DOKTORI (PhD) ÉRTEKEZÉS
BUZSÁKI KAMILLA
PANNON EGYETEM GEORGIKON KAR KESZTHELY
2011
PANNON EGYETEM GEORGIKON KAR Növényvédelmi Intézet
Herbológiai és Növényvédőszer Kémiai Osztály
Állat- és Agrárkörnyezet-tudományi Doktori Iskola
iskolavezető:
DR. HABIL. ANDA ANGÉLA egyetemi tanár, az MTA doktora
témavezető:
DR. HABIL. LEHOCZKY ÉVA egyetemi tanár, az MTA doktora
DR. HABIL. BÉRES IMRE Professor emeritus
A MANDULAPALKA (CYPERUS ESCULENTUS L. VAR.
LEPTOSTACHYUS) ELTERJEDÉSE, KÁRTÉTELE, TÁPANYAGTARTALMÁNAK VIZSGÁLATA
Készítette:
BUZSÁKI KAMILLA
KESZTHELY
2011
A MANDULAPALKA (CYPERUS ESCULENTUS L. VAR.
LEPTOSTACHYUS) ELTERJEDÉSE, KÁRTÉTELE, TÁPANYAGTARTALMÁNAK VIZSGÁLATA
Értekezés doktori (PhD) fokozat elnyerése érdekében írta:
Buzsáki Kamilla
KÉSZÜLT A PANNON EGYETEM
ÁLLAT-ÉS AGRÁRKÖRNYEZET-TUDOMÁNYI DOKTORI ISKOLA KERETÉBEN
Témavezető: Dr. Lehoczky Éva
Elfogadásra javaslom (igen / nem) …….……….
aláírás Témavezető: Dr. Béres Imre
Elfogadásra javaslom (igen / nem) …….……….
aláírás
A jelölt a doktori szigorlaton ...%-ot ért el.
Keszthely, ……… ……….
a szigorlati bizottság elnöke
Az értekezést bírálóként elfogadásra javaslom:
Bíráló neve: …... …... igen /nem
……….
aláírás Bíráló neve: …... …... igen /nem
……….
aláírás
A jelölt az értekezés nyilvános vitáján …...%-ot ért el.
Keszthely, 2011………. ……….
a Bíráló Bizottság elnöke A doktori (PhD) oklevél minősítése…...
………
Az EDHT elnöke
TARTALOMJEGYZÉK
1. KIVONATOK……….………...…3
1.1. Magyar nyelvű kivonat…….………..……….3
1.2. Angol nyelvű kivonat – Abstract……….5
1.3. Német nyelvű kivonat - Deutschsprachiger Auszug………6
2. BEVEZETÉS……….……..………..…7
3. IRODALMI ÁTTEKINTÉS………...………...9
3.1. Elnevezése és rendszertani besorolása………9
3.1.1. Elnevezése………...9
3.1.2. Rendszertana………...…9
3.2. Származása és elterjedése……….10
3.3. Gazdasági jelentősége, kártétele………...12
3.4. Morfológiája………..13
3.5. Szaporodása………...15
3.6. Az allelopátia jelensége………17
3.7. Kompetíció………18
3.8. A tápanyagokért történő versengés………...19
3.9. Védekezési lehetőségek a mandulapalka ellen……….20
4. ANYAG ÉS MÓDSZER………..….24
4.1. Laboratóriumi vizsgálatok……….24
4.1.1. Maghozam vizsgálatok……….24
4.1.2. Csírázásbiológiai vizsgálatok………24
4.1.3. A mandulapalka allelopátiája………24
4.2. Üvegházi tenyészedényes vizsgálatok………...25
4.2.1. Allelopátiás vizsgálatok………25
4.2.1.1. A mandulapalka hajtás- és föld alatti rész rmaradványának hatása a tesztnövények csírázására és fejlődésére üvegházi kísérletben………...25
4.2.1.2. A mandulapalka hajtásából és föld alatti részekből készített növénykivonat hatása a tesztnövények csírázására és fejlődésére üvegházi kísérletben………...25
4.2.2. A mandulapalka fejlődésének vizsgálata növekedésanalízis módszerével…...26
4.2.3. A mandulapalka tápelem-koncentrációjának vizsgálata………...28
4.2.4. Korai kompetíciós vizsgálat……...………...28
4.3. Eredmények statisztikai elemzése……….……….30
5. EREDMÉNYEK ÉS KÖVETKEZTETÉSEK………...31
5.1. Laboratóriumi vizsgálatok……….31
5.1.1. Maghozam vizsgálat………..………...31
5.1.2. Éves csírázási ritmus vizsgálat……….31
5.1.3. A mandulapalka hajtásából és föld alatti részekből készített növénykivonat hatása a tesztnövények csírázására laboratóriumi kísérletben………...32
5.2. Üvegházi tenyészedényes vizsgálatok………...36
5.2.1. Allelopátia vizsgálatok………..36
5.2.1.1. A mandulapalka hajtás- és föld alatti részek maradványának hatása a tesztnövények csírázására és fejlődésére üvegházi kísérletben…………...…36
5.2.1.2. A mandulapalka hajtásából és föld alatti részekből készített növénykivonat hatása a tesztnövények csírázására és fejlődésére üvegházi kísérletben…….38
5.2.2. Növekedésanalízis……….41
5.2.3. A mandulapalka tápelem-koncentrációjának dinamikája ………....49
5.2.4. Korai kompetíciós vizsgálat….……….…55
5.2.5. Tápanyagokért folytatott versengés……….………...64
6. ÖSSZEFOGLALÁS……….…….67
7. KÖSZÖNETNYILVÁNÍTÁS………...71
8. IRODALOMJEGYZÉK………...………...72
9. AZ ÉRTEKEZÉS TÉZISEI………...79
9.1. Magyar nyelvű tézispontok………79
9.2. Angol nyelvű tézispontok………..81
3
1. KIVONATOK
1.1.Magyar nyelvű kivonat
A mandulapalka (Cyperus esculentus L. var. leptostachyus Boeck.) valamennyi kontinensen megtalálható. Európában jelentős megjelenése 1980-és 1995 között volt. Magyarországon a gyomosító Cyperus esculentus L. előfordulását Dancza István figyelte meg 1993-ban.
Magyarországon jelenleg négy tájegységen, közel húsz település határában fordul elő.
A mandulapalka jelentőségét mutatja, hogy az 1970-es években a világ legfontosabb gyomnövényei között a 16. helyen állt. Napjainkban az Inváziós Növények Munkabizottsága a jelentős gazdasági kárt okozó, nemzetközi szinten kiemelt inváziós gyomnövények között tartja számon. Jelentős kártételt a tavaszi vetésű, ritka növényállományú kapás kultúrákban okoz.
Munkákban célul tűztük ki, hogy a mandulapalka biológiájáról szóló ismereteket gyarapítsuk, kiegészítsük hasznos adatokkal, amelyek később a hatékony gyomszabályozás kidolgozásához alapot nyújtanak.
Ezen szempontok alapján a következő kísérleteket végeztük:
- maghozam vizsgálat - csírázásbiológiai vizsgálat - növekedés analízis vizsgálat
- tápelem-koncentráció és tápelemtartalom vizsgálat - allelopátia vizsgálatok
- helyettesítési kompetíciós vizsgálat
Maghozam vizsgálataink során 100 db virágzat maghozamát vizsgáltuk, az átlagos magszám a vizsgált virágzatokban 155 db volt.
Csírázásbiológiai vizsgálataink során megállapítottuk, hogy a mandulapalka magok márciustól július elejéig kiegyenlítetten csíráznak, majd a csírázási erély csökken és októberre megközelíti a minimumot.
A mandulapalka növekedés analízise során vizsgáltuk a növények levélterület indexét (LAI), specifikus levélterületét (SLA), relatív növekedési sebességét (RGR), relatív levélterület növekedési arányát (RLGR) és levélterület arányát (LAR).
4 A mandulapalka tápelemvizsgálata során a nitrogén, a foszfor, a kálium és a kalcium koncentrációjának és tartalmának változását vizsgáltuk a tenyészidőszak folyamán.
A mandulapalka allelopátiáját vizsgáltuk laboratóriumban és tenyészedényes kísérletekben.
Az eredmények alapján megállapítható, hogy a mandulapalka hajtás- és föld alatti részekből készült kivonata, valamint a növénymaradványok nem befolyásolták szignifikánsan a tesztnövények csírázást a laboratóriumi és az üvegházi kísérletekben. Amennyiben a tesztnövényeket a mandulapalka friss hajtáskivonatával öntöztük, illetve a hajtásmaradványokat a tenyészedények tápközegébe kevertük, a tesztnövények fejlődésére serkentő hatást tapasztaltunk. A föld alatti részekből készült kivonat beöntözéskor és a belőle készült növényi maradvány gátolta a tesztfajok fejlődését.
Helyettesítési kompetíciós vizsgálatok során megállapítottuk, hogy a napraforgó és a kukorica esetén, fejlődésük kezdeti szakaszában nem okoz jelentős hajtás- és gyökérfrisstömeg és száraztömeg csökkenést a gyomnövény jelenléte.
