxiszómális zsírsavoxidációt és a mitokondriális oxidatív foszforiláció szétkapcsolását (Reddy és Mannaerts, 1994).
Az n-3-as zsírsavak túlzottan nagy mennyiségének felvétele is számos
QHJDWtY N|YHWNH]PpQQ\HO MiUKDW $] RSWLPiOLVW PHJKDODGy PpUWpN IRJ\DV]WiV IRNR]]D D OLSLGSHUR[LGiFLy NLYpGpVppUW IHOHO V DQWLR[LGiQVRN
iránti igényt, túlzottan lecsökkentheti az arachidoQVDYEyO NpS] G
prosztaglandinok mennyiségét és immunszupressziót okozhat (Simopoulos, 1991).
2.3.AZ ÉTKEZÉSI TOJÁS ZSÍRSAVÖSSZETÉTELÉNEK MÓDOSÍTÁSA
2.3.1. A zsírok szerepe a tojótyúk takarmányozásában
A takarmány zsírtartalma optimális esetben fedezi az állatok esszenciális zsírsavszükségletét, illetve koncentrált energiaforrást jelent. Baromfiban
HOV NpQW D OLQROVDY HVV]HQFLDOLWiViW EL]RQ\tWRWWiN 5HLVHU $ NpV EEL NtVpUOHWHN D EURMOHUHN pV WRMyW\~NRN V]NVpJOHWL pUWpNHLW LV
meghatározták. A Magyar Takarmánykódex (1990a) középnehéz tojók
UpV]pUH PLQLPiOLVDQ N|QQ\ WHVW WRMyN WDNDUPiQ\iEDQ D K PpUVpNOHWW OpVWDNDUPiQ\IHOYpWHOW OIJJ HQ-OLQROVDYDWtUHO
Az NRC (1994) ajánlása barna tojást tojó tyúkok részére min. 1,1%
linolsavat javasol. A Lohmann cég Lohmann Brown-Classic tojóhibridjének takarmányozási ajánlásában tojónként és naponta 2g
OLQROVDYDW tU HO /RKPDQQ 7LHU]XFKW *PE+ $] -linolénsavat az
HPO V|NK|] KDVRQOyDQ YDOyV]tQ OHJ D EDURPIL VHP QpONO|]KHWL GH
esszencialitása nem bizonyított, konkrét szükségleti értéket ismereteink szerint eddig csak Németországban adtak meg (Bedarfsnormen der Gesellschaft für Ernährungsphysiologie, 1999). A tojótyúkokra vonatkozó ajánlás a takarmány 1 kg szárazanyagára számítva 10 g linolsavat és 0,6 g
-linolénsavat, azaz a két zsírsav 16:1-HVDUiQ\iWWDUWMDN|YHWHQG QHN
Hazánkban a tojótyúkok takarmánya olaj- vagy zsírkiegészítést
QHPWDUWDOPD]PHUWD]iOODWRNOLQROVDYLUiQWLPLQLPiOLVLJpQ\HIHGH]KHW D WiSRN PHJIHOHO NXNRULFDWDUtalmával, továbbá a tápok szükséges
HQHUJLDWDUWDOPDHOpUKHW ]VtUNLHJpV]tWpVQpONOLV$MHOHQOHJpUYpQ\HV(8
szabályok szerint állati zsír gazdasági állatok takarmányozásában nem használható (a tej, illetve egyes esetekben a halliszt zsírtartalma kivételével). A növényi olajok felhasználása, különösen az
~MUDKDV]QRVtWRWWRODMRNpIRNR]RWWHO YLJ\i]DWRVViJRWLJpQ\HO
2.3.2 A tojótyúk lipidmetabolizmusának jellegzetességei
$] LQWHQ]tYHQ WHUPHO WRMyW\~NRN D WRMiVNpS] GpV KDWDOPDV
lipidszükségletét biztosíWy D NDNDVRNWyO pV D EURMOHUHNW O HOWpU
lipidanyagcserével rendelkeznek (Freeman, 1976). A madarakban az epe és a hasnyálmirigy vezetékei a duodenum disztális részébe torkollanak,
HOOHQWpWEHQD]HPO V|NNHODKROH]HNDYH]HWpNHNDGXRGHQXPSUR[LPiOLV
szakaszába nyílnak. A zsíremésztés szempontjából e hátrányos anatómiai elhelyezkedést a béltartalomnak a duodenum és a zúzógyomor közti visszaáramlása ellensúlyozza, biztosítva a chymusban az epesavak és a hasnyálmirigy-OLSi]PHJIHOHO HONHYHUHGpVpW6NODQpV mtsai, 1978).
