PANNON EGYETEM GEORGIKON KAR
KESZTHELY
Állattudományi és Állattenyésztéstani Tanszék
DOKTORI (Ph.D.) ÉRTEKEZÉS
Állat- és Agrárkörnyezet-tudományi Doktori Iskola Iskolavezető: Dr. Anda Angéla, D.Sc.
(Jogelőd: Állattenyésztési Tudományok Doktori Iskola Iskolavezető: Dr. Szabó Ferenc D.Sc.)
A PONTY (Cyprinus carpio L.) PIKKELYMINTÁZAT ÖRÖKLŐDÉSI MODELLJÉNEK REVÍZIÓJA
Témavezetők:
Dr. habil. Bercsényi Miklós egyetemi tanár Dr. Nagy Szabolcs egyetemi docens
Társkonzulens: Dr. Orbán László
Készítette:
Cserveni-Szücs Réka
Keszthely 2013
2
3
A PONTY (Cyprinus carpio L.) PIKKELYMINTÁZAT ÖRÖKLŐDÉSI MODELLJÉNEK REVÍZIÓJA
Értekezés doktori (PhD) fokozat elnyerése érdekében Írta:
Cserveni-Szücs Réka
Készült a Pannon Egyetem Georgikon Kar
Állat-és Agrárkörnyezet-tudományi Doktori Iskolája Keretében Témavezetők: Dr. habil. Bercsényi Miklós egyetemi tanár
Dr. Nagy Szabolcs, egyetemi docens
Elfogadásra javaslom (igen / nem) ………...
(aláírás) Elfogadásra javaslom (igen / nem) ………...
(aláírás) A jelölt a doktori szigorlaton ...%-ot ért el,
Az értekezést bírálóként elfogadásra javaslom:
Bíráló neve: …... …... igen /nem ……….
(aláírás) Bíráló neve: …... …...) igen /nem ……….
(aláírás) ***Bíráló neve: …... …...) igen /nem ……….
(aláírás) A jelölt az értekezés nyilvános vitáján …...%-ot ért el.
Veszprém/Keszthely, ……….
a Bíráló Bizottság elnöke A doktori (PhD) oklevél minősítése…...
………
Az EDHT elnöke
4
Tartalomjegyzék
Rövidítések jegyzéke ... 6
Kivonat ... 7
Abstract ... 9
Auszug ... 10
1. Bevezetés és célkitűzések ... 12
2. Irodalmi áttekintés ... 15
2.1. A ponty rendszertana ... 16
2.2. A ponty morfológiája /felépítése ... 18
2.3. A ponty eredete /elterjedése ... 20
2.4. Nemesített és vad ponty változatok ... 25
2.4.1. Tájfajták és hibridek kialakulása Magyarországon ... 26
2.4.2. Tájfajták és tenyésztett változatok pikkelyezettsége ... 27
2.5. A ponty evolúciós ugrása – a tetraploid állapot és az ebből adódó lehetőségek .. 28
2.5.1. Egy család- 3 különböző genom méret ... 28
2.5.2. Genom duplikáció a csontos halakban ... 29
2.6. Kültakaró és fenotípus ... 31
2.6.1. A pikkely és eredete: ... 33
2.7. A garatfog ... 35
2.8. A kopoltyú és részei ... 36
2.9. A ponty pikkelymintázatának genetikája anno: ... 37
2.10. Az „s” gén azonosítása ... 41
2.11. A pikkelyezettség kutatásakor eddig vizsgált és az N gén megtalálása szempontjából is jelentősebb szignál transzdukciós kaszkádok ismertetése ... 43
2.11.1. Fibroblaszt növekedési faktor transzdukciós kaszkád ... 44
2.11.2. Az ’ectodysplasin’ anyagcsereútvonal ... 49
2.11.3. Egyéb, szóba kerülhető kaszkádok ... 51
3. Anyag és módszer ... 52
3.1. Az anyapontyok kiválasztása, keresztezésük ... 52
3.1.1. Magyarországi keresztezések ... 52
3.1.2. Szingapúri keresztezések: ... 56
3.2. Az ikra felragasztása, termékenyülés számolása: ... 58
5
3.3. A kísérleti halak nevelése: ... 59
3.4. A fenotípus kiértékeléséhez szükséges egységes rendszer kialakítása ... 60
3.5. Kopoltyúfésűk tüskéinek számolása, az úszók szemrevételezése ... 62
3.6. Garatfog kiszedése, tisztítása ... 63
3.7. Eltérő pikkelyezettségű ponty változatok keresztezéséből származó utódnemzedék növekedésének vizsgálata zárt tartásban ... 64
3.8. Úszóminta gyűjtése és a nukleinsavak izolálása genotipizáláshoz, szekvenáláshoz ... 65
3.9. Szekvenálás és a jelölt gének cDNS-ének összehasonlító vizsgálata ... 66
3.10. Az alkalmazott statisztikai eljárások és programok ... 66
4. Eredmények és értékelés ... 67
4.1. A keresztezések eredményei Magyarországon: ... 67
4.2. A pikkelyezettséggel összefüggő más paraméterek, szervek vizsgálati eredményei ... 70
4.2.1. Az úszók, a kopoltyúk, valamint a garatfogak szerkezete ... 70
4.2.2. Eltérő pikkelyzettségű ponty változatok keresztezéséből származó utódnemzedék növekedésének kiértékelése ... 72
4.3. A szingapúri keresztezések kiértékelése ... 73
4.4. A Magyarországon és Szingapúrban elvégzett 19 keresztezés ... 75
4.5. Eltérés a Magyarországon és Szingapúrban végzett keresztezések bőrpontyainál: ... 76
5. Kiegészítő vizsgálatok ... 78
5.1. Genetikai vizsgálatok Szingapúrban ... 78
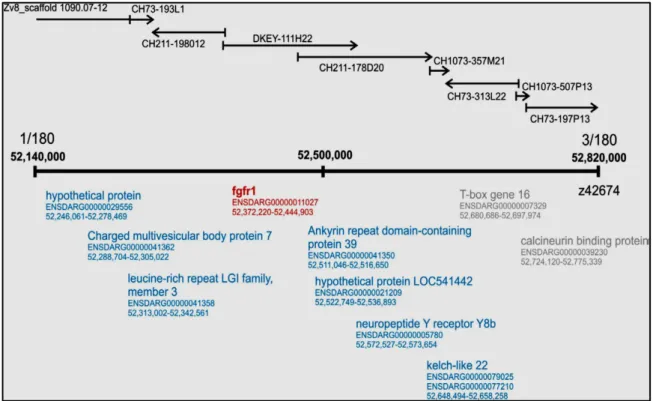
A ponty fgfr újabb paralógjainak felfedezése ... 80
5.2. Magyarországi kiegészítő vizsgálatok ... 80
5.2.1. A különböző fenotípusú pontyok pikkelyeinek alaktani vizsgálata ... 80
5.2.2. „Karcolásos kísérlet” ... 81
6. Következtetések és Javaslatok ... 83
7. Összefoglalás ... 88
Irodalomjegyzék ... 90
Tézispontok ... 108
Thesis Points ... 109
Köszönetnyilvánítás ... 110
Tudományos közlemények jegyzéke ... 111
Melléklet ... 114
6
Rövidítések jegyzéke
DAG: diacilglicerol
DDC: (Duplication- Degeneration- Complementation) duplikáció-degeneráció-komplementáció modell
EDA: ectodysplasin-A
EDAR: ectodysplasin-A receptor
EDARADD: EDAR-(associated death domain) kapcsolódó haláldomén FCR: takarmányhasznosítási érték
FGF: Fibroblast growth factor: Fibroblast növekedési faktor
Fgfr1: Fibroblast growth factor receptor 1: Fibroblast növekedési faktor receptor 1 HAKI: Haltenyésztési Kutató Intézet (Szarvas)
Ic: (incomplete scaled) nem teljesen pikkelyes IKK: IkappaB kináz (IKK)
IP3: inozitol 1,4,5-trisfoszfát Ir: (irregular) szabálytalan
Li: (linear) oldalvonalsoros, keretes m: metacentrikus
Mi: (mirror) tükrös
mRNS: hírvívő (messenger) RNS mtDNS: mitokondriális DNS NF: kromoszómakarok száma
NF-κB: aktivált B-sejtek nukleáris kappa-könnyűlánc-fokozó faktor Nu: (nude) bőr
PIT (passive integrated transponder) tag PIP2: foszfoinozitol 4,5-biszfoszfát R3: hal specifikus genom duplikáció Sc: (scaled) pikkelyes
SGR:specifikus növekedési ráta sm: szubmetacentrikus
SMMCI: (Solitary median maxillary central incisor syndrome): egy multiplex középvonalbeli rendellenesség, melyre egy, a maxillaris ív közepén lévő metszőfog utal.
st: szubtelocentrikus t: telocentrikus
TRAF6: TNF receptorhoz kapcsolt 6. faktor
7
Kivonat
A ponty (Cyprinus carpio L.) pikkelymintázat öröklődési modelljének revíziója.
Jelen értekezésben a szerző a gazdaságilag jelentős, Földünkön legszélesebb körben elterjedt édesvízi halfaj, a ponty (Cyprinus carpio L.) fenotípusos megjelenésének miértjeire, a pikkelyzet alakulásának genetikájára keresi a választ. Párhuzamot von más gerincesekben ugyanezen gének mutációi okozta rendellenességekkel, felhívva ezzel a figyelmet ezen gének fejlődésbiológiai jelentőségére is.
