Introduction and Perspectives
ANTHONY A. ALBANESE
Nutritional Research Laboratory, St. Luke's Convalescent Hospital, Greenwich, Connecticut
Perusal of the contents of the present volume will reveal that the past decade has witnessed considerable progress in our understanding of the nutrition of proteins and amino acids. It will be noted in particular that the development and application of new procedures has not only greatly expanded the field of inquiry in experimental animals, but also has made possible the conduct of exact nutritional and biochemical measurements in human subjects of all ages and under a variety of physiological and pathological conditions. Under the continuing assault of micro- and radiochemistry, we have learned that the quantitative and qualitative nutrient needs of mammals are subject to significant species (Mitchell, 1959) and individual (Williams, 1959) differences; and that the need for a specific nutrient in any single species is defined by an almost infinite number of variables; and that the protein value of foods is susceptible to an equally large number of vicissitudes. This develop- ment of minutiae, however, does not relegate to the discard the general metabolic laws of nutrient needs and waste per unit of surface area developed so ably in the first quarter of this century (Brody, 1945).
Rather, it points to the fact that maintenance of a necessary metabolic rate and function per unit of surface area varies for specific nutrients from individual to individual, and from species to species. This should come as no surprise if we consider that no two energy converters, organic or inorganic, will function with identical efficiency.
In living organisms these differences are ultimately traceable to varia- tions in available enzymes or other biocatalysts (Martin, 1958). This aspect of human chemistry is not new. It received attention and docu- mentation by Garrod as long ago as 1923, under the title "Inborn Errors of Metabolism." Today, because of the wonders of chromatography, we have micro tests for almost every conceivable organic constituent. Thus, we are able not only to ascertain the presence of classic metabolic aberra- tions (alkaptonuria, phenylketonuria) with greater frequency, but also to discover new and heretofore unsuspected biochemical lesions (Dent, 1947; Harris, 1955). Such lesions have been produced artificially by dep- rivation of a single amino acid (Albanese, 1952; Hall et al.y 1948). Ad- ministration of antimetabolites has been shown to result in similar
1
pathological lesions (Woolley, 1952) and congenital malformations (Warkany, 1958).
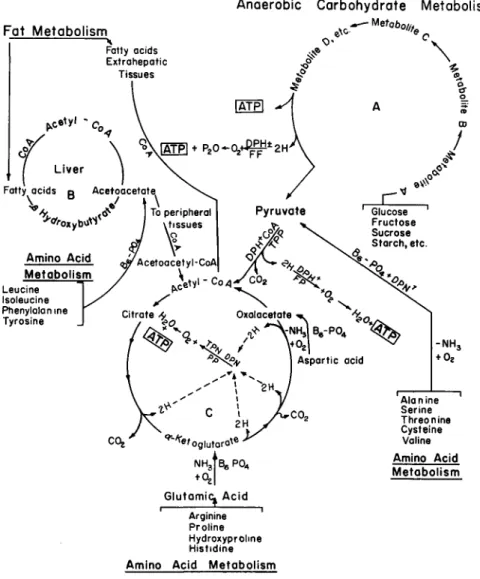
The past decade has also shown that as the periphery of specific areas of study is expanded, there occurs a rapid fusion and eventual disappear- ance of boundaries. Thus, we find that both the catabolic and anabolic phases of the metabolism of the amino acids are inextricably related to the metabolism of fats and carbohydrates (Swanson, 1959), and they cannot be studied or discussed without some reference to nutrition in general. This becomes quickly apparent from a consideration of the currently known metabolic interrelationships of the amino acids (Fig. 1).
Continued study of the nutritional implications of these metabolic link- ages may be expected in the proximate future to yield more exact knowl- edge on specific protein and amino acid needs. Accordingly, it seems worth while here to delineate briefly the current state of development of some of these relationships.
