DOKTORI (PhD) ÉRTEKEZÉS
NAGY VIKTOR
PANNON EGYETEM GEORGIKON KAR KESZTHELY
2012
PANNON EGYETEM GEORGIKON KAR
NÖVÉNYVÉDELMI INTÉZET
NÖVÉNYTERMESZTÉSI- ÉS KERTÉSZETI TUDOMÁNYOK DOKTORI ISKOLA
ISKOLAVEZETŐ:
Dr. habil. Gáborjányi Richard egyetemi tanár
TÉMAVEZETŐ:
DR. NÁDASYNÉ DR. IHÁROSI ERZSÉBET egyetemi docens
A SELYEMMÁLYVA (ABUTILON THEOPHRASTI MEDIC.) CSÍRÁZÁSBIOLÓGIÁJÁNAK ÉS KOMPETÍCIÓS
KÉPESSÉGÉNEK TANULMÁNYOZÁSA
Készítette:
NAGY VIKTOR
KESZTHELY
2012
A SELYEMMÁLYVA (ABUTILON THEOPHRASTI MEDIC.) CSÍRÁZÁSBIOLÓGIÁJÁNAK ÉS KOMPETÍCIÓS KÉPESSÉGÉNEK
TANULMÁNYOZÁSA
Értekezés doktori (PhD) fokozat elnyerése érdekében Írta:
NAGY VIKTOR
KÉSZÜLT A PANNON EGYETEM NÖVÉNYTERMESZTÉSI- ÉS KERTÉSZETI TUDOMÁNYOK DOKTORI ISKOLÁJA KERETÉBEN
Témavezető: Dr. Nádasyné Dr. Ihárosi Erzsébet
Elfogadásra javaslom (igen / nem) ………
aláírás A jelölt a doktori szigorlaton ...%-ot ért el,
Az értekezést bírálóként elfogadásra javaslom:
Bíráló neve: …... …... igen /nem
……….
(aláírás) Bíráló neve: …... …... igen /nem
……….
(aláírás)
A jelölt az értekezés nyilvános vitáján …...%-ot ért el.
Keszthely, 2012. ……….
a Bíráló Bizottság elnöke
A doktori (PhD) oklevél minősítése…...
………
Az EDHT elnöke
Tartalomjegyzék
1.Kivonatok ... 6
1.1. Magyar nyelvű kivonat ... 6
1.2. Abstract ... 8
1.3. Auszug ... 10
2. Bevezetés ... 12
3. Irodalmi áttekintés ... 13
3.1. A selyemmályva (Abutilon theophrasti Medicus) származása ... 13
3.2. Az Abutilon theophrasti elterjedése ... 14
3.3. Az Abutilon theophrasti jelentősége Magyarországon ... 15
3.4. Az Abutilon theophrasti nevezéktana és taxonómiája ... 16
3.5. Az Abutilon theophrasti morfológiája ... 20
3.6. Az Abutilon theophrasti csírázásbiológiai sajátosságainak bemutatása ... 22
3.6.1. A magnyugalom ... 22
3.6.2. A csírázásra ható tényezők ... 24
3.7. Magasabb rendű növények közötti kölcsönhatások ... 27
3.7.1. A kompetíció ... 27
3.7.2. Az allelopátia ... 29
3.8. Az Abutilon theophrasti fitokémiai karaktere ... 33
3.9. A biológiai védekezés lehetőségei a selyemmályva (Abutilon theophrasti) ellen ... 34
3.9.1. A biológiai védekezés lehetőségei kártevőkkel ... 34
3.9.2. A védekezés lehetőségei kórokozókkal ... 36
4. Anyag és módszer ... 38
4.1. Csírázásbiológiai vizsgálatok ... 38
4.1.1. A vetési mélység hatása a selyemmályva (Abutilon theophrasti) kelésére és növekedésére ... 38
4.1.2. Műtrágyák hatása a selyemmályva (Abutilon theophrasti) kelésére és növekedésére ... 40
4.2. A selyemmályva (Abutilon theophrasti) allelopatikus hatásának vizsgálata ... 42
4.2.1. Laboratóriumi allelopátia bioteszt vizsgálatok ... 42
4.2.2. Üvegházi allelopátia vizsgálatok ... 43
4.2.3. A selyemmályva (Abutilon theophrasti) fitokémiai karakterének vizsgálata ... 44
4.3. A selyemmályva (Abutilon theophrasti) kompetíciós képességének tanulmányozása . 45 4.4. A selyemmályva (Abutilon theophrasti) kártevőinek vizsgálata ... 48
5. Eredmények és értékelés ... 49
5.1. Csírázásbiológiai vizsgálatok értékelése ... 49
5.1.1. A vetési mélység vizsgálat eredményei ... 49
5.1.2. A műtrágyák kelésre és növekedésre gyakorolt hatásának értékelése ... 65
5.2. Az allelopátia vizsgálatok eredményei ... 79
5.2. Az allelopátia vizsgálatok eredményei ... 79
5.2.1. A laboratóriumi allelopátia bioteszt eredményei ... 79
5.2.2. Az üvegházi tenyészedényes allelopátia vizsgálat eredményei ... 88
5.2.3. A selyemmályva fitokémiai jellemzőinek azonosítására irányuló vizsgálat eredményei ... 100
5.3. A selyemmályva kompetíciós képességének vizsgálata petrezselyemben ... 105
5.3.1. A parcellánkénti összes gyökértermés alakulásának értékelése ... 105
5.3.2. Az első osztályú és ipari minőségű gyökértermés mennyiségi adatok értékelése 106 5.3.3. A kezelések hatása a petrezselyem gyökerek átlagos hosszára ... 108
5.3.4. A kezelések hatása a petrezselyem gyökértömegére ... 110
5.3.5. A kezelések hatása a petrezselyem föld feletti részeinek tömegére ... 111
5.3.6. A kezelések hatása a petrezselyem gyökerek tápanyagtartalmára ... 112
5.3.7. A kezelések hatása a petrezselyem hajtásainak tápanyagtartalmára ... 113
5.4. A selyemmályva kártevőinek vizsgálata ... 115
5.4.1. Helix pomatia Linnaeus 1758 – éticsiga ... 115
5.4.2. Trialeurodes vaporariorum (Westwood 1856) – üvegházi liszteske ... 115
5.4.3. Oxycarenus lavaterae (Fabricius 1787) – hársbodobács ... 115
5.4.4. Pyrrhocoris apterus (Linnaeus 1758) – verőköltő poloska ... 116
5.4.5. Rhopalus parumpunctatus Schilling 1829 – közönséges üvegszárnyú-poloska .. 118
5.4.6. Liorhyssus hyalinus (Fabricius 1794) ... 118
5.4.7. Mamestra brassicae (Linnaeus 1758) – káposzta bagolylepke ... 121
5.4.8. Helicoverpa armigera (Hübner 1808) – gyapottok bagolylepke ... 121
6. Következtetések ... 122
6.1. Csírázásbiológiai vizsgálatok ... 122
6.1.1. Vetési mélység vizsgálat ... 122
6.1.2. A műtrágyák hatása a kelésre és növekedésre ... 123
6.2. A selyemmályva allelopatikus hatásának vizsgálata ... 125
6.2.1. Laboratóriumi bioteszt vizsgálat ... 125
6.2.2. Üvegházi allelopátia vizsgálat ... 127
6.2.3. A selyemmályva fitokémiai karakterének vizsgálata ... 128
6.3. A selyemmályva kompetíciós képességének vizsgálata ... 130
6.4. A selyemmályva kártevőinek vizsgálata ... 131
7. Összefoglalás ... 135
8. Köszönetnyílvánítás ... 140
9. Irodalomjegyzék ... 141
10. Az értekezés tézisei ... 168
10.1. Magyar nyelvű tézispontok ... 168
10.2. Angol nyelvű tézispontok ... 170
1.Kivonatok
1.1. Magyar nyelvű kivonat
Hazánkban az elmúlt 50 évben néhány gyomfaj, például a selyemmályva (Abutilon theophrasti Medicus) robbanásszerű, 10-200-szoros borításnövekedést ért el az első országos gyomfelvételezéshez képest. A felszaporodásuk más körülményeket is figyelembe véve elsősorban klimatikus okokra vezethető vissza. Emelett a szaporodási stratégiájuk is segíti őket, hogy szántóterületeink nagy részén megtelepedjenek és folyamatosan károsítsanak. A negyedik országos gyomfelvételezés adatai szerint a selyemmályva a 24. helyet foglalta el a gyomnövények fontossági sorrendjében, majd a 2007-2008-ban végzett ötödik felvételezés idejére a 21. helyre lépett előre.
Az ellene való védekezés jelenleg szinte kizárólagosan a kémiai növényvédelemre alapul, de a környezetkímélőbb növényvédelem iránti igény erősödése a biológiai módszerek előtérbe kerülését vonhatja maga után.
A dolgozat célja éppen ezért az utóbbi években fontossá vált selyemmályva (Abutilon theophrasti Medic.) biológiájának és az ellene való biológiai védekezés lehetőségének tanulmányozása volt. A dolgozat tartalma témakörei szerint a következő:
- A vetési mélység és a műtrágyák selyemmályva csírázására és növekedésére gyakorolt hatásának a vizsgálata.
- A selyemmályva allelopatikus hatásának vizsgálata laboratóriumi és üvegházi biotesztekben. A selyemmályva fitokémiai karakterének tanulmányozása, a lehetséges allelokemikáliák meghatározása.
- A selyemmályva kompetíciós képességének tanulmányozása szabadföldi, kisparcellás, additív kísérletben.
- A selyemmályva kártevőinek felmérése.
