DOKTORI ÉRTEKEZÉS
KÖRNYEZETI VÁLTOZÓK HATÁSA RÁKKÖZÖSSÉGEK SZERKEZETÉRE
Készítette:
Baranyai Eszter okl. zoológus
Pannon Egyetem, Kémiai és Környezettudományi Doktori Iskola, Veszprém 8200
Egyetem u. 10
Témavezető: Prof. Dr. Padisák Judit Intézetigazgató, egyetemi tanár, az MTA doktora
Pannon Egyetem, Környezettudományi Intézet, Limnológia Intézeti Tanszék, Veszprém
Témavezető: Prof. Dr. G.-Tóth László
Tudományos tanácsadó, egyetemi tanár, az MTA doktora MTA ÖK Balatoni Limnológiai Kutatóintézet, Tihany
Pannon Egyetem
Kémiai és Környezettudományi Doktori Iskola Veszprém
2012
KÖRNYEZETI VÁLTOZÓK HATÁSA RÁKKÖZÖSSÉGEK SZERKEZETÉRE
Értekezés doktori (PhD) fokozat elnyerése érdekében
Írta:
Baranyai Eszter
Készült a Pannon Egyetem Kémiai és Környezettudományi Doktori Iskolája keretében
Témavezetők: Prof. Dr. Padisák Judit és Prof. Dr. G.-Tóth László Elfogadásra javaslom (igen / nem)
(aláírás) Elfogadásra javaslom (igen / nem)
(aláírás)
A jelölt a doktori szigorlaton ... %-ot ért el.
Az értekezést bírálóként elfogadásra javaslom:
Bíráló neve: ... igen/nem
...
(aláírás) Bíráló neve: ... igen/nem
...
(aláírás) A jelölt az értekezés nyilvános vitáján ... %-ot ért el
Veszprém, ...
a Bíráló Bizottság elnöke
A doktori (PhD) oklevél minősítése ...
...
Az EDT elnöke
1 Tartalomjegyzék
1 Tartalomjegyzék ... 3
2 Kivonat ... 5
3 Abstract ... 6
4 Zusammenfassung ... 7
5 Bevezetés ... 9
6 Irodalmi összefoglaló ... 10
6.1 Cladocerák és Copepodák rendszertani besorolása ... 10
6.2 A zooplankton kutatása a Balatonban ... 14
6.2.1 A Balaton, a zooplankton élőhelye ... 14
6.2.2 A zooplankton horizontális elterjedése a Balatonban ... 18
6.2.3 A zooplankton vertikális vándorlása ... 19
6.2.4 A turbulencia hatása a zooplanktonra ... 20
7 Célkitűzés ... 25
8 Anyag és módszer ... 26
8.1 Zooplankton mintavétel és a minták feldolgozása ... 26
8.2 A zooplankton tér- és időbeni eloszlásának vizsgálata a Balatonban ... 28
8.3 A zooplankton napszakos vándorlásnak vizsgálata ... 28
8.4 A vizsgált környezeti változók ... 29
8.4.1 Turbulencia mérés és a velociméter bemutatása ... 29
8.5 Turbulencia generátoros kísérletek ... 33
8.6 Az alkalmazott statisztikai módszerek bemutatása ... 36
8.6.1 A zooplankton tér- és időbeni eloszlásának vizsgálata ... 36
8.6.2 A mért környezeti változók tér- és időbeni mintázatai ... 37
8.6.3 Turbulencia generátoros kísérletek ... 38
9 Eredmények ... 38
9.1 A zooplankton tér- és időbeni eloszlása a Balatonban ... 38
9.1.1 A zooplankton szerkezetében mutatkozó tér- és időbeni mintázatok ... 38
9.1.2 A mért környezeti változók tér- és időbeni mintázatai ... 46
9.1.3 A környezeti változók és a zooplankton eloszlásában megfigyelt párhuzamok ... 52
9.2 A zooplankton napszakos vándorlása eltérő időjárási körülmények között 60
9.3 A turbulencia hatása a zooplanktonra kísérletes körülmények között ... 65
10 Diszkusszió ... 69
10.1 A zooplankton tér- és időbeni eloszlása a Balatonban ... 69
10.2 A zooplankton napszakos vándorlása eltérő időjárási körülmények között 76 10.3 A turbulencia hatása a zooplanktonra kísérletes körülmények között ... 79
11 Összefoglalás ... 80
12 Köszönetnyilvánítás ... 81
13 Irodalomjegyzék ... 82
14 A Tudományos tevékenység adatai ... 96
14.1 Publikációk ... 96
14.2 Prezentációk ... 97
15 Tézisek ... 98
16 Theses ... 100
17 Függelék ... 104
2 Kivonat
Környezeti változók hatása rákközösségek szerkezetére
A doktori értekezésben bemutatott kutatás célkitűzése a sekély, élénk hidrodinamikájú Balatonra jellemző turbulencia viszonyok és egyéb környezeti paraméterek feltérképezése volt, és hatásuk vizsgálata a zooplankton társulás szerkezetére, elhelyezkedésére, a különböző fajokra és egyedfejlődési stádiumaikra. A tudományos eredmények a következők:
A szerző elemezte a Balaton eltérő környezeti adottságokkal rendelkező vízterein a hőmérséklet, a Secchi átlátszóság, és a turbulencia alakulását. Kétféle módon számolta a víztestek energiadisszipációs rátáját, és a Kolmogorov-távolságot. Megállapította, hogy a Balatonban a sekély vízmélységből adódóan kevés tér áll rendelkezésre a kinetikai energia elnyelődésére. Ennek következtében gyakori az üledéket felkavaró örvények kialakulása, valamint a legkisebb örvények átmérője sokkal kisebb, a turbulens nyírófeszültségek értéke sokkal nagyobb, mint a mélyebb tavakban, illetve a tengerekben.
A szerző a tó eltérő kinetikai tulajdonságokkal rendelkező élőhelyein vizsgálta a zooplankton minőségi és mennyiségi viszonyainak alakulását. A kapott eredmények alapján megállapította, hogy a zooplankton fajok vízmélység szerinti eloszlásban egyedi sajátosságot mutattak. A kis testméretű nauplius lárvák, és a ragadozó Cyclopoidák a sekélyebb vizekben fordultak elő nagyobb mennyiségben, míg a szűrő táplálkozású Cladocera fajok és a Calanoida Eudiaptomus gracilis a mélyebb víztesteket preferálták.
A szerző Magyarországon első ízben vizsgálta a turbulencia hatását a tó zooplankton állományára, összevetve más környezeti paraméterek jelentőségével. A kapott eredmények alapján megállapította, hogy a turbulencia zooplankton közösség egészére gyakorolt hatása másodlagos volt az évszakossághoz, és a területi hatáshoz viszonyítva. A szűrő szervezetek azonban fokozott érzékenységet mutattak a turbulencia hatásával szemben, amely megmutatkozott területi elhelyezkedésükben is.
A szerző 24 órás méréssorozattal vizsgálta az eltérő turbulencia viszonyok hatását a zooplankton vertikális vándorlására. Megállapította, hogy a Daphnia cucullata x galeata, és az Eudiaptomus gracilis szélcsendes időben napi vertikális vándorlást végez. 4 cm s-1 – os RMS-turbulencia felett a vertikális vándorlás nem alakult ki. A szeles napok számát és a turbulencia viszonyokat tekintve megállapította, hogy a Balatonban a zooplankton rétegződése és vertikális vándorlása gyakran szenved zavart.
A szerző laboratóriumban mesterségesen keltett hullámzással vizsgálta a turbulencia zooplanktonra gyakorolt hatását. Megállapította, hogy a turbulenciára a kis mennyiségű lebegőanyag-tartalom mellett is a szűrő szervezetek bizonyultak a legérzékenyebbeknek, a terepi megfigyelések eredményéhez hasonlóan.
3 Abstract
The impact of environmental variables on the structure of crustacean communities
The objective of the research presented in this doctoral thesis was to map turbulence distribution and environmental parameters in Lake Balaton, a shallow, hydrodynamically highly kinetic lake, and to study their effects on the structure and location of the zooplankton assemblage, on the individual species and their developmental stages. The following new scientific results were obtained:
Temperature, Secchi transparency and turbulence in sectors of divergent environmental properties of Lake Balaton were analysed. The energy dissipation rates of water bodies and the Kolmogorov length scale were calculated by two different methods. It was established that in Lake Balaton, due to the shallowness of the water, little space is available for the dissipation of kinetic energy. Consequently, eddies stirring up sediment are highly frequent, and the diameter of the smallest eddies is much smaller and the magnitude of turbulent shearing forces is much larger than in relatively deeper lakes or in the sea.
The quality and quantity of zooplankton was studied in biotas of the lake with divergent kinetic properties.
The results obtained allow to conclude that zooplankton species follow individual distributions as a function of water depth. Nauplius larvae of small body size as well as predatory cyclopoids are present in large numbers in shallow water, whereas the filter- feeding Cladocera species and the calanoid Eudiaptomus gracilis prefer deeper water.
The author is the first in Hungary to study the impact of turbulence on the zooplankton assemblage, and to compare it to the significance of other environmental parameters.
Based on the results obtained it can be established that the impact of turbulence on the zooplankton assemblage as a whole is secondary as compared to that of seasonality and regionality. Filter-feeders, however, exhibited an enhanced sensitivity towards the effects of turbulence, which was also reflected by their spatial distribution.