I. VITAMINS AND AMINO ACID METABOLISM
From the metabolic diagram (Fig. 1) it is at once apparent that the biochemistry of amino acids is integrated with the over-all metabolism and all of the vitamins that are dietary essentials are involved in normal metabolism. In general, the role of the vitamins is that of serving essen- tial parts of the molecular structure of the enzyme systems that constitute the metabolic machinery of the cells. Many gross effects, such as lack of growth, are common to a number of vitamin deficiencies, as well as some amino acid deficiencies. It has been shown (Kinney and Follis, 1958) that some pathological lesions produced in experimental animals by amino acid deficiencies are indistinguishable from those caused by vitamin deficiencies. Goldsmith (1956) has skillfully clarified the inter- dependence of the intermediary metabolism of niacin and tryptophan and the effect of this biochemical circumstance on the nutritional needs of these two dietary essentials.
Specific evidences of even more subtle interrelationships are now on hand. Some six years ago, some babies receiving a proprietary milk formula began to show excessive irritability and, in many instances, con- vulsions (Bessey, 1957). On the basis of similar symptoms in experi- mental animals, and a consideration of the way in which the product was manufactured, a deficiency of vitamin B6 was suspected. This sus- picion was soon confirmed by the favorable response of the infants to formulas with an increased vitamin B6 content.
Biochemical studies have revealed that animals deficient in vitamin B6 excrete xanthurenic acid in the urine and this excretion is also increased after the administration of a test dose of tryptophan (Robinson, 1951).
In the absence of vitamin B6? xanthurenic acid, now known to be an abnormal product of tryptophan metabolism, is formed in increasing quantities. In babies with convulsions, xanthurenic acid disappeared after B6 therapy was instituted. In all cases except two, the level of
Fat Metabolism
Anaerobic Carbohydrate Metabolism
- Meto60
Leucine Isoleucine Phenylalamne Tyrosine
Ala nine Serine Threonlne Cysteine Valine Ami no Acid Metabolism
^•OglutarQt·
NH3f B6 P04
+o2| Glutamiq. Acid
Arginine Proline Hydroxyprolme Histidine
Amino Acid Metabolism
FIG. 1. Metabolic integration. ATP = Adenosine triphosphate; D P N + — di- phosphopyridine nucleotide (niacin in structure); T P N + + = triphosphopyridine nucleotide (niacin in structure); TPP — thiamine pyrophosphate (thiamine in structure); FP = flavoprotein (riboflavin in structure); CoA = coenzyme A (pantothenic acid in structure); Β6· Ρ 04 (vitamin B6 in structure). From Bessey (1957).
vitamin B6 required to eliminate xanthurenic acid excretion was con- siderably above that required to eliminate convulsions. Apparently the amount of vitamin B6 required for normal metabolism of tryptophan is greater than the amount required for minimum function of some as yet unknown biochemical properties of nervous tissue. These facts now form the basis for a test of vitamin B6 status in infants.
Evidences, although not as clear-cut as the foregoing, are on hand regarding the relationship of most known vitamins to protein and amino acid nutrition. The important role of vitamin Bi 2 in the biosynthesis of proteins is now generally acknowledged. A deficiency of vitamin Bi 2 reduces nucleic acid synthesis and this causes a decrease in cell division.
This has been demonstrated in human bone marrow cells from pernicious anemia patients (Glazer et al., 1954). Rats receiving Bi 2 will grow on diets deficient in choline and methionine, but containing homocystine.
This suggests that the vitamin has a function in the metabolism of labile methyl groups (Bennett, 1950). In hyperthyroid rats, Bi 2 has a protein- sparing action. This does not prevent weight loss, however, for protein is spared at the expense of other body constituents (Rupp et al., 1951).
This vitamin also permits utilization of nitrogen in animals fed a diet high in soybean protein (Catron et al., 1952; Hsu et al., 1953). Since the methionine content of soybean protein is low, this effect is most likely related to the methionine-sparing activity of Bi2, rather than to a direct effect on protein synthesis.
Less clear are the relationships of vitamins C, A, and E to protein and amino acid nutrition. The lesions which deficiencies of these vitamins induce in both experimental animals and man leave no doubt that they participate in the synthesis of many specialized tissues. For example, one of the most important roles of ascorbic acid involves the formation of collagen in teeth, bone, cartilage, connective tissue, and skin. It pro- motes normal development of the teeth, including both pulp and dentine
(Fish and Harris, 1935), but apparently does not influence the occurrence of dental caries in man. Ascorbic acid has also been reported to be essential for regeneration of damaged nerve tissue (Hines et al., 1944).