Néztük a kukorica- és a mandulapalka hajtások és a gyökerek közötti tápanyagokért folytatott versengést. Laboratóriumban vizsgáltuk a levelek és a gyökerek százalékos N, P2O5 és K2O tartalmát. Megállapítható, hogy a kukorica tápanyagfelvételét nem befolyásolta negatívan a gyomnövény jelenléte.
5 1.2. Angol nyelvű kivonat – Abstract
YELLOW NUTSEDGE (CYPERUS ESCULENTUS L. VAR. LEPTOSTACHYUS) SPREADING, DAMAGE AND INVESTIGATION OF NUTRIENT CONTENT
The yellow nutsedge (Cyperus esculentus L. var. leptostachyus Boeck.) can be found on every continents. Its considerable occurrence in Europe took place between 1980 and 1995. In Hungary István Dancza reported first time about this weed, in 1993. Presently it occurs here at four districts, on fields of near to 20 settlements.
The importance of yellow nutsedge shows the fact, that in 1970 it took the 16th place among the most important weds of the world. Presently the Working Committe of Invasive Weeds has put it on the list of ’especially invasive weed’ worldwide. It causes considerable damages mainly in the spring-sown scanty rootcrop cultures.
Our aim was to enrich our knowledge on the biology of this plant, to collect important data, which will give a basis in elaboration of an effective weed control.
For this we have made the following investigations:
Investigation on its seed production Investigation on its germination biology Growing analysis
Investigation on its nutrient uptake and nutrient concentration Investigation on allelopathy
Investigation on its ability to compete with cultivated plants.
We have investigated the seed production of yellow nutsedge in case of 100 inflorescences. In the average the seed production was 155 seed per inflorence, with a range of 0 – 466.
The seeds of yellow nutsedge started to germinate at the end of March. The germination was from March to June continuous and equal in rate, with average rates between 47 to 64 %.
Under glashouse tests the extracts of underground parts of yellow nutsedge showed an inhibition effect on the biomass of the test plants.
We have examined the change of nitrogen, phosphorus and potassium nutrient contentration in the roots and shoots of yellow nutsedge during a year.
6 1.3. Német nyelvű kivonat – Deutschsprachiger Auszug
DAS ERDMANGELGRAS (CYPERUS ESCULENTUS L. VAR. LEPTOSTACHYUS) VERBREITUNG, SCHADEN UND NÄHRSTOFF-ANALYSE
Das Erdmandelgras (Cyperus esculentus L. var. leptostachyus Boeck.) kommt an alle Kontinenten vor. In Europa tauchte es in grösseren Mengen zwischen 1980 und 1995 auf.
Sein Vorkommen in Ungarn erstmals István Dancza beobachtete in 1993. Es kommt in Ungarn gegenwärtig an vier Gegenden, an Felder von 20 Ansiedlungen vor.
Die Bedeutung der Pflanze zeigt die Tatsache, dass die Arbeitskommission für Aufsicht von Invasive Pflanzen hat es neuerdings auf internationalen Ebene als wesentliche Schaden verursachende invasives Unkraut eingereiht. Es macht wesentliche Schaden in den in Frühling ausgesäten schütteren Hackfrüchte.
Das Ziel unser Arbeit war die Kenntnisse über die Biologie von Erdmandelgras zu mehren, diese mit nützliche Daten ergänzen, die dann später Gründe für eine effektive Unkrautregelungsmaßnahme dienen können. Dafür haben wir Untersuchungen im Labor und Topfkulturen durchgeführt.
Unsere wichtigste Ergebnisse:
Wir haben festgestellt, dass die Zahl der Samen per Infloreszenz schwankt zwischen 0 und 466, durchschnittlich ist es 155.
Wir konnten feststellen, dass die Samen von März bis Ende Juni gleichmäßig keimen, im Durchschnitt 47-64 % der Samen auskeimen können.
Unser Untersuchungen an Allelopathien zeigen, dass unter Gewächshausumstände die unterirdische Teile von Erdmandelgras ein hemmende effekt an biomasseproduktion von Testpflanzen ausüben.
Wir bestimmten das Nährstoffgehalt von Erdmandelgras in Hinsicht auf Stickstoff-, Phosphor-, Kalium- und Calciumkonzentration.
7
2. BEVEZETÉS
Megközelítőleg 250000 növényfaj él a Földön (Standovár, 2001). Ebből mintegy 6700 gyomnövény befolyásolja a mezőgazdasági termelést. A világviszonylatban gondot okozó és fontos gyomnövénynek tekinthető fajok száma mindössze 200. Ebből 76 gyomfaj sorolható a világ legveszélyesebb gyomnövényei közé (Hunyadi, 2000).
A „gyom” fogalma nem tudományos fogalom, hanem köznyelvi kifejezés, és a növényeknek széles körét értik alatta (Wells, 1978). A gyomnövények definíciójának kérdésével számos szerző foglalkozott, szubjektív és ökológiai meghatározásokat különböztethetünk meg. A hazai szakirodalomban megtalálható szubjektív gyomdefiníciók közül Hunyadi Károlyét (1974) tekinthetjük a legteljesebbnek: „Gyomnövénynek nevezünk bármelyik fejlődési stádiumban levő olyan növényt vagy növényi részt (rizóma, tarack, hagyma, hagymagumó stb.), amely ott fordul elő, ahol nem kívánatos”. Az ökológusok a gyomokat a bolygatáshoz (talajművelés stb.) legjobban alkalmazkodó növényeknek tekintik.
Számos olyan gyomfaj károsít szántóföldjeinken, melyek ellen a védekezés kiemelt jelentőségű. Ezen gyomnövények közé tartoznak az inváziós növények is. Az inváziós növények (özönnövények) olyan meghonosodott nem őshonos növények, amelyeknek – gyakran igen nagy számban – létrehozott szaporítóképletei viszonylag rövid idő alatt a szülőegyedektől jelentős távolságra is eljutnak és ezáltal nagy területen képesek elterjedni.
Egyik csoportja az úgynevezett átalakító növények, amelyek az egyes ökoszisztémák és tájak jellemző sajátságait, állapotát, megjelenését vagy természetét azok jelentős területein megváltoztatják.
Magyarországon a meghonosodott neofitonok száma 150 fajra tehető, ebből 30-40 növényfaj tekinthető inváziós fajnak (Mihály - Botta-Dukát, 2004).
A hazánkban előforduló, Cyperaceae családba tartozó fajok közül kettő szerepel az inváziós fajok listáján, a Cyperus difformis és a Cyperus esculentus L., utóbbi az átalakító fajok közé is tartozik.
8 Munkákban célul tűztük ki a Cyperus esculentus L. gyomnövényről való ismereteink gyarapítását. A kitűzött cél érdekében a következő vizsgálatokat végeztük:
- Csírázásbiológiai vizsgálat - Növekedés analízis vizsgálat - Tápelemtartalom vizsgálat - Allelopátia vizsgálat
- Korai kompetíciós vizsgálat
9
3. IRODALMI ÁTTEKINTÉS
3.1. Elnevezése és rendszertani besorolása
3.1.1. Elnevezése
A mandulapalka tudományos neve Cyperus esculentus L. A világon öt változata ismert:
Cyperus esculentus L. var. sativus Boeck.
Cyperus esculentus L. var. esculentus L.
Cyperus esculentus L. var. leptostachyus Boeck.
Cyperus esculentus L. var. heermanni Kükenth.
Cyperus esculentus L. var. macrostachyus Boeck.
Az öt változat közül egy termesztett (Cyperus esculentus L. var. sativus Boeck.) és négy változat gyomosít (Schippers et al. 1995).
Az esculentus latin eredetű szó, jelentése ehető. A termesztett változat elnevezései: ehető palka, csemegepalka, gumós palka, mandulafű (Priszter, 1998). A XIX. századi Magyarország területén mondola palka (Diószegi – Fazekas, 1807) és mandulapalka (Farkas, 1866) neveken közölték. Napjainkban a mandulafű (Makai el al., 2004) nevet alkalmazza a szakirodalom gyakrabban, bár a földi mandula (Megyeri, 1992; László, 1999) elnevezés is előfordul.
Mivel a gyomosító változat hazánkban korábban nem volt ismert, ezért a termesztett változattól való megkülönböztetésre a mandulapalka elnevezést javasolta Dancza és Fischl (2000).
A gyomnövény angol neve: yellow nutsedge, német neve: Erdmandel, spanyol neve: chufa, francia neve: souchet comestible, holland neve: knolcyperus (Dancza és mtsai., 2005).
3.1.2. Rendszertana
Rendszertani besorolása az Új magyar füvészkönyv alapján (Király G., 2009):
Törzs: Zárvatermők (Angiospermatophyta)
Osztály: Egyszikűek (Monocotyledonopsida) Család: Palkafélék (Cyperaceae)
Nemzetség: Cyperus
10 A Cyperus nemzetségnek három faja, a barna palka (Cyperus fuscus L.), a rizspalka (Cyperus difformis L.) és a termesztett mandulafű (Cyperus esculentus L. var. sativus) fordult csak elő Magyarországon a mandulapalka (Cyperus esculentus L. var. leptostachyus ) 1993-as első megfigyeléséig (Simon, 1992).