A közel 80 éve Kirpichnikov és Balkashina által előjelzett két gén (s és N) megismeréséhez jelen értekezés is hozzájárul. Indukált mutációval (forward genetic screening) létrehozott tükrös zebradánió (spigeldanio) segítségével egy német- amerikai -magyar kutatócsoport pontyban is azonosította a részleges pikkelyhiányért felelős „s”
gént. A mostani tanulmány főként a másik, teljes pikkelyvesztésért felelős gén, az „N”
gén megtalálására tett kísérleteket írja le. A szerző és munkatársai különböző fenotípusú pontyokat kereszteztek, majd vizsgálták az F1 és F2 nemzedék fenotípusos megoszlását, ennek genetikai hátterét.
A kísérleteket Magyarországon és Szingapúrban párhuzamosan végezték, összesen több, mint 30 keresztezést hajtottak végre. A vizsgálatok kiterjedtek a pikkelyméretek, úszósugarak/ úszódeformitások, garatfogak, kopoltyútüskék, és egyes termelési paraméterek vizsgálatára is.
A kísérletek közben derült fény az ázsiai és európai pontyfajtákban eltérő súlyosságú genetikai deformitást okozó, teljes pikkelyhiányért felelős feltételezett
„N”gén különbözőségeire is.
A kutatás céljai az alábbiak voltak:
Nemzetközi csoportokkal együttműködve felülvizsgáljuk és pontosítsuk a ponty pikkelymintázatának genetikájáról alkotott képet, és ha lehet, olyan új modellt alkossunk, amelyik a régihez képest jobban leírja a sokféle fenotípus megjelenését, és a korábbi modellben várt letalitások elmaradását.
Nemzetközi csoportokkal együttműködve kísérletet tegyünk a pikkelymintázatért felelős másik gén (az „N” gén) felkutatására.
8
Összefüggést keressünk a pikkelymintázat és más jellemzők, mint pl. a garatfogak, a kopoltyúfésű tüskéi vagy az úszók alakulása között.
Megvizsgáljuk a megszokottól eltérő (oldalvonalsoros, bőrponty) hazai pontyok tenyésztésének jogosultságát a növekedési, vagy megmaradási mutatók tekintetében.
A kutatás eredményeit az alábbiakban lehet összefoglalni:
Kirpichnikov modelljének újragondolása- a fenotípusos besorolás kibővítése.
Különbségek kimutatása az ázsiai és európai keresztezéseknél (erősebb – gyengébb N allél) - az úszók deformitása, garatfogak számának csökkenése átmenetet mutat.
Egy új fgfr1 paralóg, az fgfr1b teljes transzkriptumának izolálása
Javaslat egy új öröklődési modellre
9
Abstract
Re-visiting the inheritance of scale pattern of common carp (Cyprinus carpio L.).
In this thesis the author deals with the genetics of the scale pattern inheritance of the common carp. The carp is one of the world's most important species of freshwater fish farming. It has a long history of domestication. Numerous breeding lines and shapes have derived from its wild form.
The author is re-visiting the original model of Kirpichnikov, that proposed four major scale pattern types connected to the genetics of common carp scale pattern inheritance and two genes (s and N) regulating scale coverage in cyprinids. The author’s research group generated a large number of crosses between different carp phenotypes involving loss-of- scale mutants of European and Asian origin and worked out a new model for the objective classifying the phenotypes of F1; F2 offspring generations. In this thesis, the author describes the ratio of scale pattern phenotypes in offspring groups. The experiments show that varieties of the so-called scattered phenotype with a larger number of non-overlapping scales often appear in offspring. Therefore, they divided the scattered type into three sub- types: irregular, incomplete scaled and classical mirror. They also analyzed the survival rates of offspring groups potentially inheriting two N alleles and found distinct differences between Asian and European crosses, indicative of the presence of a strong N allele with homozygous lethality in the former and a weaker, non-lethal one in the latter.
Crosses were performed paralelly in Hungary and in Singapore.
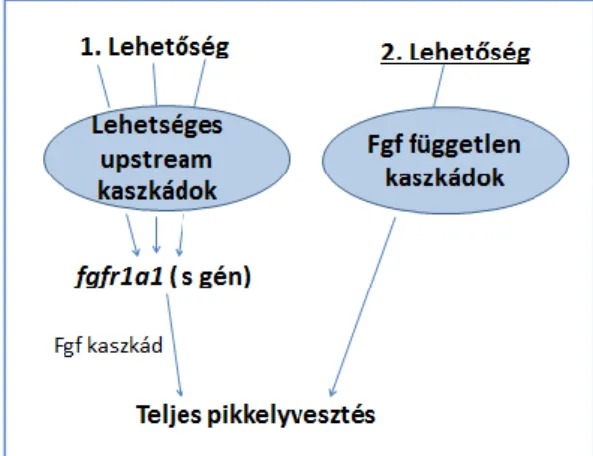
The current study describes the first steps of our continuing search towards the identification of the second gene (The “s” gene was found to be a paralog of fibroblast growth factor receptor 1, fgfr1a1.), called “N”. Important result in it, that in Singapore the research group isolated and structurally characterized a hitherto missing member of the Fgfr1 receptor family, fgfr1b, and showed that its sequence has not been mutated in nude individuals in comparison to mirrors. The research group proposes a new model that could explain the ‘deviating phenotypes’ observed in some of the crosses described above, and proposes that the new sub-types of scattered were formed due to increased levels of Fgf signals compared to mirrors and especially nudes, either due to an additional mutation in one of the FGF signaling pathway genes or, that in an upstream pathway, it functionally connected the Fgf signaling.
10
Auszug
Die Revision der Vererbungsmodell der Schuppenverbreitung des Karpfens (Cyprinus carpio L.).
Die Autorin befasst sich in ihrer Arbeit mit der Genetik der Schuppigkeit des Karpfens. Der Karpfen gehört zu den weltweit bedeutendsten Arten der Süsswasserfischzucht. Er weist eine lange Geschichte der Domestikation auf, wobei zahlreiche Linien und Zuchtformen von seiner Wildform abgeleitet wurden.
In diese Thesis revidiert die Autorin das ursprüngliche Model von Kirpichnikov welches auf vier Haupttypen, die auf der vererbten Genetik des einfach schuppigen Karpfens und 2 Genen (s und N), die die Schuppigkeit in der Abdeckung der Cyprinidis reguliert, basiert. Die Forschungsgruppe der Autorin generierte eine lange Nummer von Kreuzung zwischen den verschiedenen Phänotypen des Karpfens unter Einbeziehung des Verlusts von Schuppenmutationen in Europa und Asien und hat so ein neues Modell für die objektive Klassifizierung des Phänotyps F1 und F2 ausgearbeitet. In dieser Arbeit beschreibt die Autorin das Verhältnis von Schuppenmuster- Phänotypen von Nachwuchsgruppen. Die Experimente zeigten, dass Abarten des sogenannten gesplitterten Phänotyps mit einer grossen Anzahl nicht überlappender Schuppen öfters in den Nachkommen auftreten. Daher sind die Schuppen in drei Subspezies geteilt:
Irregular, Inkomplet und Klassisch gespiegelt. Weiters wurden auch die Überlebensraten der Nachkommenschaft von potentieller Vererbt zweier N Allel analysiert und entfernte Unterschiede zwischen Europäischer und Asiatischen Kreuzungen gefunden. Dabei wurden Hinweise auf eine Präsenz eines starken N Allel mit einer homozygoten Letalität im Asiatischen Raum, und eines schwächeren-nicht tödlicheren im europäischen Raum gefunden. Die Kreuzungsversuche wurden parallel in Ungarn und Singapur unternommen. Die aktuelle Studie beschreibt die ersten Schritte unserer fortlaufenden Untersuchung zur Identifikation des zweiten Genes genannt „N“ (das „s“ Gen wurde gefunden zum Paralog des Fibroblast Wachstum Rezeptorfaktor 1, fgfr1a1.). Das bedeutendste Resultat ist, dass die Untersuchungsgruppe in Singapur das bis dahin fehlende Glied des Fgfr1 Rezeptorfamilie, fgfr1b isoliert und strukturell charakterisiert hat, und gezeigt hat, das die Sequenz nicht in den ungeschuppten in Unterschied zur gespiegelten mutiert ist. Die Untersuchungsgruppe schlägt ein neues Modell vor, welches die abweichenden
11
Phänotypen erklären würde, und legt dar, dass die neuen geschuppten Unterarten infolge einer zusätzlichen erhöhten Stufe von Fgf Signalen entstanden sind in Vergleich zu den gespiegelten und speziell den ungeschuppten.
12
1. Bevezetés és célkitűzések
A pontyfélék (Cyprinidae) családja a legnagyobb édesvízi halcsalád. Számos étkezési, akvarisztikai, vagy kutatási szempontból fontos fajuk ismert. Különösen gyakran találkozhatunk az ebbe a családba tartozó ponttyal (Cyprinus carpio), annak díszváltozatával a koi ponttyal, az aranyhallal (Carassius auratus), és a zebradánióval (Danio rerio) (Nelson 2006) (1.ábra).
A ponty földünkön legszélesebb körben elterjedt, étkezési szempontból jelentős édesvízi halfaj (Nelson 1994), a zebradániót, egy közkedvelt akváriumi halat pedig előszeretettel használnak fejlődésbiológiai, genetikai, tumorbiológiai, viselkedésbiológiai, ökotoxikológiai kutatásokhoz (Csenki 2011).
1. ábra: A ponty és a zebradánió (Fotó: Merth János, Szücs Réka)
Míg a zebradánió a genetika és a molekuláris biológia egyik fő kísérleti állatává vált (Hill és mtsai., 2005), a ponty genetikájáról- lévén tetraploid faj- jóval kevesebbet tudunk. A legtöbb hal testét sűrű, részben átfedő pikkelysorok borítják. Ugyan, ezen pikkelyzet képződéséhez és növekedéséhez szükséges molekuláris folyamatokat már többször vizsgálták, a pikkelyzet mintázatát szabályzó genetikai mechanizmusokról meglehetősen keveset tudunk. Kutatásainkat is ennek jegyében és ennek a gazdaságilag jelentős halfajnak jobb megismerése érdekében, a pikkelyezettség és az ezzel kapcsolatos mennyiségi paraméterek öröklődésének vizsgálata céljából végeztük.
