Bentikus és planktonikus algaközösségek fotoszintézisének karakterisztikái
DOKTORI ÉRTEKEZÉS
Üveges Viktória
Témavezetı:
Prof. Dr. Padisák Judit, intézetigazgató egyetemi tanár, az MTA doktora Pannon Egyetem, Környezettudományi Intézet,
Limnológia Intézeti Tanszék
Pannon Egyetem
Kémia és Környezettudományi Doktori Iskola Veszprém
2010
Bentikus és planktonikus algaközösségek fotoszintézisének karakterisztikái
Írta:
Üveges Viktória
Készült a Pannon Egyetem Kémia és Környezettudományi Doktori Iskolája keretében Témavezetı: Prof. Dr. Padisák Judit
Elfogadásra javaslom (igen / nem)
……….
(aláírás) A jelölt a doktori szigorlaton …... % -ot ért el,
Az értekezést bírálóként elfogadásra javaslom:
Bíráló neve: ……… ………..igen /nem
……….
(aláírás) Bíráló neve: ……… ………..igen /nem
……….
(aláírás) A jelölt az értekezés nyilvános vitáján …...%-ot ért el.
Veszprém,
……….
a Bíráló Bizottság elnöke A doktori (PhD) oklevél minısítése…...
………
Az EDHT elnöke
1
Tartalomjegyzék
Rövidítések jegyzéke ... 3
Kivonat ... 4
Abstract ... 5
Zusammenfassung ... 6
1 Irodalmi áttekintés: édesvízi algaközösségek jelentısége, mőködése, vizsgálatuk ... 7
1.1 Bentikus algaegyüttesek és szabályozó tényezıi ... 7
1.2 Fitoplankton közösségek és szabályozó tényezıi ... 13
1.3 A fotoszintézis kutatások elméleti háttere ... 14
1.4 Nemzetközi bevonatalga fotoszintézis- és elsıdleges termelés kutatások ... 16
1.5 Nemzetközi fitoplankton fotoszintézis és elsıdleges termelés kutatások ... 18
1.6 Hazai alga fotoszintézis és elsıdleges termelés kutatások ... 19
2 Problémafelvetés és célkitőzés ... 20
3 Anyag és módszer ... 22
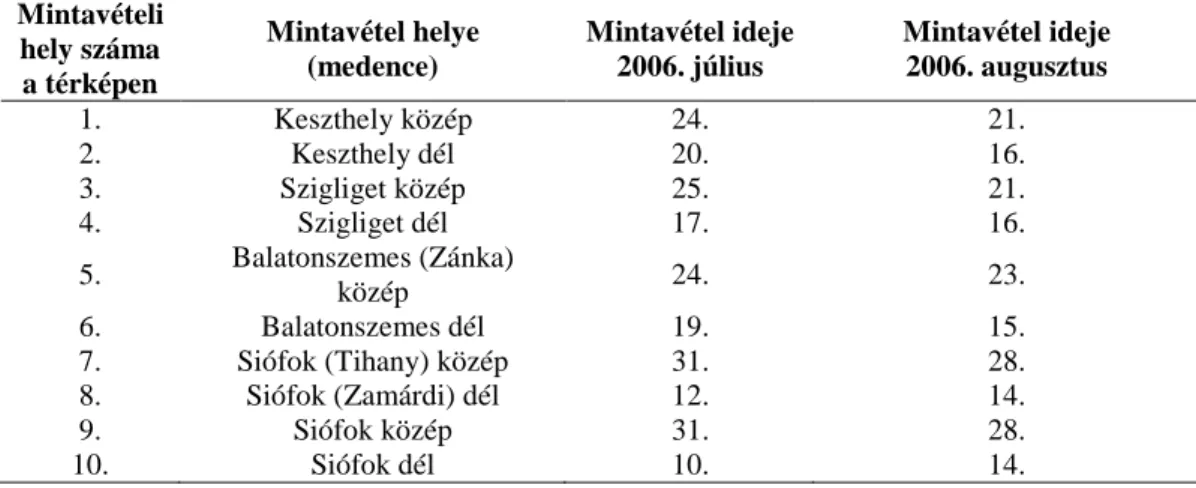
3.1 Balatoni epipszammon és epipelon fotoszintézise és elsıdleges termelése ... 22
3.1.1 A mintavétel helye és ideje ... 22
3.1.2 Inkubáló berendezés ... 23
3.1.3 Fotoszintézis mérések ... 23
3.1.4 Pigmentanalízis ... 24
3.1.5 Elsıdleges termelés becslése ... 24
3.2 Patak epiliton fotoszintézise és elsıdleges termelése ... 24
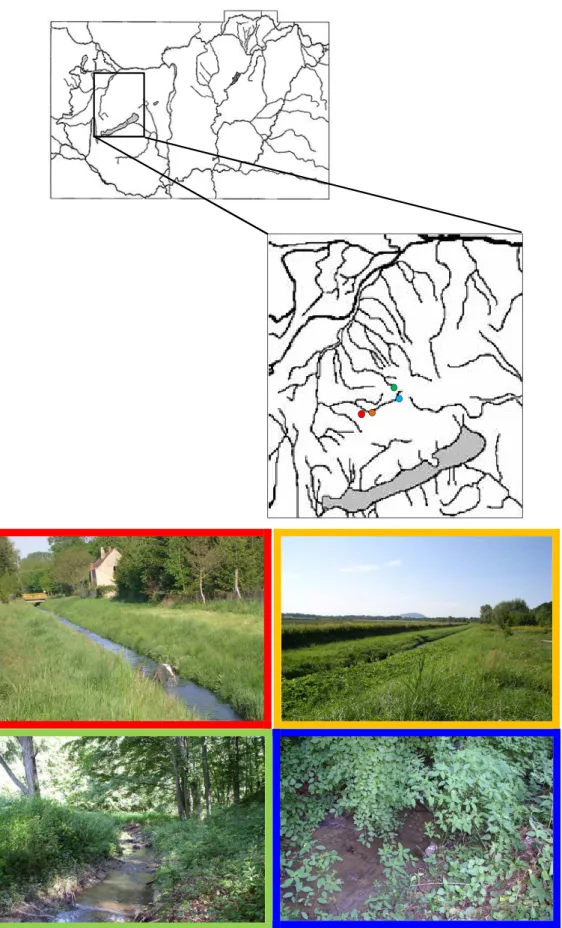
3.2.1 A mintavétel helye és ideje ... 24
3.2.2 Szubsztrátok elıkészítése, kihelyezése a mintavételi helyekre ... 27
3.2.3 Fotoszintézis mérések ... 27
3.2.4 Pigmenttartalom meghatározása ... 28
3.3 Patak epiliton elsıdleges termelésének becslése ... 29
3.4 Stechlin-tó téli Aphanizomenon flos-aquae virágzásának fotoszintézis karakterisztikája és elsıdleges termelése ... 29
3.4.1 Mintavételi hely bemutatása ... 29
3.4.2 Mintavétel, minta-elıkészítés ... 29
3.4.3 Fotoszintézis mérése ... 29
3.4.4 Fitoplankton elsıdleges termelés becslése ... 30
4 Eredmények ... 31
4.1 A Balaton epipszammon és epipelon közösségeinek fotoszintézise és elsıdleges termelése ... 31
4.1.1 Biomassza ... 31
4.1.2 P-I karakterisztika ... 31
4.1.3 Elsıdleges termelés ... 33
4.2 Patak epiliton fotoszintézise és elsıdleges termelése természetes és módosított patakszakaszokon ... 34
4.2.1 Biomassza ... 34
4.2.2 P-I karakterisztika ... 34
4.2.3 Elsıdleges termelés ... 37
4.3 Bevonat eltávolíthatósága süttıi mészkırıl ... 39
4.3.1 A korrekciós tényezı (K) ... 40
4.4 Aphanizomenon flos-aquae dominanciájú téli fitoplankton fotoszintézise és elsıdleges termelése ... 41
4.4.1 Biomassza ... 41
4.4.2 P-I karakterisztika ... 41
2
4.4.3 Fotoszintézis hımérsékletfüggése ... 43
4.4.4 Izotópos és LDO módszer összevetése ... 43
4.4.5 Primer produkció ... 43
5 Diszkusszió ... 44
5.1 A Balaton epipszammon közösségeinek fotoszintézise és elsıdleges termelése ... 44
5.2 A Torna-patak epiliton közösségeinek fotoszintézise és elsıdleges termelése ... 48
5.3 Aphanizomenon flos-aquae fotoszintézise és elsıdleges termelése a Stechlin-tóban 54 6 Összefoglalás ... 57
7 Köszönetnyilvánítás ... 58
8 Irodalomjegyzék ... 59
9 Tudományos tevékenység adatai ... 80
10 Eredmények tézisszerő összefoglalása ... 81
11 Results of the studies ... 83
12 Függelék ... 85
3
RÖVIDÍTÉSEK JEGYZÉKE CDOM: colored dissolved organic matter
DCM: Deep chlorophyll maximum
Ik: fényadaptációs paraméter (µmol m-2s-1)
PAR: Photosynthetically Active Radiation, 400-700 nm
PBmax: biomasszára vonatkoztatott maximális fotoszintézis, pl.mg C mg kl-a-1 h-1 P-I: a fotoszintézis fényintenzitás függése
Pmax: maximális fotoszintézis mértéke. Mértékegységét leggyakrabban oxigéntermelés, illetve szénfelvételben adják meg térfogat/területegységre vonatkoztatva egységi idı alatt, pl. mg C m-2h-1
α: fényhasznosítási tényezı, a P-I görbe kezdeti meredeksége, α =Pmax/Ik, pl. mg C m-2h-1 (µmol m-2s-1)-1
β: fénygátlási paraméter, β=Ps/Is (=telítési/szaturációs fényintenzitáshoz tartozó fotoszintézis/telítési fényintenzitás), pl. mg C m-2h-1 (µmol m-2s-1)-1
4 KIVONAT
Bentikus és planktonikus algaközösségek fotoszintézisének karakterisztikái
A doktori értekezésben bemutatott kutatások célkitőzése az volt, hogy megállapítsa a különbözı algaközösségek P-I karakterisztikáját, és becsülje a vízi rendszerek elsıdleges termelésében betöltött szerepüket. Erre vonatkozóan tudományos eredményei a következık:
(1) A szerzı mérte a balatoni epipszammon és epipelon fotoszintézisét laboratóriumi körülmények között, majd az eredmények segítségével becsültük a közösségek jelentıségét a tavi elsıdleges termelésben. Korábbi évek eredményeihez hasonlóan megállapította, hogy nyári idıszakban a Balaton üledékének algaközösségei a sekély viző részeken a Balaton egész területén jóval produktívabbak, mint a fitoplankton közösségek. A kapott eredmények alapján megállapítható a tó mind a négy medencéjére vonatkozóan, hogy nyári idıszakban az epipszammon jelentıs produkciója miatt a sekély, pár száz méter széles déli parti sáv a tó legproduktívabb területe.
