PANNON EGYETEM
Georgikon Mezőgazdaság-tudományi Kar, Keszthely Állattudományi és Állattenyésztéstani Tanszék
DOKTORI (PhD) ÉRTEKEZÉS
Állattenyésztési és Agrár-Környezettudományi Doktori Iskola Iskolavezető: Dr. Anda Angéla DSc
A fűrészlábú szöcskék (Saga spp.) biológiája, különös tekintettel a Saga pedo (PALLAS, 1771) fajra
Készítette:
Kolics Balázs
Keszthely
2009
2
3
A fűrészlábú szöcskék (Saga spp.) biológiája, különös tekintettel a Saga pedo (PALLAS, 1771) fajra
Értekezés doktori (PhD) fokozat elnyerése érdekében
Írta:
KOLICS BALÁZS
Készült a Pannon Egyetem Állattenyésztési és Agrár-Környezettudományi Doktori Iskola keretében
Témavezetők: Dr. Kondorosy Előd egyetemi docens, CSc
Elfogadásra javaslom (igen / nem) (aláírás)
Dr. Müller Tamás tudományos munkatárs, PhD
Elfogadásra javaslom (igen / nem)
(aláírás)
A jelölt a doktori szigorlaton ………% -ot ért el Keszthely, 2008.
Szigorlati Bizottság elnöke Az értekezést bírálóként elfogadásra javaslom:
Bíráló neve: igen / nem
Bíráló neve: igen / nem
A jelölt az értekezés nyilvános vitáján……….% -ot ért el Keszthely 2009.
Bíráló Bizottság elnöke A doktori (PhD) oklevél minősítése
Az EDT elnöke
4
T
ARTALOMJEGYZÉKKIVONAT ... 8
ABSTRACT... 9
AUSZUG ... 10
1. BEVEZETÉS ... 11
2. SZAKIRODALMI ÁTTEKINTÉS ... 13
2.1 A fűrészlábú szöcskék általános bemutatása és rendszertani áttekintésük... 13
2.2 A Saga fajok alaktana... 15
2.2.1 Fajcsoportok ... 15
2.2.2 A vizsgálatokban szereplő Saga fajok részletes morfológiai jellemzése ... 17
2.3 A Saga fajok tojásainak alaktana (ootaxonómia) ... 24
2.4 Színezet, környezethez való alkalmazkodás ... 25
2.5 Elterjedés ... 26
2.5.1 Égtáji elterjedés ... 26
2.5.2 Magassági elterjedés ... 26
2.5.3 Elterjedést befolyásoló tényezők ... 28
2.6 Egyedsűrűség ... 29
2.7 Aktivitás, napi ritmus és mozgás ... 30
2.8 Szaporodás ... 31
2.8.1 Párosodás ... 31
2.8.2 A Saga fajok kariogámiája ... 35
2.8.3 Wolbachia baktérium jelentősége a szűznemzés kialakításában ... 37
2.9 Embrionális és posztembrionális egyedfejlődés... 38
2.9.1 Tojásrakás ... 38
2.9.2 Embrionális fejlődés, nyugalmi állapot ... 39
2.9.3 Posztembrionális egyedfejlődés ... 43
5
2.10 Táplálkozás, prédaszerzés ... 44
2.11 Természetes ellenségek ... 48
2.12 Hangadás ... 49
2.12.1 A bioakusztika jelentősége, Orthoptera fajok hangadása ... 49
2.12.2 A hang megjelenítésebioakusztikai vizsgálatokban ... 50
2.12.3 A Saga fajok hangadása ... 51
3. CÉLKITŰZÉSEK ... 53
4. ANYAG ... 54
4.1 Begyűjtés és tartás ... 54
4.2 A vizsgálatokhoz használt példányok részletes adatai ... 54
5. MÓDSZER ... 55
5.1 A vizsgált példányok elhelyezése és tartási körülményei ... 55
5.2 Elterjedés ... 56
5.3 Fenológiai vizsgálatok ... 56
5.4 Tojásmorfológiai és tojásrakási vizsgálatok... 56
5.5 Bioakusztikai vizsgálatok ... 57
5.5.1 A hangfelvétel eszközei és körülményei ... 57
5.5.2 Bioakusztikai terminológia ... 58
5.6 Genetikai vizsgálatok ... 60
5.6.1 DNS kivonása, PCR és szekvenálás ... 60
5.6.2 Filogenetiai analízis ... 61
5.6.3 Morfológiai vizsgálatok a S. c. campbelli, S. c. gracilis alfajokon és a S. pedo fajon ... 63
5.6.4 A hibridizáció lehetőségének vizsgálata ... 65
5.7 Kiegészítő vizsgálatok: A S. pedo predátorok általi terjedésének vizsgálata és a kezdeti diapauza feloldása ... 67
6. EREDMÉNYEK ... 69
6
6.1 Elterjedés ... 69
6.1.1 A S. pedo magyarországi élőhelyei ... 69
6.1.2 Újonnan felfedezett lelőhelyek ... 75
6.2 Tojásrakási, tojás- és lárvamorfológiai vizsgálatok ... 77
6.3 Bioakusztikai vizsgálatok ... 80
6.3.1 A S. rammei faj éneke ... 80
6.3.2 A S. c. campbelli és S. c. gracilis alfajok énekének összehasonlítása ... 85
6.6 Genetikai vizsgálatok ... 86
6.6.1 Filogenetikai vizsgálatok... 86
6.6. Kiegészítő vizsgálatok ... 94
6.6.1 A S. pedo – S. c. campbelli – S. c. gracilis genotípusok morfometriai összehasonlítása ... 94
6.6.2 Ragadozók általi terjedés vizsgálata ... 96
6.6.3 A S. pedo és S. rammei fajok mesterséges keltetése ... 96
7. MEGVITATÁS ... 99
7.1 Elterjedés ... 99
7.2 Tojásmorfológiai és fenológiai vizsgálatok ... 100
7.3 Bioakusztikai vizsgálatok ... 101
7.3.1 A S. rammei faj hívóéneke ... 101
7.3.2 A S.c.campbelli és S. c. gracilis alfajok hangtani vizsgálata ... 102
7.4 A genetikai vizsgálatok értékelése ... 103
7.4.2 Hibridizáció lehetőségének vizsgálata a kongenerikus fajokkal ... 105
7.5 Kigészítő vizsgálatok ... 106
7.5.1 Tojásfejlődési vizsgálatok ... 106
7.5.2 Ragadozók általi terjedés vizsgálata ... 107
8. ÖSSZEFOGLALÁS ... 108
9. KÖVETKEZTETÉSEK ÉS KITEKINTÉS ... 114
10. ÚJ TUDOMÁNYOS EREDMÉNYEK ... 116
7
11. NEW SCIENTIFIC RESULTS ... 117
12. IRODALOMJEGYZÉK ... 118
13. KÖSZÖNETNYILVÁNÍTÁS ... 135
14. MELLÉKLETEK ... 136
14.1.1 A disszertáció témakörében megjelent publikációk ... 136
14.1.2 A disszertáció témakörén kívül megjelent publikációk ... 138
14.2 A vizsgálatokban felhasznált példányok befogására vonatkozó engedélyek ... 139
12.3 : A vizsgálatokban szereplő példányok részletes adatai ... 145
8
Kivonat
A fűrészlábú szöcskék (Saga spp.) biológiája, különös tekintettel a Saga pedo (PALLAS, 1771) fajra
A szerző dolgozatában a Saga nem képviselőinek biológiájával foglalkozik, kiemelve az egyetlen hazánkban is élő, védett S. pedo fajt. A szerző a faj magyarországi lelőhelyeinek eddigi legteljesebb jegyzékét állítja össze, köztük egy új lelőhellyel. Elsőként közli a fűrészlábú szöcske egyes lárvastádiumait jellemző fenológiai bélyegeket (femur és ovipositor hossz), megállapítja, hogy a faj imágóvá alakulását hat lárvastádium előzi meg. Ootaxonomiai vizsgálataiban a S. rammei és a S. c. campbelli tojására vonatkozóan elsőként közöl, a S. pedo fajra vonatkozón pedig pontosítja a korábbi morfometriai adatokat (tojás hossz- és keresztmetszet, valamint micropyle szám). A tojás keresztmetszetének változásán követi nyomon a S. rammei és S. pedo fajok embriógenezisét. A szerző szonogramos, oszcillogramos, és spektrogramos leírást közöl a S. rammei faj énekéről, mely alapján megállapítja, hogy a faj akusztikus szignálja amplitúdómodulált, ún. szokványos éneke a többi fajétól jól megkülönböztethető. Megállapítja, hogy a S. c. campbelli és S. c. gracilis alfajok éneke akusztikai bélyegek alapján egymástól elkülöníthető. A fajok ivartól függetlenül az első pár combon és lábszáron található tüskék, valamint az ugróláb (maximális) szélessége alapján főkomponens analízissel is elhatárolhatóak egymástól. Mitokondriális és nukleáris alapú markerek ezen különbséget alátámasztják, összegzésként a földrajzilag is elkülönülő alfajok faji szintre emelését javasolja. Megállapítja, hogy a S. pedo faj kialakulása a vele azonos haplotípust képező S. c. gracilis alfajból valószínűsíthető. A kládok alapján új fajcsoportokat állít fel, a természetes rokonsági viszonyokat tükrözendő. A szerző vizsgálja esetleges hibridek létrejöttét a S. pedo és a S. c.
campbelli párzásából, s megállapítja, hogy a vizsgált tojások apai örökítő
anyagot nem tartalmaznak.