The impact of different turbulence intensities on the vertical migration of zooplankton was studied in a 24-hour measurement series. It was established that Daphnia cucullata x galeata and Eudiaptomus gracilis perform daily vertical migration in calm weather.
Vertical migration did not develop at flow velocities over 4 cm s-1. In view of the number of windy days and the known distribution of turbulence, it can be concluded that stratification of zooplankton is often disturbed in Lake Balaton.
The impact of turbulence on zooplankton was also studied in the laboratory using artificially generated waves. It was established that, even at low suspended material content, filter-feeding organisms proved to be the most sensitive to turbulence, an observation that coincides with the results of field monitoring.
4 Zusammenfassung
Die Wirkung von Umweltparametern auf die Struktur von Zooplanktongesellschaften
Das Ziel der in der Dissertation vorgestellten Forschung war es die Turbulenzverhältnisse und die Umweltparameter im seichten und hydrodynamisch lebhaftem Plattensee kennenzulernen, ebenso wie ihre Wirkung auf die Struktur, die Platzierung von Zooplanktongesellschaften und auf die verschiedenen Arten und verschiedenen Entwicklungsstadien. Die wissenschaftlichen Ergebnisse sind die Folgenden:
Es wurde in den verschiedenen hydrologischen Seegebieten die Temperature, Secchi-Tiefe und Turbulenz gemessen. Die Energiedissipationsrate und die Kolmogorovdistanz wurden berechnet, die erste auf zwei verschiedene Weisen. Es wurde festgestellt, dass im Plattensee aufgrund der geringen Tiefe wenig Raum zur Absorption der kinetischen Energie zur Verfügung steht. Daher entstehen oft Wirbel, die das Sediment aufmischen.
Weiterhin ist der Durchmesser der kleinsten Wirbel viel kleiner und die turbulenten Scherkräfte viel stärker als in tieferen Seen oder im Meer.
Die qualitativen und quantitativen Veränderungen des Zooplanktons wurden in den sich durch ihre Kinetik unterscheidenden Lebensräumen des Sees untersucht. Aufgrund der Ergebnisse kann man feststellen, dass die einzelnen Zooplanktonarten entsprechend der Wassertiefe eine charakteristische Verteilung zeigten. Kleine Nauplien und räuberische Cyclopoiden kamen in größeren Mengen im seichten Wasser vor, während die filtrierenden Cladoceren und der Calanoid Eudiaptomus gracilis größere Wassertiefen bevorzugten.
Es wurde zum ersten Mal in Ungarn die Wirkung der Turbulenz auf den Zooplanktonbestand des Sees, auch im Zusammenhang mit verschiedenen anderen Umweltparametern untersucht. Die Ergebnisse zeigen, dass der Einfluss von Turbulenz auf die Zooplanktongesellschaft im Ganzen im Vergleich zur Wirkung der Jahreszeiten und der lokalen Verhältnisse zweitrangig war. Partikelfresser waren jedoch der Turbulenz gegenüber emfpindlicher, was sich auch in ihrer Schichtung zeigte.
Auch eine 24-stündige Messreihe wurde durchgeführt um den Einfluss verschiedener Turbulenzverhältnisse auf die Vertikalwanderung des Zooplanktons zu untersuchen. Es wurde festgestellt, dass Daphnia cucullata x galeata und Eudiaptomus gracilis bei Windstille vertikale tagesperiodische Wanderungen durchführt. Bei einer Strömungsgeschwindigketi von über 4 cm s-1 findet keine Vertikalwanderung statt.
Betrachtet man die Zahl der windigen Tage und die Turbulenzverhältnisse, wird ersichtlich, dass die vertikale Schichtung des Zooplanktons im Plattensee oft gestört wird.
Im Laborversuch wurde durch künstlich generierte Wellen die Auswirkung von Turbulenz auf das Zooplankton untersucht. Es konnte festgestellt werden, dass, entsprechend der Beobachtungen im Freiland, auch hier, bei geringer Schwebstoffkonzentration, die Filtrierer am empfindlichsten waren.
5 Bevezetés
A Balaton zooplanktonját zömmel a Protozoa, a Rotatoria, a Crustacea, és a Mollusca- lárvák alkotják. Kutatásunk alanyai, az alsóbbrendű rákok a tó egyik legrészletesebben tanulmányozott szervezetei közé tartoznak. Mindezek mellett, a planktonikus rákok területi és vízoszlopon belüli elterjedését befolyásoló környezeti tényezők komplex vizsgálatára eddig még nem került sor. Különösen érdekes lehet, hogy a szél dinamikai hatásának olyannyira kitett Balatonban a turbulens áramlások miként befolyásolják a rákok életkörülményeit. A Balaton nagy kiterjedésű vízfelülete lehetővé teszi, hogy a szél nagy mennyiségű energiát adjon át a víztömegnek. Sekélysége (átlagosan 325 cm) okán a kinetikai energia elnyelődésére csak kevés tér áll rendelkezésre, melynek következtében a tó vize általában erősen turbulens. Más tavakban, valamint tengerekben már végeztek kutatásokat arra vonatkozóan, hogy az élénk vízmozgás milyen hatást gyakorol a plankton fejlődésére, táplálkozására (Rotschild & Osborn 1988; Mackas et al. 1993; Alcaraz et al.
1994; Kiørboe & Saiz 1995; Lagadeuc et al. 1997; Incze et al. 2001; Lewis & Pendley 2001; O´Brien 2002, O´Brien et al. 2004). A Balatonban ilyen jellegű vizsgálatot még nem folytattak. Célul tűztük ki, hogy a tó eltérő vízmélységeiben és különböző időjárási viszonyok között megvizsgáljuk más környezeti paraméterek mellett a turbulencia hatását is a zooplanktont alkotó rákközösség egyedfejlődésére, mortalitására, és szerkezetére.
6 Irodalmi összefoglaló
6.1 Cladocerák és Copepodák rendszertani besorolása
A vizsgált kisrákok rendszertanilag Schram (1986) alapján az Arthropoda törzsbe, és a Crustacea altörzsbe tartoznak. A Cladocera rend a Phyllopoda osztály Calmanostraca alosztályába tartozik. A Copepoda alosztály a Maxillopoda osztályba sorolható, az alosztály három rendje a Calanoida, Harpactoida és a Cyclopoida rendek.
A Cladocera fajok (más néven ágascsápú rákok) kisméretű állatok, testhosszuk 0,25 és 18 mm között változik. Testük felépítése nagyon változatos, oldalról rendszerint összenyomottak, vagy karcsú, megnyúlt alakúak. Testüket kevés kivétellel két oldalról többé kevésbé átlátszó héj fedi. Szelvényeik elmosódottak, testük határozatlanul tagolt, tulajdonképpen csak fej és törzs különböztethető meg rajta. A fejükön 5 pár függelék található. A fejlett második csápok a Cladocera legjellegzetesebb szervévé, ágascsáppá alakultak. Az állat az ágascsápok ritmikus csapkodásával, „szökdelve” mozog. Általában lebegő életmódot folytatnak. A hasoldalukon páros végtagok találhatók, melyek csapkodásával előidézett vízáramlással sodorják szájukhoz a lebegőanyagokat és az apró élőlényeket. A száj szűrőrácsain fennmaradó részecskék a táplálékárkon át a garatba sodródnak, majd a bélcsatornába kerülnek. A szűrő életmódot folytató Cladocera fajok között akad ragadozó faj is, a Balatonban is előforduló Leptodora kindtii, melynek zsákmánya lehet minden 1,5 mm-nél kisebb kerekesféreg, rák vagy lárva (Manca &
Comoli 1995).
A Cladocera fajai általában egy- és kétivaros szaporodás szabályos váltakozásával szaporodnak, ismertek azonban kizárólag szűznemzéssel szaporodó populációk is. Az egy- és kétivarú szaporodásmód szabályszerű váltakozása a mostoha körülményekhez való alkalmazkodás eredménye. Ilyen módon vészelik át élőhelyük esetleg teljes kiszáradását is.
Az év nagy részében a nőstények szűznemzéssel olyan petéket hoznak létre, melyekből megtermékenyítés nélkül partenogenetikus nőstények kelnek ki.
Ha a körülmények kedvezőtlenné válnak, a partenogenetikus peték száma csökken, és közülük egyesekből hím példányok kelnek ki. Az utód nemét azok a külső hatások befolyásolják, amelyek a költőüregbe jutásig érik. A kétivaros szaporodás feltétele tehát a hímek és egyes Daphnia-féléknél az alaktanilag is eltérő megtermékenyíthető nőstények megjelenése.
A megtermékenyített tartóspete az anyaállat elpusztulásával, vagy vedlésével héjjal borított efippiumá alakul. A tartóspeték hónapokig nyugalomban maradhatnak, míg a környezeti feltételek ismét megfelelőek lesznek, ekkor az anyaállathoz hasonló utódok kelnek ki belőlük (Gulyás & Forró 1999).