(2) A szerzı Magyarországon elıször mérte egy patak epiliton közösségeinek fotoszintézisét. A laboratóriumi mérések eredményeibıl becsülte az autochton primer produkciót módosított és természetes patakszakaszok esetében. Megállapította, hogy a kísérı vegetáció hatással van az epiliton biomasszájára és ezáltal termelésére, emiatt a jó fényellátottságú, módosított szakaszok elsıdleges termelése jóval nagyobb, mint az árnyékos, természetes szakaszoké.
(3) A szerzı bizonyította a nemzetközileg elfogadott és ajánlott kefés bevonat-eltávolítás biomassza mérésekben okozott hibáját és javaslatot tett ennek kiküszöbölésére. Korrekciós tényezıt határozott meg a Torna-patak devecseri mintavételi helyére, ami által a Limnológia Intézeti Tanszék korábbi biomassza méréseinek eredményei korrigálhatóvá váltak.
(4) Meghatározta a Stechlin-tóban (Németország) jég alatt akkumulálódott Aphanizomenon flos-aquae kékalga fotoszintetikus paramétereit eltérı fényintenzitás és hımérséklet értékek esetében. Mérései alapján megállapította, hogy a közösség a felszíni vízrétegben jelentıs tömegprodukcióra képes a hımérsékleti toleranciahatárán is.
5 ABSTRACT
Photosynthetic characteristics of benthic and planktonic algal communities
The aims of the research discussed in this PhD thesis were to measure the photosynthetic characteristics of different algal communities, and to estimate the role of these communities in the aquatic primary production. The new results are the following:
(1) The photosynthesis of the epipsammon and epipelon of Lake Balaton was measured in laboratory. The precentage contribution of the epipsammon in the whole lake primary production was estimated. In the summer period of 2006, contribution of the epipsammic communities, especially in the shallow regions, was higher to total primary production than that of the phytoplankton of Lake Balaton. According to these and the former investigations, the southern shallow part is the most productive region of Lake Balaton, especially in summers.
(2) The photosynthetic activity of stream epilithon communities was measured first time in Hungary. The autochtonous production was estimated for the modified and for the natural stream sections in the laboratory measurements. The riparian vegetation affected the development of biomass and the primary production of the epilithon. The autochtonous primer production was substantially higher in the modified sections without riparian vegetation than at the natural sections with closed canopy.
(3) By brushing, algae cannot be efficiently removed from rough surfaces, therefore the brushing method can result in underestimating of chlorophyll-a content of benthic communities. The correction parameter (K) was determined from the proportion of the whole chlorophyll-a / brushed chlorophyll-a content. With the obtained K values the previous chlorophyll-a data from the same sampling station (Torna-stream) could be corrected.
(4) Photosynthesis of Aphanizomenon flos-aquae accumulated under ice at different light intensities and at different temperatures were measured. Accorrding to the measurements A. flos-aquae could reach high production in the upper 0-2 m water layer under the ice cover in the oligo-mesotrophic Lake Stechlin, Germany.
6
ZUSAMMENFASSUNG
Charakterisierung der Photosynthese benthischer und planktischer Algengemeinschaften
Das Ziel der Dissertation war es, die Abhängigkeit der Photosynthese verschiedener Algengemeinschaften vom Licht zu charakterisieren und einzuschätzen, wie wichtig diese Gemeinschaften für die Primärproduktion der Oberflächenwässer sind. Die neuen Ergebnisse für die Wissenschaft sind folgende:
(1) Die Photosynthese des Epipsammons und des Epipelons des Balatonsees wurde im Laboratorium gemessen. Der Anteil des Epipsammons an der Gesamtprimärproduktion des Sees wurde ermittelt. In der Sommerperiode 2006 war der Anteil der epipsammischen Gemeinschaften an der Gesamtprimärproduktion, speziell in den flachen Regionen, höher als der des Phytoplanktons im Balatonsee. Aufgrund unserer Ergebnisse und früherer Untersuchungen, erwies sich der südliche flache Teil des Balatonsees als der produktivste, speziell im Sommer.
(2) Zum ersten Mal in Ungarn, wurde die Photosynthese des Epilitons eines Baches gemessen. Die autochthone Primärproduktion in modifizierten und natürlichen Bachstrecken wurde unter Laborbedingungen gemessen. Die begleitende Fliessgewässervegetation hatte Einfluss auf die Biomasse und Produktion des Epilitons.
Die Primärproduktion in der modifizierten Strecke mit gutem Lichtdargebot war wesentlich höher, als in der natürlichen, beschatteten Strecke.
(3) Durch Abbürsten können die Algen nicht effektiv von rauhen Oberflächen entfernt werden. Deshalb führt diese Methode zu einer Unterbewertung des Chlorophyll-a-Gehaltes benthischer Gemeinschaften. Der Korrektur-Koeffizient (K) wurde aus dem Verhältnis von Gesamtchlorophyll-a zu abgebürstetem Chlorophyll-a ermittelt. Mit Hilfe des Korrektur-Koeffizienten wurden die ursprünglichen Daten der Messstation (Torna-Bach) korrigiert.
(4) Die photosynthetischen Parameter der sich unter dem Eis des Stechlinsees (Deutschland) akkumulierten Aphanizomenon flos-aquae-Blüte unter verschiedenen Licht- und Temperaturbedingungen wurde bestimmt. Die Gemeinschaft konnte eine hohe Produktion in der obersten Wasserschicht (0-2 m) unter Eis erreichen.
7
1 Irodalmi áttekintés: édesvízi algaközösségek jelent ı sége, m ő ködése, vizsgálatuk
Az algák a vizes élıhelyek legfontosabb fotoautotróf szervezetei. A vízben élı algákat a benépesített élıhely alapján különbözı csoportokba lehet sorolni. A fitoplankton a vízoszlop azon élılénycsoportját alkotja, mely nem képes a víz áramlatai ellenében mozogni, az áramlatokkal sodródik (Wetzel 2001). A fitoplankton jelentısége fıleg állóvizekben és olyan folyókban nagy, ahol az áramlás sebessége viszonylag alacsony.
Bentosz azon közösségek összessége, melyek valamilyen módon a vizes élıhely aljzatához kötıdnek, a szilárd-folyadék határfelületen élnek (Wetzel 2001). A perifiton olyan közösség, mely különbözı vízbe merült, a fotikus zónában található felületekhez rögzülve nı1, mikroszkopikus algák, baktériumok és gombák alkotják (Stevenson 1996a; Wetzel 2001). A felület (szubsztrát) típusa alapján a perifitont további csoportokba osztható: (a) epipelon – finom, szerves üledéken növı algabevonat; (b) epiliton – sziklán vagy kövön növı algabevonat; (c) epifiton – makrofiton szervezeteken növı algabevonat; (d) epizoon – állati szervezetek alkalmas felületén növı algabevonat; (e) epipszammon – homokos aljzaton növı, illetve benne mozgó algák csoportja (Wetzel 2001; Azim et al. 2005).
Gyorsabb folyású vizekben a perifiton különbözı csoportjai a meghatározóak a fitoplankton közösségekkel szemben (Wetzel 2001).
Az algák az élıvilág egyik legdiverzebb csoportját képezik, hatalmas szereppel bírnak a vízi rendszerekben (Stevenson 1996a). Jó ökológiai tesztszervezetek, s mivel jól definiálható ökoszisztémákban élnek, fajszámuk nagy, többségük relatíve könnyen határozható, rövid a generációs idejük, alkalmasak a különféle hipotézisek tesztelésére igen rövid idı leforgása alatt is.