Jóllehet Kirpichnikov és munkatársai már az 1930 – as években folytattak tavi kísérleteket a ponty pikkelymintázat öröklődésének megértésére, és megfigyeléseik alapján a pikkelyzet alakulására egy két gén - két allélos (N n; S s) rendszert írtak le (Kirpichnikov 1936), a gének csak napjainkban válnak ismertté. Kirpichnikov modelljét
13
a tankönyvek átvették és a halgenetikában ez lett a kétgénes öröklődés iskolapéldája (Purdom 1993., Tave 1993). Bár néhány keresztezést megismételtek és a kapott eredmények hasonlóak lettek az eredetiekhez (lásd pl.: Czucka 2002; Komen 1990b;
Nicolescu 2004), ismereteink szerint ez idáig senki nem vizsgálta újra nagyobb számú, módszeresen kidolgozott keresztezésekkel ezt a kérdéskört.
Kutatócsoportunkat két felismerés ösztönözte a modell felülvizsgálatára:
1., Kirpichnikovék rendszerében azok az egyedek, amelyekben az N allél homozigóta állapotban van, elpusztulnak. A bőrpontyok (ssNn) utódaiban pedig fenotípusos hasadásnak kell bekövetkeznie: 25% tükrös; 25% letális; 50% bőr fenotípus. Ezzel szemben egy magyarországi halfarmon az elvégzett bőr x bőr fenotípusú pontyok (2.ábra) keresztezésénél sem a Kirpichnikov modellje (Kirpichnikov 1981) szerint várt 25% letalitást, sem a „scattered”, tükrös fenotípust (Czucka 2002) nem tapasztalták.
2. ábra: A hajdúböszörményi bőrponty (Fotó: Szücs Réka)
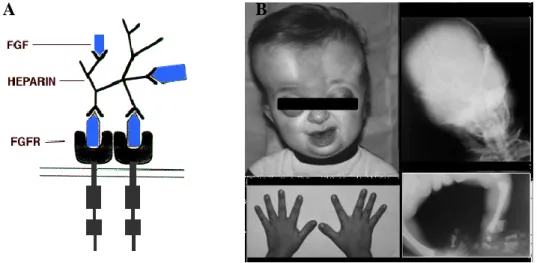
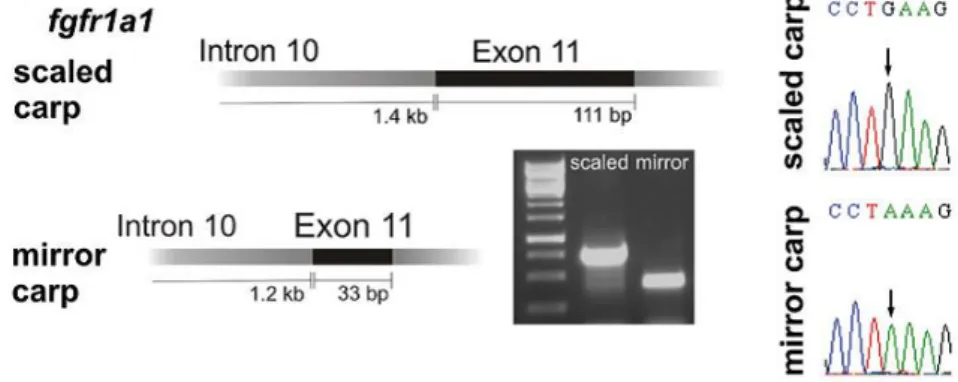
2., Ezt követte a tükrös zebradánió felfedezése, és az ezen fenotípus kialakulásáért felelős,- Kirpichnikov és munkatársai (Kirpichnikov 1981; 1999; Kirpichnikov és Balkashina 1935; 1936) által megjósolt „s” gén, a fibroblaszt növekedési faktor egyes receptor gén (fgfr1) mutációjának felfedezése.
Ennek a génnek zebradánióban két paralógja ismert: fgfr1a, fgfr1b; Pontyban eddig kettő: fgfr1a1, fgfr1a2 (Rohner és mtsai., 2009), de lévén tetraploid faj még további két paralóg jelenlétére számíthatunk.
Kirpichnikov munkája felbecsülhetetlen, mára azonban világossá vált, hogy az akkoriban általa vázolt modellnél a valóság bonyolultabb. Az „s” gén azonosítására több mint hetven évet kellett várni. Ekkor egy német-magyar-szingapúri együttműködés révén derült ki, hogy a tükrös fenotípus kialakulását a fibroblaszt növekedési faktor egyes receptorának (Fgfr1) mutációja okozza. Rohner és munkatársai megállapították,
14
hogy a pontynál is a zebradánióban talált tükrös változattal homológ fibroblaszt növekedési faktor receptor génjének mutáns változata okozza a részleges pikkelyhiányt.
2009-ben felfedezésre került továbbá az is, hogy ezt a folyamatot két, egymással nem kapcsolt gén szabályozza (Rohner és mtsai., 2009).
A dolgozatban a fenotípusos arányok pontosabb megértéséhez az elmúlt 5 év során elvégzett keresztezéseket, a pikkelyezettség kialakulásával kapcsolatba hozható elváltozásokat, a mutációja esetén a pontyfélék teljes pikkelyvesztését okozó „N” gén megtalálására tett lépéseket mutatjuk be magyarországi és szingapúri keresztezéseken keresztül. Munkánk során különböző pikkelymintázatú pontyokkal végeztünk keresztezéseket, fenotípusos megfigyeléseket, DNS mintát gyűjtöttünk, garatfogat vizsgáltunk, ezzel is segítve a genetikai háttér tisztázását és a nemzetközi kutatócsoport munkáját. Kirpichnikov modelljének nyomán az általa javasolt 4 fenotípusos változatot a pontosabb besorolások végett 6-ra bővítettük. A párhuzamos keresztezések folytán különbségeket találtunk az ázsiai és az európai keresztezéseknél az utódok túlélési rátájában, életképességükben is.
Mindezek ismeretében a főbb célkitűzéseink az alábbiak voltak:
nemzetközi csoportokkal együttműködve felülvizsgáljuk és pontosítsuk a ponty pikkelymintázatának genetikájáról alkotott képet, és ha lehet, olyan új modellt alkossunk, amelyik a régihez képes jobban leírja a sokféle fenotípus megjelenését, és a korábbi modellben várt letalitások elmaradását.
Nemzetközi csoportokkal együttműködve kísérletet tegyünk a pikkelymintázatért felelős másik gén (az „N” gén) felkutatására.
Összefüggést keressünk a pikkelymintázat és más jellemzők, mint pl. a garatfogak, a kopoltyúfésű tüskéi vagy az úszók alakulása között.
Megvizsgáljuk a megszokottól eltérő (oldalvonalsoros, bőrponty) hazai pontyok tenyésztésének jogosultságát a növekedési, vagy megmaradási mutatók tekintetében.
15
2. Irodalmi áttekintés
Napjainkban a ponty (Cyprinus carpio L.) a földünkön legszélesebb körben elterjedt édesvízi halfaj (Pintér 2002).
Feltehetőleg a legismertebb csontos hal a világon. Ez háziasítása hosszú történelmének, továbbá a kontinensek közti terjedésének köszönhető, mely eredményeképp sok vad és tenyésztett (alfaj, fajta, törzs, vagy tájfajta) változat alakult ki (Komen 1990a).
A legtöbb szakember egyetért abban, hogy a ponty Kelet-Ázsiából származik, és a miocén közepén vált el a Cyprininae alcsalád többi tagjától (Zardoya és Doadrio 1999).
Legvalószínűbb, hogy Kínából származik és később került Európába (Froufe és mtsai., 2002), Észak- Amerikába (McCrimmon 1968) és más kontinensekre.
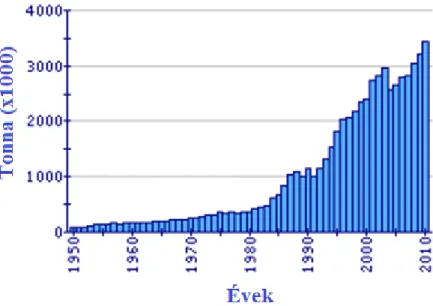
Egyike az étkezési szempontból legfontosabb édesvízi halaknak (3. ábra). A pontyfélék a világ haltermelésének több mint 30%-át adják és a ponty a harmadik legfontosabb tenyészett halfaj, 3,4 millió tonna éves világtermeléssel közel 14%-a a világ összes édesvízi haltermelésének (Xu és mtsai, 2011).
3. ábra A világ pontytermelése (FAO Fishery Statistic nyomán) http://www.fao.org/fishery/species/2957/en
A pontyfélék szinte bárhol fellelhetőek a természetben az északi félteke mérsékelt égövi zónájában és a trópusi régiókban egyaránt. Többségük édesvízi, csak kis részük él a brakkvízi folyótorkolatok környékén (Howes 1991).
16
A Japánban kitenyészett és a világon, dekoratív színeiknek köszönhetően oly népszerű aranyat érő koi-pontyok (nishikigoi -"brocaded carp") a háziasított ponty díszváltozatai.
