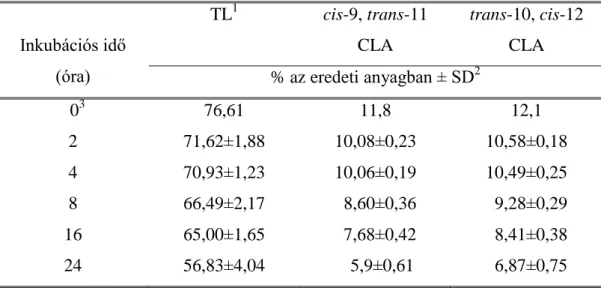
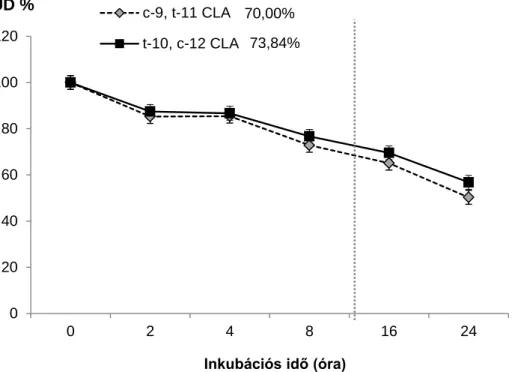
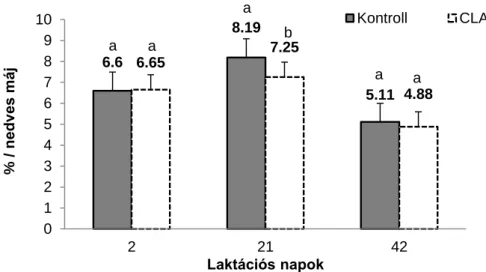
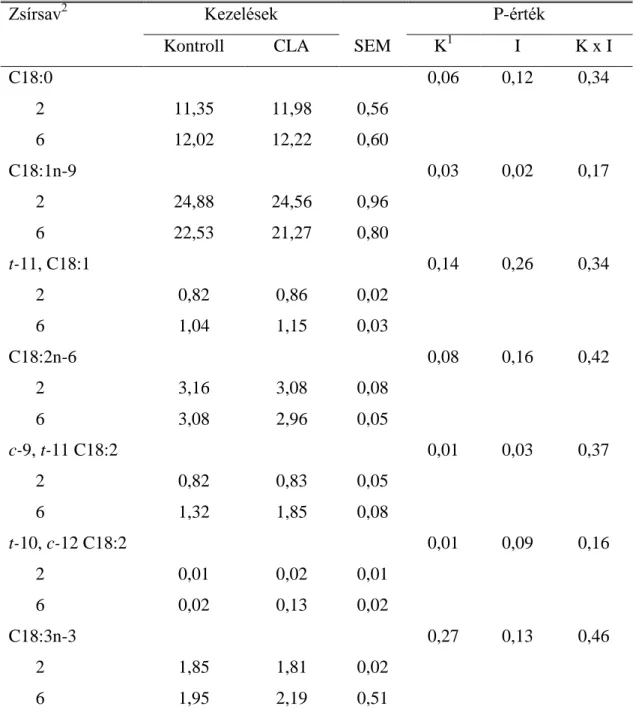
2
PANNON EGYETEM GEORGIKON KAR
ÁLLATTUDOMÁNYI ÉS ÁLLATTENYÉSZTÉSTANI TANSZÉK Állatélettani és Takarmányozástani Csoport
DOKTORI (PhD) ÉRTEKEZÉS
Készült az Állat- és Agrárkörnyezet-tudományi Doktori Iskola Keretében
VÉDETT KONJUGÁLTLINOLSAV- IZOMEREK (cis-9, trans-11;
trans-10, cis-12) SZEREPE A NAGY TEJTERMELÉSŰ KÉRŐDZŐK TAKARMÁNYOZÁSÁBAN
Készítette:
GALAMB ESZTER okl. agrármérnök
Témavezető:
DR. HUSVÉTH FERENC egyetemi tanár, az MTA doktora
KESZTHELY
2013
3 VÉDETT KONJUGÁLTLINOLSAV- IZOMEREK (cis-9, trans-11; trans-10,
cis-12) SZEREPE A NAGY TEJTERMELÉSŰ KÉRŐDZŐK TAKARMÁNYOZÁSÁBAN
Értekezés doktori (PhD) fokozat elnyerése érdekében Írta:
GALAMB ESZTER
Készült a Pannon Egyetem Állat- és Agrárkörnyezet-tudományi Doktori Iskola Keretében
Témavezető: Dr. Husvéth Ferenc
Elfogadásra javaslom (igen / nem) ..………...
(aláírás) A jelölt a doktori szigorlaton ...%-ot ért el,
Az értekezést bírálóként elfogadásra javaslom:
Bíráló neve: …... …... igen /nem
……….
(aláírás) Bíráló neve: …... …...) igen /nem
……….
(aláírás)
A jelölt az értekezés nyilvános vitáján …...%-ot ért el.
Keszthely, 2013. ……….
a Bíráló Bizottság elnöke A doktori (PhD) oklevél minősítése…...
………
Az EDHT elnöke
4
TARTALOMJEGYZÉK
RÖVIDÍTÉSEK JEGYZÉKE ... 5
1. KIVONATOK ... 6
2. IRODALMI ÁTTEKINTÉS ... 9
2.1. A KONJUGÁLT LINOLSAVAK FELÉPÍTÉSE, ÉLETTANI SZEREPE ... 9
2.1.1. A konjugáltlinolsav- izomerek szerkezete, előfordulása ... 9
2.1.2. A CLA-izomerek biológiai aktivitása ... 11
2.1.3. A CLA metabolizmusa, beépülése a szövetek lipidfrakcióiba ... 11
2.2. CLA SZEREPE A LIPIDANYAGCSERÉBEN ... 16
2.1.1. A CLA hatása a test zsírtartalmára ... 16
2.2.2. A CLA, mint a tejzsírszintézis szabályozója ... 18
2.3. A KÉRŐDZŐK ZSÍRANYAGCSERÉJÉNEK JELLEMZŐI A CLA TÜKRÉBEN ... 21
2.3.1. A tejzsírszintézis és a bendő hidrogénezési folyamatának összefüggései ... 21
2.3.2. A CLA-kiegészítés hatása a tejelő kérődzők energiaháztartására és termelési mutatóira ... 28
3. SAJÁT VIZSGÁLATOK... 31
3.1. ANYAG ÉS MÓDSZER ... 32
3.1.1. I. Kísérlet: az alkalmazott CLA-termék bendőstabilitása ... 32
3.1.2. II. Kísérlet: tejelő juhokkal végzett üzemi kísérlet ... 32
3.1.3. III. Kísérlet: tejelő tehenekkel végzett nagyüzemi kísérlet ... 32
3.1.4. Kémiai analitikai eljárások ... 39
3.1.5. Az adatok feldolgozása során alkalmazott számítások ... 41
3.1.6. Az adatok statisztikai kiértékelése ... 42
3.2. KÍSÉRLETI EREDMÉNYEK... 44
3.2.1. I. Kísérlet: az alkalmazott CLA-termék bendőstabilitása ... 44
3.2.2. II. Kísérlet: tejelő juhokkal végzett üzemi kísérlet ... 45
3.2.3. III. Kísérlet: tejelő tehenekkel végzett nagyüzemi kísérlet ... 52
3.3. EREDMÉNYEK ÉRTÉKELÉSE ... 68
3.3.1. I. Kísérlet: az alkalmazott CLA-termék bendőstabilitása ... 68
3.3.2. II. Kísérlet: tejelő juhokkal végzett üzemi kísérlet ... 68
3.3.1. III. Kísérlet: tejelő tehenekkel végzett nagyüzemi kísérlet... 71
4. ÖSSZEFOGLALÁS ... 79
5. FELHASZNÁLT IRODALMAK ... 81
6. ÚJ TUDOMÁNYOS EREDMÉNYEK ... 104
A DOKTORI ÉRTEKEZÉS TÉMAKÖRÉBEN MEGJELENT PUBLIKÁCIÓK ... 105
KÖSZÖNETNYILVÁNÍTÁS... 105
5
RÖVIDÍTÉSEK JEGYZÉKE
ADF savdetergens rost (Acid Detergent Fiber) BCS kondíciópont (Body Condition Score)
BHBA béta-hidroxi vajsav (beta-Hydroxybutyric acid) CLA konjugált linolsav (Conjugated Linoleic Acid) DIM laktációs napok (Days In Milk)
DMI szárazanyag-felvétel (Dry Matter Intake) EBAL energiamérleg (Energy Balance)
ECM energiára korrigált tejtermelés (Energy Corrected Milk production) FCM zsírra korrigált tejtermelés (Fat Corrected Milk Production)
MBW az aktív anyagcserét végző szövetek súlya, testtömeg0,75 (Metabolic Body Weight)
ME metabolizálható energia (Metabolizable Energy) MFD tejzsírdepresszió (Milk Fat Depression)
MUFA egyszeresen telítetlen zsírsav (Monounsaturated Fatty Acid) NDF neutrális detergens rost (Neutral Detergent Fiber)
NEFA nem észterifikált zsírsav (Non Esterified Fatty Acid)
NEl tejtermelésre fordított nettó energia (Net Energy for Lactation) NFC nem rosteredetű szénhidrát (Non Fiber Carbohydrates)
P4 progeszteron
PGE2 prosztaglandin-E2 PGF2α prosztaglandin-F2α
pp. ellést követő (postpartum)
PUFA többszörösen telítetlen zsírsav (Polyunsaturated Fatty Acid) RDP bendőben lebomló fehérje (Rumen degradable Protein)
RUP bendőben nem lebomló fehérje (Rumen Undegradable Protein) SFA telített zsírsav (Saturated Fatty Acid)
TG triglicerid (Triglycerides)
TL összes lipidtartalom (Total Lipid)
TMR teljes értékű takarmánykeverék (Total Mixed Ration)
6 1.