$ OLSLGHN HPXOJHiOiVD pV HQ]LPHV HPpV]WpVH D] HPO V|NpKH] KDVRQOy
módon megy végbe (Freeman, 1976). A madarakban az intesztinális
Q\LURNUHQGV]HU KLiQ\D N|YHWNH]WpEHQ D PXFRVD VHMWHNEHQ OpWUHM|Y
lipoproteinek a portális vénákba jutnak. Az emO V|N NLORPRNURQMDLYDO
homológ lipoproteineket a keringésbe kerülés módja alapján portomikronoknak nevezték el (Bensadoun és Rothfield, 1972). A portomikronok a májat elérve részleges átalakítást szenvedhetnek, majd az exrahepatikus szövetekben nagyon gyorsan lebomlanak (Hermier, 1997).
$ ]VtURN V]LQWp]LVH D] HPO V|NW O HOWpU HQ QDJ\REE UpV]EHQ D PiMEDQ MiWV]yGLN OH $ WRMiVUDNiVL LG V]DN NH]GHWpQ D] |V]WURJpQHN UHQGNtYOL
mértékben fokozzák a máj lipogenezisét. A máj szárazanyagának 10- 15%-iWNLWHY ]VtUWDrtalom a tojó madarakban akár 40%-ra is emelkedhet
6FKXPDQQ pV PWVDL $ OHJW|EE HPO V ]VtUVDYV]LQWp]LVH VRUiQ D
telített zsírsavak nagyobb arányban keletkeznek, mint a telítetlenek. A
Ki]LW\~NEDQ D] RODMVDY V]LQWp]LVH D GRPLQiQV PHJHO ]YH D WHOtWHtt palmitin- és sztearinsavét (Annison, 1983). A vérplazma lipoproteinjei a
9/'/ /'/ pV D QDJ\ V U VpJ OLSRSURWHLQ high density lipoprotein, HDL) osztályokba sorolhatók (Yu és mtsai, 1976). A plazma LDL-szintje a madárfajokban jellegzetesen alacsonyabb D] HPO V|NNHO
összehasonlítva (Annison, 1983). A tojástermelés megindulása idején a
Q LYDU~iOODWRN9/'/-szintje a vérplazmában a 0,6 g/l-U OJO-UHQ D]
ösztrogének hatására, miközben a HDL koncentrációja körülbelül
IHOH] GLN <X pV PWVDL $] HOWpU OLSRSURWHLQ DUiQ\RN HOOHQpUH D WHUPHO WRMyW\~NRN OLSRSURWHLQMHLEHQ D I|EE OLSLGIUDNFLyN KDVRQOy -ban vannak jelen, mint a fiatal jércék, kakasok vagy brojlerek esetében (Yu és mtsai, 1976).