Számos tanulmány jelent meg a ponty tenyésztéséről és genetikájáról is, például Horváth és Orbán (1995) tanulmánya a ponty genom és génmanipulációjáról; Balon (1995) írása a ponty eredetéről és háziasításáról; Nagy és mtsai (1981, 1984), Komen (1990a, 1990b) Bercsényi és mtsai. (1998), valamint Gomelsky (2003) áttekintése a ginogenezisről és az androgenezisről, az ivar szabályzásáról; Vandeputte (2003) összefoglalója a pontyok nemesítéséről. Sokak részletesen mutatják be a rokonságba tartozó más pontyféléket is (pl. Hulata 1995, 2001, Penman 2005).
2.1. A ponty rendszertana
Elnevezései (http://fishbase.org):
latin: Cyprinus carpio carpio (Linnaeus, 1758) angol: common carp, carp, wild common carp
német: r Karpfen, Karpe, Donaukarpfe, Schuppenkarpfen, Lederkarpfen, orosz: Сазан(=карп), Karp
cseh: Kapr obecný, Kapr mahujský olasz: Carpa
portugál: Carpa, Sarmão
francia: Carpe, Carpeau, Carpe commune, Carpe cuir, Carpe miroir, Feuille, Karpenn magyar: ponty, potyka, pathal, pozsár, ivadék: babajkó (Pintér K., 2002) formák és változatok: tőpotyka, csupaszponty, királyponty, tükrös ponty, stb. (Herman, 1887).
A pontyfélék sugarasúszójú csontos halak:
törzs: Gerinchúrosok - Chordata altörzs: Gerincesek – Vertebrata
osztály: Sugarasúszójú halak – Actinopterygii (ray-finned fishes) divízió: Csontos halak – Teleostei
rend: Ponty alakúak - Cypriniformes család: Pontyfélék – Cyprinidae nem: Cyprinus Linné, 1758
faj: ponty - Cyprinus carpio Linné, 1758 (http://www.fishwise.co.za)
17
A Cyprinidae család – Cyprinus genus rendszertani helye
A ponty alakúak rendjébe 6 (+2) család tartozik, köztük a pontyfélék családja (Cyprinidae).
A FishBase adatai alapján a pontyfélék családjába 210 nem és több mint 2000 faj tartozik (Froese és Pauly, 2012). ( http://fishbase.org)
o Rend Cypriniformes
Család Cyprinidae Subfamily Cyprininae
Genus Cyprinus (24 faj)
Cyprinus carpio carpio™ C. Linnaeus, 1758 - common carp
Cyprinus carpio haematopterus Martens, 1876
A Cyprinus nembe a jelenleg elfogadott rendszertan szerint 24 faj tartozik:
1. Cyprinus acutidorsalis 2. Cyprinus barbatus
3. Cyprinus carpio carpio Linnaeus, 1758 (Kottelat és Freyhof, 2007) 4. Cyprinus carpio haematopterus (Komen 1990. )
5. Cyprinus centralus 6. Cyprinus chilia 7. Cyprinus dai 8. Cyprinus daliensis 9. Cyprinus exophthalmus 10. Cyprinus fuxianensis 11. Cyprinus hyperdorsalis 12. Cyprinus ilishaestomus 13. Cyprinus intha
14. Cyprinus longipectoralis 15. Cyprinus longzhouensis 16. Cyprinus megalophthalmus 17. Cyprinus micristius
18. Cyprinus multitaeniata 19. Cyprinus pellegrini 20. Cyprinus qionghaiensis 21. Cyprinus quidatensis 22. Cyprinus rubrofuscus 23. Cyprinus yilongensis 24. Cyprinus yunnanensis
18
2.2. A ponty morfológiája /felépítése
Testalakja örökletes alapon és környezeti tényezők hatására rendkívüli változékonyságot mutat, ami a vadpontyokra és háziasított pontyokra egyaránt jellemző.
Testük szélességének, magasságának, hosszának jellemzésére, mérésére, a nemesítői munkák során is használatos mutatókat a 4. ábra szemlélteti.
A pontyok testének tipikus jellemvonásai: a fog nélküli állkapocs, a garatban lévő garatfog, mely ideális határozó bélyeg, Weber-csontok jelenléte a gerincoszlop kezdeténél (Orban és Wu , 2008). A ponty hátúszójának első úszósugara a kemény csontú, kissé ívesen hajló, belső felületén fogazott bognártüske.
A vizeinkben élő vadpontynak két formáját különböztetjük meg főként a profilindex és a fejindex eltérő értéke alapján: 1. nyurga ponty (Cyprinus carpio carpio m. hungaricus), 2. tőponty (Cyprinus carpio carpio m. acuminatus). A nyurga ponty teste nyújtott, hengeres, míg a tőponty egy magasabb hátú, rövidebb testű, oldalról viszonylag lapítottabb forma. A vadpontyok feje viszonylag nagy, szemük a testhez viszonyítva kicsi (Pintér 2002).
4. ábra: A ponty jellemzésére használt mutatók és méretek. (Pintér 2002 .nyomán). fe: fejhossz, fn:
faroknyélhossz, fsz: fejszélesség, th: testhosszúság, tm: testmagasság, tsz: testszélesség
Rendszertani bélyegek (Czuczka 2002 - Berinkei (1966) szerint):
Úszósugárképlet:
D (hátúszó) D: III (IV)/(15)16-21(22) A (farokalatti úszó) III 5 (6)
P (mellúszó) I 16-18 C (farokúszó) II 8-9
Csigolyaszám: 34-39 (36-37 a fishbase.org szerint)
mutatók
vadponty tógazdasági pontyfajta nyurga
ponty tőponty egyenes hátívű
hirtelen hátívű profilindex 3,5-4,5 2,8-3,5 2,6-3,0 2,0-2,6 keresztmetszetindex 1,4-1,8 1,5-2,0 1,5-1,8 1,8-2,2
fejindex 3,8-4,3 3,6-4,1 - -
faroknyélindex 1,4-2,0 1,6-2,2 - -
19
Pikkelyképlet: 33 ---- 40 vagy 32---41 (Harka Á. és Sallai Z. 2007).
Kopoltyútüskék: 21-29
Garatfogképlet: 1.1.3-3.1.1 = a fogak 3 sorban állnak, a belső sorban 3-3, a többiben 1-1 fog van vagy 1.2.3-3.2.1 (Harka és Sallai, 2007).
Haploid kromoszóma szám: 50 (n) (Berinkei 1966).
A száj végállású - csúcsba nyíló, harmonikaszerűen kitolható. A felső ajkán két rövidebb, a száj két szegleténél 1-1 hosszabb bajuszszál található. Orra hosszú, tompán lekerekített, homloka széles és erősen domború. A farokúszó nagy és erősen kivágott.
Cycloid pikkelyei nagyok, vastagok. Testalkatához hasonlóan színe is rendkívül változatos, a kultúrváltozatok között gyakran akadnak albínó, fekete és aranysárga példányok, koi pontyoknál fehér, kék, piros, citromsárga, narancssárga színváltozatok is.
A „vad” színezetű egyed színezete Herman Ottó szerint:
„Szín szerint a hát kékeszöld, a feketésbe húzó, az oldalak rezessárgák, néha sötétzöldesek; a hasfél fehéres; a pénzek feketés szegélyűek, mi bizonyos koczkás jellemet teremt; a hátsörényúszó szürkéssárgás, az úszószárnyak és a kormányúszók violás lehelletűek, az alsósörényúszó vörösesbe játszik, a szemcsillag aranyos. Ezek a színek azonban nem állandók s víz, táplálkozás, világosság hatása alatt sokszorosan változnak; a folyóban élő ponty sokkal világosabb, az álló, különösen mocsaras vizekben élő sötét színű, különösen oldalai sötéten aranyosak.” (Herman 1887)
Életterük az álló és lassú áramlású folyóvizek, holtágak. Kitűnően alkalmazkodtak a víz viszonylag magasabb sótartalmához is, de brakkvizekben nem szaporodnak.
Hazánk természetes vizeiben az ikrások 3-4 év, a tejesek 2-3 év után válnak ivaréretté.
Az ívás 18-22 °C-os vízben történik. Szaporodásuk 8-25 naponként megismétlődhet, ha az ívási környezet ismét kialakul. Ezért mérsékelt égövön április végétől augusztusig lehetséges ívásuk. Az ártéri területek füves rétjeit, a finom szálú hínárral dúsan benőtt, sekély vizű tórészeket választják ívóhelyként. Az ikrások testtömeg-kilogrammonként
5-6
5-6
5-6
5-6
5-6
5-6
5-6
5-6
20
150-200 ezer, 1-1,5 mm átmérőjű, szürkészöld színű ikrát raknak le. A lárvák 3-4 nap után kelnek ki, 6-7 mm hosszúak, ragasztómirigyük segítségével 4-5 napig függeszkednek a növényeken. Első táplálékként kerekesférgeket, planktonrákokat fogyasztanak, majd növekedésük során egyre inkább bentikus szervezetek képezik táplálékukat. A kifejlett egyedek mindenevők, ragadozó magatartásforma csak ritkán, főként fehérjehiányban szenvedő anyapontyoknál figyelhető meg (Györe 1995).
2.3. A ponty eredete /elterjedése
Napjainkban a ponty a földünkön legszélesebb körben elterjedt édesvízi halfaj.
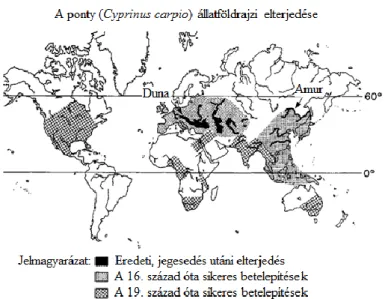
Európában és Ázsiában részben természetes terjedésének, részben az ember tudatos telepítő tevékenységének köszönheti (5. ábra), hogy a legészakibb területek kivételével gyakorlatilag mindenütt megtalálható az élőhelyi igényeinek megfelelő vizekben (Pintér 2002).