9
Abstract
Biology of the saw-legged bush crickets (Saga spp.) focusing on Saga pedo (PALLAS, 1771)
In the present survey, the author is dealing with the biology of the Saga species, focusing on S. pedo, the only protected representative of the genus living in Hungary as well. The author compiles an up-to-date habitat list of this species, with a description of a new one in Hungary. He gives first phenologic characters (length of femur and tibia) describing each larval stadia of S. pedo, furthermore determines that a S. pedo imago develops though six larval stadia.
In ootaxonomic investigations, data on the morphology of the eggs (length, width and number of micropyles) of S. rammei and S. c. campbelli were firstly given; moreover refined values for S. pedo were transmitted. The embryogenesis was traced by the changing of the diameter of the eggs of S. rammei and S. pedo.
A detailed description of the spectrographic, sonographic and oscillographic features of the calling song of Saga rammei is presented for the first time. The acoustic signal of the species is amplitude modulated and it is well distinguishable from those of the congeners. Furthermore, the songs of S. c.
campbelli and S. c. gracilis were found to be significantly different. Also, the
two subspecies can be distinguished by PCA the spines of the fore leg on the
tibia and femur and by the maximum width of the hind femur independently
from the gender. This was supported by the information gained from
mitochondrial and nuclear markers, thus the author suggests handling the two
geographically isolated subspecies as separate species. Concerning S. pedo, the
author determines, that it most likely developed from S. c. gracilis a congener
that shares the same haplotype. According to the clades, he suggests new species
groups, being more related to the natural relationships. The author investigates
the possibility of hybridization between S. pedo and S. c. campbelli, resulting
from studies confirming that no paternal DNA in the examined offspring eggs
are present.
10
Auszug
Biologie der Sägeschrecken (Saga spp.) im Fokus an Saga pedo (PALLAS, 1771)
In seiner Arbeit der Autor befasst sich mit der Biologie der Sägeschrecken, mit besonderer Beachtung an S. pedo der einzige geschützte Art seines Genus der findet sich auch in Ungarn. Der Autor stellt eine Liste von den ungarischen Fundorten dieses Artes zusammen, mit Beschreibung eines neues aus Ungarn.
Er zeigt die phänologische Merkmale jeden Larvastadium an, und stellt fest, dass S. pedo geht durch 6 Häutung um Imago zu sein. In ootaxonomischen Untersuchungen erteilt er erstens morphologische Merkmale (Länge, Breite und Zahl der Mikropülen) für die Eier von S. rammei und S. c. campbelli sowie gibt genauer Angaben für S. pedo. Der Autor verfolgt die Embryogenese von S.
rammei und S. pedo durch die Veränderung der Breite der Eier. Für den Art S.
rammei eine Beschreibung mit Sonogramm, Spekrogramm und Oszillogramm wurde gegeben; feststellbar ist, dass der Signal ist Amplitudenmoduliert und sein übliche Gesang ist artspecifisch. Der Gesang von S. c. campbelli und S. c.
gracilis lässt sich durch die von dem Autor untersuchte akustische Merkmalen
gut unterscheiden. Weiterhin, die zwei Unterarten können durch die Zahl der
Dornen an den ersten Beinpaaren und der maximalen Breite der Hinterschenkel
auch mit PCA Analysis unabhängig vom Geschlecht unterscheiden werden. Da
die von genetischem Markieren erwiesen Unterscheiden diese Ergebnisse
unterlegen, der Autor vorschlägt die taxonomischen Status der genannten
Unterarten von Unterart zu Arten zu verändern. Anschliesslich der Abstammung
von S. pedo, der Art soll sich von S. c. gracilis getrennt werden. Sowie
entschprechend der Kladen der Autor stellt neue Artengruppen auf, schon die
natürliche Verwandtschaften zeigt. Weiterhin, die Möglichkeit der Hybridisation
von einer Paarung zwischen S. pedo und S. c. campbelli wurde geprüft, mit der
Ergebnis, dass die Nachfolger Eier untersucht enthielten keinen DNA
väterlichseits.
11
1. Bevezetés
A természetvédelmi biológia napjainkban egyre jelentősebb lesz, elsősorban a fajok eltűnésének gyorsuló üteme miatt, főleg az emberi tevékenység következtében (Soulé 1985).
A rovarfajok - különösen a védett és veszélyeztetett fajok - életmódjának, biológiájának megértése a biológiai sokszínűség (biodiverzitás) megőrzéséhez elengedhetetlen, melyek sajnos jelenleg a kívánatosnál kevesebb figyelmet kapnak (Standovár & Primack 2001).
Az egyenesszárnyúak rendjének (Orthoptera) első képviselői feltehetőleg a karbon időszak közepén jelentek meg földünkön (Hennig 1981). E viszonylag kis számú rend közel 25 000 fajt számlál (Eades & Otte 2009), melyből Európában 974, (Heller et al. 1998) hazánkban pedig mintegy 120 faj él (Rácz 1998).
Maga a rend két alrendre [a csoportok rend szintre emelése kérdéses (Rácz 2001)] tagolódik: tojókampósokra (Caelifera) és a tojócsövesekre (Ensifera). Az utóbbi alrendnek része a szöcskealakúak öregcsaládja (Tettigonioidea), melynek hazai képviselői között viszonylag nagy a védett fajok száma. Ennek oka, hogy a környezeti feltételekre és zavarásra nagyon érzékenyek, speciális élőhelyhez kötődnek, sokuk - különösen a rövid szárnyú (brachypter) fajok - nehezen és lassan képesek élőhelyet váltani és menekülni. Érzékenyen reagálnak a környezeti változásokra, emiatt számos szerző (Kriegbaum 1989, Andersen et al. 2001, Szövényi 2002) hangsúlyozza jelzőfajként való alkalmazhatóságukat. A hazai egyenesszárnyúak jó indikátorfajok gyepek természetességének megállapításához (Kisbenedek & Báldi 2000, Kenyeres & Bauer 2001), továbbá közepesen nagytermetű és viszonylag könnyen határozható fajok lévén a Nemzeti Biodiverzitás- monitoring Rendszer minimális programjába is javasoltak (Kisbenedek 1997).
Az egyenesszárnyúak rendjébe sorolt fűrészlábú szöcskék (Saginae) Eurázsia legnagyobb termetű rovarfajai közé tartoznak. Populációik speciális, száraz, meleg élőhelyekhez kötődnek, ezeket szinte kizárólag hegységek sziklagyepein találják meg.
Előfordulásuk szórványos, populációik - különösen imágó korban - kis egyedszámúak (Kaltenbach 1970). Röpképtelen, kis vagilitású állatok. Táplálkozásuk az egyenesszárnyúakon belül nem szokványos: kizárólag ragadozó életmódot folytatnak, egyúttal élőhelyük csúcsragadozó rovarai.
Az eurázsiai fűrészlábú szöcskefajok ritkaságuk ellenére nem védettek, egy kivételtől eltekintve. A fűrészlábú szöcske (Saga pedo PALLAS, 1771) a 13 fajt számláló nem egyetlen magyarországi képviselője, egyúttal legnagyobb termetű rovarfajunk. A fűrészlábú szöcske
12
hazánk NATURA 2000-es, védett rovarfaja; eszmei értéke 50 000 Ft (13/2001.(V.9.) KöM rendelet). Szerepel több ország vörös könyvében, a CORINE-listán, és a Berni egyezmény II.
függelékében is. Az IUCN listáján a sérülékeny (vulnerable) kategóriába tartozik, mivel kis egyedszámú populációi könnyen kipusztulhatnak (Kisbenedek, 1997). E legészakabbra húzódó faj azonban elterjedési területén már csak foltokban, többnyire más okból is védettség alatt álló szubmediterrán élőhelyeken fordul elő. Ritkaságán kívül védettségének oka ezeken kívül még érdekes biológiája: A S. pedo Saga nemnek egyetlen olyan ismert képviselője, amelynek kizárólag szűznemzéssel szaporodó nőstény egyedei ismeretesek. Elterjedési területe a Saga nem képviselő közt a legnagyobb, mely az ivaros fajok együttes areájának többszöröse.
A faj és rokonainak biológiája, evolúciós eredete a tudomány számára számos kérdést vet fel. Ugyanakkor a genus egyes tagjait rendszertani (taxonómiai) szempontból sem hiábavaló vizsgálni a mai kor lehetőségeivel (pl.: molekuláris genetika), különös tekintettel az alfaji kategóriákra. A védett S. pedo nagyobb termete okán is modellfajként szolgálhat a védett rovarokra alkalmazható megőrzési stratégiák kidolgozásához, hasonlatosan egyes hüllőfajok példájához, mint amilyen például a rákosréti vipera (Vipera ursinii rakosiensis) megőrzési programja. A mesterséges keltetés lehetőségeinek vizsgálata, s a több éves fejlődés lerövidítése megteremtheti nemcsak a védett fűrészlábú szöcske, de az ugyancsak érdeklődést felkeltő, vele egy genusba tartozó (kongenerikus) fajtársainak oktatási célú bemutatását, megismerését, s nem utolsó sorban a faj kipusztult élőhelyekre való visszatelepítésének lehetőségét.