KIVONATOK
VÉDETT KONJUGÁLTLINOLSAV- IZOMEREK (cis-9, trans-11; trans-10, cis-12) SZEREPE A NAGY TEJTERMELÉSŰ KÉRŐDZŐK
TAKARMÁNYOZÁSÁBAN
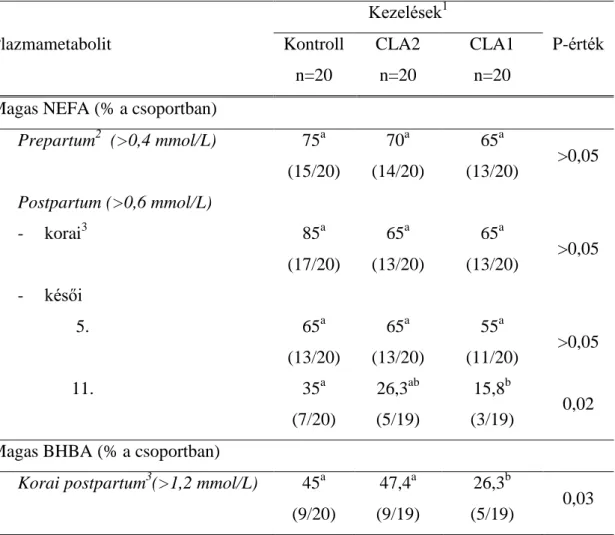
Kísérleteink során arra kerestük a választ, hogy miként befolyásolják a bendővédett konjugáltlinolsav- izomerek az intenzíven tejelő anyajuhok és tehenek tejtermelését, a termelt tej összetételét, a máj zsírtartalmát, illetve a tej- és májlipidek zsírsavösszetételének alakulását. Emellett pontos információt szerettünk volna kapni arra vonatkozóan, hogy csökkenthető-e a CLA-kiegészítés eredményeképpen a nagy tejtermelésű teheneket ellést követően jellemző, és nagy gazdasági károkat okozó negatív energiamérleg időszaka, attól függően, hogy a kiegészítést csak az ellést követően alkalmazzuk, vagy megkezdjük azt már a szárazonállás során.
Első lépésben, takarmányok bendőbeni lebonthatóságának vizsgálatára fejlesztett, in situ módszerrel határoztuk meg a további kísérleteinkhez használni kívánt CLA- készítmény izomerjeinek bendőstabilitási értékét. Ezt követően kísérletbe állítottunk 50 awassi fajtájú anyajuhot, amelyekből illesztett párok módszerével két csoportot képeztünk. Az első csoport (CLA) takarmányát 25 g CLA-termékkel egészítettük ki egyedenként, az ellést követő 2. naptól az ellést követő 42. napig. A második csoport (Kontroll) azonos energia- és nyersfehérje-tartalmú fejadagot kapott, CLA- kiegészítés nélkül. Tejelő tehenekkel végzett kísérletünket 60 holstein-fríz egyeddel hajtottuk végre azzal a különbséggel, hogy a kísérleti időszak már az ellést megelőző 21. naptól kezdődött, és a laktáció 77. napjáig tartott. A teheneket szintén illesztett párok (hármasok) módszerével osztottuk 3 csoportra, hogy lehetőségünk legyen összevetni a már ellés ellőtt megkezdett CLA-kiegészítés hatékonyságát az ellést követő CLA-kiegészítésével. A harmadik csoport CLA-kiegészítés nélküli kontroll volt. Az első csoport (CLA1) takarmányadagját az ellést megelőző 3. héttől, napi 70 g CLA termékkel egészítettük ki, egyedenként. A második csoport (CLA2) egyedei kizárólag az ellést követő időszakban részesültek CLA-kiegészítésben, amelyet az első csoporttal megegyező mennyiségben kaptak. A három kísérleti csoport takarmányadagjának energia- és fehérjetartalma mindvégig azonos volt.
7 A CLA-kezelés eredményeképpen fokozódott a tejelő juhok és tehenek tejtermelése, miközben csökkent a tej zsírtartalma. CLA-kiegészítés hatására, az anyajuhok és tejelő tehenek esetében egyaránt csökkent a máj zsírtartalma a laktáció korai, intenzív zsírmobilizációval járó szakaszában. Az ellés előtt megkezdett CLA-kezelés eredményeképpen tejelő tehenekben már a laktáció korai szakaszában kisebb mértékű volt a kondícióvesztés. Az ellést követően megkezdett CLA-kezelés hatásaként a teheneket csupán a laktáció 11. hetére jellemezte magasabb kondíciópontszám (BCS). A szárazonállás időszakában indított CLA-kezelés csökkentette a laktáció korai szakaszában a plazma NEFA-koncentrációját és a szubklinikai ketózis előfordulását. Az eredmények azt mutatják, hogy a CLA- kiegészítés alkalmazásával csökkenthető az ellést követően jelentkező lipolízis mértéke, ha az előkészítés időszakában és a laktáció korai szakaszában egyaránt alkalmazzuk a kiegészítést.
ABSTRACT
THE ROLE OF RUMEN PROTECTED CONJUGATED LINOLEIC ACID ISMERS (cis-9, trans-11; trans-10, cis-12) IN NUTRITION OF
HIGH PRODUCING DAIRY RUMINANTS
The aim of our experiments was to investigate the effect of dietary supplementation of conjugated linoleic acid isomers on milk yield, milk composition, liver fat content, and fatty acid composition of hepatic lipids and milk fat in high producing dairy ewes and cows. Moreover we intended to clarify whether CLA supplementation started prepartum or after delivery is able to decrease the length of the negative energy balance period which causes notable economic losses.
First, the ruminal stability of different CLA isomers were determined by an in situ method developed to measure feed degradability. After this, 50 multiparous Awassi ewes were allotted to two treatment groups. Diet of ewes in the CLA group was supplemented with 25 g CLA (ewe/day) containing product from d 2 to 42 postpartum while sheep in the Control group were fed an isocaloric-isonitrogenous
8 diet without CLA. The third experiment was carried out with 60 multiparous Holstein dairy cows. The experimental period started 21 days prior the expected day of calving and ended on day 77 postpartum. Animals were allotted into 3 treatment groups. The diet of CLA1 group was supplemented with 70 g (cow/day) CLA product from d 21 prepartum until 77 DIM, the diet of CLA2 was supplemented with 70 g CLA product from parturition until 77 DIM. The Control animals were fed an isocaloric-isonitrogenous diet without CLA.
CLA increased milk yield and decreased milk fat content both in sheep and cow.
During early lactation the liver fat content was lower in CLA treated sheep and cow.
Reducing the extent of body condition loss was found on week 5 and week 11 in lactation as a consequence of CLA treatment from prepartum. The postpartum CLA supplementation resulted in a higher BCS relative to Control only to week 11 in lactation. As a result of decreased BCS loss the plasma NEFA concentrations were lower and the incidence of subclinical ketosis was fewer. These results demonstrate that CLA supplementation could alleviate the body mass mobilization of high- producing dairy cows in early lactation period if the cows received CLA supplementation both during the close up and postpartum periods.
9
2. IRODALMI ÁTTEKINTÉS
2.1. A KONJUGÁLT LINOLSAVAK FELÉPÍTÉSE, ÉLETTANI SZEREPE 2.1.1. A konjugáltlinolsav- izomerek szerkezete, előfordulása
A konjugált linolsav, mozaikszóval CLA kifejezés olyan konjugált diénekre utal, amelyek a linolsav szerkezeti és geometriai izomerjeinek csoportja. A konjugált telítetlen kötésre jellemző módon, a kettős kötéseket egyes C-C kötés választja el egymástól, szemben a metilén típusú kettős kötéseket tartalmazó zsírsavakkal (pl.
linolsav; 1. ábra). A kettős kötések különböző szerkezeti helyzetekben [7, 9], [8, 10], [9, 11], [10, 12], [11, 13], magukban hordozzák mind a cis-trans, trans-cis mind pedig a cis-cis, trans-trans geometriai izoméria lehetőségét (Ha és mtsai, 1987;
Bauman és mtsai, 1999; Park és Pariza, 2007). Nomenklatúrájukban azokat a karboxilcsoporttól számított C-atomokat jelöljük, amelyeknél a kettős kötések szerepelnek (Evans és mtsai, 2002; 2. ábra).
1. ábra. A trans-10, cis-12 CLA (felül), a cis-9, trans-11 CLA (középen) és a linolsav (alul) térbeli felépítése (Pariza és mtsai, 2000)
10 A CLA-val kapcsolatos kutatások kezdetét Pariza és kutatócsoportjának vizsgálatai jelentették az amerikai Wisconsin Egyetemen (Pariza és mtsai, 1979), amikor összefüggést kerestek a marhahús karcinogén hatása és a konyhatechnikai eljárások között (hőfok, hőhatás időtartama). Meglepetésükre a sült marhahúsból származó extraktumnak antimutagén aktivitása volt megfigyelhető. Ezt a hatást a linolsavhoz hasonló szerkezetű zsírsav csoportnak, a konjugált linolsavaknak tulajdonították. Később megfigyelték, hogy linolsavból - a hő hatásán kívül - bázis által katalizált izomerizációs folyamatban is keletkezhet CLA (Ha és mtsai, 1987).
Szerkezeti felépítésüket a 2. ábra szemlélteti.
2. ábra. A linolsav, a cis-9, trans-11 CLA, valamint a trans-10, cis-12 CLA szerkezeti felépítése (Evans és mtsai, 2002)
A legjelentősebb természetes CLA-forrás a tej és kérődzők zsírja. A CLA képződésének két fő folyamata ismert, miszerint intermedierként keletkezik a bendőben a linol- illetve a linolénsavból származtatott biohidrogénezés során.
Emellett a tőgyben kimutatható Δ-9 deszaturáz aktivitásnak köszönhetően a vaccénsavból cis-9, trans-11 CLA szintetizálódhat (Griinari és Bauman, 1999a).
