1
2
UNIVERSITY OF PANNONIA GEORGIKON FACULTY
DOCTORAL SCHOOL OF ANIMAL AND AGRICULTURAL ENVIRONMENTAL SCIENCES
HEAD OF THE DOCTORAL SCHOOL PROF. DR. ANGÉLA ANDA, D. Sc.
STUDIES ON FISH PARASITIC MYXOZOANS IN LAKE BALATON, HUNGARY AND IN FRESHWATER AND MARINE BIOTOPES IN MALAYSIA
DOCTOR OF PHILOSOPHY (PhD) THESIS WRITTEN BY
MUHAMMAD HAFIZ BORKHANUDDIN
SUPERVISOR
CSABA SZÉKELY, HONORARY PROF., PhD
KESZTHELY, HUNGARY 2013
3
STUDIES ON FISH PARASITIC MYXOZOANS IN LAKE BALATON, HUNGARY AND IN FRESHWATER AND MARINE BIOTOPES IN MALAYSIA
Written By
MUHAMMAD HAFIZ BORKHANUDDIN
Written in the University of Pannonia, Doctoral School of Animal and Agricultural Environmental Sciences
Supervisor: Prof. Dr. Csaba Székely I propose for acceptance (yes /no)
Signature
The candidate has achieved………..% at the comprehensive exam, I propose the thesis for acceptance as the reviewer:
Name of reviewer: ……….yes /no
Name of reviewer: ………..yes /no
The candidate has achieved………..% at the public discussion.
Veszprém/Keszthely,
….………..
Chairman of the Committee Labeling of the PhD diploma ………..
………
President of the UCDH
4
CONTENTS
TITLE PAGE 2
APPROVAL FORM 3
CONTENTS 4
LIST OF TABLES 7
LIST OF FIGURES 7
ABSTRACT 10
KIVONAT 11
ABSTRAK 12
CHAPTER I INTRODUCTION 13
CHAPTER II LITERATURE REVIEW 17
2.1 Phylum Myxozoa 17 2.2 Class Myxosporea 18
2.3 Myxozoan Life Cycle 19
2.3.1 Myxospores and Vertebrate Hosts 21
2.3.2 Actinospores and Invertebrate Hosts 21
2.4 Myxozoan Studies in Hungary 22
2.5 Myxozoan Studies in Malaysia 24
2.6 Myxospores Characterization Methods 31
2.6.1 Morphological Examination 31
2.6.2 Genetic Information 31
2.6.3 Phylogenetic Analysis 33
5
CHAPTER III MATERIALS & METHOD 34
3.1 Malaysian Myxosporean Collections 34
3.2 Hungarian Myxosporean Collections 36
3.2.1 Collection of oligochaetes 36
3.2.2 Sampling period 37
3.2.3 Morphological investigation of actinosporean
types released 37
3.2.4 Prevalence of actinosporean infection 38
3.2.5 Histology 38
3.3 DNA Isolation, PCR, and Sequencing of Myxosporean 38
CHAPTER IV RESULTS 41
4.1 Malaysian Collections (Myxospore) 41
4.1.1 Myxobolus tambroides sp. n. 42
4.1.2 Myxobolus sp. I 43
4.1.3 Myxidium sp I 44
4.1.4 Myxidium sp II 49
4.1.5 Myxobolus ophiocarae sp. n. 49
4.2 Hungarian Collections (Actinospore) 51
4.2.1 Collective Group Aurantiactinomyxon Janiszewska, 1957 51 4.2.1.1 Aurantiactinomyxon Type 1 51 4.2.1.2 Aurantiactinomyxon Type 2 52 4.1.2.3 Aurantiactinomyxon Type 3 54 4.2.1.4 Aurantiactinomyxon Type 4 59
6
4.2.1.5 Aurantiactinomyxon Type 5 60 4.2.2 Collective Group Raabeia Janiszewska, 1955 62
4.2.2.1 Raabeia Type 1 62
4.2.2.2 Raabeia Type 2 63
4.2.3 Collective Group Neoactinomyxum Granata, 1922 65
4.2.3.1 Neoactinomyxum Type 1 65
4.2.4 Collective Group Synactinomyxon Štolc, 1899 66
4.2.4.1 Synactinomyxum Type 1 66
4.3 Elucidation of Life Cycles 69
4.3.1 Actinospore stage 69
4.3.1.1 Description of Ttriactinomyxon types 69
4.3.2 18S rDNA analysis 70
CHAPTER V DISCUSSION 74
5.1 Myxosporeans Fauna 74
5.2 Actinosporeans Fauna 77
5.3 Life Cycles Elucidation 82
CHAPTER VI CONCLUDING REMARK &
RECOMMENDATIONS 88
LIST OF NEW FINDINGS 90
ACKNOWLEDGEMENTS 91
REFERENCES 92
PUBLICATIONS LIST 104
7 LIST OF TABLES
Table 1. List of Actinosporean Collective Groups. Abbreviation: FW = freshwater, M = marine ... 23 Table 2. List of Myxosporeans collected in Malaysia. Abreviations: SP, Spore features (LSB, length of the spore body; WSB, width of the spore body; TSB, thickness of the spore body; PC, polar capsule; CP, caudal processes). The dimensions (in micrometers) of the spores are indicated as length × width × thickness; and those of the polar capsules as length × width ... 26 Table 3. Primers used for PCR and sequencing ... 40 Table 4. Number, range of total length (TL) in centimeter and weight (TW) in gram, of examined fishes. ... 41 Table 5. Descriptive data of the aurantiactinomyxon morphotypes collected in this study (measurements in µm) ... 55 Table 6. Descriptive data of the raabeia, synactinomyxon, and neoactinomyxum