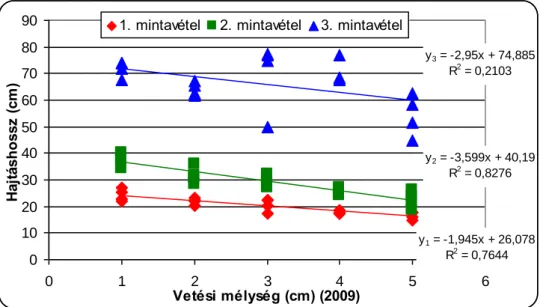
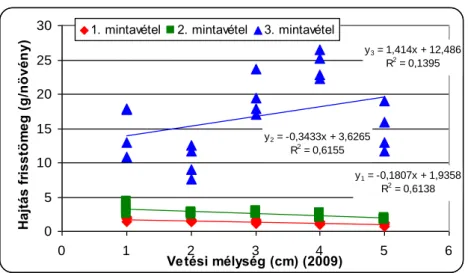
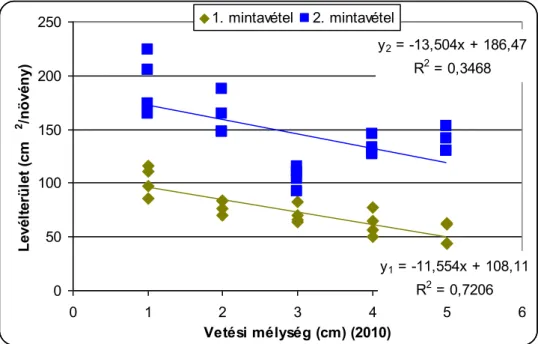
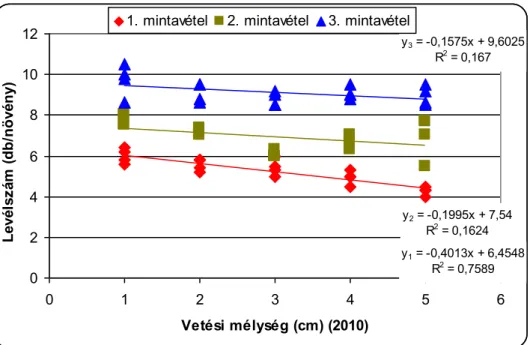
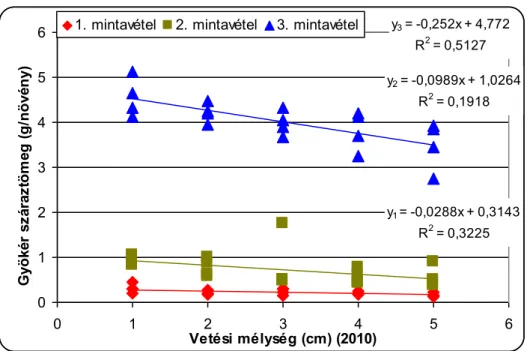
A csírázásbiológiai vizsgálat során megállapítottuk, hogy a selyemmályva számára a legkedvezőbb az 1-5cm-es csírázási mélység, de még 15cm mélyről is kicsírázik Ramann-féle barna erdőtalajon. Egy cm-es vetési mélység növekedés körülbelül 4,5%-os kelési % csökkenést okoz a selyemmályva magvaknál. A vetési mélység növelésével csökken a növények magassága, a levélterülete és a levelek száma.
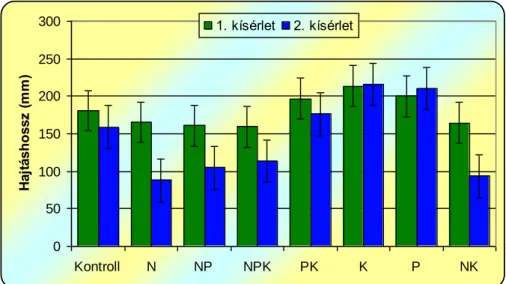
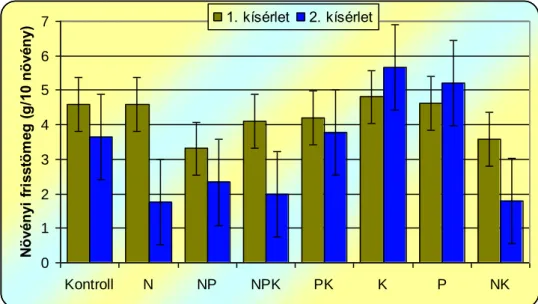
A műtrágyák selyemmályva kelésére és növekedésére gyakorolt hatásának vizsgálata során megállapítottuk, hogy a nitrogén többlet hatására csökken, a foszfor és kálium adagolásráa nő a selyemmályva kelési %-a, a hajtáshossz, illetve a hajtástömeg, valamint a levélterület a kontrollhoz képest.
Az allelopátia laboratóriumi vizsgálatok során a selyemmályva kivonatait alkalmazva gátló és serkentő hatásokat egyaránt tapasztaltunk. Megállapítottuk, hogy az allelopátikus hatás a selyemmályva esetén is koncentrációfüggő tulajdonság. A csapvizes kivonatok hatékonyabbnak bizonyultak a szerves oldószeres kivonatokhoz képest. Megállapítottuk továbbá, hogy a tesztnövények csírázási %-ának változása nem befolyásolja a hajtáshossz, illetve a gyökérhossz alakulását.
Tenyészedényes kísérletben a selyemmályva talajba kevert leveles szára magasabb koncentrációban erős gyökér- és hajtásnövekedés gátló hatású, kisebb koncentrációban kezdetben gátló, majd serkentő hatásúnak bizonyult. A talajba kevert gyökérmaradványok koncentrációtól függetlenül serkentőleg hatottak a tesztnövények fejlődésére.
SPME-GC/MS analízis vizsgálattal 23 illó, mono- és szeszkviterpenoid komponenst sikerült kimutatnunk, melyek közül 5 alkotórészt nem tudtunk azonosítani. A vékonyréteg- kromatográfiás vizsgálatban a selyemmályva magokból flavonoidot nem, de feltehetőleg 6 féle fenolkarbonsavat azonosítottunk, a mag tannintartalma csekély. A szárból rutint tudtunk kimutatni, mennyisége a 6-8 leveles fenológiai állapotban a legmagasabb.
A kompetíciós vizsgálat során megállapítottuk, hogy a selyemmályva egyedszámának növekedése a petrezselyem termését csökkenti. Az általa okozott kár azonban inkább minőségi, mint mennyiségi. A borításnövekedés hatására nőtt az elágazó gyökerek mennyisége, a gyökerek hossza és átlagos tömege. A kezelések nem befolyásolták a petrezselyem hajtástömegét, valamint a gyökerek és a hajtások tápanyagtartalmát.
A kártevőinek felmérése során 8 állatfajt írtunk le a selyemmályváról, a szakirodalmi adatokat 4 új kártevőfajjal bővítettük (Helix pomatia, Oxycarenus lavaterae, Rhopalus parumpunctatus, Mamestra brassicae). A saját eredményeket és az irodalmi adatokat figyelembe véve a selyemmályva biológiai úton történő visszaszorításában nagy szerepe lehet a Liorhyssus hyalinus üvegszárnyú poloskának.
1.2. Abstract
In the last 50 years some weed species, for example velvetleaf (Abutilon theophrasti Medicus) has reached a very high, 10-200-fold cover growth compared to the data of the first national weed survey in our country. Their accumulation is due to climatic causes, taking into account other circumstances as well. Their reproductive strategy, their biology also help them to settle and continuously cause damage on a large part of our fields. According to the data of the fourth national weed survey, velvetleaf was the 24th most important weed on the weed list, but it reached the 21st place by 2007-2008 when the fifth survey was done.
The protection against it is almost exclusively based on chemical plant protection at the moment, however since there is an increasing demand for more environmentally friendly plant protection, biological methods can come into view.
For that reason, the aim of the present thesis was to study velvetleaf (Abutilon theophrasti Medic.) which has become important in recent years, its biology and the possibility of biological control against it. The content and the topics of my thesis are the following:
- Examination of the effect of sowing depth and fertilizers on emergence and growth of velvetleaf.
- Examination of the allelopathic effect of velvetleaf in laboratory and greenhouse bioassays. Study of the phytochemical character of velvetleaf, determination of the possible allelochemicals.
- Study of the competitive ability of velvetleaf in open-field, small-parcel, additive experiments.
- Survey of the pests of velvetleaf.
Our most important results:
The increase in the sowing depth reduces emergenece percentage, shoot lenght, leaf area and the number of leaves of velvetleaf. It can emergence even from 15 centimetres depth.
From the macro-nutrient, nitrogen surplus hindered, phosphorus and potassium stimulated emergenece and growth.
During testing of the extractions prepared from different parts of velvetleaf, we experienced stimulating and interfering effects as well, aqueous extracts were proved to be the most effective. Alteration in the germination percentage of the test plants did not influence
We identified 23 volatile components in the leafy stem. We could detect rutoside in the stem. We could not find flavonoid in the seed, its tannin content is also low.
The parsley yield decreased because of the effect of the cover growth of velvetleaf, but the damage that was caused by velvetleaf was mainly qualitative. The amount of branching, waste roots increased.
We identified 8 pest species on velvetleaf, we broadened the literature data with for new pests [edible snail (Helix pomatia), mallow seed bug (Oxycarenus lavaterae), hyaline bug (Rhopalus parumpunctatus), cabbage moth (Mamestra brassicae)]. Liorhyssus hyalinus can play an important role in biological protection.
1.3. Auszug
In unserer Heimat wurde im Vergleich zur ersten Unkrautaufnahme ein explosionsartiges, 10-200faches Flächendeckungswachstum einiger Unkrautarten, zum Beispiel im Fall der Lindenblättrigen Schönmalve (Abutilon theophrasti Medic.)festgestellt.
Ihre Anhäufung ist unter Berücksichtigung anderer Faktoren in erster Linie auf klimatische Ursachen zurückzuführen. Darüber hinaus werden sie durch ihre Fortpflanzungsstrategie sowie ihre Biologie dabei unterstützt, sich auf einem großen Teil unserer Ackerfläche anzusiedeln und kontinuierlich Schäden anzurichten. Auf Grund der Daten von der vierten Landesunkrautaufnahme nahm die Lindenblättrige Schönmalve den 24. Platz in der Wichtigkeitsliste der Unkräuter ein und trat in der fünften in den Jahren 2007 und 2008 durchgeführten Aufnahme auf den Platz 21 vor.