Habár számos CLA-izomer található meg az élelmiszerekben (tej- és tejtermékekben, kérődzők húsában) a tudomány középpontjában a legnagyobb biológiai aktivitást mutató cis-9, trans-11 CLA, valamint a trans-10, cis-12 CLA-izomerek állnak (Kramer és mtsai, 1998; Sehat és mtsai, 1999; Park és Pariza, 2007). A tejzsír összes CLA tartalmának több mint 80% -át a cis-9, trans-11 CLA teszi ki, és ugyanez az
linolsav
11 arány figyelhető meg a tejtermékek lipidfrakciójában, a kérődzők húsának intramusculáris és bőr alatti zsírjában is (Parodi és mtsai, 1977; Chin és mtsai, 1992;
1994; Sehat és mtsai, 1998). Ehhez képest az említett élelmiszerekben elhanyagolható a trans-10, cis-12 CLA mennyisége (Chin és mtsai, 1994; Park és Pariza, 1998). A kérődzők termékeiben a CLA koncentrációja összefüggést mutat azok zsírtartalmával, amely általában - a takarmányozástól függően - 3-7 mg/g zsír érték körül alakul, (Chin és mtsai, 1992). Számos vizsgálatot végeztek annak érdekében, hogy kiderítsék, befolyásolható-e a raktározási illetve a gyártási folyamatokkal a kérődző állatokból származó élelmiszerekben az adott termék CLA- tartalma, de ezek az eljárások minimális változásokat eredményeztek. Emiatt kijelenthető, hogy az élelmiszerforrások CLA-tartalmát főképp a nyersanyag CLA-
koncentrációja határozza meg (Shantha és mtsai, 1995; Banni és Martin, 1998).
2.1.2. A CLA-izomerek biológiai aktivitása
A CLA-izomerek hatásmechanizmusát az emberi és állati szervezetben, az utóbbi évtizedekben széles körben tanulmányozták (Griinari és mtsai, 2006; Park és Pariza, 2007; Bauman és mtsai, 2008). A 80-as évek végén a figyelem a CLA-izomerek antikarcinogén hatására irányult.
Az első vizsgálatokat egereken végezték, és megfigyelték, hogy a CLA gátolta a benzo(a)pirén által kiváltott epidermális neoplasia kialakulását (Ha és mtsai, 1987). Később ugyanezt bizonyították emlődaganat (Ip és mtsai, 2002) és gastrointestinalis karcinoma (Ha és mtsai, 1990) esetén is. A CLA májban megállapított antimutagén hatását a rákkeltő IQ (2-amino-3-metilimidazo [4,5 f]
kinolein) aktiválását végző citokróm P-450 enzimcsalád blokkolásával magyarázták, amit egyaránt igazoltak in vivo és in vitro körülmények között. Az IQ aktiválását a májon kívül a prosztaglandin H-szintetáz végzi, és úgy vélték, hogy a CLA gátolta ezen enzim működését is (Liew és mtsai 1995). Ezt követően - a rágcsálók mellett – baromfi fajokat is bevontak a vizsgálatokba (0,5% CLA a takarmányban), és az immunstimulációt követően a CLA gátló hatását bizonyították a katabolikus folyamatokkal szemben anélkül, hogy csökkent volna az immunválsz hatékonysága (Cook és mtsai, 1993).
Patkányokkal és egerekkel végzett kísérletekben a CLA csökkentette a fajlagos takarmány-felhasználást, szabályozta a táplálóanyagok értékesülését,
12 javította az energia-háztartást (Chin és mtsai, 1994; Park és mtsai, 1997). Sertésekkel végzett takarmányozási kísérletekben is igazolták, hogy a takarmányhoz adott CLA- kiegészítés jelentősen csökkenti a szervezetben a zsírlerakódást (Dugan és mts., 1997). A takarmányhoz adott CLA-kiegészítés csökkentette a vér LDL- koncentrációját (low density lipoprotein) olyan nyulakban és hörcsögökben, amelyeket atherogén (a vér koleszterinszintjét növelő) takarmányozásban részesítettek (Lee és mtsai, 1994; Valeille és mtsai, 2004).
A legtöbb - a CLA hatásmechanizmusát vizsgáló tanulmányban - az állatok takarmányához kevert CLA-izomerek nagy százalékát a cis-9, trans-11 CLA valamint a trans-10, cis-12 CLA tette ki, hozzávetőlegesen azonos mennyiségben (40-41%, 43-45%, az összes CLA-tartalom arányában). Éppen ezért a CLA hatásai főképp az említett 2 izomernek tulajdoníthatók (Chin és mtsai, 1992; Sehat és mtsai, 1999). Ennek ellenére beszámoltak az egyéb izomerek tejzsír-szintézisben betöltött gátló hatásairól: cis-10, trans-12 CLA, trans-9, cis-11 CLA (Sæbø és mtsai, 2005;
Perfield és mtsai, 2007), viszont ezt a folyamatot mégis egyértelműen a trans-10, cis- 12 CLA-ból származtatták (Bauman és mtsai, 2008). Emellett a trans-9, trans-11 CLA-nak szerepet tulajdonítottak a vérlemezkék összekapcsolódásának gátlásában (Al-Madaney és mtsai, 2003).
A két fő izomer interakciójával kapcsolatban számos utalás található arra vonatkozóan, hogy egymással additív kölcsönhatásban állnak, egymástól függetlenül működnek, esetleg egymás antagonistái (Park és Pariza, 2007). A proliferatív folyamatokban betöltött szerepüket tekintve feltételezték, hogy összeadódnak hatásaik (Ip és mtsai, 2002; Masso-Welch és mtsai, 2004). A testzsírdepók csökkentéséért a trans-10, cis-12 CLA-t tették felelőssé, mivel ez az izomer specifikus hatású a zsírok metabolizmusában. Közvetett úton csökkentette a lipoprotein-lipáz (kulcs enzim az adipociták zsírsavfelvételében), valamint a sztearoil-CoA deszaturáz (fő szubsztrát a zsírraktárak képződése során) enzimek aktivitását, gátolva ezáltal a lipidek beépülését a rágcsálókból származó sejtvonalakban (3T3-L1). Emellett májsejt modellben (HepG2) csökkentette az apolipoprotein B szekrécióját. Ezzel ellenkezőleg a cis-9, trans-11 CLA javította a rágcsálók növekedési erélyét, mégpedig úgy, hogy fokozta a trigliceridek akkumulációját az zsírszövetbe a (Chin és mtsai, 1994; Cook és mtsai, 1999; Park és mtsai, 1999; Choi és mtsai, 2000; Storkson és mtsai, 2005). Humán vizsgálatokban
13 azt tapasztalták, hogy a trans-10, cis-12 CLA csökkenti a sejtek inzulin által előidézett glükózfelvételét és -felhasználását, rontja a teljestest inzulinérzékenységét.
Kimutatható volt a glükóztransporter-4 (GLUT4) és az acetil-CoA karboxiláz expressziójának csökkenése. Ezzel ellenkezőleg a cis-9, trans-11 CLA kis mértékben fokozta mindezeket. A trans-10, cis-12 CLA csökkentette a sejtek TG-tartalmát azáltal, hogy mérsékelte az adipociták olajsavfelvételét, majd észterifikációját.
Gátolta emellett a transportfolyamatokban jelentős kötő fehérjék működését (acyl- CoA-binding protein, adipocita zsírsavkötő fehérje). Ezzel ellentétesen a cis-9, trans-11 CLA nem játszott szerepet a sejtek zsírsavfelvételében és -felhasználásában.
Arra a következtetésre jutottak, hogy a trans-10, cis-12 CLA-izomer hiperglikaemiát, hiperlipidaemiát okozhat (Brown és mtsai, 2003a). Ezzel szemben a cis-9, trans-11 pedig pont ellentétes hatású az említett folyamatokban (Clement és mtsai, 2002).
Több kísérletben keresték a választ arra, hogy milyen módon játszanak szerepet a konjugáltlinolsav- izomerek az immunrendszer aktiválásában. DeVoney, Pariza és Cook (1999) úgy találták, hogy a trans-10, cis-12 CLA felelős a limpfociták blastogenesisének és citotoxikus aktivitásának fokozásáért, amelyet ezt megelőzően más szerzők is bizonyítottak (Wong és mtsai, 1997). Egyéb kísérletekben a cis-9, trans-11 és a trans-10, cis-12 izomerek gátló hatását bizonyították a PGE2 szintézisében, és ezzel magyarázták az immunológiai funkciók fokozódását (Cook és mtsai, 1993). Eltérő kísérleti körülmények között, több szövetben is bizonyították a CLA PGE2-képződéssel kapcsolatos gátló hatását (Sugano és mtsai, 1997; Liu és Belury, 1998). Az eikosanoidok CLA által megváltoztatott átalakulása feltételezhetően azzal magyarázható, hogy a CLA befolyásolja a konjugált eikosatrién és eikosatetrénsav metabolizmusát. Ezt az aktivitást sikerült kimutatni a juhok májsejtjeinek foszfolipidjeiben és patkányok emlőmirigyében (Banni és mtsai, 1996). Liu és Belury (1998) a CLA tumorképződés-ellenes hatását hozták összefüggésbe az eikosanoidok szintézisében játszott szerepével. Megfigyelték ugyanis, hogy CLA-kezelés hatására - szemben a linolsavas kezeléssel - változott a membránok foszfolipidjeinek zsírsavösszetétele, csökkent a PGE2 prekurzorának, az arachidonsavnak az incorporációja. May és mtsai (2011) szarvasmarhából származó luteális sejtkultúrákban végeztek in vitro tanulmányokat, hogy választ találjanak a CLA-izomerek szaporodásbiológiai folyamatokban betöltött pozitív szerepére. Eredményeik alapján a CLA-izomerek
14 hatására csökkent mind a PGE2, mind pedig a PGF2α koncentrációja, míg a P4-é nem változott.