morphotypes collected in this study (measurements in µm) ... 56 Table 7. Comparison of M. fundamentalis, M. erythrophthalmi and M. shaharomae on the basis of actinospores morphometric. ... 72 Table 8. Genetic similarity based on 18S rDNA sequence of the triactinomyxon types from the Lake Balaton and Kis Balaton reservoir to the myxospore species found from the same localities ... 73
LIST OF FIGURES
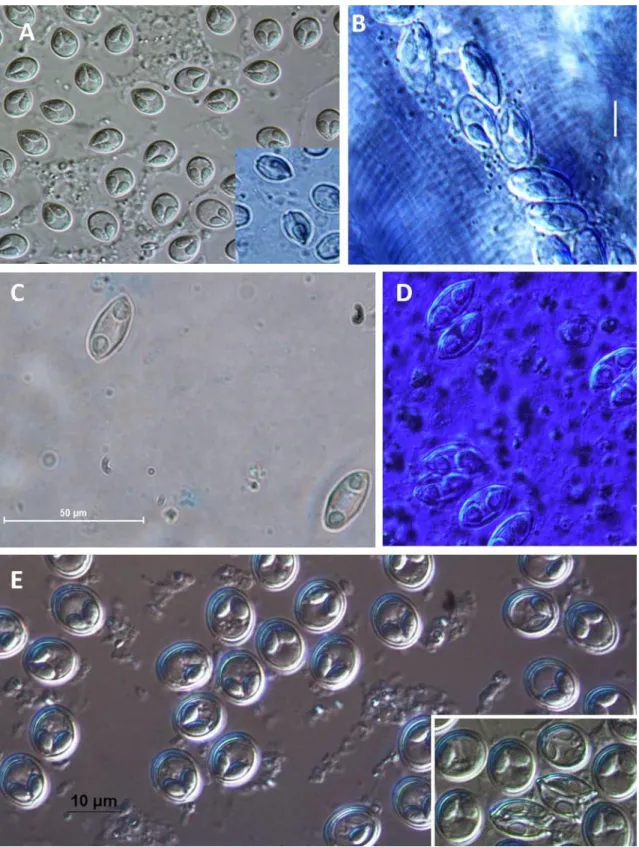
Figure 1. Diagram of the life cycle of myxosporean alternating fish and annelid hosts; A:
The polar filaments are extruded to anchor the spore to the gut epithelium, followed by opening of shell valves of myxospore. B: Gametogony. C: Sporogony of actinosporean phase. D: Mature actinospore stages develop in a pansporocyst, and actinospores are released into the water. E: Upon contact of actinospores with the skin or gills of the fish host, polar filaments extrude to anchor the spore to the skin or gills, facilitating invasion of the sporoplasms into the fish. F: Presporogonic multiplication in a cell-in-cell stage. G:
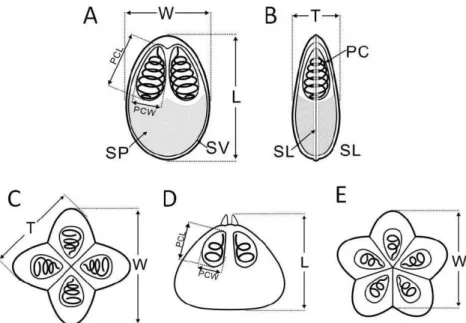
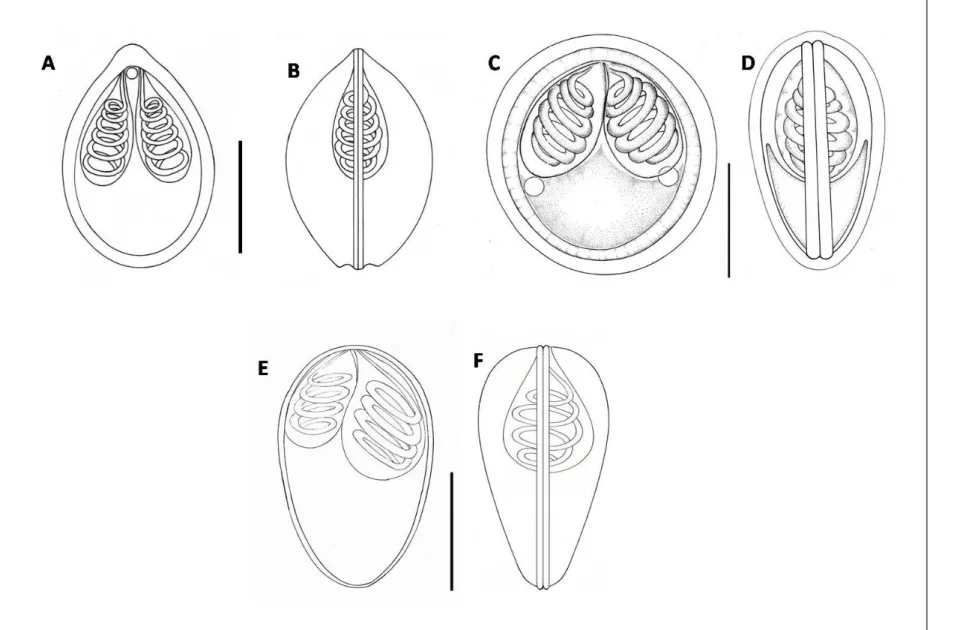
Sporogony of myxosporean phase (Adopted from Yokoyama et al. 2012). ... 20 Figure 2. Diagrams of bivalvulid (A: frontal view, B: side view) and multivalvulid (C &
E, top view, D: side view) myxosporean spores. PC: polar capsule, SP: sporoplasm, SV:
shell valve, SL: sutural line, L: spore length, W: spore width, T: spore thickness, PCL:
polar capsule length, PCW: polar capsule width (Adopted from Yokoyama et al. 2012). 32
8
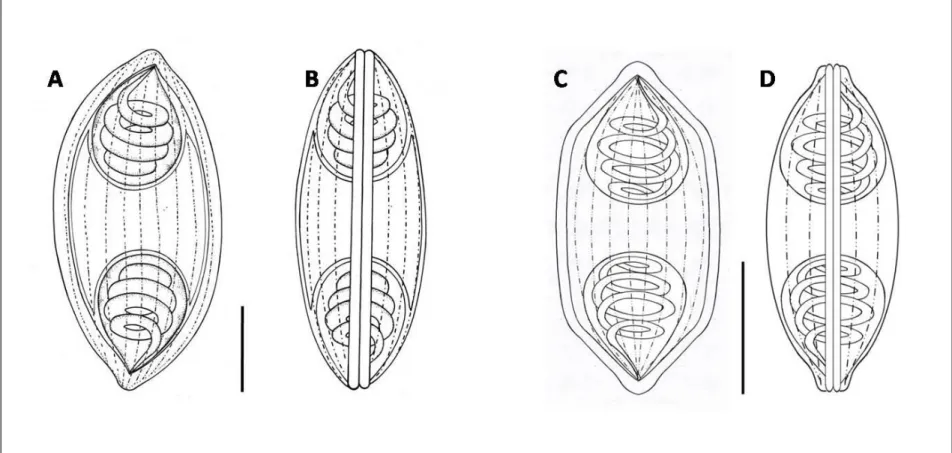
Figure 3. Diagram of actinosporean spores. A: Triactinomyxon, B & C:
Aurantiactinomyxon, D & E: Neoactinomyxum, F & G: Tetractinomyxon.B, D & F: top views , C, E & G: side view. SB: spore body, LSB: length of spore body; WSB: width of spore body; S: style; LS: length of style; WS: width of style; CP: caudal process; LCP:
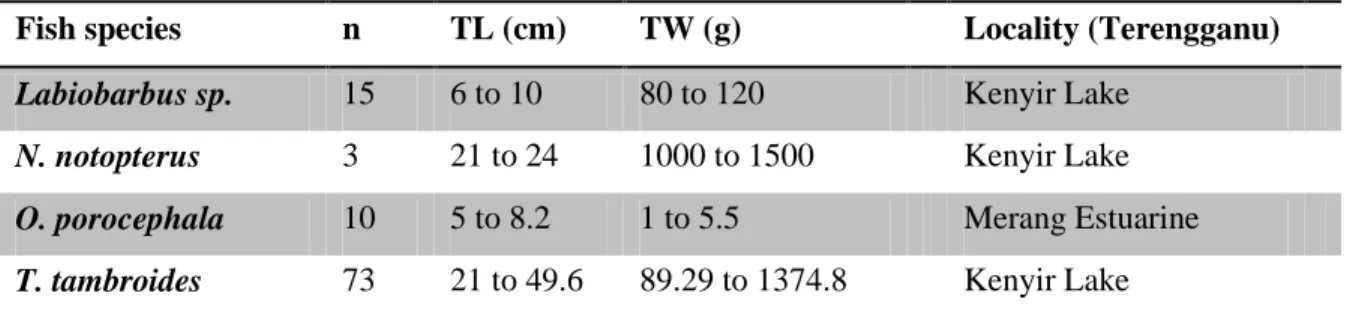
length of caudal process (regardless of curvature). LSCP: largest span of between the tips of the caudal processes; PC: polar capsule; DSB: diameter of spherical spore body (Adopted from Yokoyama et al. 2012) ... 32 Figure 4: Locations of myxozoa (myxospores) sampling in Malaysia ... 35 Figure 5: Locations for myxozoa (actinospores) sampling in Hungary ... 36 Figure 6. (A) Myxobolus tambroides sp. n.; (B) Myxobolus sp. I; (C) Myxidium sp. II; (D) Myxidium sp. I; (E) Myxobolus ophiocarae sp. n. Inset side view of spore... 45 Figure 7. Line drawings from frontal and sutural view of myxospores. (A, B) Myxobolus tambroides sp. n. ; (C, D) Myxobolus ophiocarae sp. n.; (E, F) Myxobolus sp. I. Bar = 10µm ... 46 Figure 8. Line drawings from frontal and sutural view of myxospores. (A, B) Myxidium sp. II; (C, D) Myxidium sp. I. Bar = 5µm ... 47 Figure 9. Phylogenetic tree generated by maximum likelihood analysis of the 18S rDNA sequences of myxosporeans and some related species. Numbers at nodes indicate the bootstrap values (ML) and posterior probabilities (BI). Ceratomyxa spp. were used as the outgroup... 48 Figure 10. Microphotographs of actinosporeans. A, B aurantiactinomyxon type-1; C, D aurantiactinomyxon type-2 ... 53 Figure 11. Line drawings from apical and side view of actinosporeans. A, B aurantiactinomyxon type-1; CD aurantiactinomyxon type-2. Bar = 10 µm ... 57 Figure 12. Line drawings from apical and side view of actinosporeans. A, B aurantiactinomyxon type-3; CD aurantiactinomyxon type-4; E aurantiactinomyxon type-5.