Die Unkrautbekämpfung beruht zurzeit ausschließlich auf chemischem Pflanzenschutz aber der immer stärkere Anspruch auf einen umweltfreundlichen Pflanzenschutz kann dazu
Das Ziel meiner Arbeit bestand also darin, die Biologie der in den letzten Jahren wichtig gewordenen Lindenblättrigen Schönmalve und die Möglichkeiten ihrer Bekämpfung auf biologischem Wege zu studieren. Die Arbeit beinhaltet folgende Themenkreise:
- Prüfung der Keimungstiefe und Untersuchung der Wirkung der Kunstdünger auf Keimung und Wachstum von Lindenblättrigen Schönmalven.
- Untersuchung der allelopathischen Wirkung von Lindenblättrigen Schönmalven in Labor- und Grünhausbiotests. Untersuchung des phytochemischen Charakters von Lindenblättrigen Schönmalven und Festlegung der möglichen Allelochemikalien.
- Untersuchung der Konkurrenzfähigkeiten von Lindenblättrigen Schönmalven in additiven Kleinparzellen-Freilandexperimenten.
- Erhebung der Schädlinge von Lindenblättrigen Schönmalven.
Unsere wichtigsten Ergebnisse:
Eine Erhöhung der Keimungstiefe vermindert den Keimungsprozentsatz, die Sprosslänge, die Blätteroberfläche und die Anzahl der Blätter von Lindenblättrigen Schönmalven. Sogar aus einer Tiefe von 15cm keimt die Pflanze auf.
Von den Makronährelementen hat der Stickstoffüberschuss den Wachstum und die Keimung gebremst, während Phosphor und Kalium eine belebende Wirkung darauf hatten.
Bei dem Testen von Auszügen aus verschiedenen Teilen von Lindenblättrigen Schönmalven haben wir sowohl belebende als auch bremsende Wirkungen feststellen können, wasserhaltige Auszüge erwiesen sich als am erfolgreichsten. Eine Änderung der Keimungsprozentsätze der Testpflanzen hat ihre Spross- und Wurzellänge nicht beeinflusst.
23 ätherische Komponenten haben wir aus den Blätterstängeln identifiziert. Aus den Stängeln ließ sich Rutin nachweisen.
Wegen der größer gewordenen Flächendeckung von Lindenblättrigen Schönmalven sind die Erntemengen von Petersilie zurückgefallen aber der verursachte Schaden ist eher qualitativer Art. Die Menge der verzweigenden, minderwertigen Wurzeln wurde größer.
Von der Lindenblättrigen Schönmalve haben wir 8 Schädlinge identifiziert und die Fachliteraturdaten um 4 neue Schädlinge erweitert. [Weinbergschnecke (Helix pomatia), Feuerwanze (Oxycarenus lavaterae), Glasflügelwanze (Rhopalus parumpunctatus) und Kohleule (Mamestra brassicae)]. Im biologischen Schutz könnten Liorhyssus hyalinus eine große Rolle spielen.
2. Bevezetés
Hazánkban az elmúlt 50 évben néhány gyomfaj, például a selyemmályva (Abutilon theophrasti) robbanásszerű, 10-200-szoros borításnövekedést ért el az első országos gyomfelvételezéshez képest. Ezek a fajok többnyire a melegigényes gyomnövények csoportjából kerülnek, kerültek ki, a felszaporodásuk más körülményeket is figyelembe véve, elsősorban a klimatikus okokra vezethető vissza (Szőke 2001). A gazdaságilag jelentős gyomfajok evolúciójuk során sikeresen alkalmazkodtak a gyors növekedéshez és versengéshez, a hatékony reproduktív fázis biztosításához, valamint az agrotechnikai és kémiai védekezési eljárásokhoz (Hunyadi 2000). Ezen tényezők összessége vezetett ahhoz, hogy a negyedik országos gyomfelvételezés adatai szerint a selyemmályva a 24. helyet foglalta el, a 2007-2008-ban végzett ötödik felvételezés során pedig a 21. helyre ugrott előre (Novák et al. 2009). Rohamos terjedésének következményeként napjainkban a hazánkkal szomszédos országok mindegyikében gyomosít. A selyemmályva (Abutilon theophrasti) a Malvales rendbe, a mályvafélék (Malvaceae) családba, az Abutilon nemzetségbe tartozik. A faj őshazája valószínűleg Kína, ahol mint gyógy- és rostnövény termesztették. Innen terjedt el a Balkánon át Észak –Afrikán keresztül Európába, így hazánkba is (Czimber et al. 1994).
E növényfajnak a biológiai jellemzői rendkívül sokfélék. Ezek közül az egyik legfontosabb egy gyomnövény esetében, hogy milyen a gyomnövény szaporodási stratégiája, illetve a gyom versengése a kultúrnövénnyel az ökológiai faktorokért (Hunyadi 1988). A selyemmályva mindazon kultúrnövényeknek jelentős versenytársa, amelyek teret engednek kezdeti fejlődésének, ilyenek tehát a kapás kultúrák, de a kiritkult gabonavetésekben is megjelenhet (Kazinczi 2000). A selyemmályva erős kompetitív képességű növény. Több kísérlet is igazolta terméscsökkentő hatását (Czimber et al. 1994). A kompetíciós képessége mellett azonban a selyemmályva allelopatikus hatásról is szót kell ejteni. Hazánkban a 150 allelopatikus faj közül az egyik a selyemmályva (Szabó 1997).
A selyemmályva elleni védekezésnél a keményhéjúságot és az allelopátiát feltétlenül figyelembe kell venni. A védekezést komplex módon és több évre meg kell tervezni, és végre kell hajtani. A gyommentesítési programban több szempontot figyelembe kell venni: a talajművelést, a vetésváltást és a kémiai, valamint biológiai védekezést. A három védekezési mód kombinálásával, a gyomnövény biológiai sajátosságainak ismerete mellett lehetünk csak eredményesek a selyemmályva elleni küzdelemben (Nagy et al. 2009).
3. Irodalmi áttekintés
3.1. A selyemmályva (Abutilon theophrasti Medicus) származása
A szakirodalmi adatok alapján nincs egységes álláspont a selyemmályva (Abutilon theophrasti Medic.) származását illetőleg. Egyes szerzők a faj lehetséges őshazájaként a mai Kína területét jelölik meg (Vavilov 1951; Li 1970; Spencer 1984; Tutin et al. 1986; Warwick - Black 1988), melyek szerint a selyemmályva hasznosítása Kína szerte elterjedt volt és i.e.
2000 környékére, vagy ennél is régebbre nyúlik vissza. Ezeken a vidékeken és Tibetben mint gyógy-, illetve rostnövényt termesztették (Czimber et al. 1994; Kazinczi et al. 2001). Rostja általában 69% cellulózt és 19% lignint tartalmaz (Reddy – Yang 2008). Használati tárgyak sora készült rostjából, mint például: halászháló, táska, zsák, cipő. Ezen kívül magja 15-30%- ban emberi fogyasztásra alkalmas olajokat tartalmaz (Il’in 1949), melynek gyógyhatása ismert, Schwenkfeld (1600) a következő képen írja le: „Glutinosa et vulveraria est. Recentia consolidat et glutinat” (Köptető és sebkezelő drog. Vérzéscsillapító és sebgyógyulást elősegítő hatású). A festék- és illatanyagokon kívül legnagyobb mennyiségben a poliszacharidok jelentik a felhasználás alapját, akár nutritív értékű nyálka- heteropoliszacharidokról, akár zöldenergiát adó cellulózról (és lignocellulózról) van szó.
Kína több régiójában is termesztették a selyemmályva eltérő talaj- és klímaadottságokhoz alkalmazkodott különböző fajtáit (Kirby 1963). Más források azonban Indiát jelölik meg lehetséges géncentrumként (Shaw et al. 1974; Flint et al. 1983). Stegnik - Spencer (1988) sikertelen kísérletet folytattak az Abutilon theophrasti szülővonalainak és ezzel származásának kiderítésére. Azt feltételezték, hogy a hexaploid selyemmályva valószínűleg egy diploid és egy tertraploid ősből alakult ki, ezért számos, az Abutilon génuszba tartozó szubtrópusi és trópusi fajt vizsgáltak meg, de nem találtak származást igazoló rokoni kapcsolatot a fajok között. Vizsgálatuk szerint az Abutilon theophrasti az egyetlen mérsékelt égövi géncentrummal rendelkező Abutilon faj. Így a származásával kapcsolatos eltérő vélemények annyiban biztosan megegyeznek, hogy a faj semmiképpen sem trópusi vagy szubtrópusi eredetű (Haensel 2005).
3.2. Az Abutilon theophrasti elterjedése
A növényt egészen a XVIII. század közepéig rostnövényként termesztették Európa, Észak- Amerika és Ausztrália szántóföldjein, de a kender- és gyapottermesztés előretörésével jelentősége csökkent (Haensel 2005). Észak-Amerikában egészen az 1700’as évek közepéig potenciális rostnövény volt (Warwick - Black 1986), majd a termesztésből kikerülve veszélyes gyomnövénnyé vált az Egyesült Államok (U.S. Department of Agriculture 1970, Spencer 1984) és Kanada területének jelentős részén (Ontario Ministry of Agriculture and Food 1974). Az 1950’es évektől kezdve terjedése intenzívvé vált Kanada területén is és jelenleg 4 államban fertőzi a szántóföldeket (Lindsay 1953; Montgomery 1957; Rousseau 1968; Warwick - Black 1986). Ebben a régióban többek között a kukorica-, a szója- és a gyapottermesztésben okoz dollár milliókban mérhető károkat (Spencer 1984). Természetesen a faj dél felé is terjed és Közép-Amerika térségében is, például Mexikóban problémát jelent a mezőgazdasági termelésben (Dominquez Velanzuela et al. 2003).