1.1 Bentikus algaegyüttesek és szabályozó tényezıi
A különbözı bevonatok algaközösségei számos kutatás alapjául szolgálnak. Bizonyos esetekben kiemelkedı jelentıségük van a vizes élıhelyek elsıdleges termelésében, mind álló-, mind pedig folyóvizek esetében. Minshall (1978) hívta fel elsık közt a figyelmet arra, hogy számos patakban a szerves anyag elsıdleges forrása nem a terresztris vegetáció levélhullásából ered, hanem a patakban rögzülten élı algák autochton szerves anyag termelésébıl. A bevonatlakó algák szerves anyag termelésben játszott nagy szerepét azóta további kutatások is megerısítették (pl. Vannote et al. 1980; Busch & Fischer 1981, Ostrofsky et al. 1998). Nemcsak elsıdleges termelıkként, hanem tápanyagforrásként is nagy szerepük van a vízi rendszerek tápanyagforgalmában. A makrogerinctelen szervezetek elıszeretettel fogyasztják ıket (pl. Mayer & Likens 1987; Hansson 1992;
Lamberti 1996; Munoz 2001), sıt kísérletben igazolták, hogy elınyben is részesítik az élıbevonatot pl. a patakba hulló levelekkel szemben (Webster 1983).
A bevonatalgák a víz-szilárd felület határán élnek, biokémiai folyamatok során kémiai anyagok átalakítását végzik (Lock et al. 1984). Alapvetı szerepük van a különbözı biogeokémiai körfolyamatokban és azok dinamikájában (Amblard et al. 1990; Schorer &
Esiele 1997). A szervetlen foszfor- és nitrogénformák hasznosítása során akár azok erıs napi változását is elıidézhetik (Triska et al. 1989). Az üledék-víz határon élı algák fontos nyelıi a tápanyagoknak, csökkentik azok mennyiségét a vízoszlopban (Wetzel 1996, Battin
1 Gyakran emlegetett fogalmak még a rögzült szervezetekkel kapcsolatban a német eredető „Aufwuchs”.
Ezalatt a fogalom alatt azonban már nem csak a rögzült növényi szervezeteket értjük, hanem minden olyan szervezetet, ami az alámerült szubsztrát felületéhez kötıdik, vagy azon mozog (baktériumok, gombák, algák, protozoák, különbözı állati szervezetek), de nem hatol a szubsztrát anyagába (Wetzel 2001). A másik fogalom a biofilm, melynek használata fıleg mérnöki területeken elterjedt (Wetzel 2001).
8
et al. 2003), ezáltal a víz öntisztulásában is nagy szerepet játszanak (Vymazal 1988;
Steinman & McIntire 1990).
Alacsony nitrogéntartalmú élıhelyeken is magas primer produkciót érhetnek el, hiszen a kékalgák és azon diatomák, melyek kékalgákkal élnek endoszimbiózisban, képesek a légköri N2-t ammóniummá, majd aminosavakká átalakítani (Fairchild et al. 1985; Peterson
& Grimm 1992). A diatomák és a fonalas kékalgák azáltal, hogy megakadályozzák pl. a homokos aljzat elhordását, stabilizálják az aljzatot (Biggs 1996).
Mivel gyorsan, és összetetten reagálnak a környezeti paraméterek megváltozására, nagyon jó indikátorszervezetek (Lange-Bertalot 1979; Lowe & Pan 1996; European Union 2000), gyakran alkalmazzák ıket a biomonitorozásban és az ökológiai állapot becslésre (pl. Vis et al. 1998; Gómez & Licursi 2001; Gold et al. 2002; Foerster et al. 2004; Mages et al. 2004;
Soininen & Könönen 2004; Kovács et al. 2006; Kröpfl et al. 2006).
A kovaalgák fosszilizálódó váza kiemelt proxi a paleolimnológiai kutatásokban, melyek során a múlt eseményeit igyekeznek rekonstruálni (pl. Stoermer & Smol 1999).
A bentikus algabevonatok kialakulását, biokémiai folyamatait számos tényezı befolyásolhatja. Tápanyag tekintetében a bentikus algák bizonyos esetekben versenyelınyt éleznek a planktonikus közösségekkel szemben. Áramló vizekben az áramlás folyamatosan friss, tápanyagdúsabb vízzel veszi körül a bevonatot, és csökkenti az „elhasznált”
határréteget a bevonat körül, ami pl. csendes vízben az algák körül kialakul (Borchardt 1996; Stevenson 1996b). Azonban csendesebb vízben a bevonatban nagyobb mértékő a tápanyag limitáltság, mint a fitoplanktonban, mert a bevonatban jóval szorosabban elhelyezkedı, szomszédos vagy átlapoló sejtek miatt tápanyag-szegény régiók jöhetnek létre (Borchardt 1996; Stevenson 1996b; Tuchman 1996).
Amint egy szilárd felület víz alá kerül, szinte azonnal megindul felületén a kolonizáció:
bevonat képzıdik. Elıfordulhat, hogy valamely bolygatás olyan mértéket ölt, hogy szinte teljesen lemossa a bevonatot a felületrıl. A zavarás megszőntekor, alábbhagyásakor a kolonizáció újraindul. A kolonizáció során a sejtek betelepedésének mértékét a következık befolyásolják: az algapropagulum pool mérete és típusa; immigrációs jellegek, sejtméret, sejtmorfológia; szubsztrát típusa; áramlási sebesség; fényintenzitás (Osborne 1983;
Stevenson 1983; Bothwell et al. 1989; Stevenson & Peterson 1989; Peterson & Stevenson 1990; Stevenson & Peterson 1991). Amennyiben a kolonizáció „szőz” felületen indul meg, legelıször elektrosztatikus erık miatt egy szerves vegyületekbıl álló mátrix képzıdik (Van Dam et al. 2002). Ezt követıen a baktériumok hidrofób része és a szubsztrát közt alakul ki kapcsolat (Stevenson 1983; Carrias et al. 2002). A baktériumok szekréciós/exkréciós termékei (glycocalyx, mucopolysaccharidok) elıkészítik a felületet a további kolonizációhoz. Pár nap múlva már a diatomák is képesek megtelepedni az elıkészített felületen. Az algák, baktériumok, gombák és protozoák a saját extracelluláris polimerjeikben (EPS) élnek beágyazva (Characklis & Marshall 1990). Az EPS-mátrix egy dinamikus rendszer, ami formálja és kitölti a teret a sejtek között, és felelıs a bevonat architektúrájáért és morfológiájáért (Lewandowski et al. 1994). Az algák betelepedése hasonló folyamat a szárazföldi élıhelyek szukcessziójához. A betelepülés gyorsaságát, a különbözı tapadási stratégiával rendelkezı formák egymásutániságát különbözı környezeti paraméterek befolyásolják. A szerves mátrixon legelıször a felületükkel rögzülı diatomák jelennek meg, hiszen ekkor még bıven van hely a számukra a teljes felülettel történı rögzüléshez. İket követik az apikálisan rögzülı fajok, s végül a fonalas zöldalgák (Peterson & Stevenson 1990; Tuchman 1996). A teljes felülettel rögzülı diatomák a nyírási stressznek és a legelésnek jobban ellenállnak, de az egyéb tapadási
9
stratégiával rendelkezı fajok könnyen túlnövik ıket, ezáltal hamar fény- és tápanyaglimitáltak lesznek (McCormick & Stevenson 1989).
Az algák betelepülését követıen megindul az exponenciális növekedés (1. ábra), aminek hossza a hımérséklettıl, a források elérhetıségétıl és a reprodukció típusától függ (Bothwell 1989). Olyan élıhelyen, ahol van elegendı fény, fıleg a tápanyag limitál (Borchardt 1996), míg árnyékos élıhelyen a fénynek nagyobb hatása van a tápanyagoknál (Hill & Knight 1988b).
1. ábra Bentikus algák kolonizációs görbéje. PB: biomassza csúcs; TPB a kolonizáció megindulásától a biomassza csúcsig eltelt idı (Biggs 1996)
A biomassza csúcs eléréséhez szükséges idı (TPB) nagyon változó lehet. Tápanyagdúsabb vizekben a TPB eléréséhez általában rövidebb idı kell, mert a növekedési ráták nagyobbak (Bothwell 1989). Az is elıfordulhat, hogy a tápanyagban szegényebb élıhelyen éri el hamarabb a maximális biomasszát a bevonat, ennek oka az, hogy a tápanyag limitáltság korábban bekövetkezik, ezt követıen pedig már nem növekedés, hanem degradáció tapasztalható (Biggs 1996). Hillebrand és Sommer (2000) 4-6 hétre, míg Azim és munkatársai (2002) 3 hétre becsülték a TPB elérését. A TPB nagysága ettıl is eltérı is lehet, értékét fıleg a tápanyagterhelés befolyásolja (Bothwell 1989). Több rendelkezésre álló tápanyag esetében nagyobb esély van arra, hogy a bevonat alsóbb rétegeibe is jut belıle, lehetıvé téve vastagabb bevonat kialakulását. A maximális biomassza nagyságára még a víz áramlása is hatással lehet (Horner et al. 1990).
Vannak olyan tényezık, melyek közvetlenül befolyásolják a bevonat növekedését és csökkenését. Ezek fıként a vízgyőjtık olyan jellemzıit tükrözik, mint pl. földhasználat, vegetáció. Ezeket jobban megvizsgálva képet kapunk azon tényezıkrıl, melyek szintén hatással vannak a bevonat mennyiségére és mőködésére, mint pl. a geológiai jellemzık, klíma, emberi hatások (Biggs 1990; Biggs et al. 1990) (2. ábra).