Bar = 10 µm... 58 Figure 13. Microphotographs of actinosporeans. A, B aurantiactinomyxon type-3; C, D aurantiactinomyxon type-4 ... 60 Figure 14. Microphotograph of aurantiactinomyxon type-5 ... 61 Figure 15. Microphotographs of actinosporeans. A, B raabeia type-1; C, D raabeia type-2 ... 64
9
Figure 16. Microphotographs of actinosporeans. A, neoactinomyxum; B, C, synactinomyxon... 67 Figure 17. Line drawings of actinosporeans. A raabeia type-1; B raabeia type-2; C synactinomyxon; D, E frontal & apical veiw of synactinomyxon spore body; F, G neoactinomyxum. Bar. Figs. A, B, C = 50µm; Figs. D, E, F, G = 10µm ... 68 Figure 18. Phase contrast micrographs and schematic diagrams of triactinomyxon spores.
Inset: detail of spore body. (A and B) M. fundamentalis actinospore stage; (C and D) M.
erythrophthalmi actinospore stage; (E and F) M. shaharomae actinospore stage. ... 71 Figure 19. Schematic illustration of M. erythrophthalmi life cycle: Triactinomyxon-type actinospores (A) infect the vertebrate host S. erythrophthalmus (V) in which then develop myxospores (M) that infect the invertebrate host I. michaelseni (I). ... 74 Figure 20. Schematic illustration of M. fundamentalis life cycle: Triactinomyxon-type actinospores (A) infect the vertebrate host R. rutilus (V) in which then develop myxospores (M) that infect the invertebrate host I. michaelseni (I). ... 74 Figure 21. Schematic illustration of M. shaharomae life cycle: Triactinomyxon-type actinospores (A) infect the vertebrate host A. alburnus (V) in which then develop myxospores (M) that infect the invertebrate host I. michaelseni (I). ... 74 Figure 22. Schematic illustration of T. kitauei life cycle: Aurantiactinomyxon-type actinospores (A) infect the vertebrate host C. carpio (??) (V) in which then develop myxospores (M) that infect the invertebrate host B. sowerbyi (I). ... 74 Figure 23. Schematic illustration of T. nikolskii life cycle: Aurantiactinomyxon-type actinospores (A) infect the vertebrate host C. carpio (V) in which then develop myxospores (M) that infect the invertebrate host Nais sp. (I). ... 74
10 ABSTRACT
Studies on Fish Parasitic Myxozoans in Lake Balaton, Hungary and in Freshwater and Marine Biotopes in Malaysia
There are only few data on the occurrence of parasitic myxozoans on Malaysian fishes. In this study, we report on collection of some new myxosporean species between 2010 and 2013 from this country. Four of them infected freshwater fishes; one was collected from an estuarine fish host. Besides morphological characterization of the species found, we made a molecular analysis on their 18S rDNA. In this study, we described Myxobolus tambroides sp. n. (60%) from the gills of an appreciated cyprinid, Tor tambroides collected from the Lake Kenyir Water-reservoir. Another new species elucidated was Myxobolus ophiocarae sp. n. (8.3% prevalence) found in Ophiocara porocephala (Gobiidae) collected from Merang Estuarine, Kuala Terengganu. The following specimens are yet to be described: Myxobolus sp. I. (15% prevalence) was found in the muscle tissue of a Labiobarbus sp. (Cyprinidae). Sequences of Myxobolus sp.
I resembled to M. cyprini with 94.6% similarity. Myxidium sp. I (66.7% prevalence) was found in the gall bladder of Notopterus notopterus (Notopteridae). Myxidium sp. II (15.4%
prevalence) was found in the gall bladder of Tor tambroides (Cyprinidae). For the Myxidium sp. I, partial sequences of the specimen showed 90.6% similarity to M.
cuneiforme; while complete sequences of Myxidium sp. II specimen collected from T.
tambroides showed 87.2% identity to M. anatidum. Phylogenetic analyses of the 18S rDNA were performed to estimate and analyze relationships between myxozoan entities.
In another study made in Hungary after morphological analysis, we have examined 18S rDNA sequences of actinospores isolated from oligochaetes of Lake Balaton and Kis- Balaton Reservoir and compared them with sequences of known myxospore stages. In between 2010 to 2012, we studied the natural infection of the oligochaetes, Branchiura sowerbyi Beddard, 1892, Isochaetides michaelseni Lastockin, 1936 and Nais sp. Müller, 1774. Thirteen actinosporean stages (4 triactinomyxon-types, 5 aurantiactinomyxon-types, 2 raabeia-types, 1 neoactinomyxum-type and 1 synactinomyxon-type) were found. Our molecular data showed that three types of actinospores (triactinomyxon-type 1, 2, 3) had 99.9-100% similarity to myxospores of Myxobolus erythrophthalmi, M. shaharomae and M. fundamentalis. Another complete sequence analysis of aurantiactinomyxon-type 4 collected from B. sowerbyi showed 99.4% match with Thelohanellus kitauei Egusa &
Nakajima, 1981, though the myxospores of T. kitauei have not been recorded in Hungary hitherto. In addition, partial sequence of aurantiactinomyxon-type 5 isolated from a Nais sp., corresponded to Thelohanellus nikolskii Akhmerov, 1955 showing 99.9% similarity.
Elucidation of the myxosporean fauna and their life cycle presented herein shows significant diversity among pre-investigated biotopes (Lake Kenyir, Merang Estuary, Lake Balaton and Kis Balaton). During this study, we have detected five new species of myxospores from Malaysia, we have presented new data on actinosporeans infecting oligochaetes in Lake Balaton and revealed the life cycle of five new myxosporeans in Hungary.