A Cyperus esculentus-nak öt változata ismert:
Cyperus esculentus L. var. sativus
Cyperus esculentus L. var. esculentus L.,
Cyperus esculentus L. var. leptostachyus Boeck., Cyperus esculentus L. var. heermanni Kükenth. és a
Cyperus esculentus L. var. macrostachyus Boeck. (Schippers et al. 1995).
3.2. Származása és elterjedése
A mandulapalka származási helye Afrika északi, trópusi-szubtrópusi vidéke, a trópusi Egyenlítőtől a hideg éghajlatú Alaszkáig előfordul.
A gyomosító változatok előfordulása: a Cyperus esculentus L. var. esculentus L. Dél- Európában, Ázsiában, Észak-Amerikában, a Cyperus esculentus L. var. leptostachyus Boeck.
Ny-Európában, Észak- és Dél-Amerikában, a Cyperus esculentus L. var. heermanni Kükenth.
és a Cyperus esculentus L. var. macrostachyus Boeck. az Egyesült Államokban, valamint Hollandiában jellemző (Schippers et al. 1995).
A gyomosító változatok Európában adventív növények, csupán az 1970-es évek elejétől ismertek, a Leydeni Herbárium 1975-ben gyűjtött herbáriumi lapja volt a Cyperus esculenuts var. leptostachyus gyomosító változat első bizonyító példánya. Valószínűleg az amerikai kontinensről Gladiolus szállítmánnyal kerülhetett Hollandiába. A faj azonosítása, mint új, veszélyes gyomnövény 1981-ben történt meg. A nyugat-európai populációk változat szintű azonosítása és elterjedésük vizsgálata az 1990-es évek elejétől folyik. Napjainkban az európai országok közül előfordul Németországban, Hollandiában, Belgiumban, Franciaországban, Portugáliában, Ausztriában, Horvátországban, Svájcban, Olaszországban (Dancza, 2004).
Magyarországon a gyomosító Cyperus esculentus L. spontán előfordulását Dancza István figyelte meg 1993 augusztusában, Hévíz-Keszthely határában, kukorica kultúrában, meszes lápi talajon. Ezt követően 1998-ban megfigyelték Pápasalamon határában, szintén
11 kukoricában. 2001-ben Szigetszentmiklós határában kukorica, valamint kardvirág földeken (Dancza, 2002). 2002-ben Lajosmizsén zöldségszaporító telepen, kardvirág ültetvényben, intenzív almásban. Ugyanebben az évben megtalálták Somogytúron, Somogybabodon, Vadépusztán, Karádon, Barcson. 2003-ban megjelent Dejtáron, erdészeti csemetekertben, valamint Kaposvár határában, kukorica kultúrában (Dancza 2004), (1. ábra).
Ezen adatokat kiegészíteném egy saját tapasztalattal, 2011-ben feltűnt a gyomnövény Zala megyében, Barátsziget közelében, kukorica kultúrákban.
A gyomosított terület nagysága változó: Keszthely és Hévíz határában 2 ha, Pápasalamonban 280 ha, Szigetszentmiklóson és környékén 50 ha, Somogy megyében mintegy 10.000 hektáron fertőz, ebből 2500-3000 hektáron rendszeresen védekezni kell ellene (Hoffmanné 2004).
1.ábra: A mandulapalka elterjedése Magyarországon 2004-ben (Dancza, 2004)
Valószínűsíthető, hogy hazánkba való behurcolása a fertőzött Gladiolus szaporítóanyag mellett fertőzött kukorica vetőmaggal is történt (Dancza és mtsai., 2005).
Az ötödik országos szántóföldi gyomfelvételezés (2007-2008) során a mandulapalka előfordulását is vizsgálták. Az alábbi felvételi pontokon találták meg: Kaposvár-Kaposfüred, Enying, Beremend, Ózd, Tiszatarján, Prügy (2. ábra). A felvételezés eredményei alapján az összes átlagos borítási értéket tekintve a tavasszal felvételezett kukoricában a 87. (0,0169%), a nyáron felvételezett kukoricában a 81. (0,0163%) helyen szerepel (Novák és mtsai., 2009).
12 2. ábra: A mandulapalka előfordulása az ötödik szántófoldi gyomfelvételezési pontokon
Terjedését segíti, hogy kis igényű a termőhellyel szemben, laza homok talajtól kötött agyagtalajig előfordul (Gieske, 1993). Erről számol be Fertsak (2003) is, minden talajtípuson előfordul, bár előnyben részesíti a nedves termőhelyeket. Művelt és műveletlen területen egyaránt megtalálható, a talajnedvességet tág határok között tolerálja (Jenkins et al., 1941;
Holm et al., 1977; Mulligan et al., 1976).
Hazánkban Hoffmanné (2005) Somogy megyében figyelte meg, hogy milyen talajtípusokon jelenik meg a mandulapalka. Megtalálták agyagbemosódásos erdőtalajon, Ramann-féle erdőtalajon és rozsdabarna erdőtalajon, karbonátos és nem karbonátos rétitalajon, lápos rétitalajon, lecsapolt tőzeges láptalajon, és karbonátos humuszos öntéstalajon.
3.3. Gazdasági jelentősége, kártétele
Szakirodalmi adatok alapján a mandulapalka az 1970-es években a világ legfontosabb gyomnövényei között a 16. helyen szerepelt (Holm et al., 1977).
Napjainkban az Európai és Földközi-tenger melléki Növényvédelmi szervezet IAS Panel (Inváziós Növények Munkabizottság) a legjelentősebb gazdasági kárt okozó, nemzetközi szinten kiemelt inváziós gyomnövények között tartja számon (EPPO, 2011).
Magyarországon a dísznövény szaporítóanyagok forgalomba hozataláról szóló 62/2004.(IV.27.) FVM rendelet kimondja, hogy „Az értékesítésre szánt dísznövény szaporítóanyagnak külső formáját tekintve valószínűsíthetően mentesnek kell lennie minden olyan, a minőséget hátrányosan befolyásoló károsítótól, illetve károsító jelenlétére utaló
13 tünettől vagy jeltől, amely felhasználhatóságát csökkentené, különös tekintettel az 1. sz.
mellékletben felsorolt károsítókra.” Ezen mellékletben a Cyperus esculentus a Gladiolus L., Lilium L. és Narcissus L. fajoknál egyéb károsítókén szerepel.
A mandulapalkára jellemző, hogy az árnyékot nem kedveli, sűrű állományú kultúrákban nem képes károsítani (Fertsak, 2003). Jelentős kártételt a tavaszi vetésű, ritka növényállományú kapás kultúrákban okoz. Elsősorban kukoricában, napraforgóban, burgonyában és cukorrépában károsít (Gieske, 1993).
Hazai tapasztalatok alapján, erős fertőzés esetén, a termésveszteség kukoricában akár a 30- 40%-ot is elérheti (Dancza és mtsai., 2005).
A Cyperus esculentus L. termesztett változatát, a mandulafüvet (Cyperus esculentus L. var.
sativus Boeck.) gazdasági növényként már nagyon régóta hasznosítják. A termesztett mandulafű az arabok közvetítésével került Dél-Európa országaiba. Gumója 20-24% finom étolajat, 3-7% nyers fehérjét és 30% körüli cukrot tartalmaz. Napjainkban az édes-, kozmetikai- és növényolajipar alapanyagként alkalmazza. Belőle készül a spanyolok kedvelt üdítő itala a „Horchata” (Makai és mtsai., 2008).
3.4. Morfológiája
Életformája G2-es, a talajban telelő gumós évelők csoportjába tartozik.
Tarackjai morfológiailag két típusba sorolhatók. Az egyik típus a talaj felszíne alatt vízszintesen nőve képez talajfelszín feletti hajtást. A másik típusa vízszintesen és ferdén lefelé növekedve gumót képez (Dancza és mtsai., 1995) (3. ábra).
3.ábra: A mandulapalka tarackjai
14 A tarackok mérete a 30 cm-t is elérheti. A gumók a tarackok utolsó internódiumaiból alakulnak ki. Alakjuk gömbölyded ill. ovális. Színe kezdetben fehér, majd az érés során sötétbarnává válik. Átmérője 0,3-1 cm (Dancza és mtsai., 1995) (4. ábra).
4. ábra: A mandulapalka gumói
A növény szára háromszögletű, el nem ágazó, magassága 30-80 cm (Gieske et al., 1993;
Dancza és mtsai., 1995; Fertsak et al. 2003).
Virágzata füzéres virágzatrendszer, az alapjánál elágazó. A virágzatban a fellevelek száma 3-6. A füzérek száma 3-8, hosszuk 3-10 cm. A virágok kétivarúak és két sorban ülnek a füzérketengelyen. A pelyvalevél 7-9 erű, szárnyas, a színe az aranysárgától a barnáig változó (Fernald, 1950; Gleason, 1963; Dancza és mtsai., 1995) (5. ábra).