A tojás lipidjeinek prekurzorai a májban szintetizálódó lipoforin, vitellogenin és VLDL (Davis, 1997). A tojótyúk VLDL-molekulájában a központi mag nagy részét (70-80%) trigliceridek alkotják a foszfolipidekkel (10-28%) és koleszterin-észterekkel (1-4%) együtt (Griffin és mtsai, 1982). A lipidmagot apoB100 és apoVLDL-,, MHO|OpV
apolipoproteinek veszik körbe. Egy apoB molekulára kb. 40-50 apoVLDL-II-molekula jut (Walzem és mtsai, 1999). A madarakban nem
IRUGXO HO D] HPO V|NEHQ PHJWDOiOKDWy DSR% DSROLSRSURWHLQ 'DYLV
1997). Az ösztrogének a zsírok mellett az apoB100 termelését is fokozzák és specifikusan e hormonok hatására indul meg az apoVLDL-II szintézise (Williams, 1979). A hormon hatására azonban a VLDL-molekula átlagos
iWPpU MHLVOHFV|NNHQDIHOpUH-45 nm), felületén egy apoB molekulára csupán 23 apoVLDL-II-molekula jut (Walzem és mtsai, 1999). Ez a kis
iWPpU M 9/'/\ („yolk-targeted” NpSHV D Q|YHNY SHWHVHMWHW KDWpNRQ\DQ HOOiWQL OLSLGHNNHO .LVHEE PpUHWpE O DGyGyDQ iW WXG MXWQL D IROOLNXOXV JUDQXORVD UpWHJpQHN DODSKiUW\iMiQ DPHO\ V] U NpQW P N|GLN
$ 9/'/\ PLQGHPHOHWW UH]LV]WHQV D] iWDODNtWiViW YpJ] OLSRSURWHLQ
lipázzal (LPL) szemben, mert nem tartalmazza az enzimet aktiváló apoC- II-t, valamint az apoVLDL-II az LPL specifikus inhibitora. A szöveti lebomlást elkerülve az oocyta intakt formában, receptor irányította endocitózissal veszi fel. Walzem és mtsainak (1999) feltételezése szerint a
tojótyúk veséjének proximális tubulusai „normál” VLDL-t termelnek, ami apoVLDL-II helyett apoC-t tartalmaz, s így a szomatikus LPL révén hozzájárulhat bizonyos szövetek (pl. szívizom) szabad zsírsav ellátásához.
Az említett ösztrogéneken túl a pankreatikus hormonok játszanak nagy szerepet a zsíranyagcsere szabályozásában. Az inzulin stimulálja a lipogenezis I HQ]LPMHLQHN D PDOiW-dehidrogenáznak és a zsírsav- szintetáznak az aktivitását (Hillgartner és mtsai, 1995). Tarlow és mtsai (1977) csirke hepatocytákkal folytatott kísérletében az inzulin serkentette a VLDL-szintézist. Mooney és Lane (1981) viszont csökkent apoB-
V]LQWp]LVU O pV 9/'/-szekrécióról, a májsejtekben felhalmozódó
WULJOLFHULGHNU O V]iPROW EH QDJ\ DGDJ~ LQ]XOLQNH]HOpV KDWiViUD $
glukagon az intracelluláris cAMP-koncentráció megemelése révén gátolja a zsírsavak és a lipoproteinek szintézisét (Leclerq és mtsai, 1988). A zsírszövetben lipolitikus hatású, emeli a plazma szabad zsírsav- koncentrációját (McMurtry és mtsai, 1996).
2.3.3. Az n-3-as zsírsavak hatása a tyúk lipidmetabolizmusára
$] HPO ViOODWRNEDQ pV D] HPEHUEHQ D] Q-3-as szírsavak számos kísérlet szerint csökkentik a máj saját zsírsavszintézisét, a koleszterin-észter- és
WULJOLFHULGNpS] GpVW YDODPLQW PHJQ|YHOLN D ]VtUVDYR[LGiFLy PpUWpNpW
amely hozzájárul a vér trigliceridszintjének csökkenéséhez (Harris, 1989).