A legtöbb szakember egyetért abban, hogy a ponty Kelet-Ázsiából származik, és a miocén közepén vált el a Cyprininae alcsalád más tagjaitól (Zardoya és Doadrio, 1999).
Ennek ellenére számos különböző elmélet létezik a mai pontyfajták filogeográfiájára:
Berg (1964) szerint a ponty eredetileg Eurázsia-szerte létezett, majd később keleti és nyugati populációkra oszlott a pleisztocén korban.
Balon (1995) véleménye különböző, szerinte a ponty a Fekete, Kaszpi és Aral tengerekkel határos vízrendszerből származott és innen terjedt keletre és nyugatra. A fosszilis leletek hiányában kizárta előfordulását a jégkorszak előtti Európában. Két alfajt említ: C.c carpio és C.c. haematopterus – amelyek jellegzetesen különböznek morfológiailag, mint ahogy azt korábban Svetovidov (1933) leírta. További alfajok létezését megkérdőjelezték (Orban és Wu, 2008).
Egyes szerzők, így Kirpichnikov (1967) szerint már a harmadkorban elterjedt volt az egész eurázsiai kontinens vizeiben, de a jégkorszak hatására az eredetileg egységes előfordulási terület keleti és nyugati részekre szakadhatott (Pintér 2002), amely aztán az alfajokat alkothatta (Komen 1990a).
21
5. ábra: A ponty eredeti, jegesedés utáni elterjedése/ és sikeres terjesztése (Komen 1990a).
Morfológiai különbségek alapján (Kirpichnikov 1967) négy (Pintér 2002 szerint öt) különböző alfajt (javasolt) írt le (6. ábra):
- Európai- transzkaukázusi ponty (Cyprinus carpio carpio) - Közép- ázsiai ponty (C.c. aralensis)
- Kelet- ázsiai ponty (C.c. haematopterus) - Dél- ázsiai ponty (C.c. viridiviolaceus)
(- Az indonéziai ponty- tudományos név nélkül (Pintér, 2002).)
6. ábra: Kohlman és mtsai, 2005: A ponty alfajainak megoszlása Kirpichnikov szerint (1967), (a) Európai-transzkaukázusi ponty, C. carpio carpio; (b) Közép- ázsiai ponty, C. c. aralensis; (c) Kelet- ázsiai ponty, C. c. haematopterus; (d) Délkelt- ázsiai ponty, C. c. viridiviolaceus.
Később Kirpichnikov (1999) változtatva véleményét, Balonhoz hasonlóan 2 alfajt említ: C. carpio carpio (Európa; Kaukázus; Közép-Ázsia) és C.c. haematopterus (Kelet- Ázsia). Végül a genetikai vizsgálatok nyomán a klasszikus taxonómiával együtt
22
a jelenleg létező ponty-formákat három kategóriába sorolták: (1) Európai (Cyprinus carpio carpio), (2) Távol- Keleti (C. carpio haematopterus) és (3) Délkelet- Ázsiai (C.
carpio viridiviolaceus) kategóriákba (7. ábra) (Kirpichnikov, 1999; Baruš és mtsai., 2002; Kohlman és mtsai., 2005).
Wu (1977) három alfajt ismert el különböző földrajzi elterjedésekkel: - C.c.
carpio; - C.c. haematopterus; - C.c. rubrofuscus, amelyet az irodalomban C.c.
viridiviolaceus-ként és C.c. nigroauratus-ként is említenek. A FishBase adatbázisa ezt a taxont morfológiai adatok alapján külön fajként (C. rubrofuscus), de molekuláris adatok alapján Orban és Wu (2008) alfajként említik.
Wu szerint a C.c. carpio-nak volt a legszélesebb elterjedési köre a Dunától a Volgáig, néhány populációval kiegészítve Kínának a Xinjiang Ujgur autonóm területéről. A C.c.
haematopterus a kínai Nanling Hegységtől északra eső területről származik, míg a C.c.
rubrofuscus a Nanling Hegységtől délre lévő területről.
Ma a pontyokat molekuláris genetikai alapon legalább két alfajra osztják: az európai (C. c. carpio)-ra és az ázsiai (C. c. haematopterus)- ra. Ez mikroszatellitekkel és mitokondriális genetikai adatokkal jól alátámasztott (Vandeputte 2003; Kohlmann és mtsai., 2003, 2005a; Zhou és mtsai., 2003a, 2004; Brody és mtsai., 1979; Paaver 1983;
Kohlmann és Kersten 1999; Gross és mtsai., 2002).
Ezen kívül lehetséges egy harmadik alfaj (C. c. rubrofuscus vagy C. c.
viridiviolaceus) létezése, de ez genetikai adatokkal nem megerősített (Kohlmann és mtsai., 2005b).
7. ábra: Az eurázsiai vad ponty populációk elterjedése Kirpichnikov 1999 nyomán (Flajšhans és Hulata, 2006)
1. Európai alfaj Cyprinus c. carpio
Távol- keleti alfaj Cyprinus c. haematopterus Délkelet- ázsiai alfaj Cyprinus c. viridivlaceus
23
Korábban, a közép-ázsiai alfajt (C. c. aralensis) Kirpichnikov javasolta (Kirpichnikov 1967 ). Azonban a legújabb kutatások (Kohlmann és mtsai., 2003, 2005a;
Memis és Kohlmann, 2006) azt mutatják, hogy az európai és közép-ázsiai pontyformák valóban nagyon szoros, közeli rokonságban állnak, genetikailag jórészt egyeznek.
A szerzők következésképpen az európai és közép-ázsiai pontyot is a carpio alfajba sorolják. A mtDNS szekvenciák elemzése alapján, Froufe és munkatársai (2002) arra a következtetésre jutottak, hogy az európai pontyok valószínűleg Ázsiából kerültek be.
A pontyváltozatok különböző hatások, mint pl. a földrajzi izoláció, mutációkhoz való adaptálódás, természetes,- és emberi szelekció kombinációjaként alakultak ki (Hulata 1995). A közép- és kelet- európai régióban több pontyváltozat található, ám annak ellenére, hogy kereskedelmi szempontból igen fontosak, eredetük és rokonsági viszonyaik nem tisztázottak, az állományok genetikai adatai hiányosak (Bártfai és mtsai., 2003).
Az egyik, étkezési célból legrégebben háziasított faj a ponty. Egyike a kevés halnak, amely igazán háziasított állatnak mondható (Steffens 1980).
Tenyésztésének történelme Kínában a Kr.e 5. évszázadnál korábbra, ősi kínai dokumentumok szerint a Kr.e –i 12. sz-ra nyúlik vissza (Balon 1995; 2004; Orban és Wu., 2008), bár háziasításuk csak jóval később kezdődött. A tavi haltenyészés Kínában a Shang dinasztia ideje alatt kezdődött Kr.e. 20. században. Fan Li könyve (Kr.e. 460- ban) az első írásos emlék a haltenyésztésről. A Han Dinasztia alatt (Kr. e. 206/202–Kr.
u. 220) a pontytenyésztés virágzott. A pontyot elsődleges fajként nemcsak mesterséges kis tavakban, de természetes vizekben is tenyésztették.
Az uralkodó nézet szerint (pl. Günther 1868; Chiba és mtsai., 1966; Vooren, 1972) az európai háziasított ponty elődje az ázsiai ponty volt, amelyet az ókori görögök és rómaiak idejében szállítottak Ázsiából Európába (Zhou és mtsai., 2003b). Más írások szerint: a ponty európai rasszai a dunai vad pontyoktól származnak. A legkorábbi háziasítási kísérletek az ókori Római Birodalom és a kereszténység európai elterjedésének idején történtek, és ettől a kortól kezdve jelentek meg a háziasított fajták más kontinenseken is (Flajšhans és Hulata, 2006; Horvath és mtsai., 1984; Balon, 2006).
A legrégebbi írásos dokumentumok Európából a 6. századból valók, ahogy a Day idézetében (1880) Cassiodorus említi a dunai különleges ízű, értékes pontyot, a
24
hercegek eledelét. A 13. századtól a németek és más európai haltenyésztők főleg a kolostorok környékén tavaikban a Dunából befogott vad pontyokat kezdtek tartani. A 15. századtól a halételek nagy népszerűségnek örvendtek a kelet- európai népek (cseh, lengyel, magyar) körében főként karácsony idején. A 16. század végére a háziasított dunai ponty egész Európában elterjedt, a pontytenyésztés központja Bohemia (mai Csehország) hegyvidéke lett.
Európában először kimondottan pontytenyésztésről a XII. században élt Albertus Magnus írt. Az 1500-as években a dunai ponty elérte Hollandiát, Dániát, sőt Angliába is eljutott. Poroszországból pedig hajón Svédországba is szállítottak pontyot (Bercsényi 1997). 1831-ben Franciaországból jutott Amerikába (Lachner és mtsai., 1970) és később Ázsiából és Európából pedig Kaliforniába 1872-ben és 1877-ben (Moyle 1976) Kanadába (Sarig 1966).
A ponty csak néhány Európai országban őshonos, elsősorban a Duna vízgyűjtő területén lévőkben. (8. ábra) (Magyarországon, Ausztriában, Bulgáriában, Csehországban, Szerbia, Szlovákia, stb.: fishbase.org)
8. ábra: A ponty helyzete Európában; N=őshonos; I=betelepített. ( Flajšhans és Hulata, 2006)
A ponty őshonos halaink közé tartozik, amely Belső – Ázsiából természetes úton jutott el Európába és Kelet – Ázsiába, majd később sokfelé telepítették (Harka 1997).
Ugyanakkor a jelenlegi vadon élő dunai vadponty populációk előfordulása megkérdőjelezhető, valószínűleg a vízgyűjtő terület kis részére korlátozódnak, és antropogén hatások, valamint a halgazdaságok és a tenyésztett populációk szökevényei veszélyeztetik őket. Mostanában jelentettek Törökországból néhány vad populációt, bár ezek nem őshonosak, fontos genetikai forrást jelenthetnek (Flajšhans és Hulata, 2006).