A faj az őshazájából, Ázsiából valószínűleg a Balkánon és Észak-Afrikán át került be Dél- Kelet-Európába, Dél-Oroszországba, a Földközi-tenger nyugati vidékeire és hazánkba is (Tutin et al. 1986; Warwick - Black 1986; Kazinczi et al. 2001).
A XX. század elején fertőzött takarmánykukorica, szója és napraforgó szállítmányokkal a selyemmályva magja bekerült Skandinávia területére (Hylander 1970; Hansen 1972;
Suominen 1979). Nagy-Britanniába madáreleséggel hurcolták be (Hanson - Mason 1985).
Manapság szinte Európa összes országában megtalálható, mint például: Hollandiában (Mennema 1982), Németországban (Fukarek - Henker 1983; Viehweger - Dittrich 2004), Ausztriában (Hain 2003), Svájcban (Bohren et al. 2008), Lengyelországban (Kita et al. 2003), Csehországban (Jehlik et al. 1988), Horvátországban (Hulina 1995) és Bosznia- Hercegovinában (Dikic et al. 2008). Ezen kívül megtalálható még egész Ázsiában, Észak- Afrikában, Marokkóban (Tanji - Taleb 1997), Dél-Amerikában, Ausztráliában (Hanf 1983;
Holm et al. 1979) és Japánban is (Kurokawa et al. 2003; Watanabe 2007), mindenütt veszélyes gyomnövényként tartják számon.
Terjedésének a klimatikus tényezők szabhatnak határt, a júliusi 20OC-os izoterma északi elterjedésének határa (Jäger 1991). A klíma változásával azonban északi irányba történő terjeszkedése folyamatos (Hügin 1986), valamint a kereskedelemi határok megszűnése is térhódítását elősegítő tényező. Magját különféle vetőmagokban (például: olajretek) és takarmányokban megtalálták. Mivel magja a legtöbb gazdasági haszonállat bélrendszerén
sértetlenül áthalad, így a szerves trágya és természetesen a vetőmag is a szántóterületek befertőzését eredményezheti (Meinlschmidt 2005).
3.3. Az Abutilon theophrasti jelentősége Magyarországon
A fajt a múlt század elején valószínűleg dísznövényként hozták be Magyarországra (Terpó 1987). A selyemmályva hazánkban az 1920-as évekig csak igen szórványosan fordult elő, néhány helyen volt megtalálható, mint a Közép-Duna vidékén és a Tisza mentén (Jávorka 1925).
Magyarországon, mint gyomnövényt először Priszter Szaniszló botanikus találta meg (1945-1950) Budapesten (Czimber et al. 1994). Ezt követően – mint gyomnövényről – nem találhatók feljegyzések. A hatvanas évekre a folyók árterein vagy volt árterein megjelent Belső-Somogyban, Mecsekalján, a Kisalföldön, a Duna-völgyben, Pest – Kalocsa - Baja környékén, valamint a Dráva síkon (Soó 1966). A második országos gyomfelvételezés (1969- 1971) során szántóföldön Ujvárosi Miklós is csak Szolnok megyében, Tiszaszentimrén találta meg (Ujvárosi 1971). Ujvárosi (1973) felmérése alapján elterjedése a Bükk-hegységtől északkeletre, a Sátor-hegység, valamint Zalaapáti és Gelse környékére tehető, mint útszéli ruderáliák gyomfaja. Soó (1966) is ezt az elterjedési területet jelöli meg, kiegészítve tiszántúli szórványos előfordulással. Nagyobb mértékű szántóföldi előfordulására a ’70-es évektől kezdve figyeltek fel az agrárszakemberek (Czimber et al. 1994).
A Szolnok megyei elterjedését Szautner (1984) vizsgálta. Megállapította, hogy a gyomnövény nagyobb mértékű felszaporodását már 1973-ban észlelték Törökszentmiklóson, ahol főként hibridkukoricában gyomosított. Madarász (1985) a selyemmályva Borsod-Abaúj- Zemplén megyei fertőzéséről számol be, Miskolcon már 1973-ban felfigyeltek jelenlétére, ahová valószínű fűmaggal került be. Czimber - Dömötör (1985), valamint Czimber et al.
(1987) a selyemmályva kisalföldi elterjedéséről tudósítottak, a gyomnövény térhódítása az északnyugati országrészben ekkortájt kezdődött el.
Molnár (1988) Vas megye kukorica földjeinek gyomnövényzetét tanulmányozta, és a herbicidek által szelektált gyomnövényként említi, később Lőrincz (1988) Uraiújfalu és Nick térségében észlelt 1986-ban számottevő fertőzést, majd 1987-ben további borításnövekedés volt megfigyelhető.
A harmadik országos gyomfelvételezés során (1986) már több, mint 43 ezer hektáron volt található a szántóföldön, de ebből csak 18766 ha volt kifejezetten fertőzött terület. A fertőzött terület csaknem 50%-án kukoricát, 35%-án egyéb kapás növényt és 15%-án kalászost
termesztettek (Hunyadi 1988). Mosonmagyaróváron 1986-ban a cukorrépa és szójavetésekben átlagborításával az első helyen szerepelt. Országos eloszlását tekintve ekkor a fertőzött terület mintegy 70%- a Békés, Csongrád és Szolnok megyében volt található, míg a Dunántúlon Baranya, Tolna és Komárom megyében voltak kisebb fertőzések, de Heves- és Nógrád megye kivételével más megyékben is előfordult igen kis mennyiségben.
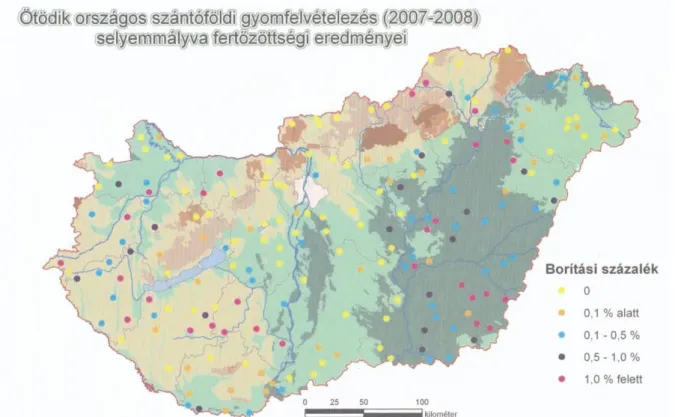
Az 1992-1993. évi cukorrépavetésekben a Dunántúlon már a 10. legveszélyesebb gyomnövény volt (Czimber et al. 1994). Térfoglalása azóta is növekedett és 1994-ben igen kellemetlen veszélyesen terjedő gyomnövényként tartották már számon. A negyedik országos gyomfelvételezés során még a 24. helyen állt, a 2007-2008-ban végzett felvételezéskor azonban már a 10. helyre lépett előre Zala megyében a kukoricavetésekben. Összességében a kukorica gyomnövényei között a 15. helyen áll a gyomok dominancia szerinti fontossági sorrendjében (Novák et al. 2009) (1. ábra).
1. ábra. A selyemmályva fertőzöttség eredményei az ötödik országos szántóföldi gyomfelvételezéskor (Novák et al. 2009).
3.4. Az Abutilon theophrasti nevezéktana és taxonómiája
A mályvafélék (Malvaceae) családja fontos gazdasági értékű fajokat foglal magában.
Közülük a legfontosabb a gyapot. A gyapotfajok (Gossypium hirsutum L. és más Gossypium-
fajok) szolgáltatják a tiszta cellulózt tartalmazó szemészeti és sebészeti vattaféléket (Gossypii lana depuratum). Újabban genetikailag módosított fajtáit egyre nagyobb mértékben termesztik Afrikában, Közép-Amerikában és Ázsia megfelelő klimatikus adottságú területein.
Idegenföldi az okra (Hibiscus esculentus L.), az okranyálkát tartalmazó meleg égövi, élelmiszernek alkalmas hibiszkuszfaj. Meleg égövi országokból beszerzett, kellemes ízű és egészséges ital készítésére alkalmas népszerű teadrog a hibiszkuszvirág (H. sabdariffa L.). A hibiszkuszok közül fontos még az illatos ábelfű (más néven ambretta). Ma érvényes tudományos neve: Abelmoschus moschatus Medic. (társneve: Hibiscus abelmoschus L.).
Indiából, Jáváról származik, trópusokon termesztik a kb. 1 méter magasra növő cserjét (Szabó 2005).
A könnyen termeszthető, sövénynek alkalmas a szíriai mályvacserje (Hibiscus syriacus L.). Ismeretes, hogy a kenaf (H. cannabinus L.) fontos rostnövény, de nálunk nem termeszthető. A kontinentális klímán biztonságosan termeszthető rostkender hivatott pótolni a vászongyártás szükségletét.
A hazai flórában igen gyakori, vadon élő mályvák a papsajtfélék. A kereklevelű mályva (Malva neglecta Wallr.) és az erdei mályva (M. sylvestris L.) levelét és virágját gyűjtik. A termesztett mórmályva (M. sylvestris L. subsp. mauritiana Thell.) antociánban (malvinban) gazdag, hivatalosan engedélyezett festőnövény.
Újra „felfedezték” a takarmánymályvát (M. verticillata L.) Közép-Európa több országában.
Alternatív növényként való termesztése nálunk is indokolt lehet.
Fontos gyógynövény az orvosi ziliz vagy fehérmályva (Althaea officinalis L.). A nálunk is termesztett mályvarózsa (Alcea = Althaea rosea L. Cav. var. atropurpurea) ugyancsak élelmiszerfestésre engedélyezett.
A hazai természetes növénytakaró értékes taxonja a Kitaibela vitifolia Willd., a nagy természettudósunkról, Kitaibel Pálról elnevezett Kitaibel-mályva (Király 2009). Kívüle akár dísznövény is lehetne még a madármályva (Lavatera) és a szép virágú, gyomnövény, a vetési varjúmák (Hibiscus trionum L.).