Az EPA-ban és DHA-ban gazdag halolaj gátolta a patkány májsejtjeinek VLDL-szintézisét és -szekrécióját in vitro körülmények között (Lang és Davis, 1990). Az n-3-as zsírsavak hatásait a zsírsavszintézis és zsírsavoxidáció aktivitására madár májszövetében eddig még nem vizsgáltáN+DUJLVpV9DQ(OVZ\N$NROHV]WHULQV]LQWp]LVHJ\LNI
enzimének, a HMG-CoA-reduktáznak aktivitása drasztikusan csökkent a csirke májában a 10% menhaden halolajjal kiegészített táp fogyasztása
XWiQ &DVWLOOR pV PWVDL 7|EE V]HU] LV EHV]iPRlt a trigliceridek
NRQFHQWUiFLyMiQDN FV|NNHQpVpU O D YpUSOD]PiEDQ pV D 9/'/-ben a menhaden halolajjal kiegészített takarmány hatására (Phetteplace és Watkins, 1990; Castillo és mtsai, 1999). Az n-3-as zsírsavakban gazdag takarmánykiegészítések után a tojás és a tojássárgája tömegének
FV|NNHQpVH PLQGHQ EL]RQQ\DO HUUH YH]HWKHW YLVV]D :KLWHKHDG pV
mtsainak (1993) elmélete szerint ez összefügésben állhat a fokozott n-3-as zsírsavbevitel miatt kialakuló alacsonyabb ösztrogénkoncentrációval a plazmában. A tojót\~NRNQDN D WRMiVNpS] GpV OLSLGLJpQ\pW NLV]ROJiOy
intenzív lipidszintézise és -WUDQV]SRUWMD HOV VRUEDQ |V]WURJpQHN KDWiViUD
jön létre. A lenmag fitoösztrogénjei ugyancsak befolyásolhatják a tojás
PLQ VpJpW pV D WRMyN WHOMHVtWPpQ\pW PHUW D V]HUYH]HWEHQ DQWLösztrogén
MHOOHJ KDWiVRNDW IHMWKHWQHN NL 7KRPSVRQ pV PWVDL .HQQHG\ pV
mtsai (1994) lenmag etetésével 30%-os csökkenést értek el jércék vérplazmájának ösztradiolszintjében. Még megválaszolásra vár, vajon ilyen esetben az n-3-as zsírsavak közvetlenül befolyásolják a hormonszintet, vagy a fitoösztrogének is hozzájárulnak ehhez.
A táp 3% menhaden-halolaj-tartalma tojótyúkokban is szignifikánsan csökkentette a trigliceridek és a koleszterin koncentrációját hat hónapig
WDUWyNtVpUOHWLHWHWpVWN|YHW HQ(Van Elswyk és mtsai, 1994). Schumann és mtsai (2000) tojótyúkokkal 28 napig folytatott kísérletükben 100 g/kg lenmag, 40 g/kg lenolaj és 100 g/kg „Dry n-3” készítmény (25% halolaj, ami 18% EPA-t és 15% DHA-t tartalmazott) alkalmaztak. A vérplazma trigliceridszintje mindhárom kezelés esetében szignifikánsan (P<0,05; az n-3 készítménynél P<0,01) csökkent a négy hét elteltével, de ez ugyanígy
MHOOHP] YROW D] iOODWL-Q|YpQ\L YHJ\HV ]VtUNLHJpV]tWpV WiSRW IRJ\DV]Wy
kontroll csoportra is (P<0,05).
Többen vizsgálták az n-3-as zsírsavak lehetséges szerepét a tojótyúk
PiMiEDQ OLSLGIHOKDOPR]yGiVVDO MHOOHPH]KHW ]VtUPiM V]LQGUyPD ÄFatty Liver Syndrome”, FLS) vagy vérzéses zsírmáj szindróma („Fatty Liver Hemorrhagic Syndrome´ )/+6 PHJHO ]pVpEHQ NLDODNXOiViEDQ (] a
NyUNpS V]yUYiQ\RVDQ IRUGXO HO WRMyW\~NRNEDQ 6FKXPDQQ pV PWVDL V YDOyV]tQ OHJ KDVRQOy IRO\DPDWRN ]DMODQDN OH D PDJDV