2.1.3. A CLA metabolizmusa, beépülése a szövetek lipidfrakcióiba
Az előző fejezetből látható, hogy a CLA-nak számos metabolikus szignál- mechanizmusban tulajdonítanak szerepet. A különböző zsírsav izomerek, esetleg metabolitjaik eltérően hathatnak az élettani folyamatokban. Fontos figyelembe vennünk tehát a CLA metabolizmusát, valamint a szövetek különböző lipidfrakcióiba történő beépülését.
Az irodalmi adatok szerint a CLA-izomerek más-más hatékonysággal épülnek be a szövetekbe lipidfrakcióiba. A különböző állatfajokkal végzett kísérletek eltérő eredményei nagy valószínűséggel az alkalmazott kiegészítések mennyiségével is magyarázhatóak. Kramer és mtsai (1998) sertések szív- és májszövetéből származó mintákban azt tapasztalták, hogy a trans-10, cis-12 CLA-izomer főként a trigliceridfrakcióba épül be. Ehhez képest a cis-9, trans-11 CLA a foszfolipidekbe inkorporálódott. Ezzel szemben Banni és mstai (2001) patkányok májából mutatták ki, hogy a cis-9, trans-11 CLA és metabolitjainak (C18:2, C18:3, C20:3) beépülése főként a neutrális zsírokba történik.
A CLA-izomerek beépültek vemhes patkányok endometriális szöveteibe, amikor takarmányukat 1,1%-ban CLA-val egészítették ki (Harris és mtsai, 2001).
Ezzel szemben tejelő tehenekkel végzett kísérletekben nem voltak kimutathatóak a CLA-izomerek a méhszövetekből, habár ez esetben az alkalmazott CLA dózis messze elmaradt a patkányokkal végzett vizsgálatokban használthoz képest (0,06% a szárazanyagban), és ennek nagy százaléka a bendőben metabolizálódott (Castaneda- Gutiérrez és mtsai, 2007). A CLA anyagcseréje hasonló az egyéb zsírsavakéhoz. Az állati szövetekben megfigyelték, hogy a CLA C-lánca elongálódhat (2 C-atomos egységekkel hosszabbodhat) és deszaturálódhat (újabb kettős kötések alakulhatnak ki), melynek eredményeképpen konjugált eikozadiének és konjugált eikozatriének keletkeznek (Banni és mtsai, 2001; Park és mtsai, 2005). Emellett az izomerek β- oxidációja által 16, 14 és 12 C-atomos konjugált, telítetlen kötéseket tartalmazó zsírsavak is képződhetnek (Ringseis és mtsai, 2006). A trans-10, cis-12 CLA hatására csökkent a C16:0, C18:1 és C20:4 zsírsavak részaránya, emellett növekedett a 22 C-atomszámú, hosszú szénláncú, telítetlen zsírsavak aránya patkányok májának
15 foszfolipid frakciójában (Sebedio és mtsai, 2001). CLA-kiegészítés hatására nőtt a tejelő tehenek follikuláris folyadék mintáiban a C18:3, C20:2 és C22:2 zsírsavak aránya (Castaneda-Gutiérrez és mtsai (2007).
Kevés információ áll rendelkezésre - a sejtekbe történő beépülés folyamatában - az egyes zsírsavak és a CLA között fennálló esetleges versengés (kompetíció) felderítésében, amely azok biológiai szerepének megértésében is segítségül szolgálhatna. Vegyük figyelembe például, hogy a zsírsavak metabolizmusa általában ugyanazon enzimrendszerek felhasználásának segítségével működik, melyek a PUFA esetében a 6, 5, 4 deszaturázok, ill. különböző elongázok. Így a linolsavból (C18:2n-6) arachidonsav (C20:4n-6) és dokozapentaénsav (C22:5n-6), az α-linolénsavból pedig eikozapentaénsav (C20:5n- 3, EPA), dokozapentaénsav (C22:5n-3, DPA) és dokozahexaénsav (C22:6n-3, DHA) keletkezik. Mivel mindkét zsírsavcsoport tagjai ugyanazt az enzimrendszert használják, így a zsírsavak metabolizmusa során a két csoport között kompetíció áll fenn (Sprecher, 1989). Az arachidonsav (C20:4n-6) és az eikozapentaénsav (C20:5n- 3) közt hasonló versengés folyik a sejtmembránba történő beépülésük során (Jump, 2002).
A CLA foszfolipidekbeni helyzetének meghatározása céljából Park és Pariza (2007) végeztek vizsgálatokat. Egérhepatocitákat foszfolipáz A2-vel reagáltattak, amely enzim specifikusan csupán az sn-2 helyzetű zsírsavak hidrolízisét végzi a foszfolipidekben. Ezt követően határozták meg a szabad zsírsavakat, ill. a lizofoszfolipid-frakció zsírsavösszetételét is. A hidrolízist követően a CLA-izomerek nagyon kis mennyisége volt mérhető az utóbbiban. Az eredményekből arra következtettek, hogy mind a cis-9, trans-11, mind pedig a trans-10, cis-12 CLA elsődlegesen sn-2 pozícióban épülnek be a szövetek foszfolipid-frakcióiba, ami az arachidonsavéval megegyező helyzet. A szerzők a CLA és az arachidonsav között negatív korrelációt figyeltek meg egerek és csirkék májában, de az izomszövetben már nem. Nagy különbség lehet tehát az eltérő szövettípusok között, amiben pedig a kezelés időtartama is szerepet játszhat. A CLA relatív kis mennyisége miatt, a CLA és a C20:4n-6 zsírsavak „helycseréjével” azonban nem magyarázható teljes mértékben az arachidonsav részarányának csökkenése. Ide kapcsolható, hogy több szerző írt a CLA-nak a ciklooxigenáz-enzimrendszer (COX) működésben betöltött
16 gátló szerepéről, amely a prosztaglandin-szintézis csökkenésére is további magyarázatul szolgál (Li és mtsai, 2005; Zhang és mtsai, 2005).
A CLA-val kapcsolatos kutatások kezdetén az izomereket szabad zsírsav formában alkalmazták, de később Terpstra és mtsai (2003) mindezt összevetették a TG-formával. A CLA mindkét kémiai forma esetén hasonlóan hatott a testösszetételre és az energiamérlegre egerekben. Park és Pariza (2007) a CLA- kiegészítéseket triglicerid, valamint diglicerid formátumban is tesztelték, és ők sem találtak különbséget - a CLA szabad zsírsavaihoz képest - a testösszetételre gyakorolt hatékonyságot illetően. Viszont a konjugált linolsavakból származtatott, nem zsírsavjellegű molekulák nem hatottak a zsírmetabolizmusra, tehát a karboxilcsoport határozza meg az aktivitásukat (Cook és mtsai, 2000). De Veth és mtsai (2004) a trans-10, cis-12 CLA izomer abomasalis infúzióját követően vizsgálták annak tejzsírcsökkentő hatását, az alkalmazott kémiai forma függvényében (metil-észter, szabad zsírsav), és nem találtak különbséget (38% és 39%). Emellett hasonló mértékben épült be az említett izomer a tejzsírba, a metil-észter és a szabad zsírsav kiegészítés mellett (18% és 19%). Mindez azt mutatja, hogy mindkét kémiai forma felszívódása hasonló hatásfokú volt.
2.2. CLA SZEREPE A LIPIDANYAGCSERÉBEN
2.1.1. A CLA hatása a test zsírtartalmára
Park és Pariza (2007) számos összetett folyamatban tulajdonítottak szerepet a CLA- nak (zsírsejtek anyagcseréjének szabályozása, a zsírsavak β-oxidációjának fokozása, a citokinek és adipokinek működésének befolyásolása), amelyek eredményeként a test zsírtartalmának csökkenését okozza. A lipidanyagcserében betöltött aktivitásért a trans-10, cis-12 izomert tették felelőssé. Megfigyelték, hogy ezen izomer átalakulása gyorsabb, mint a cis-9, trans-11 CLA-é (Park és mtsai, 1999b).
Korábban Sebedio és mtsai (1997) igazolták, hogy mindkét CLA-izomer a linolsavval megegyező módon elongálódik és deszaturálódik, éppen ezért a konjugált linolsavak az eikozanoidok prekurzorának tekinthetők. Mivel a tárgyalt CLA- izomerek szerkezete nagyban különbözik egymástól, az is feltételezhető, hogy a trans-10, cis-12 CLA-ból származó eikozanoidok funkcionálisan különböznek a cis- 9, trans-11 CLA-ból létrejövőktől (Pariza és mtsai, 2000). Ugyanezen szerzők
17 korábbi, egerekkel végzett vizsgálataikban bizonyították, hogy az eikozadién savból hevítés hatására keletkező konjugált eikozadién savak (CEA) hasonló szerepet játszanak a testösszetétel megváltoztatásában, mint maga a prekurzor CLA (Pariza és mtsai, 1998). Elképzelhetőnek tartják, hogy a CLA hatást gyakorol az eikozanoidok signálmechanizmusára, esetleg a CLA-ból származtatott eikozanoidok módosított signálokkal rendelkeznek. Mindezekből adódóan a CLA biológiai aktivitásának hatásköre kiterjedhet olyan hormonszerű mediátorok szintézisére és funkciójára, mint a citokinek (tumor-necrosis-faktor-α, TNF-α; interleukin, IL-1; Pariza, 1997). Mind a TNF-α, mind pedig az IL-1 szintézisét és működését részben az eikozanoidok, részben pedig a PGE2 által szabályozzák (Lewis, 1983). Pariza és mtsai (2000) egyértelműen igazolták, hogy a CLA módosítja a celluláris TNF-α választ. A CLA- val táplált egerek védettek voltak a TNF-α által indukált cachexia (senyvesség) ellen.
A CLA hatása a zsíranyagcserére a β-oxidáció fokozódásával is magyarázható. Rágcsálókban növelte a zsírsavak oxidációs folyamatában kulcsszerepet játszó CPT-1 enzim (karnitin-palmitoil-transzferáz-1) aktivitását (Bouthegourd és mtsai, 2002; Degrace és mtsai, 2004; Nagao és mtsai, 2005). Ezen tulajdonságok mellé sorolható a már említett lipoprotein-lipáz aktivitásának csökkentése, valamint a stearoyl-CoA deszaturáz gátlása, amelyet kimutattak több szövetben, eltérő körülmények között (Park és Pariza, 2007). Ezt a reakciót figyelheték meg tejelő tehenek tőgyében is, melynek eredménye, hogy CLA- kiegészítés hatására a tejlipidek zsírsavaiban csökken a deszaturázindex (Shingfield és mtsai, 2009).