A selyemmályva az egyéves életformájú (Therophyta, T) életformába tartozó, tavasszal kelő, nyárutói (T4) gyomfaj (Ujvárosi 1973). A Dilleniidea alosztályba, a Malvales rendbe, a mályvafélék családjába (Malvaceae), a Hibisceae tribuszba, az Abutilinea altribuszba és az Abutilon génuszba tartozó növény (Soó 1966; Czimber et al. 1994). Arisztotelész tanítványa, a görög filozófus és író Teophrastus (i.e. 372-287) a botanika megalapítójának tekinthető. Ő a görög „sida” szót használta azokra a növényekre, amelyeket ma a Sida és Abutilon génuszokba sorolunk.
Körülbelül időszámításunk után 900 évvel az arábiai orvos és filozófus, Ibn Sina (Avicenna) gyógynövényként írta le, nevéhez köthető az Abutilon nemzetség név is (Zimdahl 1989). A későbbiekben Linné a fajt a Sida nemzettségbe sorolta és Sida abutilon L. névvel illette. Valószínűsíthető, hogy a vele közeli rokonságban lévő amerikai bársonymályvával (Sida hermaphrodita L. (Rusby)) való hasonlósága miatt (Bailey - Bailey 1976; Priszter 1998). Friedrich Casimir Medicus (1736-1808) a XVII. század végén, 1787-ben rendszerezte a Malvaceae növénycsalád tagjait, és az azóta is elfogadott tudományos név: Abutilon theophrasti Medicus. az ő személyéhez köthető. Szinonim elnevezése még az Abutilon avicennaea Gaertner. (Bailey - Bailey 1976; Haensel 2005). A görög filozófus és tudós Theophrasztosz (Türtamosz vagy Tyrtamos) ékesszólása alapján kapta a Theophrasztos (=Isteni beszédű) nevet magától Arisztotelésztől. Ereszosz nevű városból származott, ezért Plumier a tiszteletére Eresia-nak nevezett el egy növénynemzetséget, melyet Linné Theophrasta-ka keresztelt át. Több növényfaj is kapta róla a nevét, mint a krétai datolyapálma (Phoenix theophrasti) és a selyemmályva (A. theophrasti) (Molnár 2009). Angol nevei a velvetleaf , elephant ears, cottonweed, china jute, flowering maple és abutilon hemp, német nyelvű elnevezése a Lindenblättrige Schönmalve és a Samtpappel (Taksonyi 1992; Haensel 2005). A magyar selyemmályva néven kívül használatos a szépasszony tenyere, sárda, selyemsárda, sárgamályva és a díszmályva elnevezés is (Czimber et al 1994).
Az Abutilon nemzetség egyes szerzők szerint 80 (Hanf 1983; 1999; Kazinczi et al. 2001), mások szerint több mint 200 fajt foglal magába (Fryxell 1988). A génuszba tartozó fajok többsége trópusi elterjedésű, találhatunk köztünk fásodó szárú cserjéket, lágyszárúakat, évelőket és néhány egyéves növényt (Jäger 1991) (1. táblázat).
A selyemmályva idegenföldi rokona a hibrid selyemmályva (A. x hybridum hort. ex Siebert et Voss). Dísznövény (kínai lámpának is nevezik), a rózsás selyemmályvának (A. darwinii) és a kerti selyemmályvának (A. pictum) a hibridje. Az A. indicum (L.) Sweet, az indiai selyemmályva paleotrópusi és szubtrópusi területeken terjedő, peszticid hatású gyomnövény (Wiersema – León 1999).
1. táblázat. Néhány Abutilon génuszba tartozó faj elterjedési területe és felhasználása (Kazinczi et al. 2001)
No. Tudományos név Felhasználás Elterjedés 1. A. asiaticum (L.) Sweet rostnövény Trópusok, Mauritius 2. A. bedfordiannum Hil. rostnövény Brazília, Ausztrália
3. A. fructicosum Guill. & Perr. Angola
4. A. glaucum Don.
Szudán
Syn. A. pannosum Schlecht
5. A. graveolens Sweet termesztett, magolaj Trópusok, Oroszország
6. A. guinense Bak. & Exell
7. Abutilon x hybridum termesztett
Kenya, Ghána (A. darwinii x A. pictum) dísznövény
8.
A. indicum Sweet. gyógynövény India, Vietnám, Ghána, Syn. A. mauritanum Medic. rostnövény
Kenya, Nigéria, Thaiföld, Kambodzsa, Indonézia 9. A. intermedium Hochst
rostnövény Trópusok, Madagaszkár Syn. A. angulatum Cav.
10. A. jackquinii Don
rostnövény Texas, Mexikó, Nyugat- India
Syn. A. lignosum Cav.
11. A. megapotamicum Spring dísznövény Brazília, Trópusok
12. A. molle Sweet. Argentína, Hawaii, USA
13. A. muticum Sweet magja élelmezési
célokra Trópusok, Szudán Syn. A. asiaticum
14. A. ochsenii (Phil) Reiche
Syn. Corynabutilon ochsenii Kearney
15. A. octocarpum Muell rostnövény Ausztrália
16. A. oxycarpum Muell rostnövény Brazília
17. A. persicum Merr
rostnövény India
Syn. A. polyandrum Roxb.
18. A. pictum Walp. dísznövény USA
19. A. polyandrum Don. rostnövény India
20. A. striatum Dik Ex. Lindl. dísznövény Brazília, USA 21.
A. theophrasti Medik. gyomnövény Észak-Afrika, Ázsia, Európa, India, Észak- Amerika (kozmopolita) Syn. A. avicennae Gaertn., rostnövény
Sida abutilon L. gyógynövény
22.
A. trisulcatum Jack
rostnövény, gyógynövény
Dél-Amerika, Mexikó, Nicaragua, Kuba Syn. A. triquetum Sweet,
Sida trisulcata
23. A. vitifolium Presl. = Corynabutilon vitifolium Kearney
3.5. Az Abutilon theophrasti morfológiája
A selyemmályva egyéves, karógyökerű, felálló hengeres szárú, 60-150 cm [80-250 cm (Jäger 1991)] magas, szórt levélállású növény (6. ábra). Az egész növény bársonyosan molyhos, levelei nagyok [7-20 cm (Warwick - Black 1988)], szíves kerekdedek, vállból hosszan, hirtelen kihegyezettek, csipkés szélűek (2. ábra). Virágai egyesével vagy kevés számban ülnek a levelek hónaljában. A virágkocsányok rövidebbek, mint a levélnyél. A csészelevelek széles tojásdadok, kihegyezettek, számuk öt. Sziromlevelei visszás szíves háromszögűek, a csészeleveleknél hosszabbak, élénksárga színűek, számuk szintén öt (3.
ábra).
2. ábra. A selyemmályva levele (Fotó: Nagy V.) 3. ábra. A selyemmályva virága (Fotó: Nagy V.)
Termése 15 kétcsőrű, tompa, körkörösen álló résztermésből összetett tok. A résztermések 1,5cm hosszúak, éretten feketék, két vagy több magvúak. (Ujvárosi 1973) (4. ábra).
A magok aszimmetrikus vese alakúak, laposak, 1mm szélesek és 2-3mm hosszúak (Warwick – Black 1988) (5. ábra). A növény magprodukciója körülbelül 8000 mag/növény (Spencer 1984). Kazinczi et al. (2007) azonban 1000 mag/növény körüli produkcióról ír, amely a növénysűrűség növelésével csökken, 5 és 10 darab növény/m2 egyedsűrűségnél csak mintegy 430 mag/növény. Ezermagtömege 6-11g (Baloch et al. 2001).
4. ábra. A selyemmályva termése (Fotó: Nagy V.) 5. ábra. A selyemmályva magvai (Fotó: Nagy V.)
6. ábra: A selyemmályva (Bíró Krisztina rajza, szerzői jogvédelem alatt)
3.6. Az Abutilon theophrasti csírázásbiológiai sajátosságainak bemutatása
3.6.1. A magnyugalom
A mezőgazdaságilag művelt talajok felső 15cm-es rétegében hektáronként kb. 4-60 millió életképes gyommag található, aminek csupán 10%-a kel ki egy évben, bár a földbe került friss magvak a legnagyobb mértékben az első évben csíráznak ki (Taylorson 1971, Beuret 1984).
A talaj magkészlete két csoportra osztható aszerint, hogy a magvak melyik évben csíráznak ki. Így beszélhetünk tranzit és perzisztens magkészletről (Thompson-Grime 1979, Thompson 1987, Walck et al. 1996).
A magnyugalom, vagy más néven dormancia, a magok azon állapota, amikor azok a számukra optimális körülmények között sem csíráznak ki (Zimdahl 1999). Így a gyomnövények ellen való védekezés gátló tényezője, hiszen a gyomok ellen csak a magvak nyugalmi állapotának megszűnése után tudunk védekezni (Magyar-Lukács 2002). Ezzel az
„időbeni terjedési stratégiával” biztosítják a gyomnövények a mezőgazdasági területek hosszú távú fertőződését (Zimdahl 1999).
A magnyugalmi állapotnak számos típusa létezik, emellett több csoportosítási szempont is elterjedt (Hilhorst-Toorop 1997).
A magnyugalom felosztására jelenleg Nikolaeva által történt csoportosítás a legelfogadottabb, mely szerint endogén- és exogén dormanciáról beszélhetünk (Toole-Toole 1941). Az exogén dormanciának három formája létezik: fizikai, kémiai és mechanikai. A fizikai magnyugalom esetében a csírázás fő akadálya a maghéj vízzel szembeni impermeabilitása, azaz a keményhéjúság. Ez esetben fásodott sejtekből álló paliszád réteg található a maghéjban (Corner 1976).
3.6.1.1. A selyemmályva dormanciája, a magvak életképessége
Az utóbbi évtizedekben néhány melegigényes, egyéves gyomfaj elszaporodása volt megfigyelhető Magyarországon. Ezen fajok között találjuk a Malveceae családba tartozó selyemmályvát (Abutilon theophrasti Medic.) is. A terjedésében - egyéb tényezőkön kívül - nagy szerepe van a faj biológiai sajátosságainak (Kazinczi et al. 1991; Nagy et al. 2009;
Novák et al. 2009).