Az ágascsápú rákokra jellemző feltűnő jelenség az évszakos alakváltozás, más néven a ciklomorfózis, melynek során az egyes testrészek egyenetlenül nőnek. Jellemző példája a Daphnia nem sisak képzése (1. ábra). Vizsgálatok szerint a ciklomorfózist a vízmozgás, a víz hőmérséklete, a fényklíma, és a tápanyag-ellátottság együttesen és külön-külön is kiválthatják (Gulyás & Forró 1999). A Balatonban a Daphnia cucullata ciklomorfózisát figyelték meg (Ponyi 1965). A ciklomorfózist a legtöbb kutató az alkalmazkodás jelenségének tartja. Többféle hipotézis látott világot magyarázatára. Az egyik szerint az állatok a testméretük növelésével a nyári kisebb viszkozitású vízhez alkalmazkodnak, ezáltal növelve alaki ellenállásukat, és lassítva süllyedésüket (Gulyás & Forró 1999).
Laforsch & Tollrian (2004a) munkájában a Daphnia cucullata sisakképzésnek lehetséges okaként a kisléptékű turbulenciát jelölte meg. Szélesebb körben elfogadott elképzelés szerint a ciklomorfózis a predációs hatás csökkentését szolgálja, mivel az átlátszó, nagyméretű sisakos formákat a vízben nehezebb észrevenni, illetve nehezebben férnek be a halivadékok szájába (Hutchinson 1967; Tollrian 1990, 1994, 1995; Tollrian & Harvell 1999; Jeschke et al. 2002; Laforsch & Tollrian 2004b).
1. ábra: A Daphnia sp. ciklomorfózisa (saját felvétel)
A Copepoda csoport (evezőlábú rákok) átlagos hossza 1-2 mm, de a legnagyobb termetűek sem haladják meg a 10 mm-t. Jellegzetes formájuk van, testük kitinizált gyűrűkből, szelvényekből áll. A fejen található 6 pár módosult végtagjuk a táplálkozásban vesz részt.
A mozgásban a toron található 5 pár végtag, az úgynevezett evezőlábak játszanak szerepet.
Váltivarúak, szaporodásuk mindig ivaros, formája pedig a Harpacticoida fajoknál esetenként megfigyelt szűznemzés kivételével heterogámia. A megtermékenyítés után a peték átlátszó burokba kerülnek, amelyek a Cyclopoida fajoknál két, a Calanoida fajoknál egy petezsákká fejlődik. A petéket sok esetben a környezeti hatásoknak (pl. kiszáradás, hideg) ellenálló burok védi. A környezeti tényezők nagymértékben befolyásolják, hogy a petékből hím vagy nőstény egyedek kelnek ki. Tartóspete képzésére a Cladocera fajokhoz hasonlóan képesek. A petékből úgynevezett nauplius lárvák kelnek ki. A Copepoda szervezeteknek 11-12 fejlődési stádium van, nauplius stádiumok után 5 copepodit, majd a kifejlett állat alakul ki.
Az evezőlábú rákok planktonikus formái mindenevők, többségük a táplálékát szűréssel szerzi meg. Táplálkozásuk során valószínűleg először érzékszerveikkel érzékelik a táplálékot, és ezt követően mechanikusan átszűrik a közeget.
A szűrés sebessége nagymértékben függ a közeg hőmérsékletétől, a táplálék mennyiségétől, és a fényviszonyoktól. A Calanoida fajok többsége valódi szűrőszervezet.
A Cyclopoida egyes fajai algaevők, mások ragadozók is lehetnek. A ragadozó fajok leggyakrabban állati egysejtűeket, kerekesférgeket, kevéssertéjű férgeket, kisebb ágascsápú- és kagylósrákokat, apróbb rovarlárvákat zsákmányolnak, de táplálékszervezeteik között más evezőlábú rákok, ill. azok naupliusait és copepoditjait is megtaláljuk. A Cyclops vicinus nőstények 62 %-ban, a hímek 89 %-ban növényevők (Adrian 1991a,b; Adrian & Frost 1993).
A Copepoda fajok jellegzetes tulajdonsága, hogy kedvezőtlen körülmények hatására élettevékenységük lelassul, látens állapotba (anabiozis) kerülnek, s így vészelik át a számukra kedvezőtlen időszakot. Ha a látens állapotban levő állat ismét megfelelő körülmények közé kerül, az élettevékenységek hamar normalizálódnak.
Mozgásukat a testalkatuk híven tükrözi. A Calanoida csoport szinte valamennyi képviselője lebegő életmódot folytat (2. ábra). Első csápjuk a testnedvek nyomása révén merevvé válik, kifeszül, így az állat csak lassan süllyed lefelé. 2-4 cm-es süllyedés után a csápok elernyednek, a test mellé simulnak, majd a lábak erőteljes csapásával az állat
„ugrik” néhány centimétert a vízben. A Cyclopoidák többsége úszó szervezet (3. ábra).
Torlábaikkal csapkodnak, percenként mintegy 60-szor. Mikor lábaikkal hátrafelé csapkodnak, ugrásszerűen ferdén felfelé lökődnek, majd kis szakaszon visszasüllyednek, miközben farokvillájuk és csápjaik kormányként működnek. A Harpacticoida fajokra a kígyózó mozgás jellemző. Törzsük oldalirányú tekergődző mozgásával haladnak a fenéken, rövidebb szakaszon úszni is tudnak.
A Copepoda fajok évszakos alakváltozása az ágascsápú rákokhoz viszonyítva jelentéktelen (Gulyás & Forró 1999).
2. ábra: Eudiaptomus gracilis (saját felvétel) 3. ábra: Cyclops vicinus (saját felvétel)
6.2 A zooplankton kutatása a Balatonban
6.2.1 A Balaton, a zooplankton élőhelye
A Balaton Közép-Európa legnagyobb édesvizű sekély tava. Területe 594 km2; hosszúsága 77 km, szélessége 1,5-12 km. Felületéhez képest a víz mélysége csekély, átlagos mélysége 3,25 m, legmélyebb pontja az ún. tihanyi kút (11 m). Vizét 20 állandó és 31 időszakos vízfolyás, valamint néhány víz alatti forrás táplálja. A felszíni vízfolyások közül a legjelentősebb a Zala folyó, amely a Keszthelyi-medencébe jutva a tó vízhozamának 50-60
%-át biztosítja. A tó mesterséges lefolyását a Sió biztosítja (Virág 1998).
A Balatonban a mély tavakra jellemző hőmérsékleti és kémiai rétegzettség nem, vagy csak ritkán és nagyon kis mértékben jelentkezik, tipikus polimiktikus tó. A víz teljes mélységében oxigéndús, gyakran oxigénben túltelített.
A vízmozgás előidézésében a szél, a légnyomás és a hőmérséklet hirtelen változása játszik szerepet. A vízmozgások általában horizontális irányúak. A vízmozgás a hullámzásban, a vízfelszín hossz- és keresztirányú lengésében, illetve a kilendülések mértékében, továbbá a köröző és átforduló, valamint a szekundér áramlásokban nyilvánul meg.
A légnyomáskülönbségek miatt a tóvíz mozgásában tólengés (seiche) jelentkezik.
Cholnoky (1897) tanulmányában ötféle szabályos ingadozást írt le, melyek között ismeretei szerint a Balatonban és egyben a Földön mért legnagyobb periódusidőjű lengés 10-12 óráig tart. A leggyakrabban előforduló hullámmagasságok 35 - 60 cm körüliek, de előfordulhatnak 2 méteres hullámmagasságok is (Muszkalay 1973). A tóra jellemző domináns észak-északnyugat irányú szeleknek köszönhetően, valamint a Balaton medencéjének alakja, elhelyezkedése következtében a hullámzás összetett hatásaként a déli partoknál széles homokpad épült, míg az északi parti sávok mélyebbek, egyúttal magasabbak.
A Balatont négy különböző vízkicserélődési idejű medence alkotja, melynek vízminőségi vonzata jelentős. A medencék trofitása a külső és a belső tápanyagterhelés függvénye, a fajlagos terhelés a Keszthelyi-medencében a legnagyobb. A fitoplankton biomasszája a tó hossztengelye mentén eltérő értékeket mutat, nyugatról keletre haladva a biomassza értéke csökken (Padisák 2002; Vörös et al. 2000). Az 1960-1980 közti időszakban a tóra az eutrofizáció volt jellemző, melyet reoligotrofizáció követett, köszönhetően részben a tápanyagterhelés csökkentésének, a Kis-Balaton szűrőhatásának (Virág 1998).
A Balaton vize (Ca, Mg) HCO3-os kemény víz, pH-ja (≈8,6) enyhén lúgos. Vize állandóan zavarosnak tűnik, átlátszósága ritkán haladja meg az egy métert, a kalcium-magnézium hidrokarbonátos jellege miatt. A fitoplankton a Balatonra jellemző pH mellett HCO3-ot tudnak szerves szénforrásul felvenni, emiatt a biogén mészkiválás intenzív. A lebegőanyag tartalom tág határok között változhat, attól függően, hogy a vízmozgások mennyi szilárd anyagot kevernek a víztérbe, és a pillanatnyi biogén mészkiválás milyen intenzív. A átlagos lebegőanyag tartalma 25 mg l-1, de amikor a szél tartósan 400 cm s-1-nál erősebben fúj, az egész tó vize felkeveredik. Viharok alkalmával a felkevert vízben a lebegő üledék mennyisége valamennyi medencében elérheti az 500-600 mg l-1 értéket (Entz & Sebestyén 1942; Entz 1983; Somlyódy 1986; G.-Tóth et al. 1999). A hullámzás az üledék felkavarásával sajátságos életviszonyokat teremt tavunkban és ezért a hullámzást nem lehet egyszerűen csak hidrodinamikai kérdésnek tekinteni (Enzt & Sebestyén 1942).