Diets high in protein increase the excretion of ascorbic acid.
Although the function of vitamin A in the chemistry of the visual purple is now clearly understood, its function in maintaining the integrity of epithelial cells and as a stimulus for new cell growth, remains obscure.
Vitamin E deficiency leads to progressive muscular dystrophy in experi- mental animals, but vitamin E has no beneficial effect, even in massive doses, on muscular dystrophy occurring in man. The investigative oppor- tunities in this area seem endless.
II. EFFECT OF SOME THERAPEUTIC AGENTS ON PROTEIN AND AMINO ACID NUTRITION
Although the number and diversity of pharmacological agents having rather well-defined effects on metabolism continues to grow, very little effort has been made to date to correlate the metabolic or nutritional disturbances with either pharmacological activity or chemical structure of these agents. The ever-increasing interest in nutrition will, of course, result in a growing need to assay new therapeutic agents, not only for their pharmacological properties, but also for their nutritional char- acteristics.
A. ANTIBIOTICS
The subject of antibiotics in nutrition was admirably reviewed by Jukes (1955). It is clear and well established from available evidence that an apparent improvement in the nutritional status of animals and humans may often be produced by adding small quantities of certain antibiotics to the diet. The mechanism by which their effects on nutrition are produced appears to be secondary to their antibacterial action.
Stokstad (1955) has discussed the effect of antibiotics on vitamin re- quirements. He notes that while a vitamin-sparing action can explain in part the growth response on diets marginal in vitamins, it does not account for the growth observed in nutritionally complete rations. The vitamin-sparing effect of the antibiotics has been observed to occur for both water-soluble and fat-soluble vitamins. A thorough consideration of the data leads Coates and Kon (1955) to believe that although the growth-promoting effect of antibiotics depends to a large extent on their antibacterial properties, their direct effect on certain metabolic processes cannot be overlooked. Hence, it is conceivable that given in the diet, they exert a small but definite pharmacological action which contributes to their beneficial effects on growth.
There are currently growing indications that the combined admin- istration of antibiotics and amino acids may help resolve some nutritional problems in technologically underdeveloped areas of the world. Because of the impracticability of supplying to many children living in tropical countries diets rich in high quality proteins, it often happens that their growth is slow; and they become stunted. Since antibiotics and other substances, such as lysine, have growth-stimulation effects on children in temperate zones, Loughlin and associates (1957-1958) decided to try these agents in chronically malnourished tropical children. Sixty-four children (6-16 years) at a rural school in Haiti were selected for study.
After their conditions had been analyzed they were fed under controlled study conditions; the children were separated into four matched groups.
In three, supplements to their diets were used in the form of 10 mg. and 50 mg. of oxytetracycline and 1 gm. of lysine, respectively. The fourth group received placebos. The study was run on a double blind basis, and lasted for 6 months. On analysis it was found that the group that received 50 mg. of oxytetracycline daily manifested the best growths. In growth factors, the group receiving lysine came out second. There were no toxic or untoward effects noted as the result of this long-range administration of oxytetracycline or lysine.
A critical evaluation of the role of nutrition in the prophylaxis and treatment of disease reveals many other areas in which proteins and antibiotics may jointly serve to improve over-all metabolism and reduce the incidence of infections (Halpern, 1955).
B. STEROIDS
The rapidly increasing availability and usage of new synthetic cata- bolic and anabolic steroids provides a limitless opportunity for study of basic metabolic problems of proteins and amino acids (Luetscher and Lieberman, 1958). Some of these possibilities are touched upon in other sections of this volume. It is clear from the evidence on hand that control of the nitrogen depletion effects of the corticoids can be achieved in part by protein supplementation of the dietary and, in some instances, by ad- ministration of small quantities of some of the essential amino acids.
Similarly, it appears that effects of some anabolic steroids may be en- hanced by amino acid fortification of the dietary. Closer scrutiny of this nutritional synergism may lead to attainment of maximal anabolism at very small dosage levels of these steroids, and thereby a reduction in the incidence of virilism and other untoward effects.