11 KIVONAT
Vizsgálatok Balatoni-, Valamint Malajziai Édesvízi- És Tengeri Biotópokon Élősködő Halparazita Nyálkaspórásokon
Malajziai nyálkaspórás paraziták vonatkozásában napjainkig csak kevés szakirodalmi adat áll rendelkezésre. Disszertációmban a már meglévő ismeretanyaghoz további adatokat szolgáltatok, és beszámolok néhány 2010 és 2013 között gyűjtött új malajziai nyálkaspórás fajról, melyek közül négy faj édesvízi hal-gazdákból, egy pedig brakkvízi halgazdából került kimutatásra. A kimutatott nyálkaspórás fajok morfológiai jellemzése mellett azok molekuláris biológiai módszerekkel való vizsgálatát (18S rDNS gén) is elvégeztem. Az első kimutatott fajt a Tasik Kenyir víztározóban élő, Malajziában magasan értékelt pontyféle, a Tor tambroides kopolyújáról gyűjtöttük, és azt Myxobolus tambroides néven új fajként írtuk le (Székely et al., 2012). A Kuala Terengganu közelében található Merang Lagunából származó Ophiocara porocephala nevű géb faj kopoltyújáról ugyancsak kimutattunk egy új nyálkaspórást (8.3% prevalencia), melyet Myxobolus ophiocarae néven írtunk le. A Tasik Kenyir Víztározóból gyűjtött további fajok leírása folyamatban van. Ezek a következők: egy Labiobarbus sp. (Cyprinidae) izomzatában élősködő Myxobolus faj (15% prevalencia). Szekvenciája 94.6%-os genetikai hasonlóságot mutatott a génbankban megtalálható M. cyprini fajjal. Notopterus notopterus (Notopteridae) epehólyagjából izolált Myxidium faj (66.7% prevalencia). Részleges szekvenciája 90.6%-os hasonlóságot mutatott a M. cuneiforme fajjal, míg a Tor tambroides (Cyprinidae) epehólyagjából kimutatott Myxidium faj teljes szekvenciája 87.2%-os genetikai hasonlóságot mutatott a M. anatidum-mal. A kimutatott új nyálkaspórás élősködők rendszertani besorolásához filogenetikai elemzéseket végeztem.
Magyarországi vizsgálataim során a Balatonból és Kis-Balatonból 2010 és 2012 között gyűjtött kevéssertéjű férgekből (Branchiura sowerbyi Beddard, 1892, Isochaetides michaelseni Lastockin, 1936 és Nais sp. Müller, 1774) több aktinospora típust izoláltunk.
Ezeket morfológiailag jellemeztük, majd molekuláris módszerekkel meghatároztuk 18S rDNS szekvenciájukat. A kapott adatokat egybevetettük a Génbankban megtalálható adatokkal. Összesen 13 aktinospora típust izoláltunk, melyekből 4 a triactinomyxon típusba, 5 az aurantiactinomyxon típusba, 2 a raabeia típusba, 1 pedig a neoactinomyxum típusba volt sorolható. Molekuláris vizsgálataink alapján az 1., 2. és 3. triactinomyxon típusok 99.9-100% hasonlóságot mutattak a M. erythrophthalmi, M. shaharomae és M.
fundamentalis nevű, Balatonból leírt nyálkaspórás fajokkal, melyek a vörösszárnyú keszeg, a küsz és a bodorka parazitái. A B. sowerbyi-ből izolált 4. aurantiactinomyxon típus teljes szekvenciája 99.4% azonosságot mutatott a Thelohanellus kitauei Egusa &
Nakajima, 1981 parazitával, jóllehet ezt a fajt még nem mutatták ki hazai halakból. Egy Nais fajból izolált 5. aurantiactinomyxon típus részleges szekvenciája 99.9% hasonlóságot mutatott a Thelohanellus nikolskii Akhmerov, 1955 fajjal. Az egyes biotópokon (Lake Kenyir, Merang Estuary, Balaton és Kis Balaton) vizsgált myxozoák előfordulása jelentős diverzitást mutatott. A munka során öt új malajziai nyálkaspórás fajt sikerült kimutatnunk, új adatokat szolgáltattunk a Balatonban élő aktinosporákról, valamint tisztáztuk a Balatonban élő halakon élősködő öt nyálkaspórás faj fejlődési ciklusát.
12 ABSTRAK
Kajian ke atas Parasitik Myxozoa Ikan di Tasik Balaton, Hungary dan di Biotop Air Tawar dan Marin di Malaysia
Terdapat hanya beberapa data mengenai kehadiran parasit myxozoan pada ikan Malaysia. Dalam kajian ini, kami melaporkan koleksi baru spesies myxosporean antara tahun 2010 dan 2013. Empat spesis menjangkiti ikan air tawar; satu species dikumpulkan dari ekosistem muara. Selain pencirian morfologi spesies yang ditemui, analisis molekular 18S rDNA juga dijalankan pada mereka. Dari kajian ini, Myxobolus tambroides sp. n.
dikenalpasti dari insang Tor tambroides yang diambil dari Tasik Kenyir (Székely et al.
2012). Satu lagi spesies baru dijelaskan adalah Myxobolus ophiocarae sp. n. (8.3%
kekerapan) telah ditemui di Ophiocara porocephala (Gobiidae) diambil dari muara Merang , Kuala Terengganu. Spesimen berikut masih belum di identifikasi; Myxobolus sp.
I. (15% kekerapan) telah dijumpai dalam tisu otot Labiobarbus sp. (Cyprinidae). Jujukan sikuensi Myxobolus sp. I menyerupai M. cyprini dengan 94.6% persamaan. Myxidium sp. I (66.7% kekerapan) ditemui di dalam pundi hempedu Notopterus notopterus (Notopteridae). Myxidium sp. II (15.4% kekerapan) ditemui di dalam pundi hempedu Tor tambroides (Cyprinidae). Bagi Myxidium sp. I, sebahagian jujukan sikuensi spesimen menunjukkan 90.6% persamaan dengan Myxidium cuneiforme, manakala jujukan lengkap Myxidium sp. II yang dikutip dari T. tambroides menunjukkan identiti 87.2% kepada M.
anatidum. Filogenetik analisis 18S rDNA telah dijalankan untuk menganggarkan dan menganalisis hubungan antara entiti myxozoan.
Selepas analisis morfologi, kajian jujukan 18S rDNA aktinospora yang diasingkan daripada oligocheta Tasik Balaton dan empangan Kis-Balaton dijalankan dan perbandingan mereka dengan jujukan peringkat myxospora dilakukan. Di antara tahun 2010 hingga 2012, kami mengkaji jangkitan semulajadi pada oligochaetes, Branchiura sowerbyi Beddard , 1892 , Isochaetides michaelseni Lastockin , 1936 dan Nais sp. Müller, 1774. Tiga belas aktinospora (4 triactinomyxon -jenis, 5 aurantiactinomyxon -jenis, 2 raabeia -1 neoactinomyxum -jenis dan 1 synactinomyxon -jenis ) telah dijumpai. Data molekular menunjukkan tiga aktinospora ( triactinomyxon - jenis 1 , 2, 3 ) mempunyai 99,9-100 % persamaan dengan M. erythrophthalmi , M. shaharomae dan M.
fundamentalis myxospores. Satu lagi analisis jujukan lengkap aurantiactinomyxon – jenis 4 menunjukkan 99.4 % persamaan dengan Thelohanellus kitauei Egusa & Nakajima, 1981, walaupun myxospores T. kitauei belum direkodkan di Hungary sehingga kini. Di samping itu, jujukan separa aurantiactinomyxon - jenis 5 dari Nais sp., sepadan dengan Thelohanellus nikolskii Akhmerov, 1955 dengan 99.9% persamaan. Myxosporeans fauna dan penjelasan kitaran hidup mereka yang dibentangkan di sini menunjukkan kepelbagaian yang ketara dalam biotopes yang disiasat (Tasik Kenyir, Muara Merang, Lake Balaton dan Kis Balaton ). Kajian ini melampirkan lima spesies baru myxospores dari Malaysia, data baru akinospora dan lima kitaran hidup baru myxozoa dari Hungary.