A selyemmályva magvai szárazon tárolva vagy a talajban több mint 50 éven keresztül megőrzik életképességüket. Talajban történő 2,5 éves tárolás után a magjai 58%-ban csíráztak (Egley - Chandler 1978), 3 év után 70%-os (Stoller - Wax 1974), 4 év után 37%-os (Lueschen - Andersen 1980), 5,5 év után 36%-os (Egley - Chandler 1983), valamint 39 év talajban töltött idő után is 43%-os csírázást tapasztaltak (Toole - Brown 1946).
Duvel 39 éves tartamkísérletében (1902-1941) 55 és 105cm talajmélységben a selyemmályva magjainak csírázása 39 év után is meghaladta a 15%-ot (Hunyadi - Kazinczi 2000).
Saját kísérletünkben azt tapasztaltuk, hogy minden esetben a sötétebb színű magvak csíráztak nagyobb százalékban annak ellenére, hogy a sötétebb magszín erősebb nyugalmi állapotot feltételezne. A nyugalmi állapot a kor előre haladtával folyamatosan megszűnik, a 24 és 21 éves magtételek csírázási eredménye jobbnak bizonyult a frissen begyűjtött magvakhoz képest (Nagy – Nádasy 2010).
Tehát a selyemmályva magja képes a művelt területeken perzisztens módon a talajban elfeküdni. Ez a tulajdonság a magok nyugalmi állapotának köszönhető (Kremer et al. 1984).
A nyugalmi állapotát több tényező befolyásolja, de a csírázás fizikai korlátja a maghéjban található sűrű paliszád sejtsor (Kremer 1986; Warwick - Black 1988).
Hyde (1954) kimutatta, hogy a hilum higroszkóposan aktivált szelepként működik, amely lehetővé teszi, hogy a mag száraz légkörben vizet veszítsen, de megakadályozza, hogy nedves légkörben vizet szívjon fel. Barton (1965) szerint a hilum szerepet játszik a keményhéjúság kialakításában. A hilum egyirányú regulációs szerepére Popcov (1953) is felhívja a figyelmet, mely tulajdonságra a pillangósvirágúakon kívül az Abutilon theophrasti esetében is rámutat.
A Malvales-rendből elsősorban a Malvaceae-család toktermésű növényfajai teremnek impermeábilis héjú magvakat (Sumner - Cobb 1967).
A keményhéjű magot érlelő növények ökológiai spektrumában a Therophiton-ok és Hemitherophiton-ok 42,6%-os részesedéssel szerepelnek, a Magyarországon található virágos és virágtalan növények ökológiai spektrumában előforduló 23,5%-kal szemben. A keményhéjúság tehát az egyéves növények jellemző tulajdonsága (Borhidi 1969). A keményhéjú magvakkal rendelkező növények flóraelem spektruma szintén eltér a magyar flóra elemeinek százalékos megoszlásától. Amíg a magyarországi flóraelemek között az eurázsiaiak 22,52%-kal szerepelnek, addig a keményhéjú magvakat termők között ezek 36%- ot tesznek ki. Ugyancsak nagyobb a mediterrán és adventív elemek jelenléte is (Czimber 1980b).
A selyemmályvát számos szerző a keményhéjúság tipikus példájaként írja le, amely a maghéj vízzel szembeni impermeábilitásából fakad (Egley - Chandler 1978; Lueschen - Andersen 1980; Horowitz - Taylorson 1984; Warwick - Black 1986; Czimber et al. 1994). A frissen gyűjtött, tökéletesen beérett, fekete színű selyemmályva magvak gyakorlatilag mind keményhéjúak. A kevésbé érett (zöld, sárgásbarna, sárga, barna színű) magvai csak kisebb százalékban keményhéjúak. Ezeknél ugyanis a köldök (hilum) és a csírakapu (mikropile) sejtjei még nem záródnak teljesen és a víz bejuthat a csírázáshoz (Steinbauer - Grigsby 1959;
Czimber et al. 1994).
Számos kémiai és fizikai módszer tudja hatékonyan feloldani a primér magnyugalmat. A legjobb módszer a magvak leforrázása (Muliken - Kust 1970; Khedir - Roeth 1981). Egyéb módszer lehet még a kénsavas kezelés és a mechanikia szkarifikáció (Khedir - Roeth 1978;
Horowitz - Taylorson 1985).
Más kutatások szerint azonban a selyemmályva esetében nem csak primér magnyugalmi állapot figyelhető meg, hanem egy második u.n. szekunder dormancia is, amely embrionális eredetű (Cardina – Sparrow 1997).
A selyemmályva csírázása szezonális jelleget mutat, a csírázás fő időszaka április végétől június közepéig tart (Meinlschmidt et al. 2004).
3.6.2. A csírázásra ható tényezők
A gyommagvak csírázási dinamikáját nagymértékben befolyásolják a különböző évjáratok hőmérséklet- és csapadékviszonyai továbbá az agrotechnikai és talajművelési eljárások (Kazinczi 2005).
3.6.2.1. A magvak talajbeli mélységének hatása a csírázásra
A különböző szántóföldi talajművelési munkák igen változatos körülményeket alakítanak ki a különböző mélységben elfekvő, különböző korú és eltérő magnyugalmú gyommagvak csírázásához. A gyommagvak között eltérő csírázási igényei miatt mindig vannak olyanok, amelyek kényszernyugalmi helyzetüktől mentesülve életfeltételeiket megtalálják és kicsíráznak (Roberts – Dawkins 1967; Máthé – Percsényi 1968; Bencze 1969).
A gyommagvak csírázásukhoz eltérő körülményeket igényelnek, így a különböző talajtípusokban is más és más gyomfajok találják meg kicsírázásukhoz szükséges igényeiket.
Az optimális körülmények a talajtípustól függően, az egyes fajok esetében eltérő mélységben
szabályozó tényezője, amely hatással van a csírázás idejére és mértékére is (Lovato – Viggiani 1974). A talajmélység növekedésével a csírázás mértéke általában csökken.
Általános szabályként elmondható, hogy minél nagyobb a mag mérete, annál mélyebbről képes kicsirázni (Froud – Williams et al. 1984; Fykse 1984). A gyommagvak számára előnyösebb, ha csak a talajfelszínhez közel eső rétegekből csíráznak ki, különben a mag endospermiumának tartalék tápanyagkészlete kimerül, mielőtt a csíranövény autotróffá válna (Roberts et al. 1987).
A csíranövények talajban történő elpusztulása nem csak az endospermium kimerülésének következménye, a selyemmályva esetében a 8 és 10 cm-es mélységből csírázó magvak nagyarányú (20 és 40%) pusztulását a Pythium ultimum gombafaj okozhatja (Davis – Renner 2007).
A fényigényes magvak, méretüktől függetlenül, csak a talajfelszínről, és mintegy 1 cm-es mélységéből képesek kicsírázni, ahová a diffúz fény eljut. A talaj felső 5 cm-es rétegéből csírázik a szántóföldi gyomfajok 90%-a. A selyemmályva azonban képes 5-10 cm-es mélységből is csírázni (Kazinczi – Hunyadi 1992; Benvenuti et al. 2001). Ennél mélyebben azon fajok magvai csíráznak, amelyek magvai mintegy 5mg tömegűek.
Annak ellenére, hogy a talajmélység a csírázás jelentős tényezője, csak mint a csírázást módosító (esetleg késleltető) tényezőt szokták figyelembe venni (Kazinczi 2005).
3.6.2.2. A műtrágyák hatása a csírázásra és növekedésre
A különböző növényvédő szerek, elsősorban herbicidek mellett a műtrágyák is jelentős csírázást befolyásoló hatással rendelkezhetnek (Blumrich 1992). Több szerző bizonyította, hogy a magvak mellé kerülő műtrágyaszemek a csírázást megakadályozó magas só- koncentrációt okozhatnak (Harris 1915; Cummins – Parks 1961; Hunter – Rosenau 1966). A csírázást befolyásoló hatás természetesen számos tényezőtől függ, mint például a műtrágya fajtája, illetve hatóanyaga, valamint a környezeti tényezők (Béres – Sárdi 1994).
Az ammónia csírázás és növekedésgátló hatásáról számolt be Colliver – Welch (1970). Azt tapasztalták, hogy a kukorica növények csírázása és fejlődése gátolttá vált a nitrogén 994 ppm koncentrációjánál. A KNO3 oldat azonban a gyommagvak nagy részénél serkentőleg hat a csírázásra. Steinbauer – Grigsby (1957) vizsgálatai során 85 faj felénél kimutatták a KNO3
serkentő hatását. Béres – Sárdi (1994) több műtrágya és koncentráció hatását tanulmányozták a búza (Triticum aestivum L.), a ragadós galaj (Galium aparine L.) és az ebszékfű (Tripleurospermum inodorum L. Schultz-Bip.) csírázására. Azt tapasztalták, hogy a ragadós
galaj csírázását az összes makrotápanyag (N, P és K), minden műtrágyában és minden alkalmazott koncentrációban csökkentette. Az ebszékfű esetében az ammónium szulfát és a triplefoszfát gátló, míg az NH4NO3 és a káliumszulfát serkentő hatásúnak bizonyult. A vizsgált fajok közül a búza reagált legkevésbé a kezelésekre. Csak a karbamid gátolta a csírázást 20%-kal 1000ppm koncentráció esetén.
Korábbi vizsgálatok kimutatták, hogy a gyomnövények tápanyagokért történő kompetíciójában a nitrogénért folyó harc a legerősebb (Keely 1965). Általánosan elmondható, hogy a gyomnövények a szerveikben átlagosan több káliumot halmoznak fel, mint a kultúrnövények és ez kiváltképp a kétszikűekre igaz (Vengris et al. 1953). A selyemmályva tápanyagtartalmát és tápanyagreakcióját számos kutató vizsgálta. Debreczeniné (1988) a selyemmályva magas nitrogén tartalmát tapasztalta, eredményei szerint a növényre a 4,5%
nitrogén, 0,4% foszfor és 2% kálium tartalom jellemző.