szénhidráttartalmú takarmányok kényszeretetésének kitett libamájban is
+HUPLHU $ WQHWHJ\WWHV FVDN D WHUPHO Q LYDUEDQ IRUGXO HO
(Van Elswyk és mtsai, 1994), de csirkékben is kiváltható 17β-ösztradiol segítségével, ami a hormonnak a májlipidózis létrejöttében játszott
V]HUHSpW YDOyV]tQ VtWL 7DNDKDVKL pV PWVDL $ WHUOHWHQ IRO\y
kutatómunka ellenére az FLHS kórokWDQD pV NyUIHMO GpVH PpJ QHP
tisztázott pontosan, s az n-3-as zsírsavakkal kapcsolatos eredmények is ellentmondónak látszanak. A tengeri halolajoknak a máj zsírtartalmát és
W|PHJpWFV|NNHQW KDWiViUyOV]iPROWDNEH0DXULFHpVPWVDL(]]HO
szemben két NXWDWyFVRSRUW LV MHOHQW V OLSLGIHOKDOPR]yGiVW WDSDV]WDOW D
tojótyúkok májában a 3% menhaden-halolaj kiegészítés következtében (Hargis és mtsai, 1991; Van Elswyk és mtsai, 1994). Schumann és mtsainak (2000) már említett kísérletében a kezelések mindegyike hasonló mértékben csökkentette a máj zsírtartalmát a kontrollhoz képest, ám egyik kezelés sem volt képes meggátolni a májban vérzések
KDHPRUKDJLD NLDODNXOiViW V W D] Q-3-as készítmény a két kísérlet
HJ\LNpEHQIRNR]WDDYpU]pVHNV~O\RVViJiW$V]HU] NIHltételezése szerint az n-3-DV ]VtUVDYDN QDJ\REE PpUWpN IHOYpWHOH N|YHWNH]WpEHQ D WKURPERF\WD DJJUHJiFLyW HO VHJtW DUDFKLGRQVDY NRQFHQWUiFLyMD OHFV|NNHQKHW Q|YHOYH D YpU]pNHQ\VpJ YDOyV]tQ VpJpW ÈOWDOiQRVDQ HOIRJDGRWW KRJ\ D WULJOLFHULGHN PiMEDQ W|UWpQ felhalmozódásának
KiWWHUpEHQDOLSLGHNV]LQWp]LVpQHNpVDPiMEyOW|UWpQ V]HNUpFLyMiQDNQHP HJ\HQO PpUWpN YiOWR]iVD iOO +HUPLHU $ NpQ\V]HUHWHWpVVHO
takarmányozott ludakban szintén kialakuló zsírmáj esetén a triglicerideknek VLDL-EH W|UWpQ EHpSülése szenved zavart (Hermier, 1997).
2.3.4. A tojás lipidjei és zsírsavösszetétele
$]iWODJRVDQ JW|PHJ W\~NWRMiVN|UOEHOOJOLSLGHWWDUWDOPD]DPL
majdnem teljes mennyiségében a tojássárgájában található (Romanoff és Romanoff, 1949). A sárgája DODSYHW HQ HJ\ RODM-víz emulziónak
WHNLQWKHW DKRO D OLSLGFVHSSHNHW HJ\ KLGUDWiOW IHKpUMHIi]LV YHV]L N|UO
(Noble, 1987). A lipidek a fehérjékkel kb. 2:1 lipid:fehérje arányú lipoprotein-komplexeket képeznek. A lipoproteineket fizikai jellegzetességeik alapján „nagy”, és a lipidek 90%-át tartalmazó
ÄDODFVRQ\´ V U VpJ IUDNFLyNUD RV]WMXN $ WRMiVViUJiMiEyO H[WUDKiOKDWy
lipidek a sárgája teljes tömegének kb. 33%-át, szárazanyagának 60-65%-
iW WHV]LN NL $ OLSLGHN KiURP I FVRSRUWMiW D WULJOLFHULGHN -65%), a foszfolipidek (25-30%) és a szabad, nem észterezett koleszterin (4-6%)
DONRWMiN $ I EE HO IRUGXOy IRV]IROLSLG PROHNXOiN D IRV]IDWLGLO-kolin (69%) és a foszfatidil-etanolamin (24%).
A tojássárgája zsírsavai között az olajsav a domináló, a zsírsavak kb.