5. ábra: A mandulapalka virágzata
Termése 1-1,5 mm nagyságú, egymagvú termés (Dancza és mtsai., 1995; Fertsak et al. 2003).
A mandulapalka terjedésének felderítését nehezíti, hogy a Cyperaceae család egy másik tagja, a zsióka (Bolboshoenus maritimus) első ránézésre igencsak hasonlít rá (6.,7. ábra).
A zsióka szára, a mandulapalkához hasonló háromszögletű, 30-100 cm magas. A virágzat megjelenésekor már jól megkülönböztethető a két növény, virágzata ernyő vagy fejecskeszerű csomó, színe vöröses vagy sötétbarna, (Simon, 1992).
15
6. ábra: A mandulapalka és a zsióka virágzata 7. ábra: Zsióka virágzat és gyökér (A fotókat Dobszai-Tóth Veronika készítette)
3.5. Szaporodása
Fejlődésmenetére a következő jellemző: az áttelelt gumók tavasszal, április elejétől, 10-12
°C talajhőmérsékletnél kezdenek kihajtani a talaj 5-10 cm-es rétegéből (Stoller et al., 1973, Gieske et al. 1993).
A tömeges kihajtás április végére, május elejére tehető. A felszínre törő hajtások 3-4 leveles állapotban jellemzően sárgás-zöldes színűek áttelelést (Gieske et al., 1993; Dancza és mtsai., 1995; Fertsak et al. 2003).
Az intenzív hajtásnövekedés 5 leveles állapottól indul meg. A tarackok ekkor már gyorsan fejlődnek. Nyár elején gyakran már gumókat is megfigyelhetünk áttelelést (Gieske et al., 1993; Dancza és mtsai., 1995; Fertsak et al. 2003).
Az anyatövek augusztus elejétől virágoznak. 12-14 órás megvilágításnál kezdődik a virágzás (Jansen, 1971). Ekkor a leveles hajtásokat képző tarack a talaj felszíne alatt, vízszintesen, a gumót képző tarackok pedig vízszintesen és ferdén lefelé növekednek. A tarackokból fejlődő hajtások újabb gumókat képeznek. Ezek a gumók szeptemberre fásodva érnek be. Áttelelnek, és tavasszal hajtanak ki áttelelést (Gieske et al., 1993; Dancza és mtsai., 1995; Fertsak et al.
2003).
Egy virágzatból több száz makkocska is képződhet. A termés kivételével fagy hatására a növény minden talajfelszín feletti része elpusztul. Ez, valamint a talajfelszín alatt elhelyezkedő gumók biztosítják az áttelelést (Gieske et al., 1993; Dancza és mtsai., 1995;
Fertsak et al. 2003) (8. ábra).
16 8. ábra: A mandulapalka fejlődésmenete (Dancza és mtsai., 1995)
Az évelő gyomnövényeknek, szemben az egyéves fajokkal, a magvakkal való szaporodás mellett igen hatékony vegetatív szaporodásmódjaik is vannak. Ennek az ivaros szaporodásmóddal szemben számos olyan előnye ismert, amely a fajok megtelepedését és felszaporodását jelentősen segíti. Az ivartalan szaporodás legfontosabb előnye, hogy az utód kezdeti fejlődéséhez több tápanyag áll rendelkezésre. Hátrányai: az utódnövény genetikai alapja teljesen azonos az anyanövénnyel, az utódok kis száma valamint a nagy távolságra való terjesztés hiánya (Hunyadi és mtsai., 2000).
Mandulapalka esetén nem beszélhetünk az utódok kis számából eredő hátrányról, Gieske (1993) leírása szerint egy anyanövény optimális feltételek esetén egy év alatt 1900 újabb növényt és akár 6900 gumót is képezhet. Nem megfelelő gyomirtás mellett négyzetméterenként 1000 db gumó is előfordulhat (Stoller et al., 1979).
A gumók többsége a talaj felső 15 cm-es rétegében fordul elő, de 46 cm-es mélység alatt is találtak gumókat (Tumbelson et al., 1961; Bell et al., 1962; Stoller, 1973). Hazánkban Doma Csaba vizsgálatai alapján a gumók 40 cm mélyre is képesek lejutni, és ezen mélységből tenyészedényes vizsgálatok alapján képesek kihajtani (Doma, 2004).
17 Maggal való szaporodásának is lehet jelentősége. Lapham (1990) a mandulapalka nagy magprodukcióját írja le, Massashusettsben 605 millió/ha, míg Zimbabwében 100 millió/ha.
Az érett magok életképessége 5-40% között volt.
Gieske (1993) által leírt vizsgálatban, a 186 virágzat magprodukcióját vizsgáló felmérésben 647 mag volt a legtöbb amit egy virágzatban találtak.
3.6. Az allelopátia jelensége
A szomszédos magasabb rendű növények egymás növekedésére gyakorolt káros hatását interferenciának nevezzük. Az interferenciában szerepet játszó kapcsolatok a következők lehetnek:
- kompetíció - allelopátia
- közvetlen módok (Mikulás, 1988).
Az allelopátiával a hazai irodalomban először Újvárosi (1973) könyvében találkozhatunk.
Az allelopátia a növények közötti mindennemű serkentő és gátló élettani illetve biokémiai kölcsönhatást takar (Molisch, 1937; Szabó, 1984). Allelopátián általában az átadó (donor) növényeknek a befogadó (akceptor) növényekre gyakorolt serkentő vagy gátló hatásait értjük (Rice, 1974).
Az allelopátiáért felelős anyagok (allelokemikáliák) megtalálhatók a növények gyökérzetében, szárában, leveleiben, a virágokban, a termésekben és a magvakban is (Béres, 2000). Az allelokemikáliák a növényekből több módon szabadulhatnak fel, így párolgással, gyökér általi kiválasztással, kimosódással és a növényi maradványok szétbomlásával. A gátló anyagok hatása sokrétű, hathatnak a tápanyagfelvételre, a sejtosztódásra, a megnyúlásos növekedésre, a fotoszintézisre, a légzésre, a membrán permeabilitására, az enzimaktivitásra (Mikulás, 1988).
A hazai allelopátia kutatás első eredményei Mikulás (1979) nevéhez fűződnek, aki a Sorghum halepense (L.) Pers. allelopatikus hatását vizsgálta. A későbbiekben Béres (1983) az Ambrosia artemisiifolia L., Kovács mtsai. (1988) a Cirsium arvense (L.) Scop., Kazinczi és mtsai. (1991) az Abutilon theophrasti Medic., Hunyadi és mtsai. (1999) a Centaurea cyanus L. allelopátiáját vizsgálták. Az utóbbi években hazai szinten számos egyéb gyomfaj
18 allelopátiája vált ismertté (Béres-Kazinczi 2000, Kazinczi és mtsai. 2004, 2009, Csontos 1997, Hódi-Gazdagné 1999, Dávid 2003).
A hazai flórát vizsgálva Szabó (1997) 150 allelopátiás hajlamú fajt említ, melyek többsége táj- és flóraidegen, meghonosított haszonnövény, de jelentős csoportot alkotnak még a zavarástűrő fajok és az agresszív kompetítorok is.
A mandulapalka allelopátiáját Dirk et al. (1980) vizsgálta. Növénykivonatok és növénymaradványok hatását üvegházi körülmények között vizsgálva megállapították, hogy allelopatikus hatással van a szójára és a kukoricára. Az allelopátiáját kiváltó vegyület valószínűleg azonos azzal a fenolos vegyülettel, amit a gumóiból ki is mutattak (Jangaard et al., 1971; Tames et al., 1973) .
3.7. Kompetíció
A kompetíció meghatározására a szakirodalom leggyakrabban Donald (1963) definícióját idézi: „A kompetícióban két vagy több szervezet mindegyike keresi a maga sajátos szükségleteinek kielégítését valamilyen adott tényezőből, amikor a közvetlen ellátás ebből a tényezőből alatta marad a szervezetek együttes igényének”.
A kompetíció lehet fajok közötti (interspecifikus), fajon belüli (intraspecifikus), genotípusok közötti (intergenotypic), genotípuson belüli (intragenotypic) és növényen belüli (intraindividuális).
A növények közötti kompetíció erősségét befolyásolja kezdetének időpontja, a versengés időtartama és a növények fenológiai állapota annak kezdetekor (Chancellor-Peters, 1974).
Azok a gyomnövények, amelyek egy időben csíráznak a kultúrnövénnyel, általában jobb kompetítorok, mint a később csírázó egyedek (Niemann, 1977; Peters-Wilson, 1983).
A kultúrnövények bizonyos ideig képesek tolerálni a gyomnövények jelenlétét, amennyiben ezt követően gyommentes periódust tudunk biztosítani a kultúrnövény számára, akkor már képes elnyomni a később fejlődő gyomnövényeket, ezáltal elkerülhető a gyomnövény – kultúrnövény kompetícióból adódó termésveszteség (Kazinczi, 2000). A tolerálható gyomversengés hossza és a szükséges gyommentes periódus hossza közötti időtartamot nevezzük kritikus kompetíciós periódusnak (Nieto et al., 1968).