10
2. ábra Az ábra azt mutatja, hogy a táj jellege hogyan befolyásolhatja egy patak azon alapvetı fizikai paramétereit, amik irányítják a bevonatalgák számára fontos biológiai folyamatokat és kölcsönhatásokat. Az
erıs kölcsönhatásokat sima nyíl, míg a gyengébbeket szaggatott nyíl jelöli (Biggs 1990 alapján)
A növekedést legfıképpen a források szabályozzák, mint a tápanyagok (az 2. ábrán a Tájhasználat kategóriába tartozik) és a fény (Vízminıség) mennyisége. A csökkenésért felelıs ellentétes folyamatokat szabályozó faktorok közé soroljuk a zavarásokat, a szubsztrát instabilitását, az erıs áramlást, az áradásokat, a lebegı anyagok általi dörzsölést (ezek mind a hidrológiai jellegő tényezık), illetve a legelést (makrogerinctelenek, halak).
Ezen faktoroknak a különbözı kombinációi különbözı folyamatokat erısítenek, és különbözı összetételő közösségek kialakulásához vezetnek. Alacsony és közepes áradási gyakoriság és kismértékő legelés mellett a növekedési folyamatok dominálnak. Ilyenkor - amennyiben van megfelelı mennyiségő tápanyag - fıleg a nyeles diatomák és a fonalas zöldalgák lesznek jellemzık a bevonatban. Azonban ha a források nem elegendıek, inkább fonalas kékalgákkal és kevesebb diatomával találkozunk. Közepes és gyakori zavarás és erıs legelési nyomás hatására csökkenés figyelhetı meg a bevonatban. Ezt a lassan növı, erısen rögzülı diatomák, (pl. Cocconeis placentula, Achnanthidium minutissimum), illetve a gyors kolonizációra képes fajok (Fragilaria vaucherie, Cymbella sp.) jelzik (Biggs 1996). Kevéssé vagy egyáltalán nem árnyékolt patakokban az áradásoknak jóval nagyobb szabályozó szerep jut, és nemcsak közvetlen hatása van. Az áradások során az alga propagulumok száma is megnövekszik (Uehlinger 1991), befolyásolja a tápanyagok koncentrációját (pl. Davies-Colley 1990; Humphrey & Stevenson 1992), csökkenti a víz átlátszóságát (Davies-Colley 1990; Hill 1996), módosíthatja a patak geomorfológiáját, az áramlási viszonyokat és a szubsztrátok méretét, eloszlását is (Jowett & Duncan 1990), illetve hatással lehet a legelı szervezetek denzitására (pl. Scrimgeour & Winterbourn 1989;
Quinn & Hickey 1990; Opsahl et al. 2003).
Vannak áradások, melyeket nem követ teljes biomassza vesztés, mert pl. az áradás gyenge, az áradás elıtti biomassza alacsony, vagy a rögzült fajok ellenállók a nyírási stresszel szemben (Biggs & Close 1989; Biggs & Thomsen 1995; Bergey & Resh 2006). Alacsony vagy közepes intenzitású áradásokat követıen a biomassza csúcs újbóli eléréséhez rövidebb idı is elég, mert az alga propagulumok abundanciája nagy. Ilyenkor a bevonat gyorsabban regenerálódik, csak a szukcesszióban került visszább. Ilyen esetekben akár öt
11
nap elteltével már diverz közösséggel találkozhatunk (Fisher et al. 1982), illetve a TPB eléréséhez elegendı lehet két hét (Stevenson 1990). Vannak olyan áradások, melyek után nagyon lassan alakul ki újra a biomassza csúcs, a folyamat akár több hétig is eltarthat.
Ennek egyik oka lehet a fajok alacsony növekedési rátája. Az is elıfordulhat, hogy ki sem alakulhat a biomassza csúcs, egy újabb áradás megint elmossa a friss bevonatot. A nagyon gyakori áradás nemcsak a bevonat mennyiségére, hanem annak összetételére is hatással lehet. Biggs és munkatársai (1998) azt tapasztalták, hogy a gyakori áradások miatt a diverzitás is csökkent a bevonatban, összevetve azokkal az évekkel, mikor nem volt olyan gyakori a bolygatás. Belátható tehát, hogy a különbözı felületeken kialakult bevonat folyamatosan változik, növekedési és degradációs szakaszok követik egymást.
Hosszabb idıskálán szemlélve a folyamatot, kisvízfolyások esetében három fı típust lehet megkülönböztetni a bevonat biomasszáját tekintve.
(1) Relatíve állandó, alacsony biomassza: állandó zavarások hatása miatt alakul ki, amik még a tápanyag hatásánál is erısebbek (Biggs 1995). Ilyen zavarások lehetnek a gyakori vagy váratlan áradások (Biggs 1995; Griffith & Perry 1995), illetve a folyamatosan mozgó üledék (iszap, homok). Amint e tényezık megszőnnek, a biomassza növekedésnek indul. Ha egy folyamatosan mozgó aljzaton izolálunk egy részt, akár 15x magasabb klorofill-a értéket is mérhetünk (Biggs & Shand 1987).
(2) Ritka fizikai zavarás ellenére alacsony biomassza: pl. ha sok makrogerinctelen legelı jelenik meg idıszakosan, és a legelés hatására alacsony biomassza marad (pl. Murphy 1984; Hill & Knight 1988a; Rosemond et al. 1993; Hillebrand &
Maria Kahlert 2001). Ilyen bevonatoknál a legeléssel szemben rezisztens fajok dominálnak egész évben (Biggs & Gerbeaux 1993; Rosemond 1994).
(3) Idıszakosan képzıdı, majd eltőnı bevonat: idıszakos változások, pl. közepes gyakoriságú áradások, vagy szezonális áradások, esetleg a fény szezonális változása, ahol sem a legelık sem az áradás nem játszik akkora szerepet (Hill &
Knight 1988a; Hill & Harvey 1990). A zavarást követıen 4-10 hétig tartó stabil hetek alatt fejlıdik ki a bevonat (Biggs & Close 1989; Uehlinger 1991) s mivel ilyenkor még nincsenek legelık, akár magas biomasszát is elérhet. A legelık megjelenésével az algák mennyisége csökken, azáltal van egy váltás az abiotikus és biotikus kontroll között. Majd egy újbóli áradást követıen kezdıdik a folyamat elölrıl.
A szubsztrát minısége is befolyásolja a rajta kialakuló biomassza mennyiségét és minıségét (pl. Tett et al. 1978; Biggs & Shand 1987; Kjeldsen 1996; Rier & King 1996;
Kjeldsen et al. 1998; Kahlert & Pettersson 2002; Bergey 2005; Flipo et al. 2007).
Állóvizek esetében például az üledéket fıleg diatomák népesítik be (Whitney & Darley 1983; Pinckney & Zingmark 1993). Azonos aljzatok esetében a tápanyag-fluxus az, ami a bevonat fajösszetételét meghatározza (Hagerthey & Kerfoot 2005). A köves aljzatok kémiája nem igazán befolyásoló tényezı az összetétel tekintetében (Bergey 2008), mert a kövek az algák számára inert felületnek tekinthetık, nem innen nyerik a tápanyagokat, hanem a vízbıl vagy a mátrixon belüli mikrobiális folyamatokból (Stevenson & Glover 1993). Természetesen vannak esetek, mikor a kı kémiája mégis hat a kialakuló közösség összetételére, pl. homokkıbıl a szilícium jóval könnyebben kioldódik, mint gránitból, ezáltal a diatomáknak kedvezıbb élıhelyet biztosít (Hiebert & Bennett 1992).
A víz kémiai összetétele hatással van a bevonatra, ami számos kutatás alapjául szolgál/szolgált. A legtöbben a tápanyagok hatását vizsgálták. Vannak, akik pozitív kapcsolatot találtak a víz tápanyag koncentrációja és a bevonat mennyisége között (pl.
12
Hillebrand & Kahlert 2001; Rosemond et al. 1993). Ismert azonban olyan eset is, hogy nem a tápanyagok, hanem más tényezı határozta meg a bevonat biomasszáját, mint pl. az alkalinitás (LaPerriere et al. 1989), vagy az áramlási sebesség (Welch et al. 1988).
Már korai kutatások bizonyították, hogy a perifiton közösségek fontosak lehetnek a szén fixálásában (pl. Wetzel 1964; Hargrave 1969; Bjöjk-Ramberg & Ännel 1985), s még relatív keskeny litorális zónájú tavaknál is jelentıs szerepet játszhatnak a szén körforgalmában (Loeb et al. 1983). Tölcsértorkolatok perifitonja akár a harmadát is adhatja a teljes produkciónak (Pinckney & Zingmarck 1993). Állóvizek esetében a fény, turbulencia, a víz kémiai összetétele illetve a legelés évszakos és napi skálán is befolyásolhatja a biomasszát és a biológiai folyamatokat (pl. Whitney & Darley 1983;
Munn et al. 1989; Biggs 1990; Blanchard & Montagna 1992; Lowe 1996; Putz 1997;
Pillsbury & Lowe 1999; Wellnitz & Ward 2000). Gruendling (1971) többváltozós korrelációs analízist végzett, mely alapján az általa vizsgált sekély tóban az epipelon biomasszája és elsıdleges termelése fıleg a fénytıl és a hımérséklettıl függött. Sok esetben találtak még összefüggést a hımérséklettel, ami nem meglepı, hiszen a hımérséklet az enzimek reakciósebességét, ezáltal pedig a biokémiai folyamatok sebességét befolyásolja (pl. Phinney & McIntire 1965; DeNicola 1996; Godwin & Carrick 2008).
S bár számos tényezı befolyásolhatja mind tavakban, mind pedig áramló vizekben a bevonat biomasszáját, összetételét és mőködését, mégis egyik kiemelkedı jelentıségő ezek közül a fény. A Napból érkezı sugárzás PAR tartománya (400-700 nm) az algák fı biológiai folyamatának, a fotoszintézisnek a motorja.