C. HYPOGLYCEMIC AGENTS
The fortuitous observation of Janbon et al. (1942) that the newer sulfonamides produced a disorder very similar to hypoglycemia, led to Loubatiere's studies (1957) on the mechanism of action of the sulfo- nylureas in diabetes, and eventually to an evaluation of these substances for the control of diabetes (Dolger, 1957). A pilot study on two diabetics given rather large doses of tolbutamide for about 2 weeks showed no changes in respiratory quotient in either subject, and a slightly negative nitrogen balance in one (Goetz, 1957). McGavack and associates (1957) observed that whereas 1 gm. daily of carbutamide did not influence the uptake of radioiodine, 2 gm. daily of carbutamide progressively de- pressed I131 uptake to 56% of the control value at the end of the ninth week of treatment.
Numerous reports have appeared implicating salicylates in carbohy-
drate metabolism, particularly the reported ability of aspirin to lower blood sugar levels in diabetics (Smith, 1953). A definite hypoglycemic effect of salicylates was demonstrated in rheumatic fever patients by Albanese and co-workers (1955). Results of the study which involved ten subjects (5-18 years) indicated that the magnitude of fasting hypo- glycemia produced by aspirin bears a direct relationship to the duration and dosage of therapy, and the age and weight of the patient. Subsequent studies by this group (Albanese, 1959) have disclosed that the hypo- glycemia of prolonged and massive salicylate therapy is sometimes ac- companied by a persistent decrease in fasting plasma amino nitrogen levels—a sensitive criterion of protein nutrition.
A further and heretofore unsuspected relationship of amino acid and carbohydrate metabolism has been described by Seltzer and Smith (1958). These investigators reported that in normal subjects and in mild or severe diabetics, tolbutamide (Orinase) and indole-3-acetic acid, a physiological end product of tryptophan metabolism, exerted identical effects on blood glucose and plasma insulin activity in the same indi- viduals.
D. PHRENOTROPIC DRUGS
The pharmacology of this rapidly growing family of substances was thoroughly reviewed in 1957 by a very able conference group (Kety, 1957). The psychotherapeutic activity of some of the agents in common use was reported to correlate well with their inhibition of oxidative phosphorylations (Fig. 1) and/or cytochrome oxidase. Preliminary ob- servations (Abood and Romanchek, 1957) have disclosed that many indole substances and derivatives of the urine of phenylketonurics are also formidable inhibitors of cellular oxidations and phosphorylations.
Recently, these aromatic substances, which are metabolically derived from tyrosine and tryptophan, were suspected of a possible role in the biogenesis of schizophrenia-producing compounds.
It is interesting to note in this connection that dietary deficits of niacin, which is also derived biosynthetically from tryptophan, have been implicated in the etiology of various psychiatric symptoms which were relieved by niacin administration (Jolliffe et al., 1940). Isoniazid, an isomer of nicotinic acid, has been observed to produce marked euphoria early in experimental trials in tuberculosis (Bennett et al., 1954).
From the foregoing, it is apparent that investigations on the bio- chemical properties of proteins and amino acids may have ramifications in many areas far removed from their nutritive functions. Also, from the preceding, as well as the pages which follow, it will become abundantly
clear that future progress of the science of nutrition will depend more and more on the mutual understanding and collaborative efforts of in- vestigators working at all levels of biochemical organization—from two- carbon molecules to the whole man.
REFERENCES
Abood, L. G., and Romanchek, L. (1957). Ann. Ν.Ύ. Acad. Set. 66, 812.
Albanese, A. A. (1952). J. Clin. Nutrition 1, 44.
Albanese, A. A. (1959). Unpublished data.
Albanese, A. A., Higgons, R. A., Avery, S. G., and DiLallo, R. (1955). Ν.Ύ. State J. Med. 55, 1167.
Bennett, I. F., Cohen, D., and Starer, E. (1954). A.M.A. Arch. Neurol. Psychiat.
71, 54.