30-40%-át jelenti. A telített zsírsavak közül legnagyobb a palmitinsav, majd ezt követi a sztearinsav mennyisége. Különösen a foszfolipid frakció gazdag 20 és 22 szénatomszámú PUFA-ban (Noble, 1987). A foszfatidil-kolinban a linolsav, a foszfatidil-etanolaminban az
DUDFKLGRQVDY pV D '+$ PHQQ\LVpJH V]iPRWWHY 6WULFNODQG $
trigliceridekben a zsírsavak sajátos elhelyezkedést mutatnak (Christie és Moore, 1972). A glicerin 1-HV KHO\]HW V]pQDWRPMiKR] N|W G ]VtUVDYDN
70 %-a palmitinsav, a 2-es pozíciót G|QW HQ D] RODMVDY pV D OLQROVDY
foglalja el, míg a 3. helyen megint az olajsav az uralkodó. A foszfolipidekben a telítetlen zsírsavak a 2-es pozícióban találhatók leginkább. Összességében a tojássárgája zsírsavösszetétele túlnyomórészt telítetlennek nevezKHW NE -es telített/telítetlen zsírsavaránnyal, ami
kielégíti az egészséges ételekkel szemben támasztott követelményeket (National Advisory Committee on Nutrition Education, 1983). Az összes n-6-os és n-3-as zsírsav aránya 8 és 12 között mozog egy átlagos tojásban, ami alig múlja felül a többek által kívánatosnak megjelölt 4:1-10:1 arányt,
V H WHNLQWHWEHQ PpJ PLQGLJ PHJHO ]L D WHMHW pV D K~VRN W|EEVpJpW $
tyúktojáshoz hasonlóan más baromfifajok tojásaiban is az olajsav és a palmitinsav mennyisége a meghatározó (Maldijan és mtsai, 1996; Noble, 1987). A fácán- és a strucctojás triglicerid frakciójában az -linolénsav szintje a nagyon magasnak számító 25%-ot is eléri. A DHA mennyisége a tyúk és a lúd tojásának foszfolipidjeiben a legnagyobb.
Az intenzív tojástermelés takarmányozási és környezeti viszonyai között a tojássárgája lipidösszetétele nagyfokú stabilitást mutat. A tojótyúk anyagcseréje a potenciális utód számára a legoptimálisabb
N|UQ\H]HW EL]WRVtWiViW HO WpUEH KHO\H]YH NpSHV D OHJW|EE NOV IDNWor változását kivédeni. A takarmány energiatartalmának a szükségletet el
QHPpU YDJ\PHJKDODGyV]LQWMHDWRMiVViUJiMDPpUHWpWEHIRO\iVROKDWMDGH
a lipidösszetételre nem, vagy csak nagyon kis mértékben hat (Noble, 1987). A szénhidrátok és a fehérjék egymáshoz viszonyított aránya szintén nem hat szignifikánsan a lipidek arányára, a sárgája összlipidtartalmát viszont megváltoztathatja (Andersson és mtsai, 1978).
$ WDNDUPiQ\ Q\HUV]VtUWDUWDOPiQDN YiOWR]WDWiVD LV FVDN NLV PpUWpN
összlipidtartalom-változást eredményezett (Ostrander és mtsai, 1960). A
PiMpVDYpUSOD]PDOLSLGSDUDPpWHUHLQHNNO|QEVpJHLQHPV]NVpJV]HU HQ MHOHQQHN PHJ D WRMiVViUJiMD HOWpU OLSLG|VV]HWpWHOpEHQ $ NO|QE|]
tyúkhibridek és -IDMWiN LOOHWYH NO|QE|] NRU~ PDGDUDN N|]|WW PiU
nagyobb eltérések lehetnek a tojássárgája lipidösszetételében (Washburn, 1979).
2.3.5. Az n-3-as zsírsavak hatása a tojástermelési paraméterekre
A takarmány 3%-os menhaden-KDORODMMDOW|UWpQ NLHJpV]tWpVpQHNKDWiViUD
egyes kutatócsoportok nem tapasztaltak változást a tojótyúkok tojástermelési mutatóiban (Hargis és mtsai, 1991; Van Elswyk és mtsai, 1995). Marshall és mtsai (1994) azonban e halolaj típus 1,5%-os alkalmazása során a tojássárgája tömegének, Gonzalez-Esquerra és Leeson (2000) a tojás tömegének csökkenéspU O V]iPROWDN EH $
heringliszt felhasználása során a tojástömeg csökkenését írták le Nash és mtsai (1995), amit korábban nem tapasztaltak a menhaden-halliszt etetésének hatására (Nash és mtsai, 1996). Az utóbbi kísérletben a 4 és
8% menhaden-halliszt szignifikánsan (P<0,05) csökkentette, a 12%-os szint pedig szignifikánsan (P<0,05) növelte a tojók testtömegét a kontroll táphoz képest. A 4%-os bekeverési szintig javult a takarmányértékesítés, nem változott viszont a tojástermelési % és a Haugh-egység. A 4%-os halolaj kiegészítés nem vezetett a tyúkok gyengébb tojástermeléséhez vagy kisebb tojástömeghez Baucells és mtsai (2000) vizsgálatában sem.