A természetben a növények leggyakrabban három fő tényezőért: a tápanyagokért, a vízért és a fényért versengenek egymással (Hunyadi, 1993).
19 Heterogén populációk kompetíciójának tanulmányozására két módszert használhatunk:
- helyettesítési kísérlet: ezekben a kísérletekben együtt kerül vetésre a két (vagy több) növényfaj, különböző arányokban keverve, a kevert állomány sűrűségét állandó értéken tartva. Ezen kísérletekben adott faj tiszta állományban tanúsított viselkedése összehasonlítható a változó arányban kevert gyomállományéval és mérhető a kölcsönös agresszivitás.
- additív kísérlet: két növényfajt nevelnek együtt, az egyik faj sűrűsége állandó, a másikét változtatják. A különböző gyomfajok egymáshoz viszonyított kompetíciós képessége mérhető (Hunyadi és mtsai., 2000).
A mandulapalka kompetícióját vizsgálva Hauser et al. (1974) megállapította, hogy 0-66%
termésveszteséget okoz a gyomnövény jelenléte olyan kukoricatáblán, ahol nem irtották a mandulapalkát.
Stoller et al. (1979) megfigyelései szerint négyzetméterenként minden 100 mandulapalka egyed 8% termésveszteséget okozott a kukoricánál.
Hoffmanné (2004) 30-40%-os termésveszteségről számol be erős mandulapalka fertőzés esetén.
3.8. A tápanyagokért történő versengés
Hazánkban a gyomnövények ásványielem-felvételét és a műtrágyák hatását a gyomnövény – kultúrnövény kompetícióra többen is vizsgálták (Tölgyesi 1969, Debreczeniné 1988, Lehoczky 1988, Radics 1990, Lukács és mtsai 1998).
Alkämper (1976) leírja, hogy a gyomnövények kezdeti kompetíciós előnyt szereznek azáltal, hogy az ásványi elemeket általában hamarabb és nagyobb mennyiségben veszik fel, mint a velük társulásban élő kultúrnövények.
A makroelemek közül különösen a nitrogénért történő versengés erős (Thrasher et al., 1962).
A jó nitrogén- ellátottság kedvező hatással van egyes gyomfajok kompetíciós képességére (Wells 1979, Dryden 1974). Az elmúlt évek nagyadagú nitrogénműtrágyázásával olyan nitrofil gyomok szaporodtak fel, melyek ma az őszi búza nehezen irtható gyomnövényei közé tartoznak (Tóth és mtsai, 1989).
A káliumért történő versengés erős kompetítoraira a nagymértékű kálium akkumuláció utal (Kazinczi és mtsai, 1998; Qasem-Hill, 1995).
20 A foszforért folytatott versengés intenzitása elmarad a másik két elemétől. A pillangósvirágúak és az egyszikű gyomnövények versengése lehet jelentős (Keeley, 1965;
Vengris et al., 1953).
A gyomnövényfajok tápelemtartalmának és a tápanyagfelvétel dinamikájának ismerete szükséges az egyes fajok tápanyagversengésben betöltött szerepének megítéléséhez. A szakirodalomban a gyomnövények kémiai összetételének vizsgálatáról először Kling (1914) munkájában olvashatunk. Korsmo (1930) könyvében 18 gyomnövényfaj tápanyagtartalmáról közöl adatokat. Singh-Singh (1939) 48 faj makroelem- tartalmát, réz- és kéntartalmát vizsgálta. Tölgyesi (1969) könyvében az egyes növénycsaládok tápelem- összetétel szerinti jellemzésére is találhatunk adatokat. Debreczeniné (1988) a gyomfajok makro- és mikroelem- tartalmáról közöl adatokat. Lehoczky (1994) 54 szántóföldi gyomnövényfaj N-, P-, K-, Ca- koncentrációját ismerteti.
3.9. Védekezési lehetőségek a mandulapalka ellen
Agrotechnikai védekezés
A mandulapalka ellen a leghatékonyabb és egyben legolcsóbb védekezés a betelepedés megakadályozása. A terjedés megelőzésének céljából fontos a gumók, valamint a szennyezett talajművelő eszközzel történő terjesztésének megakadályozása (Dancza, 2005).
A fertőzött területeken az őszi mélyszántást úgy célszerű elvégezni, hogy a gumók a talajfelszínre kerüljenek, mert ott a téli fagyok elpusztítják azokat, illetve a tavaszi talajmunkák során a kihajtott gyomnövények elpusztíthatók (Gieske et al., 1993).
Fertsak (2003) a mandulapalka elleni védekezési javaslatok között megemlít egy spanyolországi módszert, mely során a területet egy évig ugaron hagyják. Megvárják, amíg a mandulapalka növények elérik a 15-30 cm-es magasságot, ekkor lekaszálják, ezután burgonya betakarító géppel a talajfelszínre forgatják a gumókat és hagyják a napon megszáradni, végül összegyűjtik azokat és elégetik.
21 Vetésforgó
A mandulapalkára jellemző, hogy az árnyékolást nem kedveli, ezért visszaszorítására célszerű olyan növényeket illeszteni a vetésforgóba, amelyek tavasszal gyorsan záródó kultúrát létrehozva árnyékolják a talajt (Gieske et al., 1993).
Hollandiában 1984-től 1992-ig érvényben volt egy jogszabály, amely a mandulapalkával fertőzött területeken tiltotta a gyökgumósok (burgonya, cukorrépa) termesztését (Dancza, 2005).
Vegyszeres gyomirtás
A védekezési eljárások kidolgozása során meghatározó a mandulapalka biológiai sajátosságai közül az igen intenzív vegetatív fejlődési ciklus, valamint a vékony tarackok képződése. A mandulapalka biológiáját ismerve a kémiai védekezés során cél a mandulapalka kártételének minimalizálása elsősorban a gumóprodukció redukálásával. A mandulapalka életmódja miatt elsődleges, hogy a hatóanyagot a gumókhoz juttassuk. Viszont a gumók Doma (2004) megfigyelései szerint 40 cm mélyre is képesek lejutni, majd kihajtani. Ezért egy védekezés nem elégséges ellene. Vegyszeres védekezés során javasolt a glifozát hatóanyag, mely elpusztítja a gumót, és a diquat-dibromid hatóanyag, mely csak leperzseli a növényeket, ez esetben a leperzselt gumók újrahajthatnak. Csapadékszegény időjárás mellett a pre-sowing kezelések hatékonyabbak, mert a preemergens kezelések esetében nagyobb mennyiségű bemosó csapadékra van szükség ahhoz, hogy a hatóanyag a gumókhoz jusson. A mandulapalka ellen, mint ahogy a többi évelő gyomnövény ellen csak komplexen lehet védekezni. Az alapkezeléseket állománykezelésnek kell követniük (Hoffmanné, 2004).
Irodalmi adatok alapján a mandulapalka ellen használható hatóanyagok a következők:
B-csoport: ALS-gátlók
Szulfonil-karbamidok: rimszulfuron, nikoszulfuron, tifenszulfuron-metil (Ackley 1996, Hoffmanné 2004);
Imidazolin: imazaquin (Richburg 1993, Ackley 1996), imazetapir (Ackley 1996),
Flumetszulam, foramszulfuron + jodoszulfuron, tritoszulfuron + dikamba (Hoffmanné, 2004).
22 C-csoport: Fotoszintézis- gátlás a PS II. rendszerben
1,3,5-triazin: atrazin;
Karbamid: linuron;
Benzotiadiazin: bentazon (Pereira 1986/a, Stoller 1979, Hoffmanné 2004), bentazon + dikamba (Hoffmanné, 2004).
E-csoport: PROTOX gátlók
Difeniléter: oxifluorfen (Pereira, 1986/b)
F3-csoport: Plasztokinon- bioszintézist gátlók
mezotrion, mezotrion + s-metolaklór + terbutilazin (Hoffmanné, 2004)
G-csoport: EPSP szintetáz gátlók Glicin: glifozát (Pereira 1986/a)
K3-csoport: Fehérje- és nukleinsav- szintézist gátlók
Klóracetamid: acetoklór (Hoffmanné, 2004), alaklór (Dixon, 1980; Hoffmanné, 2004), dimetenamid (Hoffmanné, 2004), s-metolaklór (Obrigawitch, 1980; Dixon, 1980; Banks, 1983; Hoffmanné, 2004), propaklór (Banks, 1983), propizoklór (Hoffmanné, 2004).
N-csoport: Lipid- bioszintézis gátlás
Tiokarbamát: EPTC (Pereira, 1986/a; Banks, 1983; Stoller, 1979), butilát (Hoffmanné, 2004).
O-csoport: Hormonrendszerre ható
Ariloxi-karbonsav: 2,4-D (Pereira, 1986/a), floraszulam + 2,4-D (Hoffmanné, 2004).
Ezen herbicid csoportok közül szisztemikusak, tehát a növényben mozogva a gumókhoz is eljutók a B-csoportba (ALS-gátlók), a G-csoportba (EPSP szintetáz- gátlók) és az O- csoportba (Hormonrendszerre ható) tartozó hebicidek.