25
Olyan vad törzseket, amelyek kapcsolatban állnak az európai populációkkal, közép- ázsiai országokban pl., Üzbegisztánban is találtak (Flajšhans és Hulata, 2006).
Mára a pontyot több helyen invazív fajként tartják számon: pl. Ausztráliában (Gwilym 2009) és az USA-ban pl. Minnesota államban (regulated invasive species:
http://www.dnr.state.mn.us/invasives/laws.html).
2.4. Nemesített és vad ponty változatok
„ A TŐ PONTY. Cyprinus carpio, LINNÉ.
Fajtái:
A király vagy tükrös ponty. Ez magán viseli a tőponty minden főjegyét; de héjja nem egész, hanem csupasz helyek vannak rajta, ahol pedig megvan, ott egyenetlen, sokszor igen nagy pénzekből alakul; az oldalvonalat képzők rendesen sorakozottak, azon alúl és felül mindenféle fejlődésűek és rendetlenül állanak. Ez a BLOCH-tól leírt "Cyprinus rex Cyprinorum"; AGASSIZ "Cyprinus Carpio macrolepidotus" néven írta le.
A csupasz ponty. Minden bizonynyal a tükröstől ered akként, hogy vénségére az amúgy is rosszúl legyökerezett pénzek kihullanak s a hal egészen meztelenné válik; ez azután a
"Cyprinus carpio nudus, vel alepidotus AG."
Ezek a fajták tavakban keletkeznek és átöröklés útján tovább szaporodnak is.”
(Herman, 1887).
A genetikailag kódolt tulajdonságokat, két fő csoportba sorolhatjuk: minőségi (kvalitatív) és mennyiségi (kvantitatív) csoportba. A minőségi tulajdonságok (pl.
pikkelyezettség, a testszín, forma) főbb jellemzői, hogy fenotípusos megjelenésüket egy vagy kevés számú, de erős hatású úgynevezett oligén, major vagy főgén határozza meg és hatásukat a környezet változása egyáltalán nem vagy csak kis mértékben befolyásolja. A mennyiségi tulajdonságok (pl. testsúly, takarmányhasznosítás, ellenálló képesség) fenotípusos megjelenését az előzővel ellentétben nem egy, hanem több gén úgynevezett poligénes rendszer határozza meg, és hatásukat a környezet változása nagymértékben befolyásolja. A minőségi és mennyiségi tulajdonságok egymással kölcsönhatásban alakítják ki az adott élőlény tulajdonságait, azaz a ponty pikkelyezettsége, mint minőségi tulajdonság a mennyiségi tulajdonságokkal kapcsolatban van és így együttesen jelentős mértékben befolyásolják a haltenyésztés eredményességét (Czuczka 2002).
26
2.4.1. Tájfajták és hibridek kialakulása Magyarországon
A ponty egyike a legrégebben tenyésztett és háziasított halfajoknak a világon.
Amikor a tenyésztés kezdődött az ősi populációk vad formái domináltak. Egyesek a mai napig léteznek (a vad pontyok megnyúlt, pikkelyes formáiként) gazdagítva a helyi halfaunát, azonban a legtöbb napjainkban tenyésztett ponty különbözik őseitől.
A ponty háziasítása különböző változatok kialakulásához, egyúttal eltérő pikkelyezettségi mintákhoz vezetett. A vad fenotípus teljesen pikkelyes, torpedó alakú hal volt, de a mesterséges szelekcióval számos pikkelyváltozatot alakítottak ki az évszázadok során. Ezeket a változatokat igen megkedvelték, mivel kevesebb pikkelyük miatt főzésnél könnyebb volt tisztításuk (Michaels 1998).
Az eltérő környezeti feltételek, a tenyésztők egyéni módszerei, a tenyészállomány kis mérete és a szigorúan zárt tenyésztési rendszerek miatt a tenyészett pontyoknak (fajon belül) helyi populációi alakultak ki.
A XIX. század közepe után különböző genotípusokat fejlesztettek ki és tájfajtáknak nevezték el őket ("landraces", Bakos 1979). A XIX. század végére tehető első nemesponty honosítások idején több európai fajta került hazánkba, melyek német, cseh és dél-szláv tógazdaságokból származtak, majd a haltenyésztők gondos szelekciós munkája, és a helyi környezeti adottságok együttes hatása következtében genetikai tulajdonságaikban konszolidálódva az 1950-es években már, mint önálló tájfajták gazdagították a magyar haltenyésztést.
A halgazdaságok egy jelentős része önálló fajtával rendelkezett, így beszélhetünk a tatai pikkelyes, a hortobágyi tükrös és pikkelyes, a szegedi, a biharugrai, a varászlói, majd később a dinnyési és szarvasi tükrös pontyfajtákról. A hazai halgazdaságok pontyfajtáinak összegyűjtésével ponty fajtagyűjteményt, élő génbankot alakított ki Dr.
Bakos János Szarvason a 60-as években. A génbank az 1970-es évek közepére már 15 hazai és 14 külföldi pontyfajtának adott helyet.
A világban számos helyen működik élő génbank (pl.: Vodnany –Csehország;
Zator, Golys – Lengyelország; Nucet – Románia; Poljana – Szerbia; Zagreb – Horvátország; FCRI (Thoothukudi, Tamil Nadu) - India, de a magyarországi Szarvason működő élő génbank is világhírű.
27
2.4.2. Tájfajták és tenyésztett változatok pikkelyezettsége Magyarországi tájfajták és tenyészett változatok:
Bikali tükrös
Hortobágyi nyurga ponty Hortobágyi pikkelyes ponty Hortobágyi tükrös
Dunai vadponty Felsősomogyi tükrös Dinnyési tükrös Nagyatádi tükrös Palkonyai tükrös Gödi tükrös
Sumonyi tükrös Szarvasi 15 tükrös
Szarvasi 22 tükrös Szarvasi P 33 pikkelyes Szarvasi P31 hibrid pikkelyes Szarvasi P34 hibrid pikkelyes Szarvasi 215 hibrid tükrös Szarvasi piros
Szarvasi tükrös Szegedi tükrös Tatai pikkelyes
Tiszai nyurga (www.fao.org)
A hibridek nemesítése során leginkább az alábbi 5 fő termelési paraméter javítására irányuló tenyésztési eljárást alkalmazzák. A legfontosabb, a termelés gazdaságosságát meghatározó tulajdonságok a következők: növekedőképesség;
takarmányértékesítés; megmaradási százalék; vágóérték; a hús zsírtartalma.
Magyarországon a tükrös és pikkelyes pontyok a közkedveltek, ritkán találkozhatunk oldalvonalsoros és bőr változatokkal is. A 60- as években a bőrpontyot próbálták kiiktatni a termelésből, mondván, hogy genetikai defektusokat hordoz magában, és ezért nem felel meg a gazdaságos haltenyésztés követelményeinek.
Ugyanígy az oldalvonalsoros fajta sem közkedvelt. Magyarországon nincs minősített oldalvonalsoros tájfajta, de Lengyelországban, a Zatori Haltenyésztési Kutatóintézet génbankjában 1973 óta, génmegőrzési céllal generációk óta belterjesen tartják fenn ezt a helyi eredetű pontyot, amely a Z-102 zatori anya és a Z-105 zatori apa keresztezéséből származik. Megnyúlt testformájú oldalsoros fajta. Mivel eredeti gazdaságában sem tenyésztik, sőt az oldalvonal mentén pikkelyezett egyedeket később szelekcióval eltávolították az állományból, csak génmegőrzési céllal tartják fenn.
Teljesítményvizsgálaton a Szegedi tükrös fajtához képest csak minimális előnyöket mutatott, fenotipusos megjelenése azonban nem tette kedvelt fajtává, illetve keresztezésekor vagy tiszta tenyésztésekor az utódok fenotípusos megjelenése (az oldalsoros jelleget meghatározó genotípusból eredően) a négy lehetséges pikkelyezettségi forma között szórt.
További külföldi tájfajták és hibridek például: Cseh pikkelyes, Cseh tükrös, Amuri vadponty, Fresinet tükrös ponty, Német tükrös, Lengyel tükrös (Golys), Nasici
28
tükrös (Horvátország), Poljana pikkelyes (Szerbia), Poljana tükrös, Ropsha pikkelyes (Románia), Ukrán pikkelyes, Vietnámi pikkelyes.
2.5. A ponty evolúciós ugrása – a tetraploid állapot és az ebből adódó lehetőségek
2.5.1. Egy család- 3 különböző genom méret
A pontyfélék kromoszómakészletét citogenetikai vizsgálati módszerekkel már alaposan tanulmányozták (lásd pl. Buth és mtsai., 1991; Klinkhardt és mtsai., 1995;
Sola és Gornung, 2001). Számuk 42-től több mint 200-ig terjedhet (Buth és mtsai., 1991). Mindemellett minden pontyféle vagy 24-25 kromoszómát vagy ennek a dupláját hordozza haploid készletként. Az amur (Ctenopharyngodon idella), a pettyes busa (Hypophthalmichthys nobilis) és a fehér busa (Hypophthalmichthys molitrix) kromoszóma száma egységesen 2n=48, viszont kariotípusukban eltérés mutatkozik (Yu és mtsai. 1987).
A ponty kariotípusa 22 metacentrikus (m) + 34 szubmetacentrikus (sm) + 22 szubtelocentrikus (st) + 22 telocentrikus (t). A kromoszómakarok száma NF=156 (Rab és Collares-Pereira, 1995). A kromoszóma számban változatok lehetnek, pl. a koi pontyé lehet 101 és 102 is; Ojima és Takai 1981). Az aranyhal diploid kromoszóma száma megegyezik a pontyéval (2n=100) és a kariotípusuk is egyformának tűnik (Wu és Gui, 1999).