Patakok esetében a biomassza változása erdıs területeken gyakran összefügg a kísérı vegetáció változásaival (pl. Hill & Knight 1988b; Hill & Harvey 1990; Elósegui & Pozo 1998; Ostrofsky 1998; Figueroa-Nieves et al. 2006). A kísérı vegetáció hiányában akár ötször nagyobb biomassza értékek is mérhetık, mint a vegetáció miatt árnyékos helyeken (Hill & Knight 1988b). Állóvizek esetében az algabevonat szintén ott alakulhat csak ki, ahol az aljzat kap elég fényt. Mind álló-, mind pedig folyóvizekben a bevonathoz lejutó fény mennyiségét a vízben lebegı részek, a turbiditás nagymértékben befolyásolja (V.- Balogh et al. 2009), ezáltal hatásuk lehet a bentikus algák biomasszájára az elsıdleges termelésére egyaránt (pl. Davies-Colley et al. 1992). Fıleg állóvizek esetében a fény kulcstényezı a PAR-t hasznosító fitoplankton és a bentikus algák kompetitív interakciójában. A fitoplankton kedvezıbb helyzetben van e szempontból, mivel a bentikus algák felett helyezkedik el. Ezáltal a fitoplankton szabályozza annak a fénynek a mennyiségét és minıségét, ami eléri a bevonatot. A bentikus és a planktonikus algabiomassza között emiatt inverz kapcsolat áll fenn (Hansson 1988). Nem a fény az egyedüli szabályozó tényezı a tavak esetében sem. Amennyiben csak a fény lenne az egyedüli tényezı, ami befolyásolja a bevonat biomasszáját, akkor a fény exponenciális csökkenésével a biomasszának is exponenciálisan kellene csökkennie a mélység növekedésével. Ez nem így van. Stevenson és Stoermer (1981) azt tapasztalták, hogy a sekélyebb részeken (<9m) alacsonyabb volt a biomassza, mint a mélyebb helyeken. 9-15 méter mélység közt magasabb értékeket mértek, majd 23 méternél nagyobb mélységekben újra csökkenést tapasztaltak. A sekély részek alacsonyabb biomasszája ebben az esetben pedig nem a fényintenzitás miatt alakult ki, hanem a hullámzás hatása miatt.
Nemcsak a bevonat feletti vízréteg, hanem maga a bevonat is csökkenti a fény intenzitását, aminek eredményeképp a bevonat alsóbb rétegeibe kevesebb fény jut. A perifiton mátrix fénykioltását többen tanulmányozták (pl. Meulemans 1987; Jorgensen & Nelson 1988;
Dodds 1989, 1992). Homokos, iszapos aljzatban is történik fénykioltás, ami azt
13
eredményezi, hogy csak az üledék felsı pár mm-es rétege fotikus, itt található a bentikus algabiomassza nagy része (pl. Revsbech & Jorgensen 1983; Jorgensen & Des Marais 1986;
Revsbech et al. 1988; Nielsen et al. 1990; Dodds 1992; Pinckney & Zingmark 1993;
Barranguet 1998; Qu et al. 2004; Bányász et al. 2005).
1.2 Fitoplankton közösségek és szabályozó tényezıi
A nyílt tengerekben és mély tavakban az elsıdleges termelés szinte teljesen a pelagiális egysejtő vagy kolóniás mikroalgák alkotta fitoplankton fotoszintézisén alapszik, emiatt a mélyebb vizekben a fitoplankton közösségek a vízi tápanyaglánc alapjai. A fitoplankton mellett a bentikus algák csak olyan sekély helyeken tudnak jelentısen hozzájárulni az elsıdleges termeléshez, ahol van elég mennyiségő fény a fotoszintézisükhöz (Hill 1996).
Az utóbbi évtizedekben a globális felmelegedés miatt a világóceán fitoplanktonjának mennyisége csökkenı tendenciát mutat, egyes becslések szerint (Siegel & Franz 2010) 2010-ben 40%-kal csökkent a mennyisége 1950-hez képest.
A fitoplankton fajok mérete nagyon változó, általában a 0,5 µm-1 mm mérettartományba esik, kolóniák esetén meghaladhatja az 1 mm-t is. Három fı mérettartományt különböztetünk meg: pikoplankton (< 2 µm), nanoplankton (2-30 µm) és hálóplankton (>
30 µm) (Wetzel 2001; Lampert & Sommer 2007). A fitoplankton szervezetek maximális méretét egy adott élıhelyen a szél által keltett örvények mérete is befolyásolja. A tengerekben az utolsó örvény átmérıje is nagyságrendekkel meghaladja a sekély tavakban tapasztalhatókét, lehetıvé téve emiatt tengerekben a nagyobb mérető fajok megjelenését, aminek a fentin kívül fontos komponense a közeg és a fitoplankton fajlagos tömege közti kisebb különbség, valamint a mélyebb epilimnion is. Fotoszintetizáló szervezetek lévén a fitoplankton az eufotikus rétegben akkumulálódik. Önálló mozgásra nem, vagy egyes fajok csak igen csekély mértékben képesek, a víz áramlataival sodródnak. A víz keveredése segíti a fitoplankton szervezetek eufotikus rétegben maradását, de számos stratégiát kifejlesztettek a felsıbb rétegekben töltött idı meghosszabbítására. Egyes fajok olajcseppeket tartalmaznak, másikakban apró gázzal töltött vakuólumokat találunk, hogy csökkentsék a sőrőségüket (Wetzel 2001). Vannak fajok, melyek morfológiai adaptációval, alakrezisztenciájuk növelésének segítségével védekeznek a gyors kisüllyedés ellen (Padisák et al. 2003).
A fitoplankton biomasszáját és fajösszetételét szabályozó fontosabb tényezık a tápanyagok mennyisége és minısége, a fény, a hımérséklet és a heterotróf szervezetek aránya. A fény és a hımérséklet szezonális változása a mérsékelt övi tavakban a fitoplankton közösség fajösszetételének változását eredményezi. A mérsékelt övi tavakban tavasszal és nyáron a leggazdagabb a fitoplankton, diverzitása is ekkor a legmagasabb (Lampert & Sommer 2007). Tavasszal a növekvı fénymennyiség és hımérséklet hatására a fitoplankton szervezetek fotoszintézisének aktivitása megnövekszik, a kora tavaszi idıszakban a kisebb mérető, rövidebb generációs idejő diatomák biomassza csúcsa a jellemzı. Ezt követıen a zöldalgák, majd a nyári, késı nyári idıszakban a kékalgák válnak dominánssá a közösségekben. A fényért folyó kompetíció olyan fajok kialakulását eredményezte, melyek jól tőrik a fitoplankton önárnyékolását, képesek a csökkent fényintenzitás ellenére is aktívan fotoszintetizálni. Ezen fajok fényadaptációs paramétere alacsony, lehetıvé téve a fotoszintézist az eufotikus réteg mélyebb, kisebb fényintenzitású mélységeiben is. Ilyen faj pl. a Planktothrix rubescens, mely e tulajdonságának köszönhetıen a tápanyagdúsabb eufotikus metalimnionban, vagy felsı hipolimnionban DCM-et hozhat létre.
14
Patakok esetében a fitoplankton mennyisége elhanyagolható annak ellenére, hogy a tápanyagok ezt nem indokolnák. A patakokban vett fitoplankton mintákban talált algák legtöbbször epifiton és/vagy epiliton eredetőek (Piirsoo et al. 2007).
1.3 A fotoszintézis kutatások elméleti háttere
A fotoszintézis a bioszféra egyik legáltalánosabb és legalapvetıbb biokémiai folyamata, mely során a növények a fény energiát kémiai energiává alakítják, s közben szerves anyagot állítanak elı. A növények eme tulajdonságuk miatt tekinthetık elsıdleges termelıknek. Fotoszintézis nélkül nem alakulhatott volna ki a ma ismert szárazföldi élet. A légköri oxigén szintjének kialakításában nagy szerepe volt a cianobaktériumoknak, de az algák jelentısége azóta sem elhanyagolható. A földi légkör oxigénjének 40-50%-át ezek az élılények termelik, s bár legtöbbjük mikroszkopikus mérető, a földi összes elsıdleges termelésben jelentıs részesedéssel bírnak. A világóceán a Föld felszínének kb. 71%-át borítja s az óceánokban, tengerekben az algák a kizárólagos primer producensek. Bár a világtengerekben az algák a földi növények összes klorofill tartalmának csak 1-2%-át adják, a földi, éves összes elsıdleges termeléshez akár 30-60%-kal is hozzájárulhatnak (Jeffrey 1981; Berger et al. 1989; Falkowski 1994; Antoine et al. 1996).
Ahhoz, hogy a fotoszintézis kutatások eredményeibıl következtetéseket lehessen levonni, tisztában kell lenni a fotoszintézis molekuláris biológiai hátterével és a szabályozó környezeti tényezık hatásaival is. Továbbá vannak olyan alapvetı fogalmak, melyek elengedhetetlenek a fotoszintézis kutatások eredményeinek értelmezéséhez.
A fénynek jelentıs szerepe van a fotoszintézisben. A fotoszintézis és a fényintenzitás viszonyát az úgynevezett P-I (Photosynthesis-Irradiance) görbék szemléltetik a legjobban (3. ábra).
3. ábra P-I görbe jellemzı adatai. Zöld vonalak: fénygátolt fotoszintézis. Piros vonalak:
fényszaturált fotoszintézis. Pmax – maximális fotoszintézis; Ik – fényadaptációs paraméter; α – fényhasznosítási tényezı; β – fotoinhibíciós paraméter.