13 CHAPTER I INTRODUCTION
Several diseases and disorders have associated with fishes over the years by various pathogens. Fishes, both wild and captivated are susceptible or vulnerable to pathogens-carrying organisms such as parasites, viruses, bacteria and others. Parasites for example, possesses an ability to introduce diseases via several mechanisms such as outnumbering or sizing up their population and localized themselves in more or less important organs of the host. Other parasite might cause deprivation of food upon feeding on blood or withdraw vitamins and important substances of the host (Mehlhorn, 2001).
Some parasites have been determined as a serious threat in aquaculture and fish farming practices. Therefore, interest in fish parasitic group has intensified due to their threats and a substantial amount of research has been conducted, particularly those that cause morbidity and mortality in fish (Woo, 2006).
Although wild fish are commonly infected by parasites and they are usually evolved, yet under unusual circumstances the parasites could threaten the host populations in nature. On the other hand, when fishes are confined or clustered under relatively crowded conditions, disease outbreaks are more likely to occur. Hence, mass production of cultured fish is always accompanied by threatening parasites and pathogens agent (Rückert et al., 2008). Infection by species of Phylum Myxozoa Grasse, 1970 for example, is worthwhile to note as some of them are highly pathogenic to their hosts.
Two exclusive classes belong to Phylum Myxozoa; 1) Class Malacosporea Canning, Curry, Feist, Longshaw et Okamura, 2000 and; 2) Class Myxosporea Bütschli, 1881. Myxozoa was once classified as protozoan parasites that were later described as metazoan parasites in the last decades of 1900’, largely due to the multicellular nature of the spores (Canning et Okamura, 2004). Wolf et Markiw (1984) discovered the role of annelids (Tubificidae) as an alternative host in Myxobolus cerebralis life cycle. This finding is contradicted to the general understanding of that age, that myxozoan parasites are transmitted directly between their vertebrate hosts. Molecular data, through parsimony
14
and neighbor-joining methods, supported the position of myxozoans as metazoan lineage, as other have suggested on the basis of morphological criteria (Smothers et al. 1994). In different studies Cavalier-Smith et al. (1996) and Siddall et al. (1995) suggested that the myxozoans are Cnidarian affinity by the presence of nematocyst-like polar capsules.
Findings over myxozoan research has drawn questions of origin and evolutionary aspect as whether they originated in aquatic invertebrates and spread to fish or vice versa, hence the fundamental phylogenetic placement of the Myxozoa remains uncertain (Canning et Okamura, 2004). The utilization of annelids as alternative hosts is only appliable for the members of Class Myxosporea, while members of Class Malacosporea are exclusively, or up until now, using bryozoans as their alternative hosts.
It has been generally accepted that the life cycle of the Class Myxosporea takes place in two hosts with two developing phases called myxospore and actinospore. The myxospore phase develop in vertebrate hosts (i.e. fishes, reptiles, amphibians), with spores typically cruising in large plasmodia, called pansporocysts. Meanwhile, the actinospores develop in the epithelium of the intestinal truct of the mud dwelling annelid hosts.
Myxosporean are present at different geographic areas, particularly as fish parasites and quite rare as parasites of reptiles and amphibians (Lom et Dyková, 2006). However, the vertebrates host ranges are now extended to terrestrial mammals and also to avians. There is no clear pattern regarding host specificity and site preference within the myxozoa. Some species appear to be highly specific to their host, whilst others may be specific at the family level.
On the course of present study, particular interests were given to the Class Myxosporea of freshwater ecosystems of Hungary and Malaysia, with an addition of attention to estuarine and marine ecosystem in the latter region. In Hungary, intensive work on myxosporean was conducted since 1980's until the present time. In these years, efforts have been made to gather a collection of myxospores infecting fishes and actinospores from several species of annelid hosts. A broad myxosporean diversity was reported from Hungarian freshwater biotopes. Twenty three actinospore types from four different actinospore collective groups were recorded and distinguished exclusively by
15
morphological studies from the Lake Balaton and Kis Balaton (El-Mansy et al., 1998 Kelemen et al., 2009). Meanwhile, almost a hundred myxosporean species of various genera such as Henneguya, Hoferellus, Myxidium, Myxobilatus, Myxobolus, Sphaerospora, and Thelohanellus were collected from Hungarian fishes. Among them, Myxobolus spp. were the most dominant species, which constitute almost 90% of the collections (Molnár et Székely, 1995, 1999; Székely et Molnár, 1996; Molnár et al.
2006c). Apart from diversity compilation, experimental infections had also been conducted to reveal myxosporean life cycles in Hungary.
On the other hand, biodiversity of myxozoans is a much underestimated area in Malaysia. Nevertheless, findings in recent years on the parasitic myxozoans of both wild and cultured Malaysian fishes in the recent years (Molnár et al., 2006a, b; Székely et al., 2009a, b) showed high taxonomic diversity of the group. Minimal works, or if any, not well documented study, were conducted on myxozoan association with Malaysian fishes.
Hence, there are relatively little known facts about the species composition of parasitic Myxozoa in Malaysian environments. Therefore, knowledge of their hosts association, speciation and their regionalization is even less. The first paper concerning myxozoan infection of Malaysian fishes was written by Shariff (1982), who presented a complex morphological and pathological study on the parasites of a perciformid fish, Oxyeleotris marmoratus (Bleeker), and described a new species Henneguya shaharini Shariff, 1982.
Examinations on the myxozoan fauna of Malaysian fishes were then conducted subsequently by Molnár et al. (2006a, b) and Székely et al., (2009a, b).
The importance of parasitic myxozoan studies could not be excluded with a variety of interests and specialization regarding this topic. Molecular biological analyses, in a wide range of applications, constitute the current trends in the myxozoan study, especially in characterization and classification. Due to close morphological characteristics and similarities of some species, scientists, especially taxonomist, need to incorporate a genetic analysis into their work to support their findings. However, , morphometric, ultrastructre and other biological studies as well as experimental infections are also important branches in myxozoan research.
16
Realizing the diversity richness of myxosporeans and their potential to act as a pathogens in fish culture, the present dissertation tries to compile myxosporean collection from Hungarian and Malaysian biotopes, and including a first estuarine assessment of parasitic myxozoans in Malaysia. Principle aspect of this dissertation emphasizes on finding the parasitic spores of myxosporean infection on both fish (myxospore) and alternate annelids (actinospore) host with comprehensive morphological works. In depth coverage, essential knowledge about the biology of this class was gathered through molecular biology with particular focus on genetic analysis for identification. Eventually, phylogenetic analysis which is now crucial upon almost every aspect of biological research, were implemented to estimate and analyze relationships between myxozoan entities.