Taksonyi (1992) a selyemmályva különböző szerveinek tápelem koncentrációját tanulmányozta a növény vegetációs periódusa során több mintavételi időpontban.
Tapasztalatai szerint a nitrogén szintje a vegetáció elején a levelekben a legmagasabb (4-4,5%
körüli), majd a termésérés kezdetétől minden növényi rész nitrogén koncentrációja csökken a termések nitrogén elvonó képessége miatt.
A foszfor mennyisége a termésekben a legnagyobb, 0,45%, ezután csökkenő sorrendben a levél , a szár és a gyökér következik.
A kálium koncentráció is a termésekben bizonyult a legmagasabbnak, a levelek és a szár kálium tartalma ingadozott a vegetációs periódus folyamán.
Béres et al. (1996) a tápanyagstressz hatását vizsgálták a selyemmályva növekedésére.
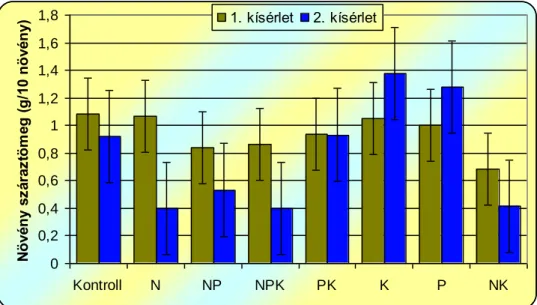
Kimutatták, hogy a tápanyaghiány az Abutilon hajtástömegében jelentős depressziót okozott a kiegyensúlyozott tápanyagellátásban részesülő növényekhez képest. A tápanyaghiány okozta stressz hatása a hajtások száraztömegében kifejezetten a N hiány esetében mutatkozott meg.
Ezt a P hiány hatása követte, legkevésbé érzékenyen a K hiányára reagált a selyemmályva. A gyökerek száraztömegében a legnagyobb csökkenést legtöbbször a P hiányos kezelések okozták. A hajtás:gyökér arány a N hiány következtében alacsonyabb volt, míg a K hiánynál növekedett.
A korábbi kísérleti eredmények értelmében elmondható, hogy a talaj gyommagkészletének csírázása a műtrágyák helyes alkalmazásával befolyásolható és a műtrágyák szakszerű alkalmazása a gyomszabályozás hatékony eszköze lehet (Béres – Sárdi 1994).
3.7. Magasabb rendű növények közötti kölcsönhatások
3.7.1. A kompetíció
A magasabb rendű növények közötti versengés, azaz a kompetíció felismerése nem újkeletű. Már a Bibliában is találhatunk utalásokat, illetve Darwin az 1859-ben megjelent
„Fajok eredete” című művében a természetben mindenütt fellelhető jelenségként írja le. Az oxfordi angol szótárban a kompetíció fogalmát már 1933-ban pontosan definiálták, mely szerint a kompetíció két vagy több egyed ugyanazon időben ugyanazon tényező megszerzésére irányuló törekvése (Kazinczi 2000). A későbbiekben a növények közötti kölcsönhatásokat interferenciának nevezték el, és két külön ágra: a kompetícióra és az allelopátiára tagolták (Harper 1977). A szakirodalomban jelenleg legtöbbet idézett definíció azonban Donald-tól (1963) származik, mely szerint a „kompetíció akkor lép fel, ha két vagy több szervezet mindegyike keresi a maga sajátos szükségleteinek kielégítését valamilyen adott tényezőből, amikor a közvetlen ellátás ebből a tényezőből alatta marad a szervezetek együttes igényeinek”. Természetes körülmények között azonban számos tényező befolyásolhatja a növények közötti interferenciát (Bleasdale 1960).
3.7.1.1. Az Abutilon theophrasti kompetíciós képessége
A selyemmályva (Abutilon theophrasti) évről–évre több gondot okoz a növénytermesztésben. Más melegigényes fajokhoz hasonlóan látványosan nőtt a borítása az utóbbi évtizedekben (Szőke 2001). Erőteljes növekedése, kiváló hő- és szárazságtűrése révén kompetíciós előnyhöz juthat egyes kultúrnövényekkel szemben (Patterson 1992).
Kompetíciós és terméscsökkentő hatását több kutató vizsgálta különféle kultúrákban, mint például: kukoricában (Czimber – Dömötör 1985; Czimber et al. 1987; Varga et al. 2000;
Werner et al. 2004; Kovács et al. 2006; Dávid et al. 2006; Dávid – Kovács 2007; Kazinczi et al. 2007), napraforgóban (Dávid et al. 2006), szójában (Gressel és Holm 1964; Retig et al.
1972; Eaton et al. 1976; Oliver 1979; Colton – Einhellig 1980; Dekker 1981; Sadeghi et al.
2002; 2004) és cukorrépában (Schweizer – Bridge 1982; Renner – Powel 1991; Bördner et al.
2004; Haensel 2005), valamint cirokban (Traoré et al. 2003).
Számos kutató szerint az Abutilon-nal fertőzött táblákon a kultúrnövények termésvesztesége a selyemmályva kompetíciós előnyéből adódik. Hagood et al. (1980), McConnaughay – Boose (1998) és Holt – Boose (2000) a vízért, míg Stoller – Wooley
(1985), Satin et al. (1992) és Jurik – Akey (1994) a fényért történő versengést helyezi előtérbe. Oliver (1979), valamint Roeth (1987) szerint a tápelemeknek a kultúrnövénytől történő elvonásában rejlik e gyomnövény erős kompetíciós képessége.
A gyomnövény zavaró hatást fejt ki a szójabab vízháztartására (Retig et al. 1972).
Csökkenti a szója szárazanyag akkumulációját (Gressel - Holm 1964; Eaton et al., 1976;
Oliver 1979; Colton – Einhellig 1980). Jelentős termés (maghozam) veszteséget okoz (Gressel – Holm 1964; Eaton et al. 1976; Oliver, 1979). Akadályozza az ág- és virágzati rügyfejlődést (Gressel – Holm 1964).
Dekker (1981) szerint már alacsony selyemmályva fertőzöttség (2,4-3,7db/m2) is csökkenti a szója szárazanyag felhalmozódását, a virág- és rügyképződést.
Stanifort - Weber (1956, in Dekker, 1981) kutatásai alapján a terméscsökkentő hatás akkor a legnagyobb, ha az A. theophrasti 10 napon belül kiemelkedik a vetésből. 130-140 db/m2 fertőzés esetén a hüvelyek száma már 47%-kal csökken.
Sadeghi et al. (2004) eredményei azt mutatták, hogy a vizsgált egyéves gyomfajok közül a selyemmályva csökkentette leginkább a szója száraztömegét, levélterület indexét és relatív növekedési rátáját.
Czimber - Dömötör (1985) az Abutilon theophrasti kártételét vizsgálták kukoricában. A gyommentes és selyemmályvával fertőzött parcellák csöves termését, illetve a kukorica és a selyemmályva talajfelszín feletti tömegét mérték és hasonlították össze. A vizsgálati eredmények alapján megállapították, hogy a gyommentes és a selyemmályvával fertőzött parcellák szemtermése között szignifikáns különbség van a gyommentes kukorica javára. A gyomos parcellákon a selyemmályva és a kukorica száraztömege, illetve a szemtermés mennyisége között megbízható negatív korrelációt tapasztaltak.
Mosonmagyaróvári vizsgálatok szerint 0,8 kg/m2 száraztömegű selyemmályva mennyiségnél a fertőzött terület átlagtermése csupán 27%-a volt a gyommentes terület átlagtermésének. Szójavetésben 42%-os terméskiesést 0,256kg/m2 selyemmályva száraztömeg okozott, cukorrépában pedig 35% kiesést 0,168kg/m2.
Weaver - Hamill (1985) által végzett kompetíciós vizsgálatok szerint az Abutilon theophrasti 6-os pH-nál 3,7 t/ha-ral csökkentette a kukorica szemtermését. A levélanalízis vizsgálatok alapján megállapítható, hogy a selyemmályva hatására a kukorica levelének tápanyagtartalmában koncentrációcsökkenés léphet fel.
Kazinczi et al. (1991) vizsgálatai szerint szabadföldi interferencia kísérletekben az Abutilon theophrasti erősebb kompetítornak bizonyult mint a szója és a kukorica. A
állományhoz képest. A szója esetében a nitrogénért folyó versengés, a kukoricánál pedig a káliumért történő kompetíció volt erős. A napraforgó ellenben jelentősen csökkentette a gyomnövény magprodukcióját és késleltette generatív fejlődését.
Werner et al. (2004) kimutatta, hogy kukorica esetén a szilázs hozam érzékenyebben reagál a selyemmályva jelenlétére, mint a szemes termény hozama.
A selyemmályvát a cukorrépa vetések veszélyes gyomnövényeként is számon tartják.
Schweizer – Bridge (1982) vizsgálatai szerint 30méter hosszúságú cukorrépa sorban 6, 12, 18, valamint 24 darabos selyemmályva sűrűség a termésmennyiséget 14, 17, 25, valamint 30%- kal vetette vissza. Renner – Powel (1991) kísérletei bebizonyították, hogy cukorrépában 7,6 méterenként 1 selyemmályva növény az a tolerálható gyomsűrűség, amely még nem okoz veszteséget.
Debrecenben, napraforgóban végzett kísérletek szerint 10 db selyemmályva/m2 növénysűrűség 37%-kal csökkentette a kultúrnövény termésének mennyiségét (Dávid et al.
2006).
3.7.2. Az allelopátia
Már régóta ismert jelenség, hogy a természetes növénytársulásokban megtalálhatók olyan fajok, amelyek képesek más fajok betelepedésének megakadályozására. Egyes növények szomszédaikra gyakorolt negatív hatása olykor annyira szembetűnő, hogy a közös tápanyagforrásokért folytatott versengéssel már egyáltalán nem magyarázható. Ilyen esetekben a növényi biomasza látványos csökkenése csak az egyik fajt érinti, a másikat nem.