13
2. Szakirodalmi áttekintés
2.1 A fűrészlábú szöcskék általános bemutatása és rendszertani áttekintésük
A fűrészlábú szöcskék Eurázsia legnagyobb termetű rovarfajai. Őseik valamikor a Jura időszak közepén jelenhettek meg más olyan csoportokkal együtt, melyek ma Eurázsia, Ausztrália és Közép-Amerika száraz élőhelyeit lakják (Harz 1969, Gorochov 1995). A fürgeszöcskék (Tettigoniidae) családjába tartozó fűrészlábú szöcskék képviselői mind morfológiailag, mind pedig életmódjukat tekintve jól elkülönülnek a többi szöcskefajoktól;
csupán a kizárólag ausztráliai hasonló fajokat magába foglaló Austrosaginae alcsalád fajai állnak még viszonylag közel hozzájuk. A Saginae alcsaládba tartozó nemek képviselői - az általában vegyes táplálkozású többi szöcskefélével ellentétben - kizárólag ragadozó életmódot folytatnak. Megjelenésüket illetően az alcsalád minden fajára jellemző az elfogott zsákmány megtartására szolgáló, legalább az első pár lábon meglévő tüskesor.
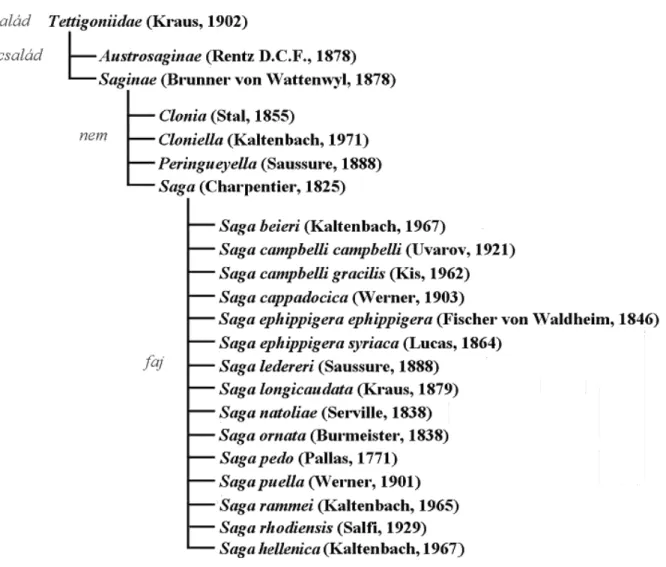
A Saginae alcsalád (Eades & Otte 2009) négy nemet foglal magába, melyek két egymástól távol eső régióban terjedtek el. A fajok döntő többsége - azaz mintegy 30 faj és alfaj - Afrika Szaharától délre eső részén él. Ezek a Clonia; Cloniella; valamint a Peringueyella genusok képviselői. A palearktikum nyugati részén csupán egyetlen nem, a Saga képviselő terjedtek el. A fűrészlábú szöcskék mindegyik faja - ellentétben a többnyire szárnyas afrikai rokonaikkal – röpképtelen.
A Saga nem (1. ábra) összesen 13 fajt számlál, melyek közül öt (S. natoliae, S.
rammei, S. campbelli campbelli és S. campbelli gracilis, S. hellenica, S. pedo) fordul elő Európában, míg a többi Kis-Ázsiában él – ideértve S. rhodiensist is, mely bár Rhodos szigetén is előfordul, elterjedési területének zöme az Anatóliai-félszigeten található (Kaltenbach 1970).
14
1. ábra: A Saga fajok rendszertani helyzete a fürgeszöcskék alcsaládjában
15
2.2 A Saga fajok alaktana
A fűrészlábú szöcske fajok elkülönítése sokáig problémát jelentett. Burr et al. (1923) még így ír: „Kevés olyan palearktikus csoport van, mint a Saga nem, amely rendszertanilag ennyire reménytelenül összekuszálódott‖. Ennek oka, hogy a korai szerzők fajleírásai nem kielégítőek, a korai határozókulcsok (Krauss 1878, Saussure 1888) összehasonlító táblái nem teljesek, valamint olyan bélyegekre építenek, melyek taxonómiai használhatósága kétséges. Werner (1905) például az első combon lévő tüskék számára és a változékony, bizonytalan színezet-különbözőségekre építette határozókulcsát; Ramme (1951) határozása már valamivel részletesebb, ám a nagyon kevés rendelkezésre álló példány (sok típuspéldányhoz nem tudott hozzáférni), valamint a rendelkezésre álló irodalom feldolgozatlansága miatt nem volt teljes. Kaltenbach (1967) már 992 vizsgált példány alapján dolgozta ki a Saga fajok elkülönítésére szolgáló morfológiai határozóbélyegeket, melyek alapján (mind a hímekre, mind pedig a nőstényekre vonatkozó) határozókulcsa 13 fajt különít el.
2.2.1 Fajcsoportok
A Saginae filogenetikailag nagyon ősi alcsalád (Burr et al. 1923, Uvarov 1929, Matthey 1941), képviselőit faji szinten csak nemrég sikerült elkülöníteni. Ennek oka, hogy nincs általános érvényű stabil morfológiai bélyeg, hanem azok egyik fajnál stabilak, míg egy másik esetében nagyon változóak lehetnek, és így ezek alkalmazhatósága a határozáshoz az egymással összehasonlítandó fajoktól függ (Kaltenbach 1967). Rokonsági viszonyokat illetően először Kaltenbach (1967) sorolta Saga fajokat fajcsoportokba (Artengruppe) - elsődlegesen a hímek alaktani bélyegei alapján. Ezek a következők:
1. Natoliae-csoport: Valószínűleg az őstípushoz a legközelebb állók csoportja, nagy és közepesen nagy méretű fajokkal. Ide tartoznak a S. natoliae, S. rhodiensis és S. ephippigera fajok. Elterjedési területük Nyugat-Ázsia és a Balkán, feltételezett géncentrumuk a Földközi- tenger keleti partvidéke. A Kaukázusban a S. ephippigera karcsúbb, kisebb termetű, pigmentáltabb változata gyakoribb, azonban ilyen formák mindenhol megtalálhatók, nincs olyan határozóbélyeg, mely alapján egy kaukázusi ephippigera alfaj elkülöníthető. A S.
ephippigera syriaca alfaj elterjedési területe az (anatóliai) Tigris és Eufrátesz folyók közti területtől Mezopotámián át Szíria északi részéig húzódik. A S. e. syriaca és a S. e.
16
ephippigera keveredési területe a Kurd-hegység vonulatai, és az Amanus-hegység déli része.
A két alfaj a fej szélességén kívül elkülöníthető még a hátsó láb keresztmetszetének és hosszának hányadosa alapján is (a határérték nagyjából 7,9 – ez alatti érték a többi bélyeg tükrében S. e. syriaca alfajra utal) (Kaltenbach 1967)..
2. Puella-csoport: Tagjai a S. hellenica, S. cappadocica, S. rammei, S. puella, S.
campbelli. A kicsit a S. natoliae-hoz hasonlító S. hellenica kivételével e fajok az előző fajcsoporttól jól elkülöníthetőek. A hímek szárnya kevésbé redukált, mint a másik két csoport esetében, a hímek cercus-a középső részén a legvastagabb. Elterjedési területük Anatólia és a Balkán. A S. campbelli faj két alfaját illetően (Kaltenbach 1967) szerint a kisebb elterjedési területtel rendelkező S. c. campbelli a szekunder alfaj, mivel a S. c. gracilis élőhelye egy jóval nagyobb területet ölel fel.
3. Ornata-csoport: A S. ornata, S. longicaudata, és S. beieri fajok képviselnek - valamelyest közel áll a Natoliae-csoport fajaihoz; a homlok dorozmás, ám megjelenésük karcsúbb, és a lábakon is kevesebb tüske található. Elterjedési centrumuk a Földközi-tenger keleti partvidéke, és Délkelet-Anatólia, feltételezett elterjedési központjuk Palesztina.
4. A Saga pedo fajt Kaltenbach (1967) különválasztotta a többi fajtól, mivel a fajcsoportokat elsősorban a hím egyedek bélyegei alapján állította fel. Habitusa leginkább a S.
c. campbelli és S. c. gracilis alfajokéhoz hasonlatos (bővebben lásd. 3.2.2 fejezet).
Feltételezett kialakulása helyéül a Kaszpi-tenger és a Fekete-tenger közötti területet jelölte meg, s kizárta a balkáni területeket, mivel egyfelől a Balkán déli része e faj elterjedési határa, másfelől populációinak egyedsűrűsége sem itt a legnagyobb.
17
2.2.2 A vizsgálatokban szereplő Saga fajok részletes morfológiai jellemzése
Saga pedo Pallas 1771
Többnyire közepes termetű (53-75 mm), karcsú faj. Kizárólag nőstény egyedei ismeretesek, ám - bár Kaltenbach (1967) a korábban talált "hímeket" határozási hibákra vezette vissza - ma is felbukkannak hímekről szóló értesülések fényképes dokumentációval (Baur et al. 2006), erről azonban nagy
felbontású fotó vagy preparátum - kétségeket kizárandó - nem hozzáférhető.
A homlok sima, rendszerint két barna homlokponttal. A pronotum-ot a mesonotum-tól vékony szegély választja el, mely csak néha kis mértékben felfelé hajló. A cercus-ok henger alakúak, többnyire rövid csúcsi foggal. A tojócső enyhe ívben az alapjától kiindulva felfelé hajló.
Némileg hasonlít a S. campbelli fajra, ám annak tojócsöve lényegesen rövidebb, egyenes, csak utolsó harmadában felfelé hajló. A S. campbelli faj ezen kívül többnyire karcsúbb termetű, és a hátsó végtagok is hosszabbak. Továbbá, amennyiben a nyakpajzs oldala fehér, mindenképpen ez utóbbi fajról van szó.