Through morphological assessment and genetic study on the 18S rDNA sequences, this study specifically embarks on the following objectives:
1. To identify the species of myxospores of Class Myxosporea infecting wild and captivated Malaysian fishes.
2. To characterize actinosporean fauna from Lake Balaton and Kis Balaton Water Reservoir in Hungary
3. To identify myxospore and actinospore pairs of Class Myxosporea from Hungarian collection materials.
17 CHAPTER II LITERATURE REVIEW
Recent findings on diversity, ecology, and pathogenicity of parasitic Myxozoan in pisciculture have attracted a significant interest. The present collection of parasitic myxosporean knowledge from this study, as limited as it may be, has significantly showed the high diversity of the group. This fact was the bases for this project, which aims to examine and assess current understanding of myxosporean fauna in freshwater (lakes) ecosystems of Hungary as well as freshwater and marine (brackish, estuarine) bitopes in Malaysia.
2.1 Phylum Myxozoa
Myxozoans are highly specialized metazoan parasites with 2-host life cycle between aquatic invertebrates and vertebrates in both marine and freshwater biotopes.
Most of myxozoans are not typically transmissible to humans and harmful to host fish;
however, several species cause diseases in cultured and wild fish which are problems for aquaculture and fishery industries worldwide (Lom et Dyková 2006, Yokoyama et al.
2012).
Transmissible myxozoan infection, particularly in susceptible populations of cultured fishes, can be devastating with significant mortality rates. For example the Tetracapsuloides bryosalmonae for proliferative kidney disease (PKD) and Ceratomyxa shasta for ceratomyxosis on farmed salmonid fish (Canning et Okamura 2004; Yokoyama et al. 2012). Other species such as Myxobolus koi, Thelohanellus hovorkai, and Sphaerospora dykovae (syn. S. renicola) are well-known pathogens in cultured common carp (Cyprinus carpio) in Europe and Asia, which cause a respiratory dysfunction, extensive hemorrhages and edema, as well as swimbladder inflammation, which led to death of affected fish (Dyková et Lom 1988, Yokoyama et al. 1997, 1998, 2012). In the western region of United States, Myxobolus cerebralis is responsible for high mortality in wild rainbow trout (Onchorhyncus mykiss) population. Investigation on the biology of M.
18
cerebralis by Wolf et Markiw (1984) has revealed the requirement of actinosporean, Triactinomyxon-type actinosporeans, as an alternate life cycle stage of the parasite. This discovery has led to a substantial increase in the understanding on myxozoan life cycle.
Subsequently, scientists have focused their attention to biological studies of actinosporeans, such as their emergence from the annelid host, the waterborne phase, mechanism of their invasion and entrance variability into fishes. Invasion process was also investigated in relation to the mechanism of host specificity in the parasites (Yokoyama et al. 2012).
Even though numerous identification and taxonomic study have been conducted on myxozoans, interest in this group has intensified along the development of fisheries aquaculture for their significant impact or risks on the cultured fish (Morris et Freeman, 2010). Phylum Myxozoa which counts more than 2200 species and in which 58 genera have been described to date, is represented by two classes, the Myxosporea, containing all the better-known genera, which alternate between fish and annelids, and the Malacosporea, which parasite bryozoans (Canning et Okamura 2004, Lom et Dyková 2006, Yokoyama et al. 2012). In the course of this study, review is mainly concerned with myxozoans of the class Myxosporea.
2.2 Class Myxosporea
Early documentations have classified the Actinosporea and Myxosporea as independent zoological classes. However, due to the finding by Wolf et Markiw (1984), taxonomical classification of actinosporean has been uncertain as scientist went through a series of disagreements over their systematic and nomenclature position (Lester et al.
1998,1999; Kent et Lom 1999). Through a careful and thorough review of the outlined proposals by Kent et al. (1994), a consensus summary on actinosporean’s nomenclature position has been agreed, followed by the suppression of the class Actinosporea, becoming a synonym of the class Myxosporea Bütschli, 1881, leaving a single class in the phylum (Kent et al. 1994, 2001; Hallett et al. 1998, 1999; Hallett et Lester 1999; Negredo et Mulcahy 2001; Özer et al. 2002). Since this time, actinosporeans were then treated as
19
“collective-group”, a term that was used to characterise different morphological types of actinosporeans.
To date, only four species were identified from class Malacosporea, whereas the rest of myxozoan species belonged to class Myxosporea. Class Myxosporea is comprised of the two orders, Bivalvulida and Multivalvulida. Bivalvulids include 52 genera with more than 2100 species described from freshwater and marine fishes, while multivalvulids contain 5 genera with more than 60 species predominantly from marine fish (Lom et Dyková, 2006, Yokoyama et al. 2012).
2.3 Myxozoan Life Cycle
It has been well accepted that as a rule the life cycle of myxosporeans takes place in two hosts and the term “myxosporean” covers both life-cycle phases: 1) the myxospore- infective stage to oligochaete host, and; 2) the actinospore - infective stage to fish hosts (Fig. 1). Several studies have confirmed that some myxozoans undergo a two host life cycle (Feist et Longshaw 2006, Yokoyama et al. 2012). These include representatives of the genera Myxidium, Sphaerospora, Hoferellus, Myxobolus, Thelohanellus and Zschokkella. In present work, interest was focused mainly on the Myxobolus spp. life cycle.
Developmental cycles of different Myxobolus spp. were for a long time studied by experimental infection of oligochaetes with myxospores and infection of the host fishes with actinospores: M. arcticus (Kent et al. 1993, Urawa 1994), M. cotti (El-Matbouli et Hoffman, 1989), M. carassi (El-Matbouli et Hoffman, 1993), M. djargini (El-Mansy et Molnár 1997a), M. hungaricus (El-Mansy et Molnár 1997b), M. portucalensis (El-Mansy et al.1998), M. bramae (Eszterbauer et al. 2000), M. pseudodispar (Székely et al. 2001), M. macrocapsularis (Székely et al. 2002), M. intimus (Racz et al. 2004), M. parviformis (Kallert et al. 2005), M. rotundus (Székely et al. 2009c).
20
Recently the tiresome and complicated experimental studies are substituted by searching the actinosporean-myxosporean pairs of the myxosporean species in DNA studies. In this studies the counter parts of a given species, M. rotundus (Hallett et al.
2005, Székely et al. 2009c); M. diversicapsularis (Hallett et al. 2005, Molnár et al. 2010) and M. wootteni (Eszterbauer et al. 2006, Molnár et al. 2010) were identified by the correspondence of the DNA of an actinosporean type and a Myxobolus sp. In most cases triactinomyxon type actinosporeans correlated with Myxosporean partners.
Figure 1. Diagram of the life cycle of myxosporean alternating fish and annelid hosts; A: The polar filaments are extruded to anchor the spore to the gut epithelium, followed by opening of shell valves of myxospore. B: Gametogony. C: Sporogony of actinosporean phase. D: Mature actinospore stages develop in a pansporocyst, and actinospores are released into the water. E: Upon contact of actinospores with the skin or gills of the fish host, polar filaments extrude to anchor the spore to the skin or gills, facilitating invasion of the sporoplasms into the fish. F: Presporogonic multiplication in a cell-in-cell stage. G: Sporogony of myxosporean phase (Adopted from Yokoyama et al. 2012).
21 2.3.1 Myxospores and Vertebrate Hosts
Myxosporean are common in freshwater and marine fishes. Of them, 3 species have been reported in Agnatha, 35 species in Chondrichthyes, and the rest in Osteichthyes.