Ezzel ellentétben Whitehead és mtsainak (1993) kísérletében a 6%-os halolaj kiegészítést fogyasztó tyúkok tojástermelése közel 10%-kal maradt el a kontroll csoportétól, és mintegy 7%-kal a 6%-os kukoricaolaj
NLHJpV]tWpV WDNDUPiQ\WIRJ\DV]WyWRMyNpWyO$ KDORODMRVFVRSRUWUpV]pU O
szintén elmaradás mutatkozott a tojástömeg-növekedés és a sárgájatömeg tekintetében az |WKHWHVNtVpUOHWLLG V]DNYpJpQ
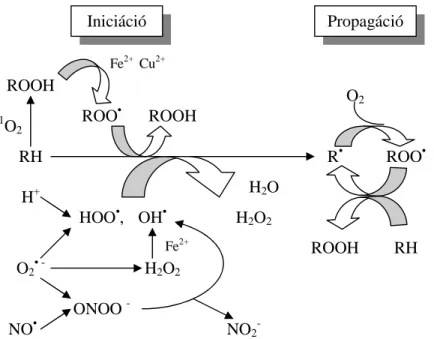
$] DOJD HUHGHW '+$ *ROGTM
QHY WHUPpN 2PHJD7HFK -ban 4 hétig 24 hetes tojókkal etetve a tojástermelés csökkenését eredményezte (Nollet, 2001). Az 5%-ban alkalmazott MaxEPA készítmény nem befolyásolta a termelési paramétereket, de 10%-os szintje már csökkentette a tojásdarabszámot (Oh és mtsai, 1994).
Caston és mtsai (1994) testtömeg- és tojástömeg-csökkenést mutattak
NL KHWHV iOODWRNEDQ pV GDUiOW OHQPDJ DONDOPD]iViW N|YHW HQ YDOyV]tQ OHJDWDNDUPiQ\HQHUgiatartalmának csökkenése miatt. Sheideler és Froning (1996) 43 hetes tyúkok testtömegének, takarmányfelvételének és tojástömegének csökkenését tapasztalta 5 és 15% egész és darált lenmag 8 hetes etetése után, változatlan tojástermelés mellett. Ezzel ellentétben a tojástermelés négy hét alatt 84%-ról 65%-ra esett vissza 15%-os egész és darált lenmag hatására Aymond és Van Elswyk (1995)
NtVpUOHWpEHQ 6KHLGHOHUpV )URQLQJ HO EEL YL]VJiODWD VRUiQ D - os lenmagarány vékonyította a tojáshéjat, és kisebb tojássárgájatömeget eredményezett. A lenmagdarák sokszor tapasztalt negatív hatása ellenére a lenolaj 9%-os szintig alkalmazva nem befolyásolta a tojástermelést, tojástömeget, sárgájatömeget és a napi takarmányfelvételt (Huygebaert, 1995). Balevi és CoskuQ KDVRQOy HUHGPpQ\HNU O V]iPROW EH
Grobas és mtsai (2001) az 5 és 10% lenolaj kiegészítés vizsgálata során szignifikánsan (P<0,05) kisebb tojástömeget mértek az azonos arányban alkalmazott szójaolajhoz képest, miközben a takarmányfelvétel és a tojástermelés nem különbözött a csoportok között. Baucells és mtsai (2000) a termelési paraméterekben nem találtak szignifikáns különbséget a 4% halolaj és 4% lenolaj kiegészítések összehasonlítása során.