6.2 A zooplanktont alkotó rákközösség szerkezete és kutatása a Balatonban
A balatoni planktonrákok vizsgálata az 1880-as években vette kezdetét Daday (1884, 1885) kutatásaival. 1897-ben „A Balaton faunája” (Entz 1897) című kötethez „Rákfélék (Crustacea)” címmel külön fejezetet írt, melyben 26 Cladocera és 12 Copepoda fajt említ.
Sebestyén 1930-as évek elejétől az 1970-es évek elejéig folytatott vizsgálatai főleg a Cladocera fajokat érinti, kiemelten a Leptodora kindtii ragadozó fajt. Megjelent cikkei a planktonrákok mennyiségi és minőségi viszonyaival, biomasszával foglalkozik (Sebestyén 1931, 1933, 1947; Sebestyén et al. 1951; Sebestyén 1953, 1955, 1960, 1964, 1970). Ponyi 1953-tól kezdve vizsgálja a Balaton rákfaunáját (Ponyi 1955, 1957, 1962; Ponyi & Tamás 1964; Ponyi 1965, 1968, 1975, 1981; Ponyi et al. 1982; Ponyi & H. Péter 1986; Ponyi 1993, 1997). Az 1970-es években többek között részletesen vizsgálta az Eudiaptomus gracilis evezőlábú faj táplálkozását. A Magyarországon kimutatott Cladocera fajok száma 95, melyek 56 %-a él a Balatonban (Forró 1997; Ponyi 1997). A Copepoda csoport esetében pedig a hazai 54 fajnak mintegy 37 % fordul elő a Balatonban.
A Cladocera fajok száma Daday (1897) által kimutatott 26-ról az évszázad közepére 66-ra emelkedett (Gulyás & Forró 1999), minden bizonyára az intenzívebbé váló kutatásoknak köszönhetően is. Ponyi (1997) vizsgálatai szerint a következő évtizedekben számuk csekély mértékben csökkent. Bizonyos fajok pl. Bosmina coregoni 1996 előtti jelenléte kérdéses, mivel korábbi mintákból nem lehetett kimutatni. Más fajok, mint pl. Moina rectirostris, az utóbbi 20-30 évben eltűntek, vagy állományuk jelentősen visszaszorult. A taxonok kicserélődése mellett, és a rendszertani pontosításoktól eltekintve az elmúlt fél évszázadban a Cladocerák faji összetétele alig változott. A Daphnia fajok szezonális formaváltozásának vizsgálatára irányuló alaposabb kutatások többek között bebizonyították, hogy a korábban D. cucullata néven ismert faj mellett a D. galeata is megtalálható a Balatonban. A legújabb kutatások során genetikai vizsgálatokkal bizonyították, hogy a D. galeata és a D. cucullata faj közös hibridje is előfordul a tóban (Nédli et al. 2007).
Daday (1897) két Calanoida és 10 Cyclopoida fajt mutatott ki a Balatonban. A Diaptomus castor és a Diacyclops nanus fajokat akkori megtalálásuk óta nem sikerült újra begyűjteni.
Dévai (1977) vizsgálatai során egy Calanoida és 21 Cyclopoida faj került elő a 80-as években. A Cyclopoida fajszám a 90-es évek végéig csupán eggyel emelkedett, a Calanoidák fajszáma nem változott. A kutatások alapján a Cyclops vicinus (Ulianine) csak 1935 után jelent meg a Tihanyi-medencében (Entz et al. 1937; Sebestyén et al. 1951;
Sebestyén 1953). A Cyclops vicinus megjelenésével egy időben szorulhatott vissza a Macrocyclops albidus (Jurine), amelyről Daday (1897) és Entz (1897) még úgy írtak, mint uralkodó fajról. 1965-től sikerült kimutatni a tó nyíltvízi planktonjában a Thermocyclops crassus (Fischer) és az Acanthocyclops robustus (G. O. Sars) fajokat. Az Acanthocyclops sp. a fokozódó eutrofizáció indikátor faja volt. A Copepoda minőségi összetételének megváltozásáért olyan emberi behatások voltak felelősek, mint a növényvédő szerek használata, a nagyobb vízszintingadozások megszüntetése, és a busa betelepítése a Balatonba (Ponyi 1981).
A kutatások eredményeiként megfigyelhető volt, hogy a zooplankton állomány mennyisége 1951-ig növekedett, melynek hátterében a Cladocera és Cyclops fajok egyedszámának változása állt. Az 1950-es és 1960-as években a zooplankton mennyisége folyamatosan csökkent, a korábbi állománynagyság hatodára, negyedére (12,4 - 20,4 egyed l-1; Ponyi 1981). 1970 és 1990 között az eutrofizáció következtében a balatoni fitoplankton elsődleges termelése a korábbi állapotához képest 4-8-szorosára emelkedett (Herodek 1984). A zooplankton tömege ezzel szemben csak a 1,5-3-szorosára növekedett, mivel a tó magas ásványi lebegőanyag tartalma, és az eutrofizáció következtében elszaporodott fonalas kékalga együttesen gátolta a zooplankton szűrését (Zánkai 1985; G.-Tóth et al.
1986; G.-Tóth 1992, 2003). 1995-re a tó visszaállt eredeti állapotára, a nyári kékalga tömegprodukciók megszűntek. A 2001-2003-as rendkívül alacsony vízállású aszályos években a rákplankton sűrűsége az egész Balatonban csökkent. A Cladocera/Copepoda arány is szignifikánsan csökkent, melynek oka a Cladocera fajok fokozott érzékenysége lehet a megnövekedett turbulens hatással és a betöményedéssel szemben (G.-Tóth et al.
2008).
6.2.2 A zooplankton horizontális elterjedése a Balatonban
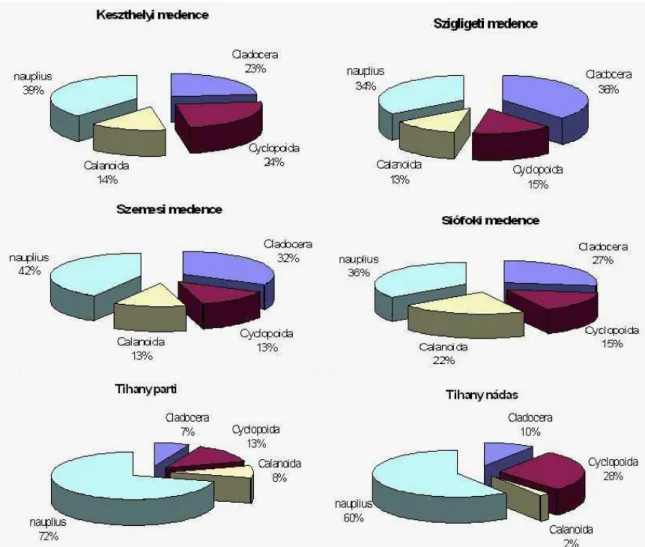
A Balaton sajátos adottságokkal rendelkezik, melynek alapja a négy medencére való tagolódása, a sekély mélység, a nagy kiterjedésű vízfelület, az uralkodó szél iránya, a befolyók egyenetlen eloszlása, és utóbbiak szennyezettségének eltérő mértéke. A Balaton területén jelentkező különbségek jogosan vetették fel annak gondolatát, hogy a planktonrák faunában horizontális eltérések lehetnek. Első ízben Sebestyén (1960) vizsgálta a planktonrákok horizontális eloszlásában és összetételében jelentkező különbségeket, melynek eredményeit az elégtelen technikai feltételek miatt csak tájékoztató jellegűnek tekinthetünk. A balatoni halpusztulás okait kutatva 1965-ben intenzív horizontális planktonvizsgálatok kezdődtek (Baron et al. 1967). Ponyi (1993) szerint a Cladocera fajok közül a Daphnia cucullata, Daphnia galeata, Diaphanosoma brachyurum, Leptodora kindtii, valamint a Copepoda fajok közül az Eudiaptomus gracilis, Mesocyclops leuckarti a nyári időszakban, a Bosmina longirostris tavasszal, a Cyclops vicinus pedig a hidegvízi időszakban az egész tóban elterjedt. A Copepoda fajok közül az Acanthocyclops vernalis szinte kizárólag a Keszthelyi-medencében élt, a Thermocyclops crassus pedig csak a Tihanyi-medencében. A planktonrákok átlagos biomassza- és egyedszám megoszlásában számottevő különbség volt tapasztalható, az eutrófabb vízterületeken a Cyclops fajok egyedszáma magasabb volt, mint más vízterületeken. Például az Acanthocyclops robustus egyedszáma lényeges emelkedést mutatott a Keszthelyi-medencében. A Daphnia cucullata állományára is jellemző volt Siófoki-medencéhez képest a Keszthelyi-medencében tapasztalható magasabb egyedszám. 1999 és 2007 közötti vizsgálat (G.-Tóth et al. 2008) alapján a tóban a Cladocera egyedszáma 1,6 és 384, míg a Copepoda egyedszáma 21,8 és 393 egyed l-l között alakult. Az egyedsűrűség a keleti tórész felől a nyugati felé haladva minden évben egyenletesen nőtt, és általában a Szigligeti-medencében mutatta a legmagasabb értéket.