Though they were mostly found in fishes, in recent years, myxosporeans were also known from other vertebrates such as frog, turtles, waterfowl (Anseriformes) and the terrestrial mammal shrews (Soricomorpha) (Prunescu et al., 2007; Bartholomew et al., 2008; Feist et Longshaw, 2006; Lom & Dyková, 2006). Lowenstine et al. (2002) reported six cases of developmental stages and unassigned spores found in inflammatory lesions of hepatic ducts in anatid ducks collected from enclosures in zoological gardens. In addition, myxozoan infections have recently been found in the liver and bile ducts of North American waterfowl, described as Myxidium anatidum, found in the lumen of afferent bile ducts in liver.
Repeated presence of myxozoan spores (Myxobolus) in the faeces of humans suffering from abdominal pain and/or diarrhea or infected with human immunodeficiency virus (HIV) have been also reported (Boreham et al. 1998, Moncada et al. 2001, Hessen et Zamzame 2004, Canning et Okamura 2004, Lom et Dyková 2006). These report led Canning et Okamura (2004) to postulate that myxozoans may under certain conditions become opportunistic parasites of homeothermic vertebrates.
2.3.2 Actinospores and Invertebrate Hosts
There have been 18 collective-groups described by far, with around 136 types which have been identified in recent years (Lom et Dyková 2006; Özer et al. 2002;
Canning et Okamura, 2004). Two additional novel collective groups were described:
Unicapsulactinomyxon-type revealed infecting the marine polychaete, Diopatra neapolitana in the Aveiro Estuary, Portugal (Rangel et al. 2011); and Triactinospomyxon- type collected from Branchiura sowerbyi of Wuhan, China (Zhai et al. 2012). Generally, the most common types described belong to Triactinomyxon- (53), Aurantiactinomyxon- (34), Raabeia- (25), Echinactinomyxon- (19) and Neoactinomyxum- (18) collective
22
groups, followed by others as listed in Table 1 (Lom et Dyková, 2006; Rangel et al. 2011;
Zhai et al. 2012).
Actinosporeans, especially of freshwater species, infect the intestinal epithelial cells of oligochaetes and emerge into the environment by defecation, whereas others species like the actinosporeans of C. shasta were released directly from the epidermis of polychaete Manayunkia speciosa into the water column (Meaders et Hendrickson 2009;
Yokoyama et al. 2012). Although actinospores are mostly reported from freshwater annelids host, such as Branchiura sowerbyi, Tubifex tubifex, Limnodrilus hoffmeisteri, Isochaetides michaelseni, Nais spp., Psammoryctides albicola and Dero digitata, they were also associated with marine worm such as Nereis diversicolor, Nereis succinae, Spirorbis spirorbis, Hydroides norvegica and Chone infundibuliformis (El-Mansy et al.
1998a, b; Özer et al. 2002; Eszterbauer et al. 2006; Canning et Okamura 2004; Køie 2000, 2002; Székely et al. 2007;Yokoyama et al. 2012). To date, the information on the effects of actinosporean infection towards the annelids host is unknown.
2.4 Myxozoan Studies in Hungary
In Hungary, intensive work on myxosporeans was conducted since 90's until the present time. In these years, efforts have been made to gather a collection of myxospores that infect fish species in Hungary, as well as the actinospores from the annelids. Broad myxosporean diversity was reported from Hungarian freshwater biotopes, especially from Lake Balaton and Kis-Balaton. Almost a hundred myxospores species of various genera such as Henneguya, Hoferellus, Myxidium, Myxobilatus, Myxobolus, Sphaerospora, and Thelohanellus were collected from Hungarian fishes. Among them, Myxobolus spp. were the most dominant species constituting almost 90% (~ 60 Myxobolus spp.) of the collections (Molnár et Székely, 1995, 1999, Székely et Molnár, 1995, 1996). Descriptions were mostly based on spore morphology (Lom et Molnár 1983, Molnár et Székely 1995, Székely et Molnár 1996, Molnár 2000) while in some cases a genetic analysis was also carried out (Molnár et al. 2006c, 2007).
23
Table 1. List of Actinosporean Collective Groups. Abbreviation: FW = freshwater, M = marine
Actinospore Collective-group
Number of Findings as:
Total No.
of Types Biotype Regular
Zoological Species
Proven or Putative Developmental
Stage
Antonactinomyxon Janiszewska, 1957 1 2 3 FW
Aurantiactinomyxon Janiszewska, 1952 6 28 34 FW
Echinactinomyxon Janiszewska, 1957 4 15 19 FW
Endocapsa Hallett, Erséus et Lester, 1999 2 0 2 M
Guyenotia Naville, 1930 1 0 1 FW
Hexactinomyxon Štolc, 1899 2 6 8 FW
Hungactinomyxon Rácz, Eszterbauer et
Molnár, 2005 0 1 1 FW
Neoactinomyxum Granata, 1922 4 14 18 FW
Ormieractinomyxon Marques, 1984 1 0 1 FW
Pseudotriactinomyxon Hallett, Atkinson,
Schöl et El-Matbouli, 2003 0 3 3 FW
Raabeia Janiszewska, 1955 3 22 25 FW
Siedleckiella Janiszewska, 1953 1 2 3 FW
Sphaeractinomyxon Caullery et Mesnil, 1904 8 0 8 FW, M
Synactinomyxon Štolc, 1899 2 6 8 FW
Tetractinomyxon Ikeda, 1912 2 4 6 FW, M
Tetraspora Hallett et Lester, 1999 2 0 2 M
Triactinomyxon Štolc, 1899 11 42 53 FW, M
Triactinospomyxon Zhai, Zhou et Gui, 2012 0 1 1 FW
Unicapsulactinomyxon Rangel, Cech, Székely
et Santos, 2011 0 1 1 M
Heliactinomyxon Bellem in Hanson et al.
2001 NA NA NA NA
24
In the recent years several species of the genus Myxobolus have been described from Hungarian cyprinids fishes (Molnár et al. 2009, 2010). Among them Myxobolus erytrophthalmi and Myxobolus shaharomae were described from Scardinius erythrophthalmus (rudd) and Alburnus alburnus (bleak), respectively (Molnár et al.
2009), while, three other new species were erected from Rutilus rutilus (roach) namely Myxobolus fundamentalis, Myxobolus wootteni and Myxobolus sommervillae (Molnár et al. 2010). Nevertheless, we could not exclude the fact on existence of other myxozoan species in Hungarian biotopes yet to be revealed.
Studies on actinosporean fauna associated with oligochaetes have been focusing on Lake Balaton, Kis Balaton reservoir, River Tisza (close to Tiszafüred), and the Temperate Water Fish Farm located at Százhalombatta near Budapest (El-Mansy et al. 1998a, b;
Eszterbauer et al. 2006). Thirty eight isolates of actinospore were evaluated in a morphological study by El-Mansy et al. (1998a, b) and the identity of fourteen actinosporeans were determined through morphology and genetic study by Eszterbauer et al. (2006). In another study, Rácz et al. (2005) described a novel actinosporean, named as hungactinomyxon, which notably differs in spore morphology from actinospore types previously described in Hungary. This latter type was also collected from the fish farm in Százhalombatta.