2.3.6. A takarmányozás hatása a tojás zsírsavösszetételére
A tojás zsírsavösszetételének változtatása takarmányozással Cruickshank (1934) kísérletéig nyúlik vissza. A takarmány 28%-os lenolaj kiegészítésével a tojás -linolénsav-tartalmát 58 mg/g tojássárgája értékre sikerült növelnie, azaz a takarmány PUFA-összetételének változtatását a tojássárgája PUFA-|VV]HWpWHOH PHJOHKHW VHQ MyO N|YHWWH $ OLQRO- és - linolénsavban gazdag olajkiegészítések megnövelték a két zsírsav szintjét a tojásban, ami együtt járt az olajsavtartalom arányos csökkenésével (Wheeler és mtsai, 1959). A tojás zsírsavösszetétele a takarmány 20 és 22 szénatomos n-6-os és n-3-as zsírsavak koncentrációjának változtatását hasonlóan jól követte (Navarro és mtsai, 1972). Az egyszeresen telítetlen olajsav szintje a takarmányban ugyancsak pozitív korrelációt mutatott a tojás olajsavkoncentrációjával (Pankey és Stadelman, 1969). A telített zsírsavak koncentrációja azonban jóval kevésbé, csak nagyon nagy arányban alkalmazott telített zsírsavakban dús zsírkiegészítés (>10%)
HVHWpQ YROW Q|YHOKHW 6XPPHUV pV PWVDL $ V]tY- és érrendszeri betegségekkel kapcsolatos epidemológiai felmérések (Sinclair, 1953;
Bang és Dyerberg, 1972) irányították rá a figyelmet az n-3-as zsírsavak szerepére az emberi egészség fenntartásában. A tojás zsírsavösszetételének változtatásában jelenleg a legfontosabb törekvés az n-3-as zsírsavak arányának növelése, ezáltal a táplálék n-6/n-3-as zsírsavarányának javítása. A „ω-3-as” vagy „n-3-DV´ WRMiV HO iOOtWiViKR]
V]iPRV WHQJHUL YDJ\ V]iUD]I|OGL HUHGHW RODM-, illetve szintetikus zsírsavkészítmény áll rendelkezésre.
$ KDORODM KDOOLV]W pV DOJD HUHGHW WHUPpNHN IHOKDV]QiOiVD VRUiQ D
tojótyúkot közvetlenül nagyobb mennyiségben EPA-val és DHA-val látjuk el. Van Elswyk és mtsai (1995) kísérletükben a 5, 10, 15, 20, 25 és 30 g/kg menhaden-halolaj kiegészítések hatékonyságát vizsgálták. Négy hét után az n-3-as zsírsavak mennyisége 8-10 mg/g tojássárgája-
NRQFHQWUiFLyW pUW HO DPLE l az EPA és DHA együttes mennyisége
PHJKDODGWD D] -linolénsavét. E kísérletben, ahogy a halolajos kísérletek többségében, a takarmány 1-nél nagyobb EPA/DHA aránya a tojásban már 0,15-re csökkent. A 1,5-3% menhaden-halolaj hatására az összes n-6- os zsírsav, különösen az arachidonsav mennyisége szignifikánsan (P<0,05) lecsökkent (Van Elswyk és mtsai, 1995). Az n-6-os zsírsavak mennyiségének 70%-os csökkenését mutatták ki Hargis és mtsai (1991) is 3% menhaden-halolaj kiegészítés alkalmazása során. A 4, 8 és 12%
menhaden-halliszt-tartalmú táp etetésének hatására a tojás sztearinsav-





![ubikinonnak, a karotinoidoknak és a glutationnak is. Az 5. ábra a Porter (1992) ál WDOOHtUWHJ\LNIHOWpWHOH]KHW UHGR[NDV]NiGIRO\DPDWRWPXWDWMD](https://thumb-eu.123doks.com/thumbv2/9dokorg/853582.45125/38.918.220.718.276.428/ubikinonnak-karotinoidoknak-glutationnak-porter-wdoohtuwhj-lnihowpwhoh-nigiro-dpdwrwpxwdwmd.webp)