A CLA lipolízisben betöltött szerepét igazolják a PPAR-α-hoz (peroxiszóma- proliferator aktiváló receptor-α) kapcsolódó megfigyelések is. A PPAR-α, mint nucleáris receptor-család - az adipociták proliferációjának és differenciálódásának szabályozása által - irányítja a lipogenezist és lipolízist (Gregorie, 1998). Korábbi kísérletekben kimutatták, hogy a CLA-kiegészítés nem idézte elő a peroxiszómák proliferációját, ha mindkét izomer jelen volt (Belury és Vanden Heluvel, 1999).
Brown és mtsai (2003b) arról számoltak be, hogy a humán preadipocitákban a cis-9, trans-11 CLA fokozza a PPAR-α expresszióját, szemben a trans-10, cis-12 CLA-val, amely csökkentette azt. Kísérleti adataikat összesítve a trans-10, cis-12 CLA csökkentette a preadipociták glükóz és lipid felvételét, oxidációját, differenciálódását azok géntranszkripciójának módosítása által, ami részben PPAR-α expresszió
18 csökkenésnek volt köszönhető. A CLA-kiegészítés hatásaként tehát fokozódik az adipociták és preadipociták apoptózisa, melynek eredménye, hogy - állat és humán modellekben egyaránt - csökken a zsírszövet tömege, és a zsírsejtek száma (Park és Pariza, 2007).
A legtöbb vizsgálatot, amelyekben a konjugált linolsavak hatásmechanizmusára keresték a választ, rágcsálókon végezték. Számos tényezőt kell azonban figyelembe venni annak eldöntéséhez, hogy miként illeszthetők be a gazdasági állatok takarmányozásába, vagy a különböző módszerek révén dúsított állati termékekbe (funkcionális élelmiszer), esetleg kapszulák formájában a humán táplálkozásba ezen, specifikus biológiai aktivitással rendelkező zsírsavak. Ha az alkalmazott CLA-mennyiségeket hasonlítjuk össze, akkor humán vonatkozásban 56 g CLA-t kellene elfogyasztania egy 70 kg-os embernek, hogy mindez egyenértékű legyen az egereknél használt adaggal (a táplálék 0,5%-a). Ehhez képest a humán tanulmányokban általában 0,7-6,8 g/nap CLA-t használtak (Malpuech-Brugere és mtsai, 2004). Sertésekkel végzett kísérletekben a takarmány 0,1-1%-ában alkalmazták a CLA-kiegészítést, melynek hatásaként a zsírlerakódás csökkenését, és soványhús arányának növekedését figyelték meg (Ostrowska, 1999). Humán kísérletekben lineáris kapcsolatot találtak az alkalmazott dózis emelése, illetve a testtömeg-csökkenés között (Whigham és mtsai, 2004). Nagy eltérések voltak az alkalmazott kezelések időtartamát tekintve is, habár ezzel kapcsolatban nem találtak szignifikáns különbségeket a CLA-alkalmazás eredményét illetően (Park és Pariza, 2007). Ezek mellett nehezen hasonlíthatók össze az egerek anyagcsere folyamatai az egyéb állatfajokéval, illetve az emberével. Zsíranyagcseréjük ugyanis gyorsabb, mint az egyéb fajoké (Terpstra és mtsai, 2001). Fontos figyelembe vennünk az adott élőlény energiamérlegét is, hiszen állatmodellek esetében ez általában pozitív (ad libitum takarmányfelvétel) volt, míg humán vonatkozásban energiaszegény táplálkozás mellett zajlottak a megfigyelések. Az ezzel kapcsolatos eredményeket összesítve a CLA hatékonyabban csökkentette a testzsír mennyiségét, ha pozitív volt az energiamérleg (Park és Pariza, 2007).
2.2.2. A CLA, mint a tejzsírszintézis szabályozója
A CLA-nak a test zsírtartalmának szabályozásában betöltött szerepét különböző kísérleti eredményekkel támasztották alá. Számos kutató foglalkozik ezen zsírsavak
19 tejzsír-szintézis gátlásában betöltött szerepével is, amelyhez fűződően Bauman és munkacsoportja a Cornelli Egyetemen (Cornell University, Ithaca, NY), illetve Griinari és munkatásrai a Helsinki-i Egyetemen (University of Helsinki, Helsinki, Finland) végeztek áttörő tanulmányokat (Griinari és Bauman 1999a és 1999b; 2006;
Bauman és mtsai, 2008).
A CLA tejzsírszintézist befolyásoló szerepe a kilencvenes évek végén került az érdeklődés középpontjába, amikor exogén úton, mint kiegészítőt keverték a tejelő tehenek takarmányához abból a célból, hogy növekedjen a tejzsír CLA-tartalma.
Ennek előzménye, hogy a CLA-nak rákellenes tulajdonsága mellett számos jótékony hatást tulajdonítottak humánélettani szempontból, így a CLA-val dúsított tej, mint
„funkcionális élelmiszer” szerepelhetett (Bauman és mtsai, 2008). Nem várt hatásként a CLA-izomerek keverékét tartalmazó kiegészítés azonnal, és drámaian csökkentette a tejzsír szekrécióját (Loor és Herbein, 1998; Chouinard és mtsai, 1999).
Összesítve a különböző kísérleti eredményeket elmondható, hogy a CLA- kiegészítést követően 4-5 nappal a tejzsírcsökkenés eléri a maximumát, és ezen értékeken marad mindvégig a CLA-adagolás ideje alatt, majd ezt követően visszaáll a kiindulási zsírszint (Bauman és mtsai, 2008). Mindez elmondható mind a néhány hetes (5 hét), mind pedig a hosszú időtartamú kezelésekre (20 hét) vonatkozóan egyaránt (Bernal-Santos és mtsai, 2003; Castaneda-Gutiérrez és mtsai, 2007), függetlenül a laktáció különböző stádiumától (Perfield és mtsai, 2002). Habár néhány szerző úgy találta, hogy a laktáció korai időszakában (postpartum 3 hét) a CLA hatására nem csökken a tej zsírtartalma (Bernal-Santos és mtsai, 2003), csupán ha nagyobb adagban alkalmazták azt (Moore és mtsai, 2004).
Szembetűnő, hogy - a testzsírraktárak csökkentése céljából adagolt mennyiségekhez (a takarmány 0,5-2%-a) képest - a CLA tejzsírcsökkentő hatása már meglehetősen alacsony adagolás mellett érvényesül (a takarmány <0,01-0,05%-a), viszont mindkét esetben dózis-függő módon érvényesül (Ostrowska és mtsai, 2003;
De Veth és mtsai, 2004). De Veth és mtsai (2004) 7 tanulmány eredményeit összegezték abból a célból, hogy összehasonlíthassák a CLA-kiegészítés hatékonyságát, és meghatározzák a CLA ED50 értékét tejelő tehenekben. Lineáris összefüggést találtak a trans-10, cis-12 izomer adagolt mennyisége, valamint a tejzsír termelésében bekövetkező csökkenésben. A maximális hatás eléréséhez szükséges
20 dózis fele 2,5 g/nap trans-10, cis-12 izomer volt, amely 25%-os tejzsírcsökkenést okozott, és a takarmány szárazanyag-tartalmának kb. 0,01%-át jelentette. A CLA- kiegészítés tehát legfeljebb 50%-al képes csökkenteni a tej zsírtartalmát. A dózisválasz-tanulmányok egyértelműen a trans-10, cis-12 izomert tették felelőssé a tejzsírszintézisben bekövetkező csökkenésért (Bauman és mtsai, 2008), emellett kijelentették, hogy a cis-9, trans-11 CLA-nak nincs hatása ugyanerre (Baumgard és mtsai, 2000a). Bell és Kennelly (2003) az említett ED50 érték kb. 20-szorosát alkalmazva (45 g trans-10, cis-12 CLA) megfigyelték, hogy a tejzsír mellett csökkent a tehenek tej- és tejfehérje-termelése is, emellett emelkedett a szomatikus sejtszám, valamint a tej klórtartalma. Az alkalmazott CLA-mennyiség hatására romlottak az állatok involúciós folyamatai is (apoptózis stb).
Peterson és mtsai (2004) szarvasmarha tőgyének epithel sejtjeiben (MAC-T sejtsor) a SREPB1 protein (Sterol-Response Element-Binding Protein) nagyarányú csökkenését figyelték meg, amikor a trans-10, cis-12 CLA által gátolta zsírsavak szintézisét. Ez a fehérjecsalád fontos regulátor szerepet tölt be a tőgy zsírszintézisében, transkripciós faktorként fokozza a lipogenetikus enzimek expresszióját (Harvatine és mtsai, 2007). Egerekkel végzett kísérletekben a SREPB1c gén szétesése 41%-os csökkenést okozott a tej zsírtartalmában, amely hasonló a CLA maximális hatásaként leírt 50%-hoz (Rudolph és mtsai, 2005 és 2007). A legújabb kutatási eredmények szerint juhokban a 1,5 g napi trans-10, cis-12 CLA kiegészítés hatására bekövetkező 23%-os tejzsírcsökkenés együtt járt az SREPB1 gén 55%-os csökkenésével (Hussein és mtsai, 2013).
A Spot14 gén (Thyroid Hormone Responsive, S14) – amely szintén szabályozó szerepet tölt be a lipogenetikus folyamatokban – expressziója ugyancsak csökkent a trans-10, cis-12 CLA kezelés, valamint a takarmányozás által előidézett MFD esetén. Sőt mindez megfigyelhető a tőgy mellett az adiposa szövetben is (Harvatine és Bauman, 2006).