Korábban a fajt Anatóliából is jelezték, ám az itt honos esetleg hasonló fajoktól is (S.
beieri, S. cappadocica, S. longicaudata) jól elkülöníthető a szubgenitális lemez kivágása, a tojócső sima alsó széle, és a lábakon található tüskék száma alapján (Kaltenbach 1967).
Saga campbelli campbelli Uvarov 1921
A legkisebb testhosszal (49,5-68,5 mm) rendelkező Saga faj, eléggé vékony, hosszú, karcsú végtagokkal. A homlok sima, rendszerint csak két pár ponttal, melyek közül a felső pár erőteljes, barna színezetű, az alsó kettő tompa, gyakran jelentéktelen. A pronotum hátsó széle a hímeknél enyhén, a nőstények esetében pedig alig észrevehetően felfelé hajló. A hímek
2. ábra: Saga pedo(a szerző felvétele)
18 szárnyának (oldalnézetben) felső pereme még a végnyúlvány előtt lefelé ível, és kimélyed. A ciripelőmező erősebben kiöblösödő és csaknem mindig ráfut valamelyest az elytra felső szegélyére. A fogak száma az első két pár lábon, a lábszáron átlagban 10,7-13 a combon 12,1- 14,9 egy sorra vonatkoztatva. A faroktoldalékok a hímek esetében változó alakúak, azonban leginkább a hellenica
típusnak felelnek meg. Ezek csúcsi része befelé hajló cercus-foggal rendelkezik, kisebb a S.
hellenica és rammei fajokénál. Hasonlóan változékony a nőstények faroktoldalékának formája is. A nőstény ivarnyílás alatti lemeze a csúcsánál mérsékelten mélyen kivágott. A tojócső karcsú, egyenes, csak a csúcsi harmadában hajlik valamelyest felfelé. Alapszínezete változékony, fűzöld, sárgásbarna olivás beütéssel, szürkésbarna, nagyon ritkán vörösesbarna.
Kevésbé fordulnak elő közöttük tarka, barna háti foltokkal rendelkező példányok, sokkal gyakoribbak viszont az oldalt fehér sávval díszített egyedek (Kaltenbach 1967).
Saga campbelli gracilis Kis 1962
Az alfaj eredetileg S. italica gracilis Kis 1962 néven került leírásra; névrevízió (Kaltenbach 1967) után Saga italica (Saga campbelli) taxon Saga campbelli gracilis-re változott - az alfaji kategóriát továbbra is megtartva. Csaknem minden bélyege megegyezik a törzsfajjal. A testhossz 49,5-63,3 mm. A fogak száma az első két pár lábon: a combon 9,9- 11,5 (S. c. campbelli: 10,7-13); a lábszáron 10,5-12,4 (S. c. campbelli: 12,1-14,9) egy sorra vonatkoztatva (Kaltenbach 1967). Kis (1962) szerint jelentősebb alfaji különbségek a következők:
- Az egyik leglényegesebb a lábak szélessége és hosszának viszonya: míg a S. c.
gracilis esetében az első végtag nagyjából 7-8-szor hosszabb annak legnagyobb szélességénél, addig a S. c. campbelli ez csak 5-5,5 szeres.
- A harmadik végtag a S. c. gracilis-nél 19,5-22-szer, a S. c. campbelli-nél pedig 11-14- szer hosszabb a legnagyobb szélességnél.
- A S. c. campbelli nőstényekre jellemző négy, háromszögletű sötét folt a S. c. gracilis
3. ábra: Saga c. campbelli (a szerző felvétele)
19
esetében hiányzik a nyakpajzsról (tergit) - ellenben két sárgásfehér csík fut a potroh mentén.
Kis (1962) szerint a S. italica (=S.
campbelli) az egykori Jugoszlávia területéről, Albániából, Görögországból, és Itália déli területeiről ismeretes. Jacobson &
Bianchi (1905) említést tesz az egykori cári Oroszország délnyugati részén való előfordulásáról is. Kis (1962) szerint ez, valamint a dobrudzsai előfordulása e fajnak arra utal, hogy eredeti előfordulása
szélesebb területet ölelt fel, mint a mai - később azonban elterjedése beszűkült, a közbülső területekről kipusztult. Kis (1962), Bureš & Pešev (1958) adataira hivatkozva megjegyzi, hogy ezen faj Bulgáriából hiányzik, s ma már csak foltokban maradt fenn Dobrudzsában és a (volt) Szovjetunió délnyugati területein. Véleménye szerint valószínűleg a dobrudzsai Saga italica populáció az ősi formából szakadt le, s az enyhébb környezeti és klímaviszonyok hatására a S. italica gracilis alfaj már régen létrejött, és rögzült. Minden valószínűség szerint a két alfaj területei már nem érintkeznek egymással, s így nem található meg a határzónákra általában jellemző átmeneti forma sem. A dél-ukrajnai területek populációinál a törzsalak ismertetőjegyei jól megfigyelhetőek, s ebből arra lehet következtetni, hogy itt nem alakult ki külön alfaj. (Kis 1962).
Saga rammei Kaltenbach 1965
A fűrészlábú szöcskék között kis-és közepes testmérettel (45-71 mm) rendelkezik, nősténye leginkább a S. cappadocica-ra hasonlít, ám nála zömökebb, erősebb végtagokkal. A homlok sima, rajta három – kivételes
4. ábra: Saga c. gracilis (a szerző felvétele)
5. ábra: Saga rammei (a szerző felvétele)
20
esetben kettő vagy egy – pár jellegzetes barázdával. A pronotum hátsó széle a hímeknél erőteljes, vastag, a nőstények esetében nagyon enyhén felfelé hajló. A hímek szárnyának (oldalnézetben) konvex felső pereme jól fejlett végnyúlványban végződik, amely a S.
hellenica vagy S. cappadocica fajéhoz viszonyítva keskenyebb. A ciripelőmező simább, mint az előző két faj esetében, és csak alig terjed tovább a szárny felső szegélyénél.
A fogak száma az első két pár lábon a combon 9,4-11,5 (átlagban: 9,5-10,4) a lábszáron 10,4- 12,4 (átlagban: 11-12) egy sorra vonatkoztatva. A faroktoldalékok (cercus) a hellenica típusnak felelnek meg, distalis végük felé erőteljesebben elvékonyodnak, a csúcsi részük befelé hajlik.
A nőstény faroktoldalékai orsó alakúak, alapjukon kiszélesednek. A nőstény ivarnyílás alatti lemeze mélyebben, kereken kivágott, némiképp hasonlít a S. cappadocica sima lemezéhez. A tojócső viszonylag rövid, meglehetősen egyenes, csupán a csúcsi harmadban felfelé görbült.
Megkülönböztetése mindenekelőtt a S. hellenica és campbelli fajoktól fontos, hiszen ezek elterjedési területe érintkezik a S. rammei fajéval.
A hímek alapján a fajok elhatárolása viszonylag egyszerűbb, mivel az elytra kialakításában a fajok mindig jelentősen különböznek.
A nőstények alapján való elkülönítés már nem ilyen egyértelmű: a S campbelli egyedei többnyire karcsúbbak: A potroh a S. campbelli esetében 17,5-24-szer hosszabb, mint annak legnagyobb szélessége; a S. rammei-nél viszont ez a szám 12,4-17, de nagyritkán elérheti a 19-et is. Ezen túlmenően a fogak száma a lábon, valamint a tojócső hossza segíthet még az elkülönítésben. A S. pedo-val összehasonlítva tojócsövük hosszáról és alakjáról elkülöníthetők (Kaltenbach 1967).
Saga natoliae Serville 1839
Közepes- és nagyméretű (51,5-88,6 mm) példányokkal rendelkező Saga faj, erőteljes fejjel és a végtagokkal. A fej dorozmás, az egyedek csaknem felénél homlokfoltokkal tarkított. A pronotum hátsó szegélye felfelé hajló (a nőstények esetében ez kevésbé kifejezett).
A hímek szárnya konvex felső szegéllyel és gyengén fejlett véglebennyel rendelkezik. A ciripelőmező sosem
6. ábra: Saga natoliae ( a szerző felvétele)
21
ér túl a szárny felső szegélyén. A hímek cercus-a szarvszerűen hajló, közepén kiszélesedő, csúcsi részén a fog befelé és enyhén lefelé hajlik. A tojócső erősen fejlett, egyenes; csak az utolsó harmadában görbül valamelyest felfelé, majdnem mindig rövidebb, mint a pronotum hosszának háromszorosa. Közel rokon faj a S. rhodiensis, melytől a nőstények a cercus-ok és a subgenitalis lemez kivágása, a hímek pedig a cercus-ok és a szárny alakja alapján jól elkülöníthetőek. A másik rokon faj, a S. ephippigera ezen bélyegek, valamint az első lábon található tüskék alapján ugyancsak jól elhatárolható. A natoliae csoport fajaira emellett jellemző a hímek szárnyának alapján található fekete folt, és a dorozmás, pontozott pofa és homlok (Kaltenbach 1967).