A fotoszintézis nem lineáris kapcsolatban áll a fényintenzitással. Alacsony fényintenzitáson a fotoszintézis lineárisan nı a fénnyel, és limitált, mert ilyenkor a fotonok száma kevesebb annál, mint amit a fotoszintetikus rendszer befogni képes. A P-I görbéknek errıl a kezdeti szakaszáról a fényhasznosítási tényezı (α) ad információt. A kezdeti szakasz után már nem lineáris a viszony a fényintenzitás és a fotoszintézis között.
Ez a szakasz a telítıdés vagy szaturáció. Itt a fotoszintézist a sötét szakasz limitálja a
15
ribulóz-1,5-biszfoszfát alacsonyabb aktivitása miatt (Rivkin 1990). A fényintenzitás további növelésénél vagy tartja az elért maximum értéket, vagy csökkenés mutatkozik a fellépı fénygátlás miatt (Hill 1996).
1.3.1 Fotoszintézis modellek
A kísérletes kutatásokkal egy idıben megjelentek azon a munkák is, melyek a gyakorlati mérések során kapott eredményeket matematikai modellek segítségével próbálták értékelni, magyarázni (pl. Steele 1962; Jassby & Platt 1976; Platt et al. 1980; Falkowski &
Wirick 1981; Eilers & Peeters 1988). Számos fotoszintézis modell létezik, emiatt ehelyütt csak néhányat soroltam fel azok közül, melyeket nemzetközileg széleskörően alkalmaznak.
További modellek segítségével pl. az in situ termelést is meg lehet határozni. Flipo és munkatársai (2004) perifitonra dolgozták ki a PROSE modellt, mely igen jól közelítette a terepi méréseket (Flipo et al. 2007). Behrenfeld és Falkowski (1997) klimatológiai változókat is beépítettek a modelljükbe, mellyel globális szinten számoltak fotoszintézist.
Létezik olyan empirikus modell, mely a klorofill-a mennyiségét és a terepen mért hımérsékletet használja fel a primer produkció becslésére (Morin et al. 1999). Elınye, hogy a legtöbb vizsgálatban ezeket a paramétereket rutinszerően mérik, és a modell kihasználja azt, hogy a termelés, mint a legtöbb biokémiai folyamat hımérsékletfüggı (Phinney & McIntire 1965; DeNicola 1996). Van de Bogert és munkatársai (2007) pedig egy olyan modellt alkottak, mely a vizsgált tó pelagiális régiójának a heterogenitását is figyelembe veszi az elsıdleges termelés becslésénél.
1.3.2 Árnyékadaptáltság
Árnyékadaptáltság tekintetében kevés kutatás és kevés adat áll rendelkezésre, annak ellenére, hogy tudjuk, egyes élıhelyeken extrém kicsi lehet a beesı fény intenzitása. Az árnyékadaptált mintákban általában az antenna pigmentek száma nı, a sötétreakció enzimjei pedig csökkennek, ezáltal az α értéke nagyobb, míg a Pmax és Ik értéke kisebb lesz.
Ezt támasztotta alá McIntire és Phinney (1965) kutatása is, melyben 40 és 110 µmol m-2 s-1 fényintenzitásokon növesztett bevonatok P-I karakterisztikáját vizsgálták. Az α értéke az árnyékadaptált mintákban magasabb volt, mint a fényen nevelt mintákban. Természetben az árnyékos helyek fotoszintézise nem haladja meg a napfényes helyek fotoszintézisét, annak ellenére sem, hogy az alacsony fényintenzitásokon nagyobb a fotoszintézis hatékonysága az árnyékadaptált közösségekben. Ennek oka, hogy az árnyékos helyeken legtöbbször nincs is akkora fényintenzitás, amin a Pmax kialakulhatna (Hill et al. 1995; Hill 1996).
1.3.3 Fotoinhibíció vagy fénygátlás
Elıfordul, hogy nagyobb fényintenzitásokon már nem növekszik tovább a fotoszintézis, hanem intenzitása csökkenést mutat. A rövid ideig tartó fotoszintézis vizsgálatokban bentikus algák esetében a P-I görbéknél akár már 400 µmol m-2 s-1-os fényintenzitáson is tapasztalták a jelenséget (pl. Hunding 1971; Boston & Hill 1991), de számos mérés van, amiben nem tapasztaltak fénygátlást (pl. Turner et al. 1983; Davies-Colley et al. 1992;
Dodds 1992). Ha árnyékos helyrıl vett mintákat teszünk ki magasabb fényintenzitásnak, gyakrabban tapasztalunk fénygátlást, mint a fényes helyrıl vett minták esetében. Hill és Boston (1991) csak erısen árnyékolt helyrıl vett mintáknál tapasztalt fénygátlást 1100 µmol m-2s-1 feletti fényintenzitásnál. A fényes helyen növı közösségekre a nagyobb fényintenzitás szelekciós nyomást jelent, a közösséget olyan fajok alkotják, melyek képesek kivédeni az erıs fény károsító hatásait. Ilyen védekezı mechanizmus lehet pl. a járulékos pigmentek számának növelése az antennarendszerben (Garcia-Pichel &
Castenholz 1991; Hill et al. 1995).
16
Ismert, hogy a bevonaton belül a fényviszonyok eltérıek lehetnek (Paul & Duthie 1989;
Dodds 1992; Pinckney & Zingmark 1993), s emiatt a különbözı fényellátottságú rétegek fotoszintézis karakterisztikája más és más lehet (Paul & Duthie 1989; Dodds 1992;
Pinckney & Zingmark 1993). A kísérletek során mérhetı a teljes bevonat fotoszintézise (pl. Howard-Williams & Vincent 1989; Davies-Colley et al. 1992; Pinckney & Zingmark 1993; Kolmakov et al. 2008), vagy akár rétegenként haladva a különbözı mélységeké külön-külön (pl. Pinckney & Zingmark 1993). A rétegenkénti mérés jóval idıigényesebb és összetettebb feladat, de hasznos, mert ennek segítségével megismerhetjük, hogy a közösség mekkora része aktív illetve inaktív a különbözı fényintenzitások esetén.
Belátható, hogy ha egy bizonyos fényintenzitás a legfelsı rétegben fénygátlást okoz, az alsóbb rétegekben a csökkent fényintenzitás miatt a közösség lehet, hogy még a szaturációt sem éri el (Paul & Duthie 1989; Dodds 1992). Elıfordul, hogy a laboratóriumi mérésekben nem mutatkozik fénygátlás a bevonatmintákban, de ez nem jelenti azt, hogy nem is létezik.
A legtöbb P-I módszer a fotikus zónába esı teljes bevonat fotoszintézisét méri, ami egy integrált eredményt ad a különbözı rétegekrıl. Ez lehet az oka, hogy a bevonat összességét tekintve nem mutatkozik fénygátlás a közösség P-I karakterisztikájában, hiszen míg a bevonat felsı rétege gátolt, addig az alatta lévı rétegek szaturációs vagy szubszaturációs fényintenzitáshoz jutnak. Amennyiben elég vastag a bevonat, akkor az alsó rétegek fotoszintézise kompenzálhatja a felsı rétegek fénygátlását (Sand-Jensen & Revsbech 1987;
Hill & Boston 1991).
A fénygátlás kialakulása idıfüggı is (Takahashi et al. 1971; Harris & Lott 1973; Marra 1978a; Falkowski 1984; Macedo et al. 2002). Ezt a fotoszintézis mérések tervezésénél figyelembe kell venni: az inkubációs idınek megfelelı hosszúságú idıintervallumot kell választani. A steady-state állapot elérését többen más idıpontra tették, de minimum egyórás inkubálás, de még jobb, ha 2 vagy több, amit javasolnak (Harris & Piccinin 1977;
Marra 1978a, 1978b; Lamberti et al. 1987; Macedo et al. 1998; Macedo et al. 2002).
A fotoinhibíció erısen függ a beesı fény hullámhosszától is, ami fıleg alacsony hullámhosszaknál jelentkezik (Vincent & Roy 1993).
1.3.4 Fényszaturáció
A szaturációs fényintenzitás általában az in situ fényintenzitás fölé esik, amibıl következik, hogy az in situ fotoszintézis erısen fénylimitált (Hill 1996). Vannak olyan esetek, mikor a P-I görbe nem éri el a telítési értéket (Kelly et al. 1974; Dodds 1992). Kelly és munkatársai (1974) szerint a telítés a fogyatkozó tápanyagok vagy egyéb limitáló tényezı miatt bekövetkezhet.
1.4 Nemzetközi bevonatalga fotoszintézis- és elsıdleges termelés kutatások
A bentikus algáknak is jelentıs szerepe lehet a vízi rendszerek szén- és tápanyagforgalmában (pl. Wetzel 1964; Mayer & Likens 1987; Hansson 1992; Lamberti 1996; Underwood & Kromkamp 1999; Munoz 2001; Wetzel 2001; Dodds 2003;
Liboriussen & Jeppesen 2003; Azim et al. 2005; Blindow et al. 2006; Van de Bogart et al.
2007; Poulíčková et al. 2008; Vadeboncoeur et al. 2001, 2003, 2006, 2008;). Ahhoz, hogy jobban megérthessük ezen algaközösségek mőködését, ökoszisztémákban betöltött szerepét, elengedhetetlenek a taxonómiai, társulásszerkezeti és fiziológiai vizsgálatok.