Harvatine és mtsai (2006; 2007) több tanulmányukban kiemelték, hogy a tejzsír az egyetlen tejösszetevő, melynek szintézisét a trans-10, cis-12 CLA-kezelés gátolja. Eredményeik szerint a rövid és hosszú szénláncú zsírsavak egyaránt szignifikánsan csökkentek tej lipidfrakciójában. Mindezt úgy magyarázták, hogy a CLA gátló hatása a tőgy lipidszintézisében specifikus, és - a de novo szintézis mellett - kiterjed a takarmány eredetű zsírsavak felhasználására is. Az említett izomer
21 hatására bekövetkező tejzsírcsökkenés együtt járt az olyan lipogenetikus enzimek mRNS-génexpressziójának csökkenésével, amelyek felelősek a zsírsavak felvételéért, transzportjáért, de novo szintéziséért és a TG-ek képződéséért (Harvatine és Bauman, 2006; Harvatine és mtsai, 2007 és 2013). Mindemellett a trans-10, cis-12 CLA-kezelés csökkentette a tejzsír cis-9 C14:1/C14:0, cis-9 C16:1/C16:0 és a cis-9 C18:1/C18:0 arányait is (Shingfield és Griinari, 2007). Az eredményeket korábbi adatokkal magyarázták, miszerint az említett CLA izomer csökkentette a Δ9- deszaturáz génexpresszióját (Baumgard és mtsai, 2002; Palmquist és mtsai, 2005). A Δ9-deszaturáz enzimrendszert tették felelőssé a cis-9, trans-11 CLA endogén szintéziséért is, amelynek prekurzora a trans-11 C18:1. Ezen folyamat aktivitását figyelték meg a kérődzők adiposa szövetében és a tőgyben. A Δ9-deszaturáz-reakció lényege, hogy egy cis kettős kötés keletkezik a 9-ik és a 10-ik C-atom között. A folyamat fő szubsztrátjai: a stearoyl-CoA és a palmitolaj-CoA. Ezen biokémiai reakció termékeként keletkeznek többek között azok a zsírsavak, amelyek a membránok fluiditását fenntartó foszfolipidek fő összetevői (Bauman és mtsai, 1999;
Palmquist, 2006).
2.3. A KÉRŐDZŐK ZSÍRANYAGCSERÉJÉNEK JELLEMZŐI A CLA TÜKRÉBEN
2.3.1. A tejzsírszintézis és a bendő hidrogénezési folyamatának összefüggései A tejlipidek összetételének alakulása
A kérődzők tejében a zsír a legnagyobb változatosságot mutató összetevő. A kérődzők tejzsírjának TG-frakciója kb. 400 különböző zsírsavat foglal magában. A tejlipidek mintegy 95%-a TG formájában van jelen, a benne található zsírsavak túlnyomó részének lánchosszúsága C4-C18 közötti. A tejlipidek fennmaradó 5%-a az egyéb lipidosztályokba sorolható, úgy mint: foszfolipiek, digliceridek, monogliceridek és szabad zsírsavak (Jensen és msati, 1991).
A kérődzők tejzsírját felépítő zsírsavak egy része, a C4-C14, valamint a C16 bizonyos százaléka a tőgy mirigyhámsejtjeiben zajló de novo szintézisből származik, amelynek prekurzora az ecetsav és kisebb mértékben a BHBA. Az egyéb C16, illetve a hosszabb szénláncú zsírsavak a vérkeringésből kerülnek a tejmirigybe, amelyek
22 lehetnek takarmány eredetűek és származhatnak az endogén zsírraktárak mozgósításából (Bauman és Davis, 1974; Palmquist, 2006).
A takarmányozás a legfontosabb környezeti tényező, amellyel hatást gyakorolhatunk a tej zsírtartalmára, egyúttal eszközül szolgál a tejtermelés, valamint a tej összetételének megváltoztatására (Bauman és Griinari, 2001). Bizonyított, hogy a tejlipidek zsírsavainak akár 90%-a is származhat a takarmányból (Palmquist és Jenkis, 1980), amely azzal magyarázható, hogy a felszívódott zsírsavak egy része a nyirokáram útján – a máj megkerülésével – jut a tőgybe, közvetlenül bekapcsolódva a tejzsírszintézisbe (Palmquist, 2006).
Az alacsonytejzsír-szindróma kiváltó okai
A takarmányozás által kiváltott, alacsonytejzsír-szindrómával kapcsolatos kutatások évtizedek óta a figyelem középpontjában állnak. Már a múlt század elején megfigyelték, hogy jelentősen csökken a tej zsírtartalma, ha a tejelő szarvasmarhák takarmányát halolajjal (Drummond és mtsai, 1924), vagy növényi olajjal (Williams és mtsai, 1939) egészítették ki. Másrészről, ha az állatokat kicsi rosttartalmú és magas abrakhányadú (HG/LR, High Grain/Low Roughage) fejadaggal etették (Powell, 1939; Loosli és mtsai, 1945), esetleg ha a takarmány rosthordozó komponenseit darálták (Powell, 1939).
Powell és mtsai (1939) elsőként figyeltek fel arra, hogy a takarmányozás által előidézett MFD mellett megváltoznak a bendőben zajló mikrobiális folyamatok.
Pozitív korrelációt tapasztaltak a bendőműködés intenzitása, és a tejösszetevők termelődése között. Mindezt legjobban a HG/LR, és a darált vagy pelletált rostkomponenseket tartalmazó takarmányfejadag etetése szemléltette, melynek hatására csökkent a bendő pH értéke, és megváltozott a keletkezett illózsírsavak moláris aránya (Van Soest, 1963). Abban az esetben, ha az említett tényezők mellett pufferrt adtak a takarmányhoz, minimális változások léptek fel a bendőműködésben, és a tejzsír aránya is normál értéken maradt (Davis és mtsai, 1964).
Hasonlóképp változott a bendőfermentáció, ha a tejzsír csökkenését halolajjal, vagy növényi olajokkal váltották ki (Storry, 1981; Doreau és mtsai, 1999).
Minimális hatással voltak azonban a tejzsírtermelésre a hidrogénezett olajok (Brumby és mtsai, 1972), és az egyéb módon bendővédetté tett olajok, avagy ha az olajkiegészítést abomasalisan alkalmazták (Rindsig és Schultz, 1974; Pennington és
23 Davis, 1975; Astrup és mtsai, 1976). Az olajkiegészítések abomasalis infúziója, esetleg bendővédett formában való alkalmazása gyakran okozott változást a tejlipidek zsírsavösszetételében, de általában mindez a tejzsírtermelés fokozódásával, mintsem annak csökkenésével járt (Doreau és mtsai, 1997).
A transzsírsav-elmélet
Moore és Williams (1963) elsőként figyelték meg, hogy a trans-C18:1 zsírsavak aránya növekedett a tej lipid-frakciójában, amikor a teheneket alacsony rost- és magas gyapotmagolaj-tartalmú takarmánnyal etették. Storry és Rook (1965) két évvel később ugyancsak beszámolt a trans-C18:1 zsírsavak részarányának növekedéséről, amikor az állatokat a klasszikus HG/LR fejadagot alkalmazták, így okozván a tej zsírtartalmának csökkenését. Mindezzel megegyező módon, duodenum fisztulával ellátott tehenekkel végzett vizsgálatok igazolták, hogy a duodénumtartalom trans-C18:1-zsírsavtartalma többszöröse volt az adott takarmányénak, amellyel kiváltották az MFD-t (Wonsil és mtsai, 1994). A kilencvenes évek végén vált csak nyilvánvalóvá, hogy a tejzsír csökkenése a tejlipidek trans-10 C18:1 zsírsavainak növekedésével áll összefüggésben, és nem az egész trans-C18:1-es csoporttal (Griinari és mtsai, 1998). Nem okozott változást a tejzsírban a trans-9 C18:1 (Rindsig és Schultz, 1974), a trans-11 C18:1 és a trans-12 C18:1 zsírsavak oltógyomorba történő infúziója sem (Griinari és mtsai, 1998).
Selner és Schultz (1980) beszámoltak arról, hogy a takarmány 500 ml olajsavval történő kiegészítése mellett sem csökkent a tej zsírtartalma, habár a trans- C18:1 zsírsavak mennyisége 3-szorosára növekedett a tejzsírban. A trans-10 C18:1 zsírsavak részarányának növekedése a duodenális folyadékban és a tejzsírban pedig minden esetben megfigyelhető volt, amikor a teheneket tejzsír csökkenést kiváltó fejadaggal takarmányozták (Griinari és mtsai, 1998; Piperova és mtsai, 2002; Loor és mtsai, 2004). Mindezért a bendőben zajló hidrogénezési folyamatokat tették felelőssé, melynek metabolitjai a trans-10 C18:1 zsírsavak, amelyek közvetlenül vagy közvetve szabályozzák a tejmirigy lipogenesisét. A folyamat kiinduló anyagai pedig olyan, többszörösen telítetlen zsírsavak, amelyeket a növényi olajok, a halolaj valamint a gabonamagvak tartalmaznak (Griinari és mtsai, 1997; 1998).
Az első közvetlen vizsgálatot, amely a fentiekben részletezett hipotézist volt hivatott igazolni, Lock és mtsai (2007) végezték, akik 95%-os tisztaságú trans-10
24 C18:1 zsírsavakat infundáltak tejelő tehenek oltógyomrába 4 napon keresztül (42,6 g/nap). Azt tapasztalták, hogy a tejben 0,47 g-ról 1,11 g-ra növekedtek a trans-10 C18:1 zsírsav izomerek (g/100 g zsírsav mennyiségben), de nem változott a tej zsírtartalma. Az alkalmazott zsírsavinfúzió túlságosan kismértékű lehetett ahhoz, hogy befolyásolja a tőgyben zajló lipogenetikus folyamatokat (Kadegowa és mtsai 2008). Shingfield és mtsai (2009) mindezt az alkalmazott kiegészítés magas olvadáspontjával magyarázták, ami 52-53ºC volt, és 15%-os hatékonysággal jelent meg a tejzsírban. Ezzel ellentétben, a szabad zsírsavakhoz képest alacsonyabb olvadáspontú zsírsav metil-észter formájában alkalmazva a trans-10 C18:1 zsírsav infúziót (37,3 g/nap), a tejzsír 19,5%-al csökkent (Shingfield és mtsai, 2009). Ezt támasztja alá az a megfigyelés, hogy kisebb mértékű volt a trans-C18:1 zsírsavak felszívódása a duodénum és az ileum között, ha hidrogénezett faggyút is tartalmazott a takarmány, aminek az olvadáspontja magasabb volt az állat testhőmérsékleténél, melynek következtében csökkent a micellák oldhatósága (Glasser és mtsai, 2008).