Saga ephippigera ephippigera Fischer de Waldheim 1846
A legnagyobb Saga faj, ugyanakkor a legváltozatosabb méretű is, közepes termetű példányai is ismeretesek;
testhossza 56 és 113 mm között változhat. A hím előhátának hátsó szegélye erősen vagy mérsékelten felfelé hajló; nőstények esetében ez kevésbé kifejezett. A homlok dorozmás felületű, esetenként homlokfoltokkal. A hímek cercus-a
vastag, belső felülete egyenes vagy enyhén kiöblösödő, csak a csúcs előtt vékonyodik el valamelyest. A hímek szárnya erősen redukálódott; a ciripelőmező sosem ér túl a szárny felső szegélyén. A tojócső erőteljes, hossza a testhossztól függetlenül változó hosszúságú (Kaltenbach 1967)..
Saga ephippigera syriaca Lucas 1864
Nagyon hasonló a másik alfajhoz, ám annál tömzsibb és robusztusabb megjelenésű.
Testhossza 71- és 112,5 mm között változhat. A homlok nagyon széles (11-17mm). A nyakpajzs, különösen a nőstények esetében nagyon gyakran csak enyhén felfelé hajló. A hátsó comb, hosszához viszonyítva a vastagabb, mint a névadó alfaj esetében. Sötét tergitmintázatú egyedek ritkábban fordulnak elő, csak a magasabban fekvő régiókban
7. ábra: Saga e. ephippigera (Amir Weinstein felvétele)
22
találhatóak kifejezetten sötét színezetű példányok is. A S. e. ephippigera alfajtól elkülönítése sokszor elég nehéz és csak számos példány figyelembevételével lehetséges.
Saga hellenica Kaltenbach 1967
Kis- és közepesen nagyméretű (48-78 mm) Saga faj. A homlok felülete sima, elvétve előfordulhat finom pontozás. Az állatok túlnyomó részénél két vagy három pár barnásfekete homlokpont található. A pronotum hátsó szegélye a hímek esetében erős, felfelé hajló – ám kisebb mértékben mint a S. natoliae esetében; nőstény példányok esetében ez enyhébb, nem kifejezett. A hím elytra viszonylag nagy, oldalnézetben konkáv felfutású felső szegéllyel. A véglebeny jól fejlett, csúcsukon háztetőszerűen zárulnak össze.
A cercus-ok a hímek esetében konvex belső széllel rendelkeznek, a középső részükön
megvastagodottak. A csúcsi fogak befelé és valamelyest előre hajlanak. A nőstény cercus-ai orsó formájúak. A tojócső, különösen annak alapi részén, erőteljes, utolsó harmadában alsó felén enyhén felfelé hajlik. Hasonló fajoktól (pl. S. natoliae, S. campbelli, S. rammei) a hím elytra és a cercus-ok alakja alapján jól elkülöníthetőek. A nőstények esetében ezen bélyegeken kívül a tergitek mintázata, alakja használható az elkülönítéshez (Kaltenbach 1967).
Saga cappadocica Werner 1903
Kis- és közepes méretű (43,4- 64,5 mm) példányokkal rendelkező Saga faj. A homlok sima, néha egy pár homlokpont látható. A hímek esetében
8. ábra: Saga hellenica (a szerző felvétele)
9. ábra: Saga cappadocica (a szerző felvétele)
23
a pronotum hátsó szegélye erősen felfelé hajló, a nőstények esetében a mezonotum-tól csak egy gyenge szegély határolja. A hím elytra jól fejlett véglebenyekkel rendelkezik, a ciripelőmező nagy ívű, mindig túlér a szárny felső szegélyén. A hím cercus-ok belső oldala konvex, majdnem egyenes lefutású, középső részén a legvastagabb; a csúcsi fog befelé hajlik.
A nőstények cercus-a orsó vagy kúp formájú, alapi részén a legszélesebb. Habitusát tekintve a S. rammei fajhoz hasonlít, ám hímjei a szárnyak; nőstényei a homlokfoltok (a S. cappadocica esetében hiányoznak, vagy jelentéktelenek) alapján jól megkülönböztethetők tőle. A hasonló anatóliai fajoktól (S. puella, S. longicaudata) a hímek a szárny alakja, a nőstények pedig a tojócső alapján különböztethetők meg (a S. puella tojócsöve az alapjától kiindulva fogazott, a S. longicaudata esetében az ovipositor lényegesen hosszabb) (Kaltenbach 1967).
S. ornata Burmeister, 1839
Példányai közepesen nagyméretűek (57-80 mm). Karcsú termetű Saga faj, hosszú végtagokkal. Az első és középső lábak tüskéi alapjuknál keskenyek. A homlok pontozott vagy redőzőtt. A hím elytrája legfeljebb a
utóhát (metanotum) közepéig ér, a véglebeny alig felfedezhető. A ciripelőmező erősen ívelt és a szárny felső részén valamelyest túlnyúlik. A nőstény cercus-ai hengerformájúak, ám nagyon változékonyak. Közeli rokon faj a S. ledereri, melytől nőstények a lábszáron található fogak száma, a hímek a szárny kiképzése alapján határolhatók el (Kaltenbach 1967).
10. ábra: Saga ornata (Guy Haimovich felvétele)
11. ábra : Néhány Saga faj tojócsöve: 1: S. rammei, 2: S. cambelli, 3:S. hellenica, 4 S. ephippigera, 5: S.
ornata, 6: S.pedo (Kaltenbach 1967) nyomán
12. ábra : Néhány Saga faj ivarnyílás lemeze: 1: S. rammei, 2: S. campbelli, 3:S. hellenica, 4 S. ephippigera, 5: S. ornata, 6: S.pedo (Kaltenbach 1967) nyomán
24
2.3 A Saga fajok tojásainak alaktana (ootaxonómia)
A földbe lerakott Saga tojás többnyire barnás, esetenként zöldes színű, alakilag a Decticus, Antaxius, Anconotus, Thyreonotus nemek ovális, szimmetrikus tojásaival mutat kapcsolatot (Hartley & Warne 1972); kemény, átlátszatlan tojáshéjjal (chorion) rendelkezik.
A nagysága fajonként különböző, bár nem akkora mértékben, mint maguk a kifejlett állatok (Kaltenbach 1970). A szélesség és hosszúság aránya a mediális tengelynél 5:1, mely mérettől független; a tojásnak megkülönböztethető egy kisebb görbülettel rendelkező alapi, valamint egy csúcsi vége, ahol a spermiumok bejutását szolgáló képletek, az ún. tojáskapuk (mikropyle) zöme elhelyezkedik Sänger & Helfert (1994).
A Saga fajok tojása fajra jellegzetes. Hosszúsági méreteikről három fajnál először Kaltenbach (1970) adott ismertetést: S. ephippigera: 11,5-13 ; S. natoliae: 11-12 ; S. c.
campbelli: 8,5-9,2 ; S. pedo: 10-12 mm. Tömegadatot elsőként Hartley & Warne (1972) közölt: S. pedo: 40-60 mg.
Részletesebb leírást a tojás hosszának vonatkozásában Sänger & Helfert (1994) közölt az alábbi fajokra: S. pedo 9,9±0,3mm (n=74) ; S. natoliae 9,8±0,3mm (n≈100); S.
ephippigera 12,6±0,2mm (n≈100).
A S. pedo tekintetében továbbá Krištin & Kańuch (2007) közölt még hosszúsági és keresztmetszeti adatokat: hossz: 11,9 mm ; szélesség 3,8 mm ; tömeg: 41 mg.
A spermium bejutását szolgáló, tojáson található tojáskapuk (mikropyle) (Romoser &
Stoffolano 1994) számát illetően adatokat először Mazzini (1976) közölt, elsőként megállapítva, hogy a S. pedo tojásainak egy részén (4,4%, n=45) a tojáskapu és az azt körülvevő kráter hiányzik.
Ezt követően részletesebb adatokat Sänger & Helfert (1994) közölt:
mikropylék száma: S. ephippigera: 11,2±3,3 (min. 4, max. 27); S. natoliae: 8,9±2,9 (min. 4 max. 17); A S. pedo esetében a tojások 27%-ban hiányzik a mikropyle, átlagos számuk: 1,8
±1,8 (max.7). A tojáskaput körülvevő kráter átmérője: S. pedo és S. natoliae 25-30 µm ; S.
ephippigera 10-15 µm, maga az ennek mélyén ülő korong alakú mikropyle nyílás: S. pedo és S. natoliae 2-3 µm; S. ephippigera 1-2 µm.
25
2.4 Színezet, környezethez való alkalmazkodás
A Saga fajok színezete élő állapotban többnyire zöld, ritkábban barna vagy sárgászöld, valamint esetenként feketés foltokkal tarkított többnyire a tor, fej, és a potroh háti részén. A rajzolat többnyire örökletes, ám nem kis részben külső tényezők általi befolyás alatt is áll.
A színezet függ elsőként attól, hogy az állat melyik fajhoz tartozik. Így a S. natoliae és a S. pedo fajokra jellemző, hogy egyszínűek, többnyire zöldek, csak kivételes esetben fordulnak elő teljesen sárga vagy barna egyedek, ezzel szemben a S. rhodiensis fajnak barna példányai gyakoribbak. A S. rammei fajnak zömmel tarka mintázatú egyedei fordulnak elő.
Fekete foltokkal tarkított példányok a S. ephippigera ephippigera és S. ephippigera syriaca esetében a leggyakoribbak (Kaltenbach 1967).