Bennett, M. A. (1950). /. Biol Chem. 187, 751.
Bessey, O. A. (1957). /. Am. Med. Assoc. 164, 1224.
Brody, S. (1945). "Bioenergetics and Growth/' Reinhold, New York.
Catron, D. V., Richardson, D., Underkofler, L. A., Maddock, H. M., and Friedland, W. C. (1952). /. Nutrition 47, 461.
Coates, M. E., and Kon, S. K. (1955). Abstr. 3rd Intern. Congr. Biochem. Brussels pp. 448-452.
Dent, C. E. (1947). Biochem. J. 41, 240.
Dolger, H. (1957). Ann. Ν.Ύ. Acad. Set. 71, 275.
Fish, E. W., and Harris, L. J. (1935). Brit. Dental J. 58, 3.
Garrod, A. E. (1923). "Inborn Errors of Metabolism," 2nd ed. Oxford Univ.
Press, London and New York.
Glazer, H. S., Mueller, J. F., Jarrold, T., Sakurai, K., Will, J. J., and Vilter, R. W.
(1954). /. Lab. Clin. Med. 43, 905.
Goetz, F. C. (1957). Ann. Ν.Ύ. Acad. Set. 71, 46.
Goldsmith, G. A. (1956). /. Am. Dietet. Assoc. 32, 312.
Hall, W. K., Bowles, L. L., Sydenstricker, V. P., and Schmidt, H. L., Jr. (1948).
J. Nutrition 36, 277.
Halpern, S. L. (1955). Ann. Ν.Ύ. Acad. Sei. 63, 147.
Harris, H. (1955). Abstr. 3rd Intern. Congr. Biochem. Brussels pp. 467-473.
Hines, H. M., Lazere, B., Thomson, J. D., and Cretzmeyer, C. H. (1944).
/. Nutrition 27, 303.
Hsu, J. M., Stern, J. R., and McGinnis, J. (1953). Arch. Biochem. Biophys. 42, 54.
Janbon, M., Chaptal, J., Vedel, A., and Schaap, J. (1942). Montpellier med. 21-22, 441.
Jolliffe, N., Bowman, K. M., Rosenblum, L. A., and Fein, H. D. (1940). J. Am.
Med. Assoc. 114, 307.
Jukes, T. H. (1955). "Antibiotics in Nutrition." Medical Encyclopedia, New York.
Kety, S. S. (1957). Ann. Ν.Ύ. Acad. Set. 66, 417.
Kinney, T. D., and Follis, R. H., Jr. (1958). Federation Proc. 17 ( 3 ) , 1-162.
Loubatieres, A. (1957). Ann. Ν.Ύ. Acad. Set. 71, 4.
Loughlin, E. H., Joseph, A. A., and Alcindor, L. (1957-1958). Antibiotics Ann.
1957/58, 99.
Luetscher, J. A., Jr., and Lieberman, A. H. (1958). A.M.A. Arch. Intern. Med.
102, 314.
McGavack, T. H., Seegers, W., Haar, H. O., Enzinger, J., and Erk, V. O. (1957).
Ann. NY. Acad. Sei. 71, 268.
Martin, G. J. (1958). "Clinical Enzymology." Little, Brown, Boston, Massa- chusetts.
Mitchell, H. H. (1959). This volume, Chapter 2.
Robinson, F. A. (1951). "Vitamin B Complex." Wiley, New York.
Rupp, J., Paschkis, K. E., and Cantarow, A. (1951). Proc. Soc. Exptl. Biol. Med.
76, 432.
Seltzer, H. S., and Smith, W. H. (1958). /. Lab. Clin. Med. 52, 945.
Smith, M. J. (1953). /. Pharm. and Pharmacol. 5, 81.
Stokstad, E. L. R. (1955). Abstr. 3rd Intern. Congr. Biochem. Brussels pp. 445-448.
Swanson, P. P. (1959). This volume, Chapter 8.
Warkany, J. (1958). /. Am. Med. Assoc. 168, 2020.
Williams, R. J. (1959). This volume, Chapter 3.
Woolley, D. W. (1952). "A Study of Antimetabolites." Wiley, New York.