A bendőben zajló hidrogénezési folyamat
A takarmányozásban alkalmazott gabona magvak lipid összetevőit főképp glikolipidek és foszfolipidek alkotják, melyekben a telítetlen zsírsavak nagy részét a linolénsav (C18:3n-3) és a linolsav (C18:2n-6) teszik ki. Ezzel ellenkezőleg az olajos magvakban a zsírfrakció többnyire trigliceridek formájában van jelen, melynek fő zsírsavai a linolsav és az olajsav (C18:1n-9). Mindezek a bendőbe kerülésüket követően két fő átalakuláson esnek át. Elsőként hidrolizálódnak az észter kötések, amelyet a mikrobális eredetű lipáz enzim katalizál, majd a második lépésben a hidrogénezési folyamat által telítődnek a telítetlen kötések (Dawson és Kemp, 1970;
Dawson és mtsai, 1977). Ez utóbbiért főképp az előgyomrokban működő baktériumok felelősek, a protozoáknak csupán kis szerepük van ebben (Harfoot és Hazlewood, 1988).
Kepler és mtsai (1966) kizárólag a Butyririvibrio fibrisolvens baktériumnak tulajdonítottak hidrogénezési kapacitást. Kemp és Lander (1984) a biohidrogénezésben betöltött szerepük alapján 2 csoportba sorolták a baktériumokat.
Az „A”-csoportba sorolt baktériumok hidrogénezik a linolsavat és az α-linolénsavat, a folyamat fő terméke a trans-11 C18:1. A „B”-csoportba sorolt baktériumok fő szubsztrátként a trans-11 C18:1 zsírsavat használják, amelyből sztearinsavat
25 állítanak elő (C18:0). A telítetlen zsírsavak intenzív bendőbeli átalakulása az oka annak, hogy a sztearinsav adja a duodenális folyadék zsírsavainak nagy részét.
Mindez annak ellenére így van, hogy a kérődzők takarmányaiban főképp 18 C- atomszámú, többszörösen telítetlen zsírsavak szerepelnek, melyek a takarmányozás függvényében: linolsav és olsajsav (intenzív takarmányozás), linolénsav (legeltetés) (Bauman és Lock, 2006).
3. ábra. A linolsav bendőben zajló hidrogénezési folyamata normál feltételek mellett, és MFD-t (Milk Fat Depression) kiváltó takarmányozási körülmények között (szaggatott nyíl; Bauman és Lock, 2006)
Doreau és Ferlay (1994) kérődzők bendőbeni zsíranyagcseréjére vonatkozó vizsgálataik szerint a cis-9 kettős kötés izomerizációjának eredményeképpen trans- 10, cis-12 CLA keletkezik. Ezt követi a cis-12-es kötés telítődése, amely a trans-10 C18:1 zsírsavakat adja. Griinari és mtsai (1999a) lineáris kapcsolatot találtak a tejzsír trans-10 C18:1 zsírsav és a trans-10, cis-12 CLA koncentrációja között (r2=0,70).
Megfigyelték, hogy a tejzsír csökkenésével arányosan növekedett a tej lipid frakciójában a trans-10, cis-12 CLA koncentrációja, amikor a teheneket HG/LR fejadaggal takarmányozták, vagy a takarmányukhoz napraforgó olajat adtak.
Végeredményül, a bendő ökoszisztéma változását eredményezik a speciális, MFD-t kiváltó takarmányozási módszerek, melynek köszönhetően fokozódik a bendőben a
módosított bendőfermentáció
26 trans-10, cis-12 CLA termelése, ami aztán a felszívódást követően megjelenik a tejzsírban és az egyéb szövetekben is (3. ábra; Bauman és Lock, 2006). Ugyanezt támasztják alá a húsmarhákkal végzett kísérletek, amelyek jól szemléltetik, hogy abban az esetben, amikor a kérődzőket magas abrakarányú fejadaggal etették - azaz intenzíven hizlalták -, zsírszövetükben magasabb volt a trans-10, cis-12 CLA aránya a legeltetett, vagy magas rosttartalmú takarmányokkal etetett szarvasmarhákéhoz képest (Dhiman és mtsai, 1999; Griinari és Bauman 1999a).
A szintetikusan előállított konjugáltlinolsav-izomerek keverékének (trans-10, cis-12 CLA, cis-9, trans-11 CLA) abomasalis infúzióját követően dózisfüggő módon tapasztalták a tejzsír csokkenését. Az említett kezelés 50%-os tejzsírcsökkenést okozott, ha 30 g napi adagban alkalmazva (Chouinard és mtsai, 1999). Ehhez képest, a trans-10, cis-12 CLA viszonylag kis mennyiségben való adagolását követően (3,5 g/nap) 25%-al esett vissza a tej zsírtartalma (Baumgard és msati, 2000a). Ugyanezt az izomert napi 6g-os mennyiségben, postruminálisan infundálva, 41,4%-os tejzsírcsökkenést idézett elő (de Veth és mtsai, 2004). A cis-9, trans-11 CLA 10 g/nap dózisban sem hatott a tejzsír szintézisére az oltógyomorba való infúzióját követően (Baumgard és mtsai, 2000b), habár a tejzsír CLA izomerjeiben meghatározónak tekinthető <2%, míg a trans-10, cis-12 CLA csupán <0,06% (az összes zsírsav százalékában; Bauman és mtsai, 2000).
A trans-10, cis-12 CLA különböző takarmányozási módszerek eredményeképpen keletkezik, mint a bendő fermentációs folyamatainak intermedier vegyülete, és inhibitor szerepet tölt be a tejmirigyben zajló zsírszintézisben (Bauman és Griinari, 2001). A cis-10, trans-12 (Sæbø és mtsai, 2005) és a trans-9, cis-11 CLA-izomerek ugyancsak intermedierként képződnek a linolsav bendőbeli hidrogénezését követően (Wallace és mtsai, 2007), és szintén gátló hatásúak a tőgy lipogenetikus folyamataira, viszont nem magyarázzák a takarmányozás által kiváltott MFD-t, de legalábbis ez idáig még feltáratlan a mechanizmusuk (Shingfield és Griinari, 2007).
Minden, tejzsír csökkentésére irányuló takarmányozási vizsgálat alkalmával fokozódott a bendőfolyadék trans-10 C18:1 zsírsavainak mennyisége (Shinfield és Griinari, 2007), és ugyanúgy a tejlipidek trans-10 C18:1 zsírsavtartalma is (Loor és mtsai, 2005; Shinfield és Griinari, 2007). Az azonban nem bizonyított, hogy ez közvetlen hatás eredményeképpen érvényesül-e, avagy amiatt, hogy szintézisük a
27 bendőben együtt jár olyan CLA-izomerek (trans-10, cis-12 CLA, cis-10, trans-12 CLA, trans-9, cis-11 CLA) keletkezésével, amelyek bizonyítottan gátolják a tőgy lipogenetikus folyamatait. Ez nem azt jelenti, hogy a trans-10 C18:1 zsírsavaknak nincs szerepe a tejzsírszintézisében, de mindenesetre hatásfokukat tekintve 40-50- szer kisebb, mint a trans-10, cis-12 CLA-é (Shingfield és mtsai, 2009).
A védett CLA-kiegészítés hatása különböző kérődző fajokban
Az előzőekből látható, hogy a CLA-izomerek kellő mértékű felszívódásának érdekében, azokat a bendőbeni átalakulás ellen védeni szükséges.
A kérődzők tejzsírszintézise nagy hasonlóságot mutat, bár bizonyos zsírkiegészítések eredményeképpen jelentkező változások - a tejelő tehenekhez képest - különböznek tejelő juhokban és kecskékben (Chilliard és mtsai, 2003).
Tejelő tehenekben a 20-25%-os tejzsírcsökkenés eléréséhez 2-2,7 g/nap trans-10, cis-12 CLA postruminális adagolása szükséges, amely az említett izomer 10-14 g/nap per os adagolt mennyiségével biztosítható (de Veth és mtsai, 2004). Tejelő tehenekkel végzett transzformációs hatékonysági vizsgálatokat az állatok MBW- jének figyelembe vételével adaptálták tejelő juhokra. Megállapították, hogy napi 2,4 g trans-10, cis-12 CLA adagolását követően 0,4 g mennyiség jut el a juhok oltógyomráig, ami hasonló arányú tejzsír csökkenést (23%) váltott ki, mint tehenekben a fent említett, 10-14 g/napos dózis (Lock és mtsai, 2006). A lipid kapszulázott CLA-kezelés hatására a trans-10, cis-12 CLA 2,6-7,9%-os transzformációs hatékonysággal épült be tejelő tehenek tejzsírjába (Perfield és mtsai, 2004). Az ugyanilyen módon bendővédetté tett CLA kiegészítést alkalmazva tejelő juhoknál, 3,8%-os hatékonyságot figyeltek meg (Lock és mtsai, 2006). Mindez tejelő kecskékben – az MBW alapján meghatározott CLA-dózis alkalmazását követően – csupán 1,8% volt, és 5% -os tejzsírcsökkenést idézett elő (Lock és mtsai, 2008).