A színezet és mintázat kialakulása, fejlettsége függ az életkortól is. Fiatal lárvákon szinte egyáltalán nem találunk mintázatot, idősebb lárváknál is csak ritkán. A S. natoliae esetében (Kaltenbach 1970) szerint az általában zöldessárgás imágókkal szemben az egyszínű zöld imágók általában gyengébbek, feltehetőleg a tirozin-anyagcsere zavarának következtében. Az idősödő imágókon a legtöbb fajnál megfigyelhető a fokozottabb pigmentáció megjelenése.
A hímek és nőstények is eltérnek: a S. rammei faj nőstényei tarkább mintázatúak; a S.
ephippigera és S. natoliae esetében viszont a hímek pigmentációja fokozottabb (Kaltenbach 1967).
Meghatározó hatása van az adott élőhelyen elérhető hőmérsékletösszegnek. A S. pedo Közép-Európában csaknem mindig zöld színű; Kaltenbach (1970) Alsó-Ausztriában 13 év alatt (n=52 lárva és imágó) csupán egyetlen szürkésbarna egyedet figyelt meg. Matthey (1941) egy érdekes, szürkéslilás példányt említ. Padewieth (1900) Zenggnél (mai Horvátország) nagy számban figyelt meg szalmasárga és barna egyedeket.
A Saga ephippigera esetében a pigmentáltság mértéke déli elterjedési területén kisebb, észak felé haladva viszont egyre nagyobb mértékű, melynek köszönhetően az északibb élőhelyeken élő populációk pigmentáltabb egyedei a nap hőjének jobb hőkihasználása miatt előnyben vannak kevéssé pigmentált társaikkal szemben (Kaltenbach 1970).
Fontos hatása van végül a környezetnek is. A Saga fajok többnyire alkalmazkodnak a környező növényzet alapszínéhez (Pantel 1886, Lang 1930, Ramme 1951, Nagy 1965).
Kaltenbach (1970) fogságban tartott Saga fajokon (S. c. campbelli és S. rhodiensis) is megfigyelte, hogy ezen állatok lárva és fiatal imágó korban néhány nap alatt teljesen, vagy
26
közel egyszínűek lesznek környezetükkel. Még idősebb imágók is tudnak színük változtatásával a környezethez igazodni, igaz, itt már csak részben képesek annak alapszínével azonosulni. Tehát ezen alkalmazkodási képesség a Saga fajoknál nem kötődik a vedlésekhez (Kaltenbach 1970).
2.5 Elterjedés
2.5.1 Égtáji elterjedés
A Saga genus minden képviselője Eurázsiában él, s. Észak-déli elterjedésük Kurgántól (Sverinogolovskoje, 54º 30' É) a Júdeai fennsíkig (Beit Jimal, 31ºÉ 43') húzódik. Elterjedésük keleti határa Ujguriában (Xinjiang, Kína) (Huang 1987), nyugati irányban pedig az Atlanti- óceán partja Portugáliában (Vila Nova de Milfontes, 8º 47' Ny) található (Kaltenbach, 1970).
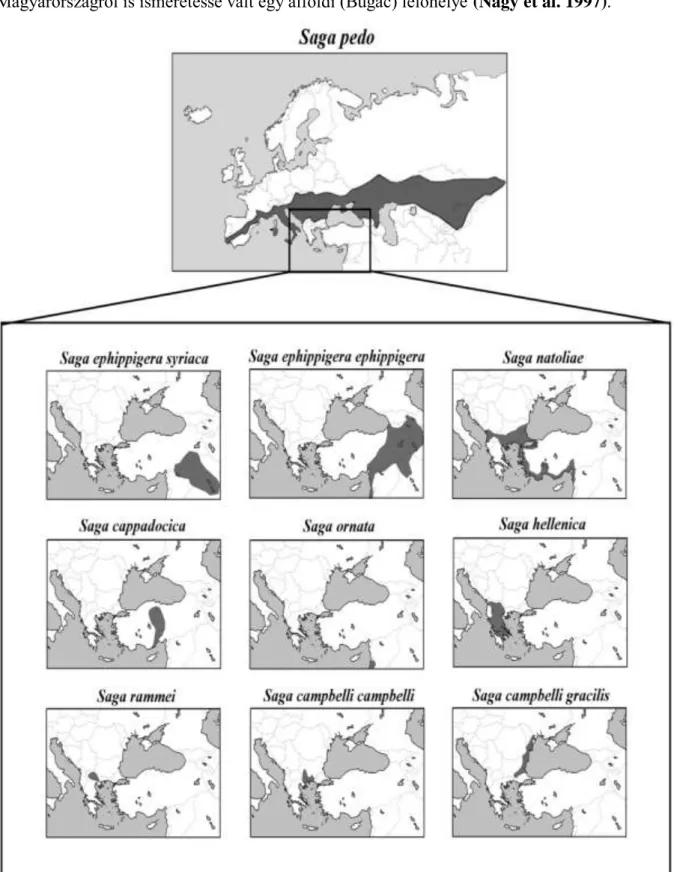
Azonban két faj van csupán, mely Ázsiában és Európában is él: a S. pedo és S. natoliae. E nagy kiterjedésű területen a fajok és populációik jórészt menedékekben ún. refúgiumokban léteznek, így többnyire szigetszerűen fordulnak elő - kivételt képez az elterjedési centrum területe, ahol gyakran összefüggő populációk találhatók (Willemse 1996). A vizsgálataimban szereplő fajok elterjedési területeit a 13. ábra mutatja be.
2.5.2 Magassági elterjedés
A Saga fajok zömmel hegyeken, dombokon, és fennsíkokon élnek, átlagban 150-600 m tengerszint feletti magasságban, ám néha ennél jóval magasabban is, ha a klimatikus viszonyok igényeiket kielégítik.
A Saga fajok közül legnagyobb magasságban a S. ephippigera fordul elő (1200-2750 m). A többi faj élőhelye zömmel 2000 m magasság alatt található (S. rammei: 750-1000 m ; S.
natoliae: 1000-1700 m, S. cappadocica: 1400-1600 m). A legnagyobb elterjedési területtel rendelkező S. pedo tengerszint felett általában 1500 m magasságig, néha magasabban is található: legmagasabb ismert előfordulási helye 1700 m magasságban található (Trpejca, Macedónia) (Kaltenbach 1967). Európában azonban többnyire 900m alatt található meg (Kaltenbach 1967): A Felvidéken (Szlovákia) 220-585 m (Krištin & Kańuch 2007);
Magyarországon 110-940 m (Nagy 1960), Svájcban 480-1680 m (Thorens & Nadig 1997) magasságban észlelték előfordulását. Olaszországban és Szlovéniában a tengermellékről is kimutatták (Fontana & Cussigh 1996), és a 90’-es években – nagy meglepetésre –
27
Magyarországról is ismeretessé vált egy alföldi (Bugac) lelőhelye (Nagy et al. 1997).
13. ábra: A dolgozatban szereplő fajok elterjedési térképe (a szerző munkája Kaltenbach 1967 és Huang 1987 alapján)
28 2.5.3 Elterjedést befolyásoló tényezők
Elterjedésüket elsősorban a klimatikus viszonyok, a talajféleség, valamint az adott élőhelyen rendelkezésre álló táplálék és a vegetáció határozza meg.
A Saga fajok kivétel nélkül melegigényes állatok, a S. pedo kivételével a 40º-tól délebbre fordulnak csak elő, emellett a fenti elterjedési területnek is zömmel a keleti részén élnek (Kaltenbach 1970).
Gyakran jelölik meg a Saga fajok élőhelyét úgy, mint dombok, hegyek dél- délkeleti tájolású száraz, sziklagyepei (Jaus 1934, Ebner 1951, Harz 1957). Hasonlóan azonban a Mantis religiosa (L.) fajhoz, a S. pedo nem kimondottan xerofil, azaz nem a csaknem teljesen kopár, gyér növényzetű napos területeket kedveli (mint pl. egyes sáskafajok), hanem inkább a meleg, ám viszonylag dúsabb vegetációhoz kötődik, ahol a növényzet párologtatása már egy speciális mikroklímát biztosít. E mikroklíma kialakításában a talajféleségnek is szerepe van, mely a Saga fajok esetében a tojásrakás közege is, tehát e vonatkozásban is befolyásoló tényező. A S. pedo többnyire meszes talajú élőhelyekhez kötődik (Jaus 1934, Müller 1924, Kaltenbach 1970).
E fajt mezőgazdasági művelés alatt álló területeteken is megtalálták, így pl. lucernamezőn (Bérengiuer 1905); Spanyolországban egy búzamezőn aratás előtt és után (Pantel 1886);
Ausztriában kukoricaföldről is bekerült az aratással (Lang 1930). Egy anatóliai Saga faj is került már elő hasonló helyről, melyet Ramme (1951) talált Ersin környékén egy lencseföldön. Azonban Közép-Európában újabb adatok szerint művelés alatt álló területekről hiányzik (Malicky 1963; Nagy 1960, 1965, Kaltenbach 1990), illetve Dél-Európában is előfordulása túlnyomóan a háborítatlan száraz sztyepplejtőkre korlátozódik (Willemse 1996).
Élőhelyen belül kisebb eltérések is megfigyelhetőek az egyek fajoknál: Míg a S. natoliae előnyben részesíti a magasabb bokrokat, s nem ritkán (pl. a déli órákban) azok belsejébe húzódik, a S. campbelli faj jórészt a magasabb füveken található meg (Kaltenbach 1970).
Részletesebb információ a S. pedo fajról áll rendelkezésre: Krištin & Kańuch (2007) megfigyelései szerint felvidéki (Szlovákia) élőhelyein a legtöbb egyedet (66%) 10-30 cm-es növényzeten találták meg, kisebb részük (19%) fordul elő csak 30 cm-nél magasabb vagy 10 cm-nél alacsonyabb vegetációjú területen (15%).