6.2.3 A zooplankton vertikális vándorlása
A zooplankton horizontális és vertikális eloszlását, és az azt alakító fizikai és biológiai tényezők konkrét hatását sokan vizsgálták (pl. Alcaraz & Saiz 1991; Folt & Burns 1999;
Pinel-Alloul et al. 1999, 2004; George & Winfield 2000; Rinke et al. 2007). A vertikális vándorlás formája faj- és élőhelyfüggő, és különbség tapasztalható a populáción belül a különböző korú, lárvaállapotú, eltérő nemű egyedek között is, de évszakos változások is előfordulnak. A vándorlás leggyakoribb formája a normál vándorlás (nocturnal migration), amikor napnyugtakor a felszín felé, hajnalban lefelé vándorolnak a szervezetek, így felszín közelben éjjel találjuk a legtöbb egyedet. Egyes esetekben megfigyelhető az ezzel ellentétes fordított vándorlás (reversed migration), amikor felszíni tömörülés a nappali órákban tapasztalható, este a mélyebb rétegekbe húzódnak az állatok, és reggel, illetve a délelőtt során vándorolnak a felszín közelébe. További, az előzőektől eltérő típusok is ismeretesek, például a szürkületi vándorlás (twilight migration) amikor a nap során kétszer, alkonyatkor és hajnalban jelentkezik felszíni tömörülés (Cushing 1951; Hutchinson 1967;
Wetzel 1975; Raymont 1983). A zooplankton vertikális eloszlását és vándorlását a korábbi munkák elsősorban a foto- és a geotaxisra vezetik vissza (Ringelberg 1987, 1999; De Meester 1993; Moore et al. 2000). De megemlítik az aktivitásban és úszásban megfigyelhető viselkedési mintázatokat (Folt & Burns 1999) és a szél által keltett áramlás és az azt követő turbulencia nagyobb léptékű összetevői következtében bekövetkező passzív transzportot (George & Winfield 2000). Vizsgálatok bizonyítják, hogy a hőmérséklet, a táplálék koncentráció horizontális és vertikális gradiensei szerinti aktív orientáció is fontos szerepet játszik (George 1983; Lampert et al. 2003). A napi vertikális vándorlás egyik okaként a predáció elkerülését is említik (Zaret & Suffern 1976; Stich &
Lampert 1981; Kvam & Kleiven 1995), mivel a halak által kibocsátott feromonokat, kairomonokat bizonyos zooplankton fajok kemoreceptorokkal érzékelik (Tátrai & de Bernardi 1992; Van Gool & Ringelberg 1998; Ringelberg 2000; Hülsmann et al. 2004;
Van Donk 2006), bár a vertikális vándorlás olyan tavakban is megfigyelhető, amelyekben nem élnek halak (Kohout & Fott 2000). Egyes Daphnia fajokról megállapították, hogy a mozgásuk irányításáért a fényerősség relatív változása felelős (Ringelberg 1987, 1999;
Van Gool & Ringelberg 1998; Moore et al. 2000).
Néhány szerző úgy találta, hogy a szél keltette hullámok befolyásolják a zooplankton vertikális eloszlását (Megard et al. 1997; Easton & Gophen 2003; Karsten et al. 2007).
A Balatonban Entz & Sebestyén (1942), Ponyi & H. Péter (1986), Ponyi et al. (1982), Homonnay (2004), valamint saját korábbi tanulmányaink (Baranyai & G.-Tóth 2007) szerint a zooplankton vertikális rétegződése múlékony ugyan, de hosszan tartó szélcsendes időszakban kialakul. A Balaton zooplankton állományának korai vizsgálatai alkalmával megfigyelték, hogy csendes időben borult ég alatt egyes Copepoda és Cladocera fajok nagy mennyiségben gyülekeztek a felszín közeli vízrétegekben. A vihar azonban a társulást a mélység mentén homogenizálta (Daday 1884; Entz & Sebestyén 1942). Sebestyén (1933) a balatoni Leptodora kindtii vertikális elhelyezkedésének alsó határát a tó fenékhez közel állapította meg, és napi ritmikus vándorlást csak a felszín közeli másfél méteres vízrétegben tapasztalt. A hullámzás megváltozatta e napi ritmust, de a napi vertikális vándorlás fő hajtóerejének a fényváltozást tartotta. Ponyi & H. Péter (1986) hosszan tartó szél és hullámzásmentes időben folytatott vizsgálatai során az Eudiaptomus gracilis copepodit és adult stádiumainak zöme nappal 1 méteres vízmélységben tartozódott, az éjszaka során le, illetve felfelé vándorlás révén egy kiegyenlítődés következett be az egyes vízrétegek között. A nauplius lárvák nappal a felszín közelében tartózkodtak, éjszaka egy intenzív lefelé vándorlás folytán minden rétegben egyenletesen oszlottak el. A Balatonhoz hasonlóan sekély tihanyi Belső-tóban, amely átlagosan 150 cm mély a Copepoda és a Cladocera fajok napszakos változásában két szakaszt különböztethető meg, egy nappali mélységi sűrűsödési és egy éjszakai felfelé vándorlási periódust (Ponyi & Tamás 1964).
6.2.4 A turbulencia hatása a zooplanktonra
Az élénk hidrodinamikájú Balaton gyakran felkeveredő, magas ásványi anyag tartalmú üledéke befolyásolja a szűrő zooplankton szervezetek táplálkozását. A vízben a táplálékszemcsék szuszpendált formában vannak jelen, amelyeket az állatnak össze kell gyűjtenie. Ezért a bentikus és a pelágikus organizmusok között sok a szűrű szervezet. A Cladocera rákok többsége szűrő szervezet, de a Copepoda rákok képviselői közül is sokan ezzel a táplálkozási formával jutnak táplálékhoz. A filtráláshoz szükséges egy vízáramlás, ami a táplálékszemcsét az állat szűrőkészülékéhez szállítja.
Ez a vízáramlás keletkezhet a víztömeg önálló mozgásából, vagy az állat saját maga hozza létre. A szűrű szervezetek táplálkozási szokásaiban két eltérő módot figyelhetünk meg. Az egyik esetében a szűrés teljesen mechanikus folyamatként megy végbe. A „fésűfogakon”
folyamatos vízáramlás közben minden megfelelő nagyságrendű részecske fennakad, tekintet nélkül arra, hogy az adott részecske táplálék vagy sem. A másik típus esetén azonban az állatok aktívan vadásznak, és a szűrendő táplálékot első csápjuk érzősörtéivel vagy tapintással, ill. áramlás érzékelése útján, vagy a már néhány mm távolságból érzékelt kémiai ingerek hatására válogatják ki.
A legtöbb Cladocera levéllábai egy bonyolult szívó-nyomó-pumpát képeznek, aminek a segítségével a víz a harmadik és a negyedik pár lábakon található szűrőfelületen keresztülpréselődik (4. ábra).
4. ábra: A Cladocerák szűrőkészüléke (Lampert & Sommer 1999)
A Copepoda rákok némelyikét bár a szűrő szervezetek közé soroljuk, de a szó szoros értelmében nem filtrálnak, hanem egyes táplálékszemcséket az általuk keltett vízáramból veszik fel (G.-Tóth & Zánkai 1985). A Daphnia fajokkal ellentétben a Copepoda rákok szelektív szűrők. Szűrőkészülékük kisebb, és nem kiszűrik vele a táplálékszemcsét, hanem megragadják. A száj módosult végtagjai vízáramlatot keltenek. Ha tehát például egy alga sodródik a Copepoda felé, akkor az állat a vízáramot olyan irányba hajtja, hogy az alga a második állkapocs közelébe kerülhessen, amivel aztán megragadhatja. Tehát a felvehető táplálékszemcse mérete nem a szűrőkészülék lyukméretétől függ, hanem attól, hogy hogyan képes megragadni és elfogyasztani a Copepoda a táplálékát. A Copepoda rákok többnyire nagyobb táplálékszemcséket fogyasztanak, mint a Cladocera szervezetek. A táplálékszemcsék maximális méretét a rákok rágójának vagy a carapax nyílás (Cladocera) résszélessége határozza meg. A kisebb méretű Cladocera rákoknál ez a felső határ 20 µm, a nagyobb Cladocera és Copepoda rákoknál pedig 50 µm.