A fotoszintézis működését zavarók a C-csoportba (Fotoszintézis- gátlás a PS II. rendszerben) és az F3-csoportba (Plasztokinon- bioszintézis gátlók) tartozó herbicidek, hatásukra a levelek elszáradnak, elhalnak. Az E-csoportba (PROTOX gátlók) tartozó herbicidek leperzselik azokat a növényi részeket, melyekre rákerülnek. A K3-csoportba (Fehérje- és nukleinsav-
23 szintézist gátlók) tartozó herbicidek csak a kelő magvakat károsítják, csíranövény- pusztulást okoznak. Az N-csoportba (Lipid- bioszintézis gátlás) tartozó herbicidek a telítetlen zsírsavak bioszintézisét gátolják. Ezen herbicidek használata a gumókat nem károsítja, azok újrahajthatnak. (Kádár, 2010).
24
4. ANYAG ÉS MÓDSZER
4.1. LABORATÓRIUMI KÍSÉRLETEK
4.1.1. Maghozamvizsgálat
Somogytúr határában, 2006-ban gyűjtött virágzatok maghozamát vizsgáltuk. A virágzatokból kézzel eltávolítottuk a magokat, majd megszámoltuk. 100 db virágzatot vizsgáltunk meg.
4.1.2. Csírázásbiológiai vizsgálatok
2005. szeptember 14-én Somogytúr határában gyűjtöttük a virágzatokat, melyekből az érett magokat kézzel eltávolítottuk, majd szobahőmérsékleten, papírzacskóban tároltuk. 2005.
október 4. és 2006. október 11. között, 2 hetente 100 db mag felhasználásával, 4 ismétlésben, Petri-csészében, nedves szűrőpapíron vizsgáltuk a mandulapalka magjainak csírázását. A csíráztatás szobahőmérsékleten, természetes fényben zajlott. A szűrőpapírokat nem hagytuk kiszáradni. A magokat 2 hétig figyeltük, majd feljegyeztük a csírázott magvak számát. A csírázás kezdetét a gyököcske megjelenése jelentette.
4.1.3. A mandulapalka allelopátiája
A mandulapalka hajtásából és föld alatti részeiből (gyökér, tarack, gumó) készített növénykivonatok hatását vizsgáltuk a tesztnövények csírázására laboratóriumi körülmények között. Tesztnövényként kukoricát (Zea mays L.), napraforgót (Helianthus annuus L.), fehér mustárt (Sinapis alba L.), cukorrépát (Beta vulgaris L. convar. Crassa Alef. provar. altissima Döll.) és parlagfüvet (Ambrosia artemisiifolia L.) használtunk. A kísérletben használt mandulapalka növények Somogytúr határában kerültek begyűjtésre. A növénykivonat készítése során 12,5 g Cyperus esculentus hajtást vagy föld alatti részt 100 ml vízzel összeturmixoltunk, 1 napig állni hagytuk, majd leszűrtük és hűtőben tároltuk. A növénykivonatok készítésének módszerét szakirodalmi adatok felhasználásával dolgoztuk ki (Dirk et al., 1980; Torma és mtsai., 2004; Kazinczi és mtsai., 2009).
25 A növénykivonatokat 1, 2, 5, és 10-szeres hígításban alkalmaztuk. . A Petri-csészékbe a kukorica és a napraforgó terméséből 25 db-ot, a fehér mustár és a parlagfű magjából valamint a cukorrépa terméséből 50 db-ot helyeztünk szűrőpapírra 3 ismétlésben. A Petri-csészéket szobahőmérsékleten helyeztük el. 11 nappal később megszámoltuk a kicsírázott magokat. A kísérletet 2007. október 3-án állítottuk be.
4.2. ÜVEGHÁZI TENYÉSZEDÉNYES KÍSÉRLETEK
4.2.1. Allelopátiás vizsgálatok
4.2.1.1. A mandulapalka hajtás- és föld alatti rész maradványának hatása a tesztnövények csírázására és fejlődésére
A kísérletben használt mandulapalka (donor) növények Somogytúr határában kerültek begyűjtésre. Az előzőleg kiszárított növényi maradványokat a tenyészedény tápközegébe kevertük június közepén (800g növényi maradvány/10kg földkeverék). A kísérlet módszerét szakirodalmi adatok felhasználásával dolgoztuk ki (Kazinczi és mtsai., 2009).
Tápközegként virágföld-homok 2/3-1/3 arányú keverékét alkalmaztuk. Tesztnövényként a laboratóriumi kísérletben is alkalmazott kukoricát, napraforgót, fehér mustárt, cukorrépát és parlagfüvet használtuk. A kísérletet 2006. szeptember 19-én állítottuk be üvegházban.
Tenyészedényenként 50-50db termést ill. magot vetettünk, 0,5kg-os tenyészedényeket használtunk. Minden kezelésben négy ismétlést alkalmaztunk. A kelést egy hét múlva értékeltük. A kelési % értékelése után tőszám- beállítást végeztünk úgy, hogy minden tenyészedényben 4 db növényt hagytunk meg további vizsgálatra. A később kelt növényeket feljegyeztük és eltávolítottuk. A tesztnövényeket 5 hétig neveltük, majd megmértük a hajtások friss- és száraztömegét.
4.2.1.2. A mandulapalka hajtásából és föld alatti részekből készített növénykivonat hatása a tesztnövények csírázására és fejlődésére
A mandulapalka növényeket Somogytúr határában gyűjtöttük. A növénykivonat készítése során 12,5g mandulapalka hajtást vagy föld alatti részt 100ml vízzel összeturmixoltunk, 1 napig állni hagytuk, majd leszűrtük és hűtőben tároltuk. A növénykivonatokat két módszerrel
26 juttattuk ki. Az egyikben a növénykivonatot a tesztnövényekre permeteztük naponta kézi permetezővel, míg az öntözés vízzel történt. A kijuttatott dózis naponta 10ml növénykivonat tenyészedényenként.
A másikban a növénykivonatokkal öntöztük a táptalajt, ezek a növények öntözővizet nem kaptak. A kijuttatott dózis naponta 30ml növénykivonat tenyészedényenként.
Tesztnövényként kukoricát, napraforgót, fehér mustárt, cukorrépát és parlagfüvet használtunk.
A kísérletet 2007. augusztus 14-én állítottuk be, 4 ismétlésben. A tápközeg virágföld-homok 2/3-1/3 arányú keveréke volt. Tenyészedényenként 50-50db termést ill. magot vetettünk, 0,5 kg-os tenyészedényeket használtunk. A kicsírázott növényeket augusztus 19-én megszámoltuk, majd tőszámbeállítást végeztünk, minden tenyészedényben 4db növényt hagytunk meg további vizsgálatra. A később csírázott növényeket feljegyeztük és eltávolítottuk. A növényeket 5 hétig neveltük, majd megmértük a friss- és a száraztömeget.
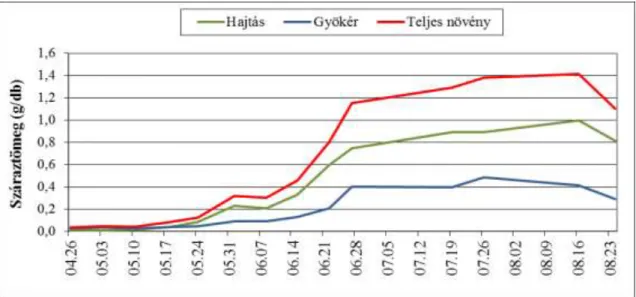
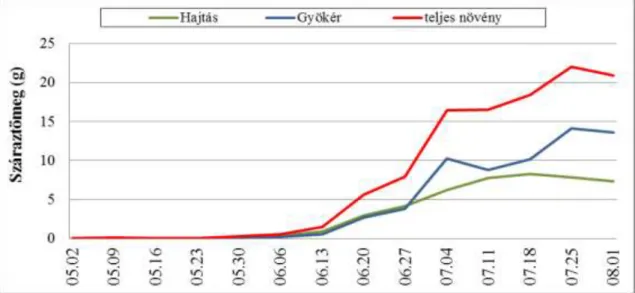
4.2.2. A mandulapalka fejlődésének vizsgálata növekedésanalízis módszerével
A mandulapalka fejlődésének vizsgálatához talajkultúrás kísérletet állítottunk be 2007.
április 19-én, 5 ismétlésben. A növényeket dézsákban neveltük, a dézsákat a szabadban helyeztük el, tápközegként virágföld : homokos vályog talaj 1:1 arányú keverékét használtuk.
Dézsánként 25db gumót ültettünk el, majd a virágzás végéig hetente dézsánként 1db növényt és szaporulatát vettük ki. A növényeket igény szerint öntöztük. A tenyészidőszakban összesen 16 alkalommal történt mintavétel, 2007. április 26. és augusztus 24. között.