A fiziológiai kutatások története is több évtizedre nyúlik vissza, melyek egyik szegmensét alkotják a fotoszintézis kutatások. Már korán felismerték, hogy a bevonatlakó algák vizsgálata is fontos a vízi rendszerek tápanyagforgalmának teljes megértéséhez (Wetzel 1964; Hargrave 1969). Ennek ellenére az édesviző tavak litorális régiójában élı bevonatalgák szerepére kevés figyelmet fordítottak. Még kevesebb kutatás foglalkozott
17
folyóvizek bevonatalgáinak elsıdleges termelésével, de vannak korai munkák folyóvizek bevonatát illetıen is (pl. Odum 1956; McIntire & Phinney 1965). A tavak és folyóvizek bevonatának fiziológiai vizsgálata fıleg azon közösségekre irányult, melyek szilárd felületen, azaz kövön, illetve a makrofita vegetáción alakultak ki (pl. Wetzel 1964; Sumner
& McIntire 1982; McCormick & Stevenson 1991; Lowe 1996; Müller 2000; Hillebrand &
Kahlert 2001; Blindow et al. 2006; Liboriussen & Jeppesen 2006; O’Reilly 2006). Ez patakok, gyorsabb folyású vizek esetén kézenfekvı, hiszen ezekben az erısebb áramlások miatt a laza üledék folyamatos „vándorlása” miatt nem alakul ki epipelon és epipszammon közösség (Azim et al. 2005), vagy annak jelentısége csekélyebb a stabil felületen kialakult bevonaténál (Biggs & Shand 1987). Tavaknál azonban annak ellenére, hogy az epipelon és epipszammon biomasszájának és elsıdleges termelésének vizsgálatai bizonyították jelentıségüket, elenyészı azon kutatások száma, melyek ezekkel a közösségekkel foglalkoznak (Round 1960; Khondker & Dokulil 1988; Cyr 1998; Liboriussen & Jeppesen 2003; Migné et al. 2004; Vadeboncoeur et al. 2006; Poulíčková et al. 2008).
Az utóbbi 20-30 évben nıtt meg azon kutatások száma, melyek behatóbban tanulmányozzák a bevonatlakó algákat mind társulástani, mind pedig fiziológiai szempontból. A P-I kutatások fı célja a legtöbb esetben az, hogy megismerjék a közösség ökofiziológiai válaszát a fény megváltozására, továbbá becsüljék az in situ fotoszintézist.
A legtöbb tanulmány fıleg az elsı célból készül, kevesebben használják a kapott eredményeket in situ fotoszintézis becslésre (pl. Hill et al. 1995).
Az ökofiziológiai jellegő kutatások számának növekedésében jelentıs szerepet játszott a mérımőszerek fejlıdése, és kifejlesztett mérési módszerek sokfélesége. A lassabb, klasszikus analitikai módszereken alapuló méréseket (mint pl. a Winkler módszer; pl.
Hansmann et al. 1971) felváltották a gyorsabb mőszeres mérések (pl. Langdon 1984; Sand- Jensen & Revsbech 1987; Uehlinger & Naegeli 1998; Hanson et al. 2003). A kontrollált laboratóriumi méréseket pedig a kutatók által fejlesztett berendezések segítették (pl.
McIntire & Phinney 1965; Hansmann et al. 1971; Steinman et al. 1990; Tóth & Herodek 2009).
Patakok bevonatának tanulmányozása során is alapvetı kérdés, hogy in situ végezzük a méréseket, vagy laboratóriumban. A terepi méréseknél nehézséget okoz az, hogy a körülmények állandóan változnak. A fényintenzitást szinte lehetetlen kontrollálni, ezért kevés olyan in situ mérés van, ahol a fényintenzitás változik a többi tényezı konstans marad. Vannak olyan kísérletek, ahol nyitott rendszerben mérik in situ a termelést (LaPerriere et al. 1989; Parkhill & Gulliver 1998; Bott et al. 2006; Uehlinger 2006) általában a CO2–fluxus vagy O2 koncentrációk alapján. Ezek jóval bonyolultabbak és kevésbé megbízhatók, mivel itt számolni kell a gázkicserélıdéssel is (Bott et al. 1978). A zárt kamrák már alkalmazhatók in situ és laboratóriumban is (pl. McIntire et al. 1964;
Hickey 1988; Winterbourn et al. 1988; Munn & Brusven 2004; Uehlinger & Brock 2005;
Reid et al. 2006). Elınyük, hogy mind izotópos mind pedig oxigénes módszer alkalmazható bennük.
Magának a fotoszintetikus aktivitásnak a nyomon követése is többféle módon történhet.
Lehet mérni az O2 koncentráció változását (pl. Odum & Odum 1955; Thomas & O’Connell 1966; Hansmann et al. 1971; Lamberti et al. 1987; Dodds 1992; Dodds et al. 1999), CO2- fluxust (Migné et al. 2002; Davoult et al. 2004; Spilmont et al. 2006), 14C felvételt (pl.
Steemann-Nielsen 1952; Ostrofsky et al. 1998), 32P-PO4 felvételt (Riley 1956), H218
O felvételt (Grande et al. 1989), fluoreszcenciát (Gorbunov et al. 2000; Corno et al. 2008;
Kolmakov et al. 2008). Patak bevonatoknál a legtöbb módszert fitoplankton módszerekbıl adaptálták.
18
1.5 Nemzetközi fitoplankton fotoszintézis és elsıdleges termelés kutatások
A XVIII. század második felében írták le elıször a fotoszintézis jelenségét. 1850-es évekig több kutató azon fáradozott, hogy minél többet megtudjon a fotoszintézis biokémiai hátterérıl, és hogy megoldja a növények ısi rejtélyét (Rabinowitch 1945). A XIX. század elején már laboratóriumi kísérleteket végeztek a szárazföldi növények fotoszintézisét illetıen, mely kiváló alapul szolgált az édesvízi és tengeri fotoszintézis kutatásokhoz. A XIX. század utolsó évtizedeiben, Európában és Észak-Amerikában a vízi elsıdleges termelés, mint környezeti kémiai folyamat megértése számos kutatót foglalkoztatott (Chambers 1912). A legtöbb kutatást édesvizekben végezték, ahol a teljes szén-dioxid tartalom vagy természetesen változott, vagy manipulálták. A kutatások kb. fele kis tavakban, míg a másik fele üveg palackokban mérte a szén-dioxid és/vagy az oxigén koncentráció változását (Barber & Hilting 2002). Ezek az úttörı kutatások egyértelmően bizonyították, hogy a növényi biomassza növekedése szükségszerően szén-dioxid felvétellel és oxigéntermeléssel jár.
1910-1940 közti idıszakban számos kutatás irányult olyan kvantitatív módszerek kifejlesztésére, mely lehetıvé tette a növényi fotoszintézis mértékének mérését. Két alapvetı módszert ismertek, az oxigén koncentráció mérését Winkler-módszerrel, illetve a szén-dioxid felvétel mérését a pH változás nyomon követésével. Mindegyik módszert folyamatosan továbbfejlesztették, hogy alkalmasak legyenek a különbözı közösségek fotoszintézisének és elsıdleges termelésének mérésére. Ebben az idıszakban a mérések fı célja az óceánok és különbözı tavak primer produkciójának becslése volt. Sötét és világos palack módszerrel nyomon tudták követni a különbözı vízmélységek oxigén termelését és légzését. A Winkler-módszer kevésbé volt érzékeny alacsony biomassza esetében, így nagyon hosszú inkubációs idıt kellett választani ahhoz, hogy a koncentrációváltozások mérhetıek legyenek. Ezen idıszak jeles képviselıje volt Riley, akinek analitikai gondolkodása sokat segített az oxigénes módszer fejlesztésében. Vele egy idıben jelentek meg Steemann-Nielsen munkái is, aki Riley-vel ellentétben inkább módszeres, újító kísérletes kutató volt. Rávilágított arra, hogy az eddig alkalmazott módszerek tökéletesen jók a termelés idıbeli változásának kimutatására, de nem megfelelıek az elsıdleges termeléssel kapcsolatos mennyiségi kérdések megválaszolásához.
1940-es években Steemann-Nielsennek nagyszerő lehetısége adódott, munkáját Koppenhágában, a nukleáris tudományok központjában folytathatta. Számos kutatás és azok eredményei segítségére voltak a 14C izotópos fotoszintézis mérés kifejlesztésében (Steemann-Nielsen 1952). Ezt követıen számos kutatásban alkalmazták, és alkalmazzák ma is a radiokarbon módszert fotoszintézis mérésére. Ezt követıen jelent meg a világon az elsı, különálló kötet, mely a vízi elsıdleges termeléssel foglalkozott (Steemann-Nielsen &
Cushing 1958). Az 1950-1970-es idıszakban az ismert elsıdleges termelés kutatók közül szinte Riley volt az egyedüli, aki motiváltságot érzett arra, hogy az oxigénes módszerrel mérjen.
A XX. század második felében a mérések túlnyomórészt radioizotópos módszerrel történtek, de egyre több kutatásban alkalmazták újra az oxigénes módszert. A mérések kivitelezését felgyorsította az elektrokémiai oxigénszondák megjelenése. Sajnos ezek még mindig kevésbé voltak érzékenyek, mint az 14C módszer, de segítségükkel a közösségek légzése is mérhetıvé vált (Wetzel & Likens 2000).