Fontos figyelembe vennünk, hogy a kérődzők takarmányozásában alkalmazott zsírsav-kiegészítések átalakulását az adott zsírkészítmény bendőstabilitásán kívül, azok postruminális emészthetősége is befolyásolja (Wu és Papas, 1997). Ebből adódóan a kecskéknél tapasztalt, csökkent CLA-hatékonyság az állatok eltérő emésztési folyamataival magyarázható, ami adódhatott a CLA intenzívebb bendőbeni átalakulásából, és/vagy a vékonybélből történő alacsonyabb mértékű felszívódásából (Lock és mtsai, 2008). A trans-10, cis-12 CLA 17,8%-os
28 transzformációs hatásfokát írták le kecskékben az adott izomer duodenális infúzióját követően (Lock és mtsai, 2008), ami hasonló a tejelő teheneknél tapasztalt 21,7%- hoz (de Veth és mtsai, 2004). Mindez azt mutatja, hogy kecskék bendőjében nagyobb mértékben hidrogéneződnek a takarmányozás útján bevitt CLA izomerek, mint a tehenekben.
2.3.2. A CLA-kiegészítés hatása a tejelő kérődzők energiaháztartására és termelési mutatóira
A laktáció kezdeti szakaszában a takarmánnyal felvett energia általában nem fedezi a tejelő tehenek energiaigényét. A tejtermelés valamint a létfenntartás együttes energia-szükséglete felülmúlja a felvett szárazanyag energiatartalmát. Ezért ebben az állapotban az állatok energiamérlege negatív (Bell, 1995; Grummer, 1995).
Mindezek következtében az endogén raktárak mobilizálásából származó szabad zsírsavak felhasználásával fedezik a hiányzó energiát (Bauman és Currie, 1980), amely extrém esetben számos metabolikus zavar és betegség okozója (ketózis, zsírmáj szindróma) az ellés körüli időszakban (Gröhn, 1995; Drackely és mtsai; 1999 és 2001). Emellett rontja a szaporodási teljesítményt (Butler, 2000 és 2003; Lucy, 2003).
Az adiposa szövetben TG formájában raktározott zsír szubsztátul szolgál az energiatermelő folyamatokban a negatív energiamérleg időszakában (Husvéth és mtsai, 1982; Bell, 1995). Ilyenkor a raktározott TG-ek glicerinre és szabad zsírsavakra bomlanak le (lipolízis), majd a NEFA a keringésbe áramlik, és ott a vérplazma albumin frakciójával komplexet képezve a máj és más szervek felé szállítódik (Husvéth és mtsai, 1982; Rukkwamsuk és mtsai, 1998). A vérplazma NEFA-koncentrációja a zsírmobilizáció mértékével, és ezáltal a máj TL- és TG- tartalmával, valamint a ketonanyagok felszaporodásával szorosan összefügg (Husvéth és mtsai, 1982; Elek és mtsai, 2008). Az plazma élettani határértéknek tekinthető NEFA-koncentrációja az ellést megelőző három hét alatt ≤0,4 mmol/l, a laktáció korai szakaszában pedig ≤0,6 mmol/l (Eicher, 2003). A nagymértékű NEFA- beáramlás fokozza a májsejtekben a lipogenezist (TG-szintézis) és a ketogenezist (acetecetsav, BHBA és aceton képződés). A zsírsavak – a májsejtek mitokondriumába kerülésüket követően – a β-oxidáció által acetil-CoA-ra bomlanak.
A nagy mennyiségben keletkező acetil-CoA – a glükoneogenezisben elhasznált
29 oxálecetsav hiánya miatt – nem tud belépni a citromsavciklusba, melynek hatására fokozódik a ketontesttermelés (Zammit, 1990). A nagy tejtermelésű tehenek a laktáció 3. és 4. hetében a legfogékonyabbak a ketózis kialakulására, melynek szubklinikai és klinikai formája egyaránt ismert (Simensen és mtsai, 1990;
Ingvarsten, 2006). Abban az esetben, ha a vérplazma BHBA-koncentrációja eléri az 1,2 mmol/l-t, szubklinikai ketózisról beszélünk, viszont ha mindehhez a plazma alacsony glükózkoncentrációja társul (≤2,5 mmol/l), klinikai tünetekké manifesztálódik (Simensen és mtsai, 1990; LeBlanc és mtsai, 2005). Ilyenek a szárazanyag-felvétel és a tejtermelés hirtelen csökkenése, bódultság, kondícióvesztés (Drackely és mtsai, 2001).
Tejelő kérődzőkben a CLA-izomerek, különösen a trans-10, cis-12 CLA befolyásolják az energia-metabolizmust, az MFD kiváltása útján (Baumgard és mtsai, 2000b; Lock és mtsai, 2006 és 2008; Bauman és mtsai, 2008). Gyakran a kisebb mértékű tejzsírszintézis által „megspórolt” energia tej termelésére fordítódik, amely ezáltal – alacsonyabb tejzsírtartalom mellett – nagyobb tejtermelésben mutatkozik meg (Bernal-Santos és mtsai, 2003; de Veth és mtsai, 2006). Mivel a tejtermelés energiaszükségletének több, mint 50%-a fordítódik tejzsírszintézisre (Tyrrell és Reid, 1965), a CLA-kiegészítés alkalmazása tejelő tehenek takarmányában egyedülálló lehetőséget nyújt a laktáció korai szakaszában - a mérsékelt energia leadás révén - javítani az energiamérleget, ezáltal a reprodukciós mutatókat (Griinari és Bauman, 2006).
Bernal-Santos (2003) és mtsai 30,4 g/nap Ca-szappan formában védetté tett CLA-val egészítették ki tejelő tehenek takarmányát a várható ellést megelőző második héttől a laktáció 20 hetén keresztül. A tejtermelés növekedését tapasztalták (3 kg/nap) a tej zsírtartalmának csökkenése mellett (12,5%). Mindezt azonban csupán a laktáció második hetétől. Nem változott a tehenek szárazanyag-felvétele, energiamérlege és a tej fehérjetartalma. Nem találtak szignifikáns különbségeket az állatok energiaállapotát jellemző vérplazmamutatókban (NEFA, BHBA, glükóz), valamint a máj triglicerid- és glikogéntartalmában. Eredményeiket azzal magyarázták, hogy a CLA-kiegészítés hatására csökkent a tejzsírtermelés, és az így megspórolt energia-leadás révén az állatok több tejet termeltek, mintsem hogy az energiamérlegük javult volna.
30 Castaneda-Gutiérrez és mtsai (2005) hasonló kémia formában és dózisban alkalmaztak CLA-kiegészítést (31,6 g/nap) a várható ellést megelőző második héttől az ellést követő kilencedik hétig, amely 11%-os tejzsír csökkenést idézett elő tejelő tehenekben. A 63,2 g-os CLA adagolás 21%-ban csökkentette a tej zsírtartalmát.
Mindkét csoport esetében csupán a laktáció harmadik hetétől volt megfigyelhető a CLA tejzsírszintézist gátló hatása. A tejtermelésben és a tej egyéb összetevői között nem találtak különbséget. A magasabb dózisú CLA-kiegészítés csökkentette a tejjel leadott energia mennyiségét, viszont nem változott az állatok energiamérlege, testsúlya és kondíciója. A laktáció első hetében szignifikánsan alacsonyabb plazma NEFA-értékeket mértek a kisebb mennyiségű CLA-kiegészítés hatására, viszont nem tapasztaltak különbséget az inzulin- és glükózszintekben, valamint a máj triglicerid- tartalmában sem.
31
3. SAJÁT VIZSGÁLATOK
A szakirodalmi adatok ismeretében olyan új tudományos eredmények elérése volt a célunk, melyek segítségével közelebb kerülhetünk a szintetikusan előállított CLA- izomerek kérődzőkben lejátszódó hatásmechanizmusának megismeréséhez, különös tekintettel az üzemi körülményekre. Ennek okán szükségünk volt egy olyan, CLA- izomereket magas hányadban tartalmazó zsírkészítményre, amely ellenáll a bendőben zajló biohidrogénezési folyamatoknak, ennek köszönhetően képes bekapcsolódni a gazdaszervezet intermedier anyagcseréjébe. Ezt követően arra kerestünk választ, hogy az említett linolsavizomerek etetését követően miként változnak az intenzíven tejelő kérődzők termelési mutatói, valamint a termelt tej összetétele üzemi körülmények között. Tekintettel a CLA-izomerek hatásait jellemező faji különbségekre, vizsgálatainkba kiskérődzőket és tejelő teheneket egyaránt bevontunk.
Az I. kísérletben in situ körülmények között határoztuk meg a vizsgálatainkhoz használni kívánt CLA-készítmény izomerjeinek bendőstabilitási értékét.
A II. kísérlet alkalmával vizsgáltuk az I. kísérletben tesztelt, magas bendőstabilitási értékeket mutató CLA-termék etetésének hatását az intenzíven tejelő anyajuhok tejtermelésére, tejösszetételére, a máj zsírtartalmára, illetve a tej- és májlipidek zsírsavösszetételének alakulására.
A III. kísérletben a CLA-etetés optimális időszakát kívántuk meghatározni nagy tejtermelésű Holstein-fríz állományokban. Ennek érdekében összehasonlítást végeztünk a CLA-izomerek hatását illetően a már előkészítés alatt alkalmazott CLA- kiegészítés, illetve a kizárólag ellést követően induló CLA-kiegészítés között.
Tanulmányoztuk a CLA-izomerek etetésének hatását a nagy tejtermelésű tehenek termelési mutatóira, a máj zsírtartalmára, valamint a metabolikus állapotot leginkább jellemző vérparaméterek alakulására. Emellett vizsgáltuk a tej- és májlipidek zsírsavösszetételének változásait.
![linolsav; 1. ábra). A kettős kötések különböző szerkezeti helyzetekben [7, 9], [8, 10], [9, 11], [10, 12], [11, 13], magukban hordozzák mind a cis-trans, trans-cis mind pedig a cis-cis, trans-trans geometriai izoméria lehetőségét (Ha](https://thumb-eu.123doks.com/thumbv2/9dokorg/853270.45103/9.892.304.625.668.1097/kötések-különböző-szerkezeti-helyzetekben-hordozzák-geometriai-izoméria-lehetőségét.webp)