A legtöbb szerző szerint a pontynak 2n=100 kromoszómája van, de vannak ettől eltérő publikációk is melyek szerint 2n=98 (Al-Sabti 1987, 1986; Arkhipchuk 1999) vagy 2n=102 (Brzuska 1988) és 2n=104 (Post 1965; Hinegardner és Rosen, 1972; Wolf és mtsai.,1969;Arkhipchuk 1999; Ohno és mtsai., 1967; Sola és mtsai., 1993).
A zebradánió kariotípusa: 4m+16sm+ 30st (NF=70) (Sola és Gornung, 2001). A zebradánió haploid kromoszóma száma 25; majdnem, mint az amurnak és a busáknak, de a genom mérete jelentősen nagyobb, közel a ponty és az aranyhal újra duplázódott genommérete (1. táblázat).
29 Magyar
név Latin név*
Haploid kromoszóma
szám
Kromoszóma készlet
Genom méret
Haploid CV***
(pg) Pettyes
busa
Aristichthys nobilis
(Richardson, 1845) 24 2 kicsi 1,04
(0,01) Amur
Ctenopharyngodon idella
(Valenciennes, 1844) 24 2 kicsi
1,02 (0,03) Fehér busa
Hypophthalmichthys molitrix
(Valenciennes, 1844) 24 2 kicsi 1,01
(0,02) Zebradánió Danio rerio (Hamilton, 1822) 25 2 közepes 1,78
(0,25) Ponty
Cyprinus carpio (Linnaeus,
1758) 50 2 közepes 1,79
(0,10) Széles
kárász
Carassius carassius
(Linnaeus, 1758) 50 2 közepes
1,99 (0,21) Aranyhal
Carassius auratus auratus
(Linnaeus, 1758) 50 2 közepes 1,78
(0,14) Japán
ezüstkárász
Carassius auratus langsdorfii (Temminck &
Schlegel, 1846)
U** 3 nagy 2,55
Ezüstkárász
Carassius gibelio (Bloch,
1782) 50 3 nagy 2,92
(0,21)
*: www.fishbase.com U**:ismeretlen ***: http://www.genomesize.com/
1. táblázat: A pontyfélék kromoszóma száma és genom mérete (Orban és Wu, 2008 nyomán).
2.5.2. Genom duplikáció a csontos halakban
A valódi csontos halak alkotják a gerinces fajok körülbelül felét (legalább 26000 fajuk van), ebből következtethetünk nagy változatosságukra is (Lah 2012).
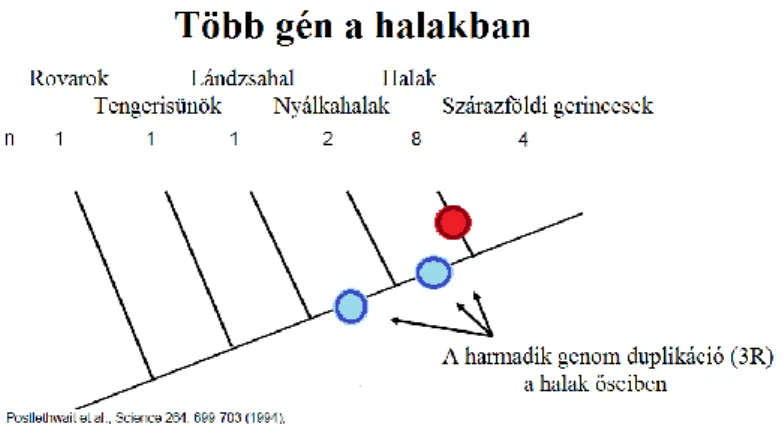
A gerincesek fejlődésének kezdetén kb. 450 millió éve két egész genomra kiterjedő duplikáció történt (Holland és mtsai., 1994; Sidow, 1996; Skrabanek és Wolfe 1998), majd kb. 300-350 millió évvel ezelőtt a csontos halak ősénél egy újabb, hal- specifikus harmadik genom duplikáció (3R duplikációként is ismert) történt (Postlethwait és mtsai., 1998; Yokoi és mtsai., 2007; Christoffels és mtsai., 2004). Ez az esemény, amely a csontos halakat egy extra paralóg készlettel ruházta fel, feltételezhetően, legalább részben felelős biológiai sokféleségükért, fajgazdagságukért (Amores és mtsai. 1998; Meyer és Van de Peer 2005; Taylor és mtsai. 2001). A mai halak genomjában ezeknek a paralógoknak kb. 30%-a maradt meg (Lah 2012).
30
9. ábra: Két egész genomra kiterjedő duplikáció a gerincesek ősében és a halspecifikus 3. genom duplikáció. (Postlethwait és mtsai., 1994; Wittbrodt és mtsai., 1998 nyomán)
A pontyoknak és az aranyhalaknak - köszönhetően egy tetraploidizációs eseménynek (Kb. 58 millió évvel ezelőtt (Zhang és mtsai., 1995)) kétszer annyi kromoszómájuk van, mint a többi pontyfélének. David és mtsai., (2003) szerint ez körülbelül 16 millió évvel ezelőtt történt, Larhammar és Risinger (1994) vizsgálatai szerint kevesebb, mint 16 millió éve. A tetraploidizációnak köszönhetően 100 - (104) kromoszómájuk van, ami kétszerese más pontyfélékének (Ohno és mtsai., 1967) és a sejtmagjaiban található DNS mennyisége is közel duplája más diploid halakénak (Ohno és Atkin, 1966). A duplikálódott lókuszok többsége több millió évig (generációkon keresztül) kifejeződő képes maradt, ezalatt sok mutáció halmozódhatott fel bennük, melyek aztán aminosav cserékhez vezettek (Larhammar és Risinger, 1994).
A poliploidizáció kedvező hatással volt egyes halfajok kialakulásásra és az evolúciós fejlődésére. Ezek a halak, bár szélesebb genetikai alkalmazkodóképességgel rendelkeznek, mégis diploid társaikhoz hasonló ökológiai fülkét töltenek be.
Fennmaradási előnyük abban rejlik, hogy a genetikai fejlődés során a megnövekedett genetikai állományból az adott élőhelyen a legkedvezőbb gének, allélok maradtak meg, míg a feleslegesek szelektálódtak. A poliploid genetikai túlterhelés azzal küszöbölhető ki, hogy az eredetileg duplikálódott szabályozó gének az evolúció során alkalmazkodtak a diploid státuszhoz, és úgy irányítanak, mintha egy diploid génkészletük lenne, de adott esetben minden egyes génen 2 allél helyett 4 közül választhatnak. Kettő dolgozik, kettő meg szabadon mutálhat, akár a letális irányban is (Váradi 2000; Volff 2005).
Általános tendencia, amit teljes genom duplikáció után tapasztalhatunk, hogy a duplikálódott gének közül sok elveszik darabolódás (kromoszómatörés vagy átrendeződés) következtében, vagy pszeudogénekké válnak.
31
A manapság legnépszerűbb modell, amely a duplikáció fenntartását magyarázza a DDC (Duplication- Degeneration- Complementation) modell (Freeling 2009).
Feltételezi, hogy rögtön a génduplikáció után (pl. a kromoszómaátrendeződések miatt;
vagy káros mutációk miatt) a cisz-szabályozó régió részben elveszik, ez szubfunkcionalizációhoz vezet (10. ábra), így mindkét kópiára szükség van az eredeti funkció betöltéséhez (Rohner 2010).
Bár sok ilyen duplikálódott gén elveszik az evolúció során, ismert az is, hogy az extra génkészlet megtartja a hasonló funkciókat, vagy elég változékony ahhoz, hogy új funkciókat szerezzen (Force és mtsai.,1999).
10. ábra: A duplikálódott génpárok három lehetséges sorsát mutatja. A kis téglalapok az egyedi funkciójú szabályozó elemeket jelölik, a hosszúkás téglalapok pedig az átíródott régiókat. A tömör téglalapok a gén érintetlen régióit jelölik, míg az üres téglalapok null mutációt jelölnek, a háromszögek pedig új funkció kialakulását jelölik. Bal oldalon a degeneratív mutációk miatt a gén nem működik, pszeudogénné vált. Középen a szabályozó régió egy új funkcióval bővült (neofunkcionalizáció), kedvező mutáció történt, így megmaradt a kópia. Jobb oldalon a két paralóg kiegészíti egymás működését (szubfunkcionalizáció), mindkét példány elengedhetetlen a genom teljes működéséhez (Force és mtsai., 1999, Loh és mtsai., 2004).
2.6. Kültakaró és fenotípus
A halak kültakarója lehet csupasz, vagy pikkelyekkel borított. A dolgozatban elsősorban az utóbbi csoporttal foglalkoztunk, pontosabban ezzel összefüggésben a mutációt jelentő, szokványostól eltérő pikkelymintázattal – a pikkelyzet részleges vagy teljes hiányával. A természetben is megfigyelhető jó néhány fajnál a pikkelyzet
32
csökevényesedése (pl. harcsáknál, angolnánál). A ponty alakúak rendjében is van legkevesebb 13 olyan faj (pl. Danionella translucida, Sawbwa resplendens, Phreatichthys andruzzii (Cavallari és mtsai., 2011)), amelyeknek nincs pikkelyük (Rohner 2010; Nelson 2006). Meglehetősen keveset tudunk a pikkelyzet fejlődésének genetikai alapjairól és a természetben található változatairól.