A kísérletet 2008-ban megismételtük. Üvegházban, tenyészedényben neveltük a növényeket, 5 ismétlésben. Tenyészedényenként 1db növényt és szaporulatát neveltük. Tápközegként virágföld : homokos vályog talaj 1/3 : 2/3 arányú keverékét használtuk. A növényeket igény szerint öntöztük. A kísérletet április 25-én állítottuk be, a tenyészidőszakban 13 alkalommal történt mintavétel, 2008. május 2. és augusztus 1. között. Hetente 5db növényt és szaporulatát távolítottuk el.
Minden értékelésnél mértük a levélterületet (LI-3000-es levélterület mérővel), a hajtás és földalatti részek (gyökér, gumó, tarack) hosszúságát (cm), frisstömegét (g), majd szárítás után (40°C –ra beállított szárítószekrényben) a száraztömegét (g). A vizsgálatok során az anyanövényt és szaporulatát egy egységként kezeltük. A növekedésanalízis vizsgálatánál a mandulapalka száraztömegben kifejezett növekedését vizsgáltuk a növények tömegét és
27 asszimiláló rendszerének nagyságát mérve. A nyert adatokból – a Virágh (1980) által értelmezett növekedési mutatók alapján - a következő növekedési indexeket számítottuk:
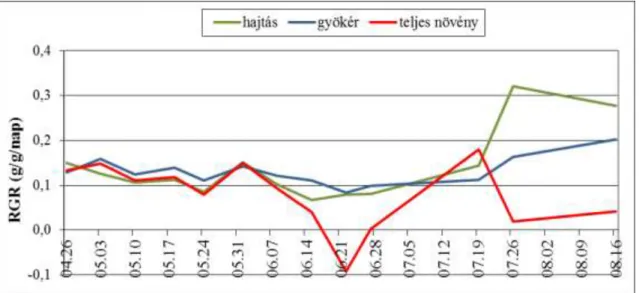
- RGR (relative growth rate), relatív növekedési sebesség. Kifejezi a primer szervesanyag- tartalom időbeni felhalmozódásának sebességét (arányát), a növekedés ütemét. Egységnyi tömegre vonatkoztatott tömeggyarapodás egységnyi idő alatt:
RGR = ln (W2 / W1) (g/g/nap) t2 – t1
- RLGR (relative leaf growth rate), relatív levélterület növekedési arány. Növényenkénti összes levélterület növekedésének rátáját akkor kapjuk meg, ha az RGR meghatározásánál alkalmazott képletben száraztömeg helyett levélterület (A) adatokkal számolunk:
RLGR = ln (A2 / A1) (cm2/cm2/nap) t2 – t1
- LAR (leaf area ratio), levélterület – arány. Az asszimiláló rendszer nagyságának, a fotoszintetikus kapacitás mértékének mutatója, a növény levélborítottságának morfológiai jellemzője. A teljes asszimiláló terület és a növény teljes tömegének hányadosaként értelmezett index:
LAR = A1 + A2
(cm2/g)
W 1 + W
2
- LAI levélterület index: A növényállomány egységnyi területére jutó levélterület. A növényenkénti levélterület és a tenyészterület közötti arány, levélterület egységnyi tenyészterületre vetítve:
LAI = LA (cm2/ cm2)
P
- SLA specifikus levélterület arány. Azt mutatja, milyen levélstruktúra képződik a rendelkezésre álló szárazanyagból – egy nagy specifikus levélterület jelzi a vékony leveleknek viszonylag nagy területét. A növény összes levélterülete és az összes levél száraztömege közötti arány:
SLA = LA (cm2/g)
LW
28 4.2.3. A mandulapalka tápelem - koncentrációjának vizsgálata
A tápelemtartalom meghatározásához a 2007-es növekedésanalízis- vizsgálat során gyűjtött mintákat használtuk fel. A mintákat szárítószekrényben, 40°C-on súlyállandóságig szárítottuk, majd papírzacskóba helyeztük a felhasználásig. A tárolás szobahőmérsékleten történt. A tápelemtartalom meghatározását a Növényvédelmi Intézet Herbológiai és Növényvédőszer Kémiai Osztály laboratóriumában végeztük el, az Agrokémiai gyakorlatok módszertana szerint (Debreczeni B.-né, 1986). Az összes nitrogéntartalom meghatározására Kjeldahl módszerrel Parnass-Wagner vízgózdesztillációt alkalmaztunk. Az összes foszfortartalom meghatározására (SPEKOL-1l) spektrofotométert alkalmaztunk. Az összes kálium és kalciumtartalom meghatározása lángfotométerrel (FLAPHO-4) történt.
4.2.4. Korai kompetíciós vizsgálat
A vizsgálatok során helyettesítési módszert alkalmaztunk, kidolgozása De Wit (1960) nevéhez fűződik. Ezekben a kísérletekben együtt kerül vetésre a két vizsgálandó növény, különböző arányokban keverve, az állomány sűrűségét állandó értéken tartva. Ezáltal adott faj tiszta állományban tanúsított viselkedése összehasonlítható a változó arányban kevert állományéval és mérhető a kölcsönös agresszivitás. Továbbá figyelhető az inter- illetve az intraspecifikus kompetíció (Hunyadi és mtsai, 2000).
Azt vizsgáltuk, hogy a napraforgó és a mandulapalka, valamint a kukorica és a mandulapalka különböző arányokban elvetve hogyan hat egymás fejlődésére a tenyészidőszak első 4 hetében. A növényszám alakulás a következő volt:
5db napraforgó (tiszta állomány), 5db mandulapalka (tiszta állomány), 4db napraforgó + 1db mandulapalka, 3db napraforgó + 2db mandulapalka, 2db napraforgó + 3db mandulapalka, 1db napraforgó + 4db mandulapalka.
5db kukorica (tiszta állomány), 5db mandulapalka (tiszta állomány), 4db kukorica + 1db mandulapalka, 3db kukorica + 2db mandulapalka, 2db kukorica + 3db mandulapalka, 1db kukorica + 4db mandulapalka (9. ábra).
29 Kezelések (növény arányok) és rövidített jelölésük a későbbi ábrákon
5db kukorica 5K 5db napraforgó 5N
5db mandulapalka 5M 5db mandulapalka 5M
4db kukorica – 1db mandulapalka 4K-1M 4db napraforgó – 1db mandulapalka 4N -1M 3db kukorica – 2db mandulapalka 3K-2M 3db napraforgó – 2db mandulapalka 3N -2M 2db kukorica – 3db mandulapalka 2K-3M 2db napraforgó – 3db mandulapalka 2N -3M 1db kukorica – 4db mandulapalka 1K-4M 1db napraforgó – 4db mandulapalka 1N -4M
9. ábra: Alkalmazott kezelések
Mivel a növényeket öntöztük, a fényért és a tápanyagért folytatott versengés hatásait tudtuk vizsgálni. Néztük a hajtások és a gyökerek közötti kompetíciót.
Üvegházi tenyészedényes kísérletet állítottunk be, 2006 júniusában. Tenyészedényenként 1kg homokos-vályog talajt használtunk. A talajt Sávoly község határában gyűjtöttük (10. ábra).
Minden tenyészedényben 5 darab növényt neveltünk különböző kombinációkban. A napraforgó és a kukorica 95%-os csírázási aránya miatt mindenhol a szükségesnél eggyel több magot vetettünk, majd a kelés utáni napokban tőszámbeállítás történt. A mandulapalka- gumókat előhajtattuk, bizonytalan hajtási százaléka miatt. Az öntözés naponta igény szerint és hetente egyszer súlyra történt. A növényeket 4 hétig neveltük. Vizsgáltuk a növények hajtásának és gyökerének friss- és száraztömegét, a hajtáshosszúságot és a levélszámot.
A kísérletben használt napraforgó: Tellia RM. Korai érésű, középmagas-magas, erős, stabil szárú, igen nagy tányérral rendelkező hibrid. Állami elismerés éve: EU-fajtalista.
A kísérletben használt kukorica hibrid: KWS 328. Zöld száron érő lófogú szemeskukorica, jellemző rá a kiváló stressztűrés, átlag feletti szárszilárdság. Állami elismerés éve: 2003.
A kísérletben használt mandulapalka- gumók Somogytúr határában kerültek begyűjtésre, majd tisztítás után Petri-csészében, nedves szűrőpapíron lettek hajtatva.
Humusz % 2,47
pH H2O 6,27
Ásványi N mg/kg talaj 16,91 mg/kg talaj AL-oldható P2O5 137 mg/kg talaj AL-oldható K2O 346 mg/kg talaj
10. ábra: A kísérleti talaj tulajdonságai
30 Ezen vizsgálatok növénymintáit felhasználva megnéztük a kukorica- és a mandulapalka- hajtások és a gyökerek közötti tápanyagokért folytatott versengést is. Laboratóriumban vizsgáltuk a levelek és a gyökerek százalékos N-, P2O5- és K2O- tartalmát. A kevert állományokat a tiszta állományokhoz hasonlítottuk az értékelés során.
4.3. EREDMÉNYEK STATISZTIKAI ELEMZÉSE
A kísérleti adatokat ANOVA SPSS statisztikai programcsomag segítségével értékeltük, egytényezős varianciaanalízist (szignifikáns differencia számítás) végezve.