A Balaton ásványi lebegőanyag tartalma az üledék gyakori felkeveredése és a biogén mészkiválás miatt magas, 10–600 mg l-1 száraz tömeg. A lebegő, elsősorban karbonát részecskék méreteloszlása 0,2–10 µm, amely a Daphnia-félék táplálék méretspektrumába (Geller & Müller 1981; Lampert & Sommer 1999) esik. A tó lebegő anyagának összetétele szerint a Balatonban élő Daphnia–féléknek több ezer ásványi szemcsét kell visszatartaniuk ahhoz, hogy egy vízigomba egyedet, 100-1000 szemcsét ahhoz, hogy egy detritusz részecskét, 50–500 szemcsét ahhoz, hogy egyetlen algaegyedet, s legalább egy szemcsét ahhoz, hogy egyetlen baktériumsejtet bekebelezhessenek (Entz & Sebestyén 1942; Entz 1979, 1981; G.-Tóth 1982, 1984, 1988). A balatoni Daphnia cucullata és Daphnia galeata béltartalom összetételének fény- és scanning elektronmikroszkópikus vizsgálata alapján a Balaton nagy ásványi lebegőanyag tartalma a Daphnia-félék számára táplálkozási problémákat jelenthet. Az átvizsgált béltartalom főleg 2–5 µm-es kalcium- és magnéziumkarbonát kristályszemcsékből állt. A béltartalmakból kimutatott algaszám és a fitoplankton vízben mért sűrűsége közötti pozitív lineáris kapcsolat pedig az algafogyasztás limitáltságát bizonyította az alga sűrűség által, pedig az önmagában többszöröse volt a táplálék felvételi felső küszöbkoncentrációnak (G.-Tóth 1982, 1984, 1988; G.-Tóth et al. 1987). Zánkai (1981) kimutatta, hogy a Balatonban a Daphnia–félék béláthaladási ideje 2–20-szor rövidebb, mint az abiszesztonban szegény tavakban.
Ugyanakkor, a ballaszt táplálék miatt megnövekedett bélürülési ráta az emésztésre fordított időt, és így feltehetően annak hatékonyságát is csökkentheti. A balatoni Daphnia galeata intenzív abioszeszton szűrésére jellemző, hogy az ásványos béltartalom időnként az állatok teljes száraztömegének 40%-át is elérte (G.-Tóth et al. 1986). Jól ismert a Daphnia- féléknek az a képessége, hogy a szűrés intenzitását a tápanyag függvényében változtatni tudják, így változó tápláléksűrűség esetén is közel azonos mennyiségű táplálékot vesznek fel bizonyos határok között (Cannon 1933; Rigler 1961; McMahon & Rigler 1965;
Lehman 1976; Watts & Joung 1980). Vizsgálatok bizonyították, hogy az állatok szűrése az abioszeszton koncentráció növekedésével rohamosan csökken (G.-Tóth 2000). Hasonlóan reagált tehát, mint tényleges táplálék koncentráció változásra, amely automatikus szűrési módjukból fakadt. G.-Tóth (2000) számításai szerint a zooplankton a Balatonban az abiszesztonon kívül saját száraz tömegének 10%-át kitevő baktériumot, 50%-ának megfelelő algát és 4,8-szeresét kitevő detrituszt szűrt ki naponta.
A fitoplankton és a baktériumok asszimilációs hatásfokát 25%-nak (Zurek 1976; Infante 1978; Holm et al. 1983), a detrituszét 10%-nak (Saunders 1972; Wetzel 1983; Arruda et al.
1983) véve a napi asszimilált mennyiség 60,5 testtömeg %-nak adódik. Muck és Lampert szerint (1984) a Daphnia-félék 19°C-on saját testtömegük mintegy 85%-ának megfelelő mennyiséget asszimilálnak naponta. Azt is figyelembe véve, hogy a balatoni állatoknál az emésztésre 2-20-szor kevesebb idő jut, mint az abioszesztonban szegény tavakban, arra a következtetésre kell jutnunk, hogy a Balatonban a Daphnia-félék nem jutnak kellő mennyiségű táplálékhoz.
Ghilarov (1985) a Moszkva közelében található Glubokoye-tóban, amely abioszesztonban szegény, a D. galeata posztembrionális fejlődési idejét nyáron 5, máskor 5-22 napnak találta. G.-Tóth (2000) kísérletei azt mutatták, hogy a balatoni D. galeata kifejlődési ideje 20–23,5°C-on szintén 5-5,5 nap lehetne akkor, ha nem lenne abioszeszton a vízben, csak a táplálék. Minden 10 mg l-1 abioszeszton növekedés azonban 2,6 órával nyújtotta meg a posztembrionális fejlődési időt az effektív táplálék viszonylag állandó koncentrációja mellett. Hasonló értelmű változást mutatott a fekunditás csökkenése is a növekvő abioszeszton koncentráció mellett. Lampert (1978), Geller (1985), Duncan et al. (1987) és mások szerint optimális táplálékellátottság mellett a D. galeata-hoz hasonló méretű Daphnia-félék fekunditása 7-10 pete nőstényenként. G.-Tóth vizsgálatai alapján az abioszeszton-fekunditás funkcionális összefüggés szerint a balatoni D. galeata fekunditása csak a tavi plankton és detritusz táplálékán is elérné a 6-7 pete nőstényenként értéket, ha nem lenne jelen az abioszeszton (G.-Tóth et al. 1986). Annak jelenlétében azonban rohamosan csökken a fekunditás, 6-10 mg l-1 abioszeszton mellett 2-5, 40-50 mg l-1 mellett 1-3, 200-250 mg l-1 mellett pedig 1-2 pete értéket kapunk nőstényenként. Présing (1986) nőstényenként 2,2 – 2,6 pete értékben állapította meg a Balatonban gyűjtött D. galeata fekunditását. Zánkai és Ponyi (1997) későbbi vizsgálatai során a Daphnia fajok (D. galeata és D. cucullata) átlagos nőstényenkénti peteszámát Keszthelynél 2,6, míg Tihanynál 1,2 találták.A hordaléktól gyakran zavaros vizekben általános tapasztalat, hogy az abioszeszton növekedésekor csökken a zooplankton sűrűsége, eltűnnek a Cladocera fajok, vagy a zavaros vizű időszakban a zooplankton ki sem fejlődik (Thienemann 1911; Berner 1951; Herzig 1979; Kerfoot 1980). Nagy valószínűséggel kimondhatjuk, hogy a Balatonban a Daphnia-félék krónikusan gyér állományának és alacsony fekunditásának oka a magas abioszeszton tartalom.
A vízben élő mikroszkópikus méretű állatok számára létfontosságú lebegőképességük fenntartása. A víz sűrűségéhez a zooplankton lipidek szervezeten belüli felhalmozásával (Campbell & Dower 2003), úszással, és felület nagyobbító képletek kialakításával alkalmazkodik. Erős turbulens áramlások esetén a zooplankton számára nehézségbe ütközhet a lebegőképesség fenntartása. Az áramlás a napi vertikális vándorlást végző planktonszervezeteket kedvezőtlenebb vízmélységbe sodorhatják, minek következtében jobban ki vannak téve a predációs nyomásnak, és a táplálékhiánynak.
A tavak turbulenciáját legnagyobb részben a szél keltette energia alakítja, hatására a vízben turbulens kinetikai energia disszipálódik. Az energia disszipálódásának rátáját a kinematikai viszkozitás és a vízmélység szabja meg. A turbulencia energiaspektum függvényében általában három jellemző tartomány van. Az első a nagyméretű örvények tartománya. A második, mikor a mozgási energia átadódása révén az örvény struktúrája a nagyobb méretűből a kisebbek felé tolódik el, de nem történik jelentős energiaveszteség.
Ilyenkor a kinetikai energia még nem vész el, mivel a nagyobb örvényből nagyobb számú kisebb örvény alakul ki, melyeknek ugyan kisebb az energiájuk, de összességében kiadják a nagyobb örvényből adódó mennyiséget. A több, egy adott térfogaton belüli kisebb örvény keletkezése következtében azonban térfogategységenként nézve fajlagosan egyre nagyobb lesz az örvények közötti érintkezés, szükségképpen a súrlódó, nyíródó határfelület. Ennek következtében harmadik lépésben eljutunk az olyan kis méretű örvények keletkezéséhez, melyeknél a nyírófelület olyan nagy lesz, hogy a viszkózus erők kezdenek dominálni a tehetetlenségi erőkkel ellentétben, és az energia disszipálódik (lásd pl. Sokoray-Varga & Józsa 2006).
A turbulencia zavaró hatását a zooplankton fejlődésére, illetve a víztérben való vertikális elhelyezkedésére már bizonyították (Mackas et al. 1993; Lagadeuc et al. 1997; Incze et al.
2001). Kimutatták, hogy a planktonszervezetek nagysága egy adott víztestben nem lehet nagyobb a turbulens áramlások legkisebb örvény átmérőjénél (Kolmogorov-távolság).
Ellenkező esetben egy kritikus érték felett az érzékenyebb planktonszervezeteket a rájuk ható nyírófeszültség roncsolja (Margalef 1978; Reynolds 1992a,b, 1998; O’Brien 2002;
O’Brien et al. 2004). A tavi plankton méreteloszlása ezért tolódik el tartós turbulens hatásra a kisebb mérettartomány felé (Herzig 1979; Alcaraz & Saiz 1991; Moisander et al.
2002).
A gyenge áramlás elősegíti a ragadozó fajok prédaállatokkal való találkozási esélyeit, a lebegő algák és baktériumok sodródását a szűrő planktonszervezetek szűrőkészülékéhez, valamint az állatok mozgását. Vizsgálatok bebizonyították, hogy egyes tengeri Copepoda fajok nauplius lárváinak fejlődési ideje másfélszer-kétszer rövidebb ideig tart turbulens, mint a pangó vízben (Saiz & Alcaraz 1991; Visser & Stips 2002; Visser et al. 2009). Az erősebb áramlási viszonyok azonban csökkentik a ragadozó fajok predációs sikerét (Rothschild & Osborn 1988; Jonsson & Tiselius 1990; MacKenzie & Leggert 1991), a napi vertikális vándorlást végző fajok estében gátolja a vándorlást, így erősebb predációs nyomásnak és táplálékhiánynak lehetnek kitéve. A szűrő szervezetek estében az erős áramlás zavarhatja a szűrés folyamatát, mivel a turbulens örvények interferálhatnak az állat által keltett táplálékszerző örvényekkel (Rotschild & Osborn 1988; Alcaraz et al. 1994;
Kiørboe & Saiz 1995; Visser & Stips 2002; Visser et al. 2009).