29
2.6 Egyedsűrűség
Régebben, különösen a Saga pedo-t illetően a szerzők kiemelték e faj alacsony egyedszámát (Kollar 1833, Redtenbacher 1900) közép- és nyugat-európai vonatkozásban.
Azonban sem a Saga pedo, sem a többi faj valójában nem ennyire ritka. Elsők közt Horváth (1916) utal arra, hogy e ritkaság – a Saga pedo esetében – csak látszólagos*. Későbbi szerzők (Jaus 1934, Kaltenbach 1970) alaposabb vizsgálata megmutatta, hogy e faj egyedei több 10 egyedes példányszámban vannak jelen élőhelyeiken. A legtöbb példányt (36 egyed) egyidejűleg egy ausztriai élőhelyen találták (Berg & Zuna-Kratky 1997).
Az vitathatatlan, hogy – különösen imágó állapotban – többnyire kis egyedszámú populációkban fordulnak elő (Nagy et al. 1983, Vidlička et al. 2002), rájuk akadni meglehetősen nehéz, a terület nagyságától is függően nem ritkán csak több órás keresés után kerül szem elé. Ebből is adódóan tehát lehetetlen lenne egy populáció abszolút létszámát meghatározni élőhelyének tönkretétele nélkül, bármely fejlődési állapotról is legyen szó, azaz egyedűrűség tekintetében is csak relatív, avagy látszólagos egyedsűrűségről beszélhetünk (Kaltenbach 1970).
A relatív egyedsűrűség napszakoktól eltérően változhat az egyes fajoktól függően.
Például a S. pedo leginkább csak meleg és napsütéses napokon mutatkozik, ezzel szemben a S. natoliae a nap meleg és napsütéses óráiban inkább visszahúzódik a bokrokba.
Az egyedsűrűség eloszlása az adott élőhelyen fajfüggő: a S. natoliae inkább egybefüggő közösségeket alkot, ám az egyedsűrűség kicsi. Ezzel ellentétben a S. campbelli campbelli populációi kisebb szórványokban fordulnak elő, melyekben ugyanakkor nagy egyedsűrűségben találhatók meg (Kaltenbach 1970).
A kis egyedsűrűség okaként Campbell (Burr et al. 1923) a Saga fajok kannibalizmusát jelölte meg. Kaltenbach (1970) később fontosabbnak tekinti a helyi tényezőket. Így, a talajféleség (tojásrakáshoz való alkalmassága), a helyi növényzet, illetve mikroklimatikus feltételek befolyásolják leginkább a fajok elterjedését.
*Dobiasch Ferenc megfigyelését írja le, Klaričevac környékén: egy alkalommal, amikor a település környékén egy sziklagyep kigyulladt, Saga pedo (S. serrata néven) egyedei ―száz számra‖ menekültek az égő bozótos területről, ahol korábban a fajt csak ―egyes példányokban‖ találta.
30
2.7 Aktivitás, napi ritmus és mozgás
A legkorábbi adatok a Saga fajok napi aktivitásával kapcsolatban szinte kizárólag a Saga pedo-val kapcsolatos megfigyelésekre szorítkoznak, és meglehetősen ellentmondásosak abban, hogy a Saga pedo illetve a Saga fajok nappali, avagy éjjeli aktivitással rendelkező fajok-e. Ugyancsak nem egyértelműek a megállapítások arról, hogy a faj fény- vagy melegkedvelő. Az éjjeli vagy nappali aktivitást illetően az ellentmondás zömmel annak tudható be, hogy az állat aktivitását annak megtalálási időszakához, annak megtalálhatóságához mérték (Kaltenbach 1970).
Kaltenbach (1970) ennek tisztázására javasolja az aktivitás alapjául venni a mozgás intenzitását, a ciripelést, táplálkozást, párzást, valamint a tojásrakás időszakát. Befolyásoló tényezőként külső és belső tényezőket nevez meg. Külső tényezőként a nappali fény-árnyék viszonyt, a nappalhossz változását, mikroklimatikus tényezőket; belső tényezőként pedig a fejlődési fázist és a fiziológiai állapotot tekinti. Vizsgálataiban megállapította, hogy júliusban a S. natoliae szinte mindig csak este 6 óra után kerül szem elé, hozzátéve, hogy ugyanazon élőhelyen (Kavalla) ugyanabban az évben a S. c. campbelli napközben ugyancsak nem volt megtalálható. Azonban ugyanezen élőhelyen egy másik évben a S. c. campbelli 9:00 és 17:00 közötti időszakban rendszeresen fellelhető volt jóval üdébb növényzet esetén.
Kaltenbach (1970) szerint az idősebb lárvák, valamint imágó állapotban az első négy hétben mutatják a Saga fajok a legnagyobb aktivitást minden jellemző tényezőt összegezve. A napi ritmust illetően is eltérések tapasztalhatók a júniusi és az októberi aktivitás (S. natoliae) között: júniusban akár már 16 órakor előjönnek, a hímek viszont csak 17 órakor kezdenek ciripelni. A legmelegebb időszakban, július és augusztusban rendszerint csak 18 óra után mutatkoznak. Szeptemberben és októberben ez már kevésbé figyelhető meg.
A tojásrakási aktivitást illetően már Matthey (1941) leírja, hogy az a S. pedo esetében mindig este vagy éjjel történik. Schall (2002) több S. pedo egyeden történt megfigyelése alapján írja, hogy a tojásrakás időszaka zömmel az esti órákra tehető, de néha a kora délutáni órákban is megtörténik. Soha nem figyelt meg azonban tojásrakást a reggeli vagy délelőtti órákban. Először a talajt a tojásrakásra kiszemelt területen végigtapogatja, majd tojócsövével próbát végez. A tojásrakás Schall (2002) megfigyelései szerint az imágóvá vedlés utáni 14- 16. napon veszi kezdetét.
A röpképtelen Saga fajok mozgása talajon, egyenletes terepen jellegzetes, szabályszerűen történik: Először a bal első és jobb középső lábával mozdul előre; azután a
31
jobb első és bal hátsóval mozdul, végül a bal középső és jobb hátsó lábakat emeli. E mozgási ritmus állandó; megegyezik a talajlakó állatok, pl. futóbogarak mozgásával. Az általában akadályokkal tűzdelt élőhelyen (pl. növényzeten közlekedés) viszont a terephez igazodik a lábak mozgása. Különleges, a Saga fajokra jellemző egy óvatos mozgás, amikor hirtelen ráijesztést követően felvett kataleptikus állapot után igyekszik továbbállni. Ekkor a lábsorrend tekintetében szabályos mozgási formát követ ugyan, ám lábait a talajra helyezés előtt előre- hátra himbálva halad lassan és vontatottan. Nagyobb folyamatos zavarás esetén pl. a S. pedo potrohát égnek emelve fut, melyet nagyobb ugrásokkal szakít meg. Ugrásukat használják továbbá támadás során, amikor a prédát már kellő távolságra, kb. 10 cm-re megközelítik:
ekkor egy kisebb, jól irányzott ugrással vetik rá magukat. Hasonlóan kisebb távolságról ugranak az észlelt nőstényre is (Kaltenbach 1970).
Az idősödő szöcskék aktivitása fokozatosan mérséklődik: táplálékfelvételük csökken, mozgásuk lassabbá válik, az állat magát is mind ritkábban, kevésbé alaposan tisztogatja. A testen fokozatosan nekrotikus foltok jelenhetnek meg. A palpus-ok, valamint a csápok végei elhalnak, leszáradnak. Életük vége felé a fűrészlábú szöcskék erőtlenné válnak, az állat egyre kevésbé képes függőleges irányú mozgásra, majd már csak nehézkesen vonszolja magát.
Pusztulásuk oka tehát leginkább végelgyengülés; az életfolyamatok folyamatos lassulása, majd leállása következik be (Kaltenbach 1967).
2.8 Szaporodás
2.8.1 PárosodásKét Saga faj, a S. ephippigera és a S. natoliae párzásáról először Boldyrev (1915) közölt adatokat, hozzávetőleges pontossággal. Ezt követően a Saga fajok párzási etológiájával kapcsolatban Kaltenbach (1970) közölt részletes leírást a S. natoliae, S.
rhodiensis és S. campbelli fajokra vonatkozóan. Ebben megállapítja, hogy a fűrészlábú szöcskék párzása az egyes fajokra jellemző viselkedési sajátosságokkal bír. Megfigyeli, hogy a Saga fajok esetében a nőstény nem mutat pozitív phonotaxis-t a ciripelő hím felé, ellentétben a legtöbb nem duettező (Robinson & Hall 2002; Rössler et al. 2006) szöcskefajjal. A S. campbelli faj egy példánya esetében Kaltenbach (1970) megfigyeli, hogy a hím rendes ciripelése halkabb, mint amikor ugyanazon hím egy nőstény közelében található, s végül leghangosabbá ténylegesen párzás közben válik. Ebből következőleg felveti, hogy a
32
csalogatáson kívül, nagyobb szerepe lehet a hangnak a nőstény támadási reakciójának leküzdésében is, melyet erősít egy megfigyelése is, melyben egy sérült szárnyú, alig hallható ciripelést produkáló hím egyed közeledése egy nőstény állatnál támadási reakciót váltott ki. A párzásra való érettség kivételes esetben röviddel (a S. ephippigera egy egyedénél mintegy 7 órával) az utolsó vedlés után is bekövetkezhet már, ám általában legalább 24 óra telik el, míg a hím a friss imágó nőstényt elfogadja Boldyrev (1915).