31
5. EREDMÉNYEK ÉS KÖVETKEZTETÉSEK
5.1. LABORATÓRIUMI KÍSÉRLETEK
5.1.1. Maghozam vizsgálat
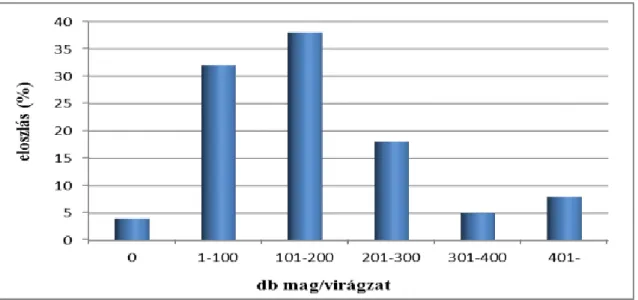
100db virágzat maghozamát vizsgáltuk meg. A vizsgált virágzatok 4%-a nem tartalmazott magot. A legnagyobb arányban (38%) 101 és 200 db között változott a magszám. Az átlag magszám 155 db volt. A legtöbb mag, amit találtunk 466 db volt (11. ábra).
Gieske et al. (1993) cikkében beszámol egy kísérletről melyben 186 virágzatot vizsgáltak, leírja, hogy nem minden virágzatban találtak magot. Vizsgálatukban 647 db mag volt a legtöbb, amit találtak.
11. ábra: A mandulapalka maghozamának vizsgálata 2006-ban
5.1.2. Éves csírázási ritmus vizsgálat
A gyűjtött magok életképesség- vizsgálatát az MGSZH Magkórtani Laboratóriuma végezte el, TTC teszttel, az eredmény 56% volt.
A mandulapalka magok március végén kezdtek el csírázni. Márciustól június végéig kiegyenlített, 47-64%-os csírázási átlagokat számoltunk. Júliusban a csírázási százalék kezdett lecsökkenni és októberben megközelítette a minimumot (12. ábra).
32 12. ábra: A mandulapalka magok éves csírázási ritmusa laboratóriumi körülmények között
5.1.3. A mandulapalka- hajtásából és föld alatti részekből készített növénykivonat hatása a tesztnövények csírázására laboratóriumi kísérletben
A napraforgó reagált a legérzékenyebben a mandulapalka hajtás- és föld alatti rész- kivonatokra. A növénykivonatok valamennyi koncentrációban csökkentették a csírázását, szignifikáns volt a csökkenés a hajtáskivonat egy és kétszeres hígításában (13. ábra). Hasonló hatást írtak le korábban az Asclepias syriaca levél kivonatáról (Kazinczi és mtsai. 2004).
0 1020 30 40 5060 70 8090 100
Csírázási %
napraforgó
13. ábra: A mandulapalka hajtás- és föld alatti részek kivonatának hatása a napraforgó csírázására laboratóriumi kísérletben (kontroll = kezeletlen, 1X = egyszeres hígítás, 2X = kétszeres hígítás, 5X = ötszörös hígítás, 10X = tízszeres hígítás)
SzD5%=18,2
33 A mandulapalka- hajtás kivonat egy-, két- és ötszörös hígításban csökkentette a kukorica csírázását (14. ábra). Hasonló hatást figyeltek meg az Asclepias syriaca levél kivonatával végzett kísérletben (Kazinczi és mtsai., 2004), valamint a Cirsium arvense növénykivonatokkal végzett kísérletben (Kovács és mtsai., 1988; Béres és Csorba, 1992). A mandulapalka föld alatti rész kivonata két- és ötszörös hígításban növelte a kukorica csírázását (10. ábra). A Cirsium arvense gyökérkivonataival végzett kísérletben hasonló hatást figyeltek meg (Kazinczi és mtsai., 2004).
0 10 20 30 40 50 60 70 80 90 100
Csírázási %
kukorica
14. ábra: A mandulapalka hajtás- és föld alatti részek kivonatának hatása a kukorica csírázására laboratóriumi kísérletben (kontroll = kezeletlen, 1X = egyszeres hígítás, 2X = kétszeres hígítás, 5X = ötszörös hígítás, 10X = tízszeres hígítás)
SzD5%=5,5
34 A parlagfű csírázását a növénykivonatok valamennyi koncentrációban növelték (15. ábra).
0 10 20 30 40 50 60 70 80 90 100
Csírázási %
parlagfű
15. ábra: A mandulapalka hajtás- és föld alatti részek kivonatának hatása a parlagfű csírázására laboratóriumi kísérletben (kontroll = kezeletlen, 1X = egyszeres hígítás, 2X = kétszeres hígítás, 5X = ötszörös hígítás, 10X = tízszeres hígítás)
A mandulapalka hajtás és föld alatti rész kivonatok minimálisan befolyásolták a fehér mustár csírázását. Szignifikáns eltérést a föld alatti rész kivonatának egyszeres hígítása okozott (16.
ábra).
0 10 20 30 40 50 60 70 80 90 100
Csírázási %
fehér mustár
16. ábra: A mandulapalka hajtás- és föld alatti részek kivonatának hatása a parlagfű csírázására laboratóriumi kísérletben (kontroll = kezeletlen, 1X = egyszeres hígítás, 2X = kétszeres hígítás, 5X = ötszörös hígítás, 10X = tízszeres hígítás)
SzD5%=4,4 SzD5%=5,2
35 A cukorrépa csírázását mind a hajtás-, mind a föld alatti rész- kivonatos kezelések csökkentették, szignifikáns eltérés a hajtáskivonat kétszeres hígításánál volt (17. ábra).
0 10 20 30 40 50 60 70 80 90 100
Csírázási %
cukorrépa
17. ábra: A mandulapalka hajtás- és föld alatti részek kivonatának hatása a parlagfű csírázására laboratóriumi kísérletben (kontroll = kezeletlen, 1X = egyszeres hígítás, 2X = kétszeres hígítás, 5X = ötszörös hígítás, 10X = tízszeres hígítás)
SzD5%=10,6
36 5.2. ÜVEGHÁZI TENYÉSZEDÉNYES KÍSÉRLETEK
5.2.1. Allelopátia vizsgálatok
5.2.1.1. A mandulapalka hajtás- és föld alatti részek maradványának hatása a tesztnövények csírázására és fejlődésére üvegházi kísérletben
Frisstömeg
A mandulapalka- hajtást tartalmazó tápközegben nevelt növények frisstömege minden esetben növekedett a kontrollhoz képest. A parlagfű frisstömege 1,5-szerese, míg a kukorica, a napraforgó és a mustár frisstömege 2-szerese volt a kontrollnövények értékének, ezek az eltérések szignifikánsak voltak.
A mandulapalka föld alatti rész- maradványokat tartalmazó tápközegben nevelt növények frisstömege kisebb volt, mint a kontrollnövényeké. A kukorica 38%-kal, a napraforgó 35%- kal, a mustár 68%-kal volt kisebb tömegű a kontrollhoz képest, ezek az eltérések szignifikánsak voltak (18. ábra)
18. ábra: A mandulapalka hajtás- és föld alatti részek maradványának hatása a tesztnövények frisstömegére, üvegházi tenyészedényes vizsgálatban
SzD5%= 1,66 SzD5%= 0,7 SzD5%= 0,19 SzD5%= 1,51 SzD5%= 2,02
37 Száraztömeg
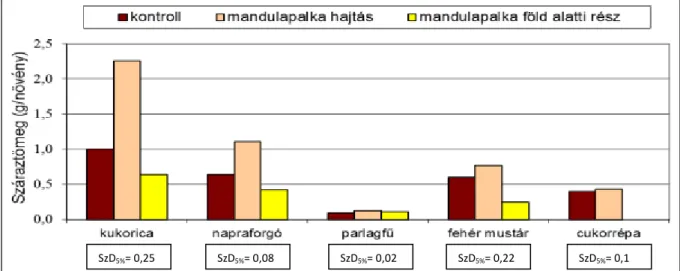
A mandulapalka- hajtást tartalmazó tápközegben nevelt növények száraztömege a frisstömeghez hasonlóan minden esetben nagyobb volt a kontrollhoz képest. A parlagfű és a napraforgó száraztömege 1,5-szerese, a kukorica száraztömege 2-szerese volt a kontrollnövények értékének, a növekedések szignifikáns volt.
A mandulapalka föld alatti rész- maradványokat tartalmazó tápközegben nevelt növények száraztömege kisebb volt a kontrollnál. A kukorica 37%-kal, a napraforgó 33%-kal, a mustár 67%-kal volt kisebb száraztömegű, az eltérések szignifikánsak voltak (19. ábra).
19. ábra: A mandulapalka hajtás- és föld alatti részek maradványának hatása a tesztnövények száraztömegére, üvegházi tenyészedényes vizsgálatban
Kelési %
A mandulapalka- hajtást és föld alatti részeket tartalmazó tápközegben nevelt növények kelési adatai nem tértek el szignifikánsan a kontroll növényektől (20. ábra).
SzD5%= 0,25 SzD5%= 0,08 SzD5%= 0,02 SzD5%= 0,22 SzD5%= 0,1