7 Célkitűzés
Kutatásunk célja a sekély, élénk hidrodinamikájú Balatonra jellemző turbulencia viszonyok és egyéb környezeti paraméterek feltérképezése, hatásuk vizsgálata volt a zooplankton társulás szerkezetére, elhelyezkedésére, a különböző fajok és egyedfejlődési stádiumaik fejlődési sebességére. A vizsgálatok kiterjedtek a tó eltérő környezeti adottságokkal rendelkező víztereire, valamint laboratóriumban hullámzás generátor műszerrel folytatott kísérletekre. A kutatási irányok a következők voltak:
1. A zooplankton minőségi és mennyiségi viszonyainak elemzése a Balaton eltérő kinetikai tulajdonságokkal rendelkező élőhelyein (pl. nádas, litorál, nyíltvíz);
2. A turbulencia a tó zooplankton állományára gyakorolt hatásának feltérképezése más környezeti paraméterek (pl. hőmérséklet, vízátlátszóság) jelentőségéhez képest;
3. A zooplankton vertikális vándorlásának 24 órás vizsgálata eltérő turbulencia viszonyok mellett (erősen szeles és szélcsendes időben);
4. Kísérleti rendszerben a turbulencia hatásának vizsgálata a különféle Cladocera és Copepoda rákok egyedfejlődésére és mortalitására.
8 Anyag és módszer
8.1 Zooplankton mintavétel és a minták feldolgozása
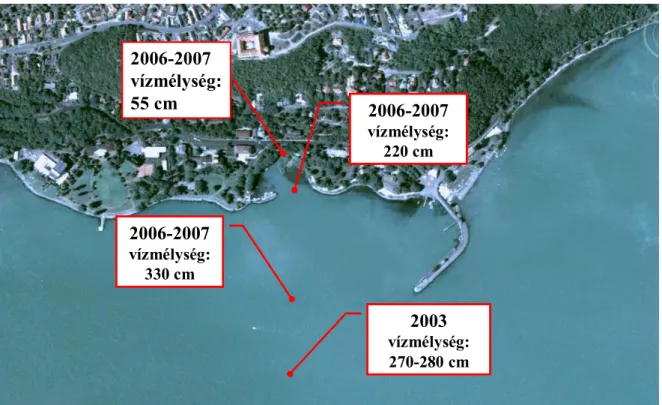
A rákplankton vizsgálata többféle nehézséget hordoz magában. Az egyik az állatok napszakos mozgása. Az állóvizekben napszaktól függően különféle mélységben kialakuló rajokban élhetnek, ezért ha meg akarjuk figyelni az elterjedésüket, akkor a teljes vízoszlopot reprezentáló mintát kell gyűjteni. Ugyanakkor testméretük apró, legfeljebb 5 mm-esek, életpályájuk viszonylag rövid, gyorsan ivaréretté válnak, akár már az első héten képesek utódokat létrehozni (pl. Naidenow 1998), mely indokolttá teszi a gyakori mintavételt (Ponyi et al. 1982; Bothár 1988; Bothár & Kiss 1990). Ezért, valamint az állatok évszakos változása, a vizsgálandó élőhelyek változatossága miatt heti mintavételezést végeztünk a tó eltérő sajátságú medencéinek különböző vízmélységű pontjain (5., 6. ábra).
Mintát a felszíntől lefelé félméterenként, 60 µm lyukbőségű planktonhálóval ellátott 50 cm magas és 34 liter térfogatú Schindler-Patalas-féle mintavevővel vettünk. Az egyes mintákat 100 cm3 térfogatban tömörítve 3,5 % végkoncentrációjú formalinnal tartósítottuk és Zeiss- Opton fordított planktonmikroszkóppal dolgoztuk fel. Minden esetben 5 ml-es almintákat véve minimum három párhuzamos számolást végeztünk. A meghatározás Gulyás & Forró (1999), Einsle (1993) és Flöβner (2000) határozókönyvei alapján történt. Az egyedsűrűséget egyed l-1 egységben adtuk meg. Eltérő időpontokban a különböző mérőpontokon összesen 119 alkalommal végeztünk gyűjtést.
2006-2007 vízmélység:
220 cm
2003 vízmélység:
270-280 cm 2006-2007
vízmélység:
330 cm 2006-2007 vízmélység:
55 cm 2006-2007
vízmélység:
220 cm
2003 vízmélység:
270-280 cm 2006-2007
vízmélység:
330 cm 2006-2007 vízmélység:
55 cm
5. ábra. Mérőpontok az MTA ÖK Balatoni Limnológiai Kutatóintézet (Tihany) előtt
6. ábra: Mérőpontok a Balaton hossztengelye mentén (Keszthelyi-, Szigligeti-, Szemesi-, Siófoki-medence)
8.2 A zooplankton tér- és időbeni eloszlásának vizsgálata a Balatonban
A balatoni zooplankton szerkezetét 2006. áprilisától 2007. októberéig heti gyakorisággal vizsgáltuk különböző vízrétegekben az MTA ÖK Balatoni Limnológiai Kutatóintézet (BLKI) előtti litorálban három helyen: a nádasban (GPS adatok: 46o54′50.97″N, 17o53′33.65″E, vízmélység 55 cm), valamint onnan 25 m (GPS adatok: 46o54′52.7″N, 17o53′36.3″E, vízmélység 220 cm), illetve 200 m távolságban (GPS adatok: 46o54′51.4″N, 17o53′46.4″E, vízmélység 330 cm). (5. ábra). Emellett vizsgáltuk a zooplankton közösségek szerkezetét a Balaton hossztengelye mentén 5 mérőponton: Keszthely (vízmélység: 310-330 cm), Szigliget (400-410 cm), Zánka (430-440 cm), Tihany (410-430 cm), Siófok (430-460 cm) (6. ábra) térségében, szintén vízrétegenkénti bontásban havi gyakorisággal.
8.3 A zooplankton napszakos vándorlásnak vizsgálata
2003-ban két 24 órás vizsgálat zajlott, szélcsendes, derült időben a Kutatóintézet előtt, a parttól kb. 800 m távolságban (vízmélység 270-280 cm).
A mérésekre augusztus 4.-5.-én és augusztus 25.-26.-án került sor, 12, 15, 19, 23, 3, 7, 10, 13, 15 órai időpontokban. 2003-ban turbulencia mérés még nem volt.
2007-ben augusztus 18.-19.-én, 2 mérőponton, a Kutatóintézet előtt 220 és 330 cm vízmélységű vízben 17, 19, 3.30, 6.30, 11, 14.30 órakor történtek gyűjtések és turbulencia mérések. A 2007-es vizsgálat időpontját időjárási prognózis alapján úgy választottuk ki, hogy alkalmunk nyíljon a nap során egy előre jelzett, és beköszönő erős szél hatását megfigyelni a balatoni zooplankton vertikális szerkezetére. Az erősen szeles időjárás 18.- án este 20 óra után alakult ki. A vizsgálatokat a különböző vízrétegek turbulenciája mérésével is kiegészítettük. Mind a mintavétel, mind a minták feldolgozása, valamint a turbulencia mérés megegyezett a heti mintavételeknél alkalmazott módszerekkel. 2007-ben az egyik mérőponton két mintavétel kimaradt az erős hullámzás miatt.
A 24 órás mérések során a korábbi irodalmi adatokra támaszkodva (Entz & Sebestyén 1942; Ponyi & Tamás 1964; Ponyi & H. Péter 1986) a Daphnia cucullata x galeata, és az Eudiaptomus gracilis vertikális vándorlást vizsgáltuk. Meghatároztuk a zooplankton populációkra jellemző mindenkori átlagos tartózkodási mélységet (mean residence depth:
MRD; Armengol & Miracle 2000):
∑
∑
×=
i i i
N d MRD (N )
ahol Ni az i-edik mélységben mért egyedkoncentráció, és di az i-edik minta mélysége. A kapott MRD érték a populáció képzeletbeli súlypontját adja meg, azt jelzi, hogy a populáció nagy része milyen mélyen tartózkodik az adott időpontban.
8.4 A vizsgált környezeti változók
A zooplankton mintavétellel párhuzamosan a következő környezeti paramétereket vizsgáltuk Horiba U-10 típusú vízanalitikai mérőműszer segítségével: vízmélység, hőmérséklet, vezetőképesség, pH. Secchi-koronggal vizsgáltuk a víz átlátszóságát. Mértük továbbá a különböző vízrétegekben a turbulencia értékeket (lásd lentebb). A turbulens kinetikai energiadisszipációs ráta és a Kolmogorov-távolság számításhoz a szélsebesség adatokat az Országos Meterológiai Szolgálat Siófoki Obszervatóriumától szereztük be.