Boldyrev (1915) a S. ephippigera faj esetében „udvarlást‖ is említ: a nősténytől még nagyobb távolságban lévő hím potrohát behajlítja, és felveszi a párzáskor megszokott pózt, majd ezt követően egymás csáppal történő „letapogatása‖ következik. Kaltenbach (1970) szerint a balkáni fajok esetében is megfigyelhetők hasonló viselkedési sémák, azonban szerinte ez nem egy esetleges násztánc része, sokkal inkább átmenet félelemérzet és szexuális viselkedés között; ebből kifolyólag a nőstény is először könnyen prédaként vagy ellenségként fedezi fel a hímet. Kaltenbach (1970 szerint első kontaktus a hím és a nőstény között mindenképpen a csápok általi intenzív, kölcsönös letapogatás, majd ezt követi a hím ciripelése és közeledése. A párzást megelőzően a nőstény néha agresszíven léphet fel párjával szemben, megsebezheti és prédaként el is fogyaszthatja (Sänger 1987).
A párzás kezdetével a hím állat légzése intenzívebb lesz. Maga a párzás a hímnek a nőstényre ugrásával veszi kezdetét; majd a hím, testét begörbítve megkezdi a női ivarnyílás keresését, mely változó hosszúságú lehet, a terepi akadályoktól, illetve a hím egyed ügyességétől függően, általában 2-10 perc. A subgenitális lemez felnyitása, majd pedig a behatolás után elhelyezi a spermatofort. Ezt követően fajtól függően a nőstényt őrizheti, vagy odébbáll. A párzás után valamivel (S. natoliae: 20-24 óra, S. ephippigera: 3-17 óra) a nőstény a hímtől kapott spermatophort leeszi magáról (Kaltenbach 1970).
A párzásnál az ugrás, a jellegzetes pozíció felvétele, majd a subgenitális lemez megkeresése, a spermatofor elhelyezése, azonos minden Saga faj esetében. Az egyes fajoknak vannak - csak az adott fajra jellemző - etológiai sajátosságai is.
A S. natoliae hímje párzás közben rágójával megragadja a nőstény tojócsövét. Párzás után a hím őrzi a nőstényt még nagyjából fél óráig rágójával finoman megragadva potrohának alsó részén a nőstényt. A nőstény e nyugalmi állapot közben lábait széttárva mozdulatlan marad, s amennyiben szabadulni próbálna, a hím rágójával és lábaival erősebben megragadja.
E mellett – különösen, ha az előzőek nem jártak sikerrel - ciripelhet is, mellyel már szinte biztosan maradásra bírja párját (Kaltenbach 1970).
33
A S. ephippigera párzása közben nem fogja rágójával párja tojócsövét, és a hím phallus-ét nem húzza vissza, ellentétben a S. natoliae és S. rhodiensis fajokkal (Kaltenbach 1970).
A S. campbelli és a S. rhodiensis párzásának menete: a kopuláció végeztével a hím elég gyorsan odébbáll; a párzás menete, póza pedig a Saga fajokra fentebb említett általános jellemzőkkel megegyező.
Kaltenbach (1970) szerint a S. natoliae és a többi faj először általa részletesen közölt párzási etológiai sajátosságai a következő összefüggésekre világítanak rá:
A S. natoliae faj hímje kivételes módon rágójával ragadja meg párját. Ennek oka feltehetőleg, hogy a potrohvégi cercus-ok a nem másik három vizsgált fajával ellentétben nem rendelkeznek megvastagodott mediális résszel. Így a tojócsőbe kapaszkodás lehet az, ami segít megakadályozni a hím párzási pozícióból való kicsúszását. Mindazonáltal ez nagyobb erőkifejtéssel jár, így ez lehet az oka annak, hogy a hím párját néha csak egy órával a párzás után hagyja el, vagyis nagyobb párzás utáni nyugalmi állapot szükséges.
Mind a hímek, mind pedig a nőstények ismételt párzásra képesek. A S. ephippigera hímje két napon belül háromszor is képes párzani, 10 perccel a sikeres párzás után pedig már ismét párzásra késznek mutatkozik Boldyrev (1915). Ugyanazon Saga pár egyedei, párzás után közvetlenül újra közel kerülve először a hím, majd a nőstény is védekező állásba helyezkedik A frissen párzott Saga nőstény spermatophorral természetszerűleg újabb párzásra nem képes, s egy hím esetleges próbálkozása sikertelen (Kaltenbach 1970).
Különös érdeklődésre tarthat számot a Saga fajok fajidegen párzása. Kaltenbach (1970) a következő megfigyeléseket tette e vonatkozásban:
S. pedo Pallas (♀) × Saga campbelli campbelli Uvarov (♂)
Öt párzási kísérletből három sikeres, ebből egy a S. pedo egy természetes élőhelyén (Mödling, Ausztria) történt. A párzás a S. c. campbelli párzásának szokványos módján történik. Két esetben a hím a fajidegen nősténnyel való párzási próbálkozást fél óra múltán feladta. Egy esetben azonban ismételt párzás történt 6 nappal később, ugyanazon S. pedo nősténnyel.
Saga pedo Pallas (♀) × Saga natoliae Serville(♂)
Saga campbelli campbelli Uvarov (♀) × Saga natoliae Serville(♂)
34
A S. natoliae hímet mindkét faj nősténye azonnal partnerként érzékelte, mozdulatlanul megmerevedve, szokványosan passzívan viselkedett. Mindazonáltal a jóval nagyobb termetű hím próbálkozása a méretbeli és anatómiai (phallus, cerci) különbségek miatt eleve kudacra volt ítélve. A S. natoliae hím e próbálkozás után azonos fajú nősténnyel sikeres párzást folytatott.
Kaltenbach (1970) szerint interspecifikus párzás esetében:
- A rokon fajok hím és nőstény összepárzásának bizonyos fajok esetében nincsenek ökológiai korlátai, azonban nagyon valószínű, hogy a megtermékenyítésnek már igen.
- A S. pedo esetében is érvényes, hogy - a rokon fajokhoz hasonlóan – az egészséges (nem sérült szárnyú) fajidegen hím párzási szándékkal történő közeledésére engedően reagál.
- Hasonló nagyságú partnerek esetében a párzás során a spermatofor is elhelyezésre kerül, nagyon eltérő nagyságú fajok esetében viszont ez nem lehetséges, minthogy a szexuális inkompatibilitás rögzült tulajdonság, valamint a phallus és a női genitália nyílása anatómiailag nem hozható kapcsolatba. Ennek kapcsán Kaltenbach (1970) hozzáfűzi, hogy érdekes ezen összefüggésben a nagyon változatos nagysággal rendelkező S.
ephippigera faj esete, mivel e fajnál a testhossz 55-113 mm között nagyon tág határok közt változhat.
Lemonnier-Darcemont & Darcemont (2007) szerint a Saga rammei Kaltenbach (♂)
× Saga pedo Pallas (♀) párzásából hibridek születnek, köztük hímek. Az F1 nemzedék fertilitásáról azonban nincsenek adatok. Morfológiailag ezen pároztatásból származó hím (n=3) különböző morfológiai bélyegek szempontjából rokon fajokra hasonlít: habitusa tekintetében a S. rammei és S. campbelli fajokra inkább, mint a S. pedo-ra. A cercus-okat és az elytra alakját illetően leginkább a S. rammei-re, a nyakpajzs (tegmen) és az ugróláb hossza tekintetében a S. rammei és S. campbelli fajokra hasonlít; a testhossz és pronotum vonatkozásában nem elkülöníthető (Lemonnier et al. 2008).
35 2.8.2 A Saga fajok kariogámiája
A legtöbb palearktikus Tettigoniidae faj kromoszómakészlete 2n= 31 (X0, ♂); illetve a 2n= 32 (XX, ♀) formát mutat, a kromoszómák akrocentrikus elrendezésben találhatók (Hewitt 1979, Warchałowska-Śliwa 1998, Warchałowska-Śliwa et al. 2005). A Saga nem képviselőinek kromoszómakészlete viszonylag hasonló (Warchałowska 2007). Az ősi típusnak felel a S. cappadocica: 2n=31 (♂) (Matthey 1950) és a S. ornata : 2n=31 (♂) (Matthey 1946) kromoszómaszáma.
E típustól eltérő viszont három fajé: S. natoliae, 2n=29 (X0, ♂), 2n=32 (XX, ♀); S.
hellenica: 2n=29 (X0, ♂), 2n=30 (XX, ♀), S. rhodiensis: 2n=29 (X0, ♂), 2n=34 (XX, ♀), ezen túlmenően pedig egy metacentrikus autoszómájuk, és egy X submetacentrikus ivari kromoszómájuk van. E három faj allopatrikus elterjedésű (Warchałowska et al. 2007).
Mindez a fenti három faj közelebbi rokonságára utal, s egyúttal eltér a Kaltenbach (1967)- féle morfológiai alapú csoportosítástól (Warchałowska et al. 2007).
A S. pedo kromoszómaszáma: 4n =68 (♀); 6 pár metacentrikus és 28 pár akrocentrikus kromoszómával (Matthey 1939, 1941, 1948, Goldschmidt 1946), ez által a nem egyetlen tetraploid képviselője (Matthey 1946, 1948).