A magyarázatot az ún. allelopátia jelensége adja meg (Szabó 1984; Béres 2000; Brückner – Szabó 2001). Molish (1937) a növények és mikroorganizmusok közötti összes biokémiai kölcsönhatást allelopátiának nevezte. Ez a görög szóösszetétel kölcsönös ártalmat, egymás elszenvedését jelenti, az allelon (=kölcsönös, egymás) és a pathos (=ártalmas, elszenvedni) szavak összetételének eredményeként.
Muller (1966) allelopátián az egyik növénynek, vagy mikroorganizmusnak a másikra gyakorolt közvetlen vagy közvetett káros hatását értette, mely a környezetben jutó kémiai vegyületek létrehozása által valósul meg.
Rice (1974) „Allelopathy” címmel monográfiát jelentetett meg, melyben az allelopátiát egyik növény (beleértve a mikroorganizmusokat is) másikra gyakorolt közvetlen vagy közvetett káros hatásaként értékeli, melyért a növényből környezetébe jutó vegyületek felelősek.
1999-ben Thunder Bay-ben a II. Allelopátiás Világkongresszuson a fenti kritikák figyelembe vételével megfogalmazták az allelopátia jelenlegi definícióját: „Minden olyan folyamat amelyben növények (algák, baktériumok, gombák, vírusok) által termelt szekunder metabolitok befolyásolják a mezőgazdasági és biológiai rendszert” (IAS Newsletter 1999).
Reese (1979) az allelokemikáliákat olyan nem tápanyag természetű kémiai anyagokként határozta meg, amelyek a környezetbe jutva befolyásolhatják más növények növekedését és egészségét.
Ezek az allelokemikáliák, vagy szekunder, másodlagos anyagcseretermékek az adott donor növénytől függően sokfélék lehetnek. Az allelopátiás szempontból jelentős hatóanyagok nagyrészt olyan vegyületcsoportokat foglalnak magukban, melyek egyes kémiai tulajdonságaik és fiziológiai hatásaik tekintetében hasonlóságot mutatnak. Allelopátiás szempontból különösen a következő vegyülettípusok hatásosak:
1, Fenoloidok (polifenolok, fenolsavak, kumarinok, naftokinonok, antrakinonok)
2, Terpenoidok (monoterpének, szeszkviterpén-laktonok, diterpének, triterpenoid szaponinok)
3, Poliinek
4, Azotoidok (alkaloidok, kromoalkaloidok, cianogén glükozidok)
5, Glikozinolátok (Evenari 1949; Vancura 1964; Whittaker - Feeny 1971; Rice 1974;
Brückner – Szabó 2001).
Az allelokemikáliák a környezetbe kimosódás, párolgás, gyökerek általi kiválasztás és a növényi maradványok elbomlása során jutnak (Wilson - Rice 1968; Parenti - Rice 1969; Rice 1974; Putnam - Duke 1978; Inderjit - Dakshini 1991, 1992, 1995).
Hatásmechanizmus szempontjából ezeknek az anyagoknak a rendszerezése meglehetősen bonyolult (Siegler 1996). Hathatnak a növényi sejtnövekedésére (Zobel – Lynch 1997), a membránok permeabilitására (Baziramakenga et al. 1995; Calera et al. 1995; Friebe et al.
1997), a növényi táplálkozásra (Pellisier 1993; Baar et al. 1994; Boufallis –Pellisier 1994; Yu – Matsui 1997), a fotoszintézisre (Baziramakenga et al. 1994), a növény vízháztartására (Penuelas et al. 1996), valamint a növények enzimaktivitására, a fehérje-, a zsírsav- és a porfirin szintézis blokkolásán keresztül (Prasad – Subhashini 1994).
3.7.2.1. Az Abutilon theophrasti allelopátiája
A selyemmályva gátló hatását számos szerző az allelopátia jelenségének tulajdonítja. A selyemmályva allelopatikus tulajdonságait több kultúrnövényen vizsgálták, pl. napraforgó, kukorica, szója, lucerna, paradicsom, stb., ahol csírázást és növekedést gátló hatását is értékelték (Gressel – Holm 1964; Sterling et al. 1987; Kazinczi et al. 1991, 2001). Elsősorban a növényi maradványok csírázás- és fejlődésgátló hatásáról számol be Klevorn – Peters (1979), Bhowmik – Doll (1980, 1982, 1984) és De Felice et al. (1984, 1988).
Az allelopatikus hatás valószínűleg a vízfelvétel gátlásán és a klorofill mennyiségének csökkenésén keresztül valósul meg (Colton – Einhellig 1980).
Gressel – Holm (1964) a mag vizes kivonatának csírázás- és növekedésgátló hatásáról ír, sterilizált és nem sterilizált talajon is kimutatták a selyemmályva kivonatok gátló hatását paradicsom csíranövényeken. Dekker – Megitt (1983) 50%-os termésveszteséget tapasztalt szója kultúrában, amelyet elsősorban a selyemmályva gyökeréből származó gátló hatású vegyületeknek tulajdonított. Sterling et al. (1987) szerint az allelopatikus hatás előidézője a levélnyélen és a száron lévő mirigyszőrökből kivonható vízoldható exudátum.
Az allelopatikus hatást okozó vegyületek többfélék lehetnek. Elmore (1980a) a selyemmályva esetében a savas frakciónál találta a legnagyobb aktivitást, mely fenolokat tartalmazott. Ezek gátló hatását Colton - Einhellig (1980) is kimutatták retek csíranövényeken. Ezen kívül Elmore (1980b) szerint a selyemmályva allelopatikus hatását a magban lévő szabad aminosavak is okozzák. Véleménye szerint a magban magas koncentrációban lévő arginin kifejtheti ezt a hatást, továbbá flavonoidok is (Paszkovski – Kremer 1988). Elmore (1980a) vizsgálatai szerint az arginin a saláta magjaira csírázásgátlóan hat, tehát elképzelhető, hogy a selyemmályvánál is hasonló hatása lehet.
Retig et al. (1972) azt tapasztalta, hogy az Abutilon theophrasti magjainak csírázása során keletkező diffúz anyagok rendellenességet okoznak a paradicsom és a káposzta gyökerein. A gyökerek megnagyobbodtak, a gyökerekben lévő parenchyma-sejtek számának és méretének növekedése következtében. A szerzők szerint a növények gyökereinek anatómiai elváltozásai változásokat okozhatnak a táplálék- és vízfelvételben.
Gressel és Holm (1964) vizsgálatai szerint a selyemmályva magjának vízben oldódó kivonatai különböző mértékben gátolják a lucerna, a retek és a fehérrépa csírázását. A magkivonatok faj- és variánsspecifikusnak bizonyultak. Az érintetlen, intakt Abutilon magcsírák gátolták a retek és a paradicsom csírázását és gyökérfejlődését. A gyomnövény
levélszöveteiből vett kivonatokról is kimutatták, hogy gátló vegyületeket tartalmaznak, de nem feltétlenül ugyanazokat.
Colton – Einhellig (1980) azt tapasztalták, hogy az Abutilon theophrasti növénymaradványai nem gátolták a szójapalánták növekedését 0-28 napig, de csökkentették a szója gyökérnodulációját. A palánták növekedését a vízben oldódó levélkivonatok gátolták. A kivonatok fokozták a szójalevél diffúz ellenállását és csökkentették a levél vízfelvevőképességét, ennélfogva a kezelt növények vízstresszben szenvedtek. Ezenkívül a kivonatok csökkentették a szójapalánták „A” és „B” klorofillját, valamint az össz-klorofillt.
Dávid – Radócz (2002) a különböző mértékű tápanyag utánpótlásnak az inhibitorok termelésére gyakorolt hatását vizsgálták laborban. Perliten nevelt műtrágyázott és nem műtrágyázott selyemmályva növények kivonatait tesztelték cukorrépa csíranövényeken csapvíz és híg sósavas kivonószerek felhasználásával. A hajtáskivonatok különböző mértékben gátolták a cukorrépa csíranövényeket. A csírázás és az 50 magból kelt növények által elért összes gyökérhosszúságot értékelve azt tapasztalták, hogy a tápanyag utánpótlás nélkül nevelt egyedek gátló hatása erősebb volt vizes kivonatok esetében, mint amelyek műtrágyát kaptak. Fordított jelenséget tapasztaltak a savas kivonatoknál. A gyökérből készített oldatok a csírázást általában enyhén gátolták, a gyökér növekedését viszont nem.
Vizsgálták emellett a víz utánpótlás hatását is az allelopatikus anyagok termelésére.
Megfelelő vízutánpótlásban részesülő selyemmályva esetében a gátló hatás vizes kivonószerrel volt erősebb, savas és lúgos esetben enyhébb. A vízhiányban nevelt selyemmályva esetében viszont a savas és lúgos kivonószer hatására kismértékben csökkent a gyökérprodukció a vizes hajtáskivonathoz képest.
Keszthelyi kísérletek kimutatták, hogy a selyemmályva levél- és gyökérkivonatának, elhalt szár-, levél- és gyökérmaradványainak csírázás és növekedésgátló hatása van (Kazinczi et al.
1991). Selyemmályvából készített kivonatok allelopatikus hatását vizsgálták (Kazinczi et al.
2008) parlagfűre (Ambrosia artemisiifolia L.). Megállapították, hogy tenyészedényes kísérletben a selyemmályva különböző részeiből készített vizes kivonatok alacsony koncentrációban serkentették a parlagfű növekedését, a stimuláló hatás erősebb volt, ha a parlagfű növények a kivonatot gyökéren keresztül vették fel, mely során a kivonatokat tápanyagforrásként hasznosították. Magasabb töménységben a vizes kivonatok a parlagfű gyökérnövekedését jobban gátolták, mint a csírázást.
Šćepanović et al. (2007) az A. theophrasti föld feletti friss hajtásainak és leveleinek, valamint külön a gyökér vizes oldatának allelopatikus hatását tanulmányozták a kukorica