Pikkelyeik a hüllőknek, madaraknak és néhány ízeltlábúnak is van, amelyek azonban számos szempontból eltérnek egymástól. Így például a lepidopterák (lepkék) esetében a szárnyakat zsindelyszerűen átfedő kitinpikkelyeket találunk; ez az úgynevezett „hímpor”, amiről a rend a nevét kapta — a Lepidoptera szó jelentése:
pikkelyes szárnyú. A pikkelyek nemcsak pigmentáltak, de vastagságuktól és elrendeződésüktől, sűrűségüktől, finom lemezes, vagy rácsos szerkezetüktől függően különböző fizikai színeket produkálnak (irizálás stb.). Ezt elősegítik a pikkelyekben található különféle kutikuláris képződmények. A fizikai és kémiai színek kombinációi egyes fajokat rendkívül dekoratívvá tesznek. Ha lekopnak róla a pikkelyek, a szárny üvegszerűen áttetszővé válik.
A hüllők fontos ismertetőjegye, a szarupikkely az epidermisz elszarusodott sejtjeiből áll, bennük véredények és idegek sohasem végződnek (A Pallas Nagy Lexikona sorozat, 1893). A hüllők és madarak esetében is akárcsak a szőrök, az epidermiszből fejlődnek és keratinból vannak, míg a halak pikkelyei az irhában lévő tasakokban indulnak fejlődésnek és nem keratin az alkotójuk (Rohner 2010). A dolgozatban a halak pikkelyeire fókuszálunk, de mint látható ez a pikkely kifejezés is meglehetősen tág.
A halak pikkelyeinek több típusa van (11. ábra):
Placoid (dermal denticles) - porcos halak pikkelyei (cápák, ráják). Szerkezetileg azonosak a gerincesek fogaival. Alapjuk egy csontlemez, belsejében fogszerűen dentinből, felszínén virodentinből áll. Az ilyen típusú pikkelyek nem tudnak növekedni.
Ganoid – pl. zománcos halak (Polypterus bichir bichir) pikkelyei – Tokalakúaknak is lehetnek- rombusz alakúak, izopedin, cosmin, ganoin rétegből állnak, nevüket a ganoin rétegről kapták, melytől fényesek és kemények lesznek.
Cosmoid – a tüdőshalak és a maradványhalak pikkelyei. Növekvő pikkelyek.
Valószínűleg a placoid pikkelyek összeolvadásából keletkeztek.
33
Elasmoid - a valódi csontoshalak pikkelyei. Vékony, hajlékony, átlátszó kollagén csontlapocskák (Bertin 1944). A pikkelyek folyamatosan növekednek, a rárakódott újabb rétegek gyűrűket alkotnak, ezt használják az életkor meghatározásához. A pikkelyek jól regenerálódnak, lehetséges interspecifikus transzplantációjuk is.
Az elasmoid pikkelyeknek két fajtája van, mindkettő cserépszerűen átfed:
- ctenoid (pl. a sügéralakúaknál) apró fogacskák borítják
- cycloid (pl. a pontyféléknél) simább, nincsenek rajta fogacskák,
kerekded. „A héj nem terjed a fejre, pénzei nagyok, kerekdedek; szélük hártyás.” A ponty is ilyen pikkelyekkel rendelkezik.
- A pisztrángsügéren mindkettő elasmoid pikkelyféle megtalálható (Kiss 2000; Rohner 2010).
11. ábra: A különböző pikkelyformák sematikus rajza (Rohner 2010): a) placoid pikkelyek b) ganoid pikkelyek c,d) elasmoid pikkelyek c) cycloid pikkelyek d) ctenoid pikkelyek
A pikkelyek fontos szerepet játszanak a halak élettanában, a védelemben és az új környezethez való alkalmazkodásban. (Sire és mtsai., 1997)
2.6.1. A pikkely és eredete:
Első pillantásra úgy tűnik, hogy a szőrnek, a fogaknak és a pikkelyeknek nem sok közük van egymáshoz, eltekintve, hogy mind a gerincesek „függelékei” (szőr és pikkelyek borítják a testfelszínt szabályos elrendeződésben), de élettani szempontból és evolúciósan is eltérnek egymástól. A fogak és a halak pikkelyei ugyan morfológiailag különbözőek, de mindketten az ősi külső váz elemei és feltételezhetően közös őstől
34
származnak. Kondo és munkatársai (2001.) munkájukban a szőrzetet és a pikkelyeket jelölik meg ugyanilyen vonatkozásban. Morfológiailag és evólúciósan a fogak és a szőrzet is meglehetősen különböznek, de mindkettő fejlődésében részt vesznek bizonyos szabályozási folyamatok, mint például a Hedgehog, a Bone Morphogenetic fehérje és Wnt jelátviteli utak, kaszkád rendszerek.
Kondo és munkatársai (2001) által publikált cikk bizonyítékul szolgál egy, a halak pikkelyének képződéséhez, a szőr és a fogazat normális fejlődéséhez is szükséges jelátviteli út meglétéhez (Sharpe, 2001) (12. ábra).
A medaka (Oryzias latipes) rs-3 lókuszán történt mutáció pikkelyzete csaknem teljes elvesztéséhez vezet. Kimutatták, hogy az rs-3 lókusz az ectodysplasin-A receptor (EDAR)-t kódolja, amely az emlősöknél a szőrzet fejlődésének indulásakor szükséges.
Azonosítottak egy újszerű transzpozont, ami az EDAR első intronjába helyezve rendellenes mRNS érést okoz. Munkájuk bizonyította, hogy az EDAR szükséges a halak pikkelyzetének fejlődéséhez, és azt sugallja, hogy ez egy evolúciósan konzervált molekula, amely a gerincesekben az epithelialis függelékek fejlődéséhez is szükséges (Kondo és mtsai., 2001).
12. ábra: Lehetséges evolúciós kapcsolatok a gerincesek ektodermális és dermális függelékei között, és alapvető struktúrájuk. A szőr follikulusai ektodermális származékok, úgy vélik, hogy hüllők pikkelyeiből alakultak ki. A fogak és a csontos halak pikkelyei dermális eredetűek, feltehetőleg közös őstől, az ősi halak páncélzatából származnak. Az EDA/EDAR jelátviteli rendszer szükséges a szőrzet, a pikkelyzet és a fogak fejlődéséhez is (Sharpe, 2001.).
Az eddigi zebradánió tanulmányok azt mutatják, hogy a pikkelyek a késői ontogenezis során fejlődnek (kb. 30 nappal a termékenyülés után) közel az epidermiszhez. Az első pikkelyek a 8-9 mm-es testhosszúság elérésekor jelennek meg.
35
A pikkely fejlődésének kezdete függ a kortól, hőmérséklettől, tápláléktól, és az egyedsűrűségtől (akváriumban) de általában 25-30 nappal a termékenyülés után indul meg. (Sire és mtsai., 1997; Rohner 2010).
Pontynál a pikkelyzet kifejlődése a kopultyúfedő csont mögött indul meg körülbelül 16-18mm testhosszúság elérésekor. A teljes pikkelyzet kialakulása 25 mm testhosszúság felett valósul meg. A pikkelyzet tömege 1 kg-os pontynál a testtömeg mintegy 4,5%-a (Kiss 2000).
Szabályos elrendeződést mutató pikkelyzet esetén (pikkelyes ponty) a test oldalán le kell számolni a hossztengelyre merőlegesen álló pikkelyoszlopokat, valamint a hossztengellyel párhuzamosan, a hátéltől az oldalvonalig és az oldalvonaltól a hasélig futó pikkelysorokat (Az így kapott értékek a pikkelyképletet adják, ami felhasználható a fajok elkülönítéséhez, határozásához).
A ponty pikkelyképlete: 33 ---- 40
2.7. A garatfog
A pontyfélék határozóbélyegei a garatfogak, amelyek az utolsó (4.) kopoltyúív módosulásából létrejött garatcsonton találhatók. Ezek a fajra jellemző számban, alakban és elrendeződésben vannak jelen. Garatfoga van a csíkféléknek is (Harka és Sallai 2007). Erős fogszerű képleteivel a darabos eledel aprításában vesz részt, szerepük van továbbá a víz kipréselésében, a nyelés elősegítésében. A fogak váltódnak. Állhatnak egy sorban (pl. Tinca, Carassius, Vimba) két sorban (pl. Gobio, Phoxinus, Pelecus) vagy három sorban (pl. Barbus, Cyprinus).
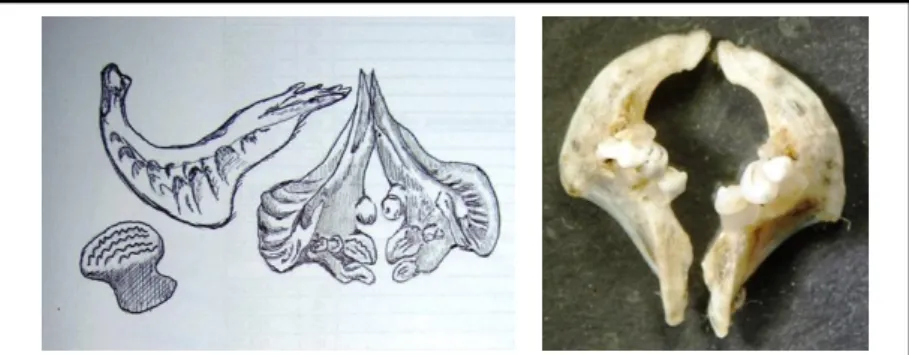
A garatfogak száma - a bal, valamint a jobb oldali csonton a soronként található fog szám - adja a garatfog képletet. A ponty garatfog képlete: 1.1.3-3.1.1. (13.ábra).
13. ábra: A ponty garatfoga (rajz: Szücs Réka a Dictionary of Ichthyology; www.briancoad.com alapján/
Fotó: Szücs Réka)
5-6
5-6
5-6
5-6