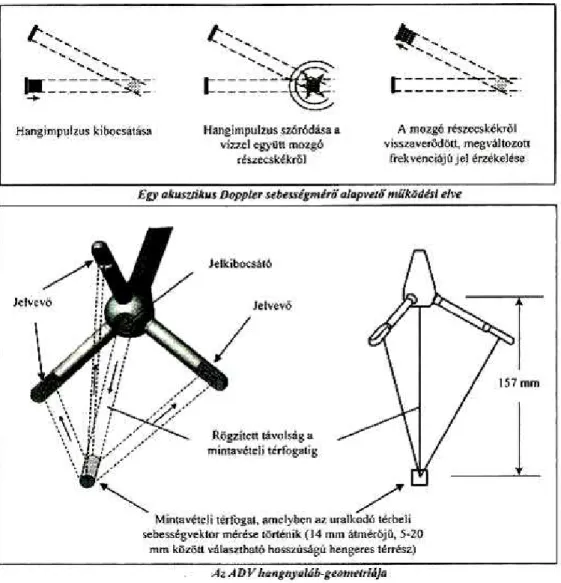
8.4.1 Turbulencia mérés és a velociméter bemutatása
A turbulencia mérés a zooplankton mintavétellel térben és időben párhuzamosan történt, 16 MHz-es MicroADV 3 D SonTek típusú Akusztikus Doppler Velociméterrel (ADV) (7.
ábra).
7. ábra: MicroADV 3 D SonTek velociméter
A műszer adatai a következők:
Paraméter 16-MZh Micro ADV
Mintavételi ráta (Hz) 0,1-tól 50-ig
Mintavételi térfogat (cm3) kb. 0,3
Mintavételi térfogattól való távolság (cm) 5
Felbontóképesség (cm s-1) 0,01
Programozható sebesség értéktartomány (cm s-1) 3, 10, 30, 100, 250
Pontosság a mért sebesség 1%-a, ± 0,25 cm s-1
Az Akusztikus Doppler Velociméter a víz sebességét az úgynevezett Doppler-effektus révén méri (8. ábra). Az akusztikus mérőműszerek előnye, hogy nem mechanikus, forgó alkatrészek révén mérik a sebességet, hanem a hang különféle összetételű vízben való terjedési sebességére alapozva, így a műszer mérési pontossága sokkal jobb. Másik előnye, hogy az akusztikus jel kibocsátása révén anélkül képes mérni, hogy a vizsgálandó közeget közvetlenül zavarná (Sokoray-Varga & Józsa 2006).
8. ábra: A velociméter működése és felépítése. Forrás: Sokoray-Varga & Józsa (2006)
A velociméter egy jelkibocsátó és –vevő műszerfejből, energiaellátó tápegységből és vezérlő, jelfeldolgozó és memóriaegységből áll, és számítógéphez csatlakoztatva lehet működtetni. A műszer belső iránytűje és dőlésérzékelője lehetővé teszi, hogy az eredményeket földi koordinátarendszerbe transzformálva lehessen rögzíteni.
A velociméterrel kapott regisztrátumok a vízzel együtt mozgó részecskék Descartes-féle 3 térdimenzióban (X, Y és Z) történő egyidejű elmozdulási sebességét adják. Ezekből kell kiszámítani a részecske tényleges elmozdulási irányát és sebességét (az áramlási sebességvektort, mint a tér három irányában mért komponensek eredőjét). Erről a SonTek kezelőprogram először egy sebesség eloszlási függvényt ad meg az előre megválasztott, esetünkben a 10 perces mérési időszakokra. Ebből számítható a vízáramlások átlagos sebessége a mérések ideje alatt.
A mérések során a műszer szondafejét a kívánt mélységnek megfelelően egy 5 m hosszú rúdra rögzítettük, amelynek a végén nehéz vasgallérral ellátott ásófej helyezkedett el. A szondafej az aljzattól és a rúdtól való megfelelő távolságát egy csúsztatható fémrögzítés biztosította. A szondafej rögzítése után a mérések alkalmával a csónakból a rudat a függőlegesen az üledékbe döftük. A vízoszlop fél méteres mélységeiben 10-10 perces mérést végeztünk a zooplankton mintavétellel párhuzamosan 50 Hz-en.
A turbulenciát a 3D turbulens sebesség-fluktuációk négyzetes középértékének gyökével (Root Mean Square, RMS-turbulence) jellemeztük és cm s-1 mértékegységben adtuk meg (Tennekes & Lumley 1972; Reynolds 1992b).
[ ] ( ) ( )
1 ' /
2 2 ' 2
−
= −
=
∑ ∑
n
n V V V
Vx
RMS x x x
A velociméterrel kapott 3D sebesség-idősorokat WinADV szoftverrel dolgoztuk fel.
A 3D sebesség-idősorokra közvetlenül alkalmazott RMS számítás a turbulencia mellett jelen levő áramlási összetevők közül a hosszú periódusidejű lengések és a köröző áramlások hatását lényegileg kiszűri. Nem képes azonban kiszűrni a hullámzást, amely nagyságrendjében a Balatonon másodperces periódusidejű felszínmozgást kísérő periodikus, az egyes vízrészecskékre nézve orbitális pályán zajló, jelentős sebességekkel kísért vízmozgás-összetevő. A 3D sebesség-idősorok dekomponálása a közelmúltban tett sikeres kezdő lépéseket Homoródi et al. (2012).
A turbulens kinetikai energiadisszipációs rátát és a Kolmogorov-távolságot különböző szélsebességek és vízszintek mellett Tennekes & Lumley (1972) és MacKenzie & Leggett (1993) nyomán számoltuk ki.
A turbulens kinetikai energiadisszipációt (ε, m2 s-3) a mintavételi pontok átlagos vízmélységével (Z) és a szélsebességgel (W) számoltuk MacKenzie & Leggett (1993) után:
( )
z W s
m m W z
C W w a
D
3 6 3
2 3 3
2 3
10 045 , 001 6
, 0
1 4
, 0
−
−
−
×
=
×
×
= ρ ε ρ
A szél sebességét (W) a következő értékekben adtuk meg: 0,5; 1; 2; 4; 8; 16 és 32 m s-1. A tó átlagos mélységeiként mintavételi pontjaink átlagos mélységével számoltunk, azaz 0,5;
2,2; 3,3; 4; 4,3; 4,45 m-rel. A levegő sűrűsége (ρa) 1,2 kg m-3, a víz sűrűsége (ρw) 998 kg m-3, a vízfelszín és a szél közötti közegellenállási tényező (CD) 0,0015 és a von Karman állandó 0,4.
A turbulens kinetikai energiadisszipációs rátát a mintavételi pontok közepes vízmélységében a Taylor-egyenlettel (Luettich & Harleman 1990) is meghatároztuk:
1
3 −
×
= A RMS l ε
ahol az RMS az ADV velociméter által mért RMS-turbulencia, az A állandót 1-nek, és a helyi hosszmértéket (l) 25 cm-nek feltételeztük, ami a Balaton átlagos hullámmagassága (G.-Tóth et al. 2011).
A Kolmogorov-távolságot (ήk, mm) a disszipációs rátából (ε) és a kinematikus viszkozitásból (ν = 1,1 10-6 m2 s-1) számoltuk Tennekes és Lumley (1972) után:
4 1 3
=
′ ε ηk v
8.5 Turbulencia generátoros kísérletek
O’Brien et al. (2004) nyomán a Pannon Egyetem Alkalmazott Gépészet Intézeti Tanszékének (Veszprém) közreműködésével turbulencia generátort (9. ábra) terveztettünk és készítettünk, hogy a turbulencia zooplanktonra gyakorolt hatását laboratóriumi körülmények között is igazoljuk, illetve kísérletileg is bizonyítsuk.
9. ábra: Turbulencia generátor 10. ábra: A mesterséges turbulencia keltésére szolgáló rácsszerkezete
2006. októbere és 2007. szeptembere között kilenc, 18-21 napos kísérletsorozatot hajtottunk végre a 64x64x96 cm-es, turbulencia generátorral ellátott kísérleti, valamint egy kontroll akváriumban.
A kontroll és a kísérleti akváriumokat 80 cm-es magasságig a Balatonból származó vízzel töltöttük fel, melyet 24 órán át ülepítettünk, és a kiülepített ásványi lebegőanyagot szívással óvatosan eltávolítottunk. A víz felső rétegében mozgott vertikális irányban a turbulencia generátor műanyag rácsozata (lyukbőség 5,8 x 5,8 cm) 1,033 s-1 frekvenciával (Giovanni elektromos motor), 5 illetve 10 cm-es amplitúdóval (10. ábra). A kísérleteket megelőzően 3D Akusztikus Doppler Velociméterrel mindkét amplitúdó alkalmazása mellett feltérképeztük az akvárium turbulencia viszonyait, amelyet 10 cm-es alapkiosztás mellett bizonyos rácspontokat kihagyva egymástól 10-20 cm távolságra elhelyezkedő képzeletbeli rácspontok mentén mértünk. Az előzetes mérések alapján a 10 cm-es amplitúdójú rácsmozgás használata mellett döntöttünk, mely leginkább modellezte a Balatonra jellemző turbulencia viszonyokat. Az akvárium vízoszlopában így az RMS- turbulencia értékek, a kiszámított energiadisszipációs ráta, és a Kolmogorov-távolság a Balaton közepes mozgású vizének felelt meg.