19
1.6 Hazai alga fotoszintézis és elsıdleges termelés kutatások
A nemzetközi irodalom igen gazdag a fotoszintézis, illetve elsıdleges termelés kutatása terén, de a magyarországi publikációk száma elenyészı. Hazánkban Böszörményi és munkatársai (1962) munkája tekinthetı úttörınek e téren, hiszen nemcsak az elsık közt mértek fotoszintézist, hanem ık voltak az elsık, akik 14C izotóppal mérték azt. A 1970-es években Herodek és Tamás (1976) közölt adatokat a balatoni fitoplankton termelésérıl, Herodek és Oláh (1973) pedig in situ mérték a balatoni fitoplankton és fitobentosz elsıdleges termelését. Ezt követıen az 1990-es évekig kevés olyan munka volt (Herodek et al. 1982; Vörös 1989), ami vízi közösségek fotoszintézisével foglalkozott, pedig a Balaton eutrofizálódása során megnıtt alga biomassza megfelelı „kísérleti alany” lett volna.
Szerencsére a foszforterhelés csökkentésével sikerült a folyamatot megfordítani, és a Balatonban megindult az oligotrofizálódás (Padisák et al. 2006a). A 90’-es évek közepétıl aztán újra megindultak hazánkban a fotoszintézis kutatások. Ezen kutatások fıleg állóvizek (Balaton, Fertı-tó, szikes tavak), illetve a Kis-Balaton (Vörös 1994; Vörös et al. 2003b) és a Duna (Vörös et al. 2000) algaközösségeire irányultak. A vizsgálatok jelentıs része csak a fitoplanktonra koncentrált (Vörös & V-Balogh 1997; Vörös et al. 2000; Pálffy & Vörös 2003; Somogyi & Vörös 2006, Somogyi et al. 2007; Ágyi et al. 2008; Vanyovszki et al.
2008; Vörös et al. 2008; Ágyi et al. 2009; Somogyi et al. 2009a; Somogyi et al. 2009b;
Somogyi et al. 2010), de megjelentek olyan tanulmányok is, melyekben további közösségeket is tanulmányoztak (Vörös 1994; Vörös et al. 2001, 2002, 2003a, 2004;
Kovács et al. 2005; Kovács et al. 2006; Üveges et al. 2007; Pados et al. 2009; Üveges et al.
2009; Vörös et al. 2010).
A felsorolt publikációk közül mindösszesen négy jelent meg nemzetközi folyóiratban, ami azt jelenti, hogy elenyészı számban folytak ugyan fotoszintézis kutatások hazánkban, de azok nemzetközi dokumentációja gyér.
Kiemelném azt, hogy - mint a vízfolyások algaközösségeinek fotoszintézisével kapcsolatos fenti felsorolásból is kitőnik - a kisvízfolyások primer produkciójáról egyetlen munka sem született még idáig hazánkban. Mindezeket összevetve megállapítható, hogy a nemzetközi fotoszintézis kutatásban Magyarország fehér foltként szerepel.
20
2 Problémafelvetés és célkit ő zés
Manapság a Víz Keretirányelvnek köszönhetıen egyre nagyobb figyelem irányult, irányul a különbözı algaközösségekre, ami fıleg a társulás összetételének megismerését célozza.
Elenyészı az ökofiziológiai, ezen belül is a fotoszintézis kutatások száma. Fotoautotróf élılények révén az algák a vízi rendszerek legfontosabb elsıdleges termelıi, így nem elég csak a különbözı közösségek fajösszetételét ismerni, vizsgálni kell azok mőködését is. A környezeti paraméterek megváltozása mind a fajösszetétel, mint pedig a fiziológiai sajátságok változásában is nyomon követhetı. Fontos annak ismerete, hogy a különbözı környezeti paraméterek (pl. hımérséklet, fényviszonyok) megváltozása hogyan befolyásolja az elsıdleges termelık mőködését, s ezáltal az egész vízi tápanyagforgalomban betöltött szerepüket.
A korábbi években történtek fitobentosz biomassza vizsgálatok a Balaton különbözı területein, azonban az elsı, egész tóra kiterjedı mennyiségi felmérésre klorofill-mérésre 2000-ben került sor (Vörös et al. 2001). Végeztek becsléseket arra vonatkozóan is, hogy a planktonikus és a bentikus algák milyen mértékben és arányban járulnak hozzá az elsıdleges produkcióhoz a Balaton három medencéjében (Vörös et al. 2002, 2003a, 2004), mivel errıl korábban nem volt semmilyen információ. Az elsıdleges produkció méréséhez a siófoki térségbıl (Vörös et al. 2002, 2003a), a Keszthelyi-medencébıl (Vörös et al. 2002, 2003a) és a Balatonszemesi-medencébıl (Vörös et al. 2004) győjtöttek mintákat. Az egész tóra kiterjedı felmérés ez idáig nem készült az elsıdleges termelést illetıen.
A balatoni bentikus algák produkciójának in vitro meghatározását eleinte fotoszintetron készülékben végezték (Vörös et al. 2002, 2003a, 2004) 14C felvétel (Steemann-Nielsen, 1952) mérésével. A készülék a helyszíni inkubáció hátrányait (idıigényesség, egyszerre csak egy helyszínen lehet mérést végezni, idıjárásfüggı) kiküszöbölte, mivel a méréseket szabályozott körülmények között, ismert mérési paraméterek beállításával lehetett elvégezni. A Vörös és munkatársai (2003a, 2004) által alkalmazott fotoszintetron maximális megvilágítása 400 µmol m-2s-1 volt. In situ fényintenzitás méréseink alapján, a déli parton (25-100 cm vízmélységnél) az üledékre esı fényintenzitás elérheti a közvetlen a vízfelszín alatt mérhetı (100%) fény intenzitásának 50-60 %-át, azaz 1000 µmol m-2s-1 nagyságot is. Mivel a fotoszintetronban a méréseknél csak 400 µmol m-2s-1 maximális fényintenzitás volt elérhetı, szükségessé vált egy olyan berendezés megalkotása, mely lehetıvé teszi a különbözı eredető minták fotoszintézisének mérését a természetben elıforduló legnagyobb fényintenzitás értékek közelében is.
Kisvízfolyások esetében az emberi beavatkozás fıleg a medermódosításokban szembetőnı.
Mivel hazánkban semmilyen korábbi munka nem vizsgálta kisvízfolyások, illetve azokon belül a módosított, illetve a természetes patakszakaszok primer produkcióját, így ebben a témában minden munka hiánypótlónak tekinthetı.
2009 és 2010 telén a németországi Stechlin-tó fitoplanktonjában vízvirágzást okozott az Aphanizomenon flos-aquae kékalga (Padisák et al. 2010). A jelenség szokatlan, mivel ez a faj a sekély eutróf vizeket kedveli (a Stechlin átlagosan 26 m mély, és oligo-mezotróf), valamint 20°C feletti hımérsékleten maximális a populációnövekedése (pl. Uehlinger 1981; Yamamoto 2009; Yamamoto & Nakahara 2009a). Az, hogy egyre magasabb szélességi körökön jelenik meg, ismert (Yamamoto 2009; Mehnert et al. 2010). Arról is van információ más tavak esetében, hogy télen nagy biomasszával képviseltetheti magát (Yamamoto 2009; Yamamoto & Nakahara 2009b), akár a jég alatt is (Tsukada et al. 2006).
Vannak laboratóriumi mérések a faj fotoszintézisét és növekedését illetıen (Uehlinger 1981; Gibson 1985; Robarts & Zohary 1987; Huisman et al. 1999; Butterwick et al. 2005),
21
de ezeket tenyészetekkel végezték, és a kísérletek célja a növekedés és a fotoszintézis optimumának megállapítása volt. Ezen kísérletek egyike sem közelítette a Stechlin-tóban mérhetı téli in situ hımérsékletet (< 1 °C), a legalacsonyabb alkalmazott hımérséklet 6°C volt, pedig egy jóval magasabb hımérséklet optimummal rendelkezı faj toleranciahatárainak megismerése elengedhetetlen ahhoz, hogy a téli megjelenését megérthessük.
A fentiek alapján a következıket tőztem ki célul:
1. Balaton üledékéhez kötıdı nyári algabevonat fotoszintézis karakterisztikájának és elsıdleges termelésének meghatározása laboratóriumban in situ mérhetı fényintenzitás értékeken. Ennek megvalósításához:
a. epipszammon/epipelon fotoszintézis fényfüggésének mérése laboratóriumban, fotoszintetikus paraméterek meghatározása;
b. tavi elsıdleges termeléshez való hozzájárulás becslése;
c. összefüggés megállapítása az epipszammon/epipelon elsıdleges termelése és a tó különbözı paraméterei (trofitás, fényklíma) között
2. Patak epiliton fotoszintézisének és elsıdleges termelésének meghatározása. Ennek érdekében:
a. mintavételi eljárás kidolgozása;
b. biomassza mérés megbízhatóságának növelése;
c. epiliton fotoszintézis fényfüggésének mérése laboratóriumban, fotoszintetikus paraméterek meghatározása;
d. módosított és természetes szakaszok elsıdleges termelésének összehasonlítása, eltérések okainak feltárása.
3. Stechlin-tóban (Németország) tapasztalt szokatlan téli Aphanizomenon flos-aquae vízvirágzás során begyőjtött fitoplankton minta fotoszintézis karakterisztikájának és elsıdleges termelésének meghatározása. Ennek érdekében:
a. minták fotoszintézis fény- és hımérséklet függésének mérése laboratóriumban, fotoszintetikus paraméterek meghatározása;
b. in situ elsıdleges termelés meghatározása.