2.5 Myxozoan Studies in Malaysia
Most of Malaysian myxozoan species described by Molnár et al. (2006a,b) and Székely et al. (2009a,b), originated from the East Coast Region in the state of Kelantan and Terengganu, while a single species described by Shariff (1982) was found in Selangor state, on the West part of Peninsular Malaysia. Investigations of myxozoans of both wild and cultured Malaysian fishes (Molnár et al. 2006a, b; Székely et al. 2009a, b) have showed high taxonomic diversity from fresh waters system, but little attention was turned at this time to the myxozoan infections of marine or brackish water biotopes in Malaysia.
25
Around 19 species have been described in Malaysia using a combination of morphometrics and molecular techniques (Shariff 1982; Molnár et al., 2006a, b; Székely et al., 2009a, b). They could be assigned to nine Myxobolus, six Henneguya, three Hennegoides and one Thelohanellus spp. that were found from wild and cultured fish species (Table 2). Of these numbers, eight myxozoans were recorded from wild fishes;
thirteen were associated with cultured fishes; while one species parasitized both wild and cultured fishes (Table 2). Some species infect fish that are economically important as a fishery resource (Hemibagrus nemurus, Pangasius hypothalamus, Tor tambroides, Leptobarbus hoevenii, Barbonymus gonionotus, Trichogaster trichopterus) in Malaysia, though no general alterations were mentioned that could cause chronic pathogenic to infected fishes (Molnár et al., 2006a,b; Székely et al., 2009a,b). Among the prominent location where myxozoan were reported (Székely 2009b) was Kenyir Lake freshwater reservoir which hosted various appreciated fish species that attracts sport fishing practitioners.
Overall, studies on Malaysian myxozoan are still limited and this is largely due to the fact that, there is lack of interest and poor documentation on this particular group.
Hence, it remains unknown how large the areas colonized by this group of parasite in the Malaysian aquatic biotopes. In this study we discuss findings on the myxosporean in terms of their fauna, phylogenetic relationships and their life cycles.
26
Table 2. List of Myxosporeans collected in Malaysia. Abreviations: SP, Spore features (LSB, length of the spore body; WSB, width of the spore body; TSB, thickness of the spore body; PC, polar capsule; CP, caudal processes). The dimensions (in micrometers) of the spores are indicated as length × width × thickness;
and those of the polar capsules as length × width No. Species Site of
Infection Spore Original Host(s) Type of
host Locality
1
Henneguya shaharini Shariff, 1982
Gills
Immature SP, broad bodies & short caudal processes; mature SP, spindle-shaped in anterior view & lenticular in lateral view, 11.3 (0.6- 12.3) x 3.3 (2.8-3.9) x 2.9 (2.5-3.4); PC 5.9 (5.6- 6.7) x 1.2 (1.1-1.7); CP 23.7 (21.8-28.0)
Oxyeleotris
marmoratus Wild Selangor & Tasik Kenyir, Terenganu
2
Henneguya shariffi Molnár et al., 2006a
Gill filaments
SP symmetric, elongated in shape, 11.7 (10.3- 11.9) x 3.71 (3.6-3.8) x 3.7 (3.6-3.8); elongated shape PC 5.48 (5.1-6.6) x 1.05 (0.9-1.2); short CP 7.54 (5.9-10.3)
Pangasius
hypophthalmus Cultured K.Terengganu, Terengganu
3
Henneguya mystusia Sarkar, 1985
Gill filaments
SP elongated, 12.17 (11-13) x 3.35 (3-3.7) x 2.21 (2-2.25); elongated PC 4.5 (4-5) x 1.01 (0.9-1.2);straight CP 17 (15-20)
Hemibagrus
nemurus Cultured K. Terengganu, Terengganu
4
Henneguya hemibagri Tchang et Ma,
1993
Gill lamellae
SP elongated 10.03 (9-11.2) x 2.91 (2-4) x 2.06 (1.8-2.5);PC located one behind the other, 1.35 (1.3-1.5) x 1.35 (1.3-1.4); strongly bifurcated CP, 17 (15-19)
Hemibagrus
hemibagri Cultured K. Terengganu, Terengganu
27
Hemibagrus
nemurus Wild Tasik Kenyir,
Terengganu
5
Henneguya basifilamentalis Molnár et al., 2006b
Basal crypts of hemibranchia
SP wide, tapering anteriorly & posteriorly, 14.06(13-15) x 6.91(6-7.5) x 4.81(4-5);
elongated PC of different sizes, larger PC 6.52 (5-7)x2.81 (2-3), smaller PC 5.08(4-
5.6)x2.6(1.3-2); very thin CP, 26-38
Hemibagrus
nemurus Cultured K. Terengganu, Terengganu
6
Henneguya daoudi Székely et al., 2009a
Gills
SP elongated, 12.3 (11.4-12.8) x 7.3(6.7-7.6) x 5.3(4.7-5.7); elongated PC 5.9(5.2-6.2) x 2.7(2.4-3.3); straight or slightly bent caudal appendages CP 29.4 (27-31)
Trichogaster
trichopterus Cultured Macang, Kelantan
7
Hennegoides pangasii Molnár et al., 2006a
Gill filaments
SP asymmetric in frontal view, one side is slightly vaulted while the other is strongly vaulted, 27.3 (24–30) x 12.6 (12–13.8) x 11.6 (10.2–12); PC of different sizes, larger PC 13.9 (11.4–15) x 6.5 (6–7.2), smaller PC 11.5(10–
12.2) x 5.1 (4.8–6); CP 64.5(48–72)
Pangasius
hypophthalmus Cultured K. Terengganu, Terengganu
8
Hennegoides malayensis Molnár et al., 2006a
Gill rays
SP asymmetric in frontal view, one side of the spore body strongly vaulted; the other one is straight, 13.7(12.5–14.4) x 6.83 (6.4–7.5) x 5.1(4.2–6.3); anterior PC 5.66(5.1–6.2) x 3.1(3–
Pangasius
hypophthalmus Cultured K. Terengganu, Terengganu
28
3.3), posterior PC 5.95 (5.7–6.3) x 3.3(3.1–3.5);
CP 39.1(36–40.5)
9
Hennegoides berlandi Molnár et al., 2006a
Gill filaments
SP asymmetric in frontal view, one side of the spore strongly vaulted; the other one is straight, 8.5(8–9) x 2.8(2.5–3) x 2.98(2.8–3.2); PC 2.8(2.5–3) x 1.84(1.5–2); CP 43.6(40–50)
Pangasius
hypophthalmus Cultured K. Terengganu, Terengganu
10
Thelohanellus zahrahae Székely et al., 2009a
Gills
SP large, drop-shaped both in frontal and sutural view, tapered towards the anterior pole,
23.8(21.7-26.3) x 9.0(8.5-9.4) x 7.6(7.5-7.9); PC 9.9(7.9-10.8) x 6.3(5.3-6.6)
Barbonymus
gonionotus Cultured Machang, Kelantan
11
Myxobolus baskai Molnár et al., 2006a
Gills
SP ovoid in frontal view,rounded at the posterior end & slightly tapered at the anterior end, in lateral view the spores are lemon-shaped, 14.2(13.5–15) x 10.64(10.5–11) x 6.83(6.5–7.2);
PC 6.35(5.5–7) x 4.33(3.5–5);presence of caudal appendages on the posterior end of the spores
Pangasius
hypophthalmus Cultured K. Terengganu, Terengganu
12
Myxobolus pangasii Molnár et al., 2006a
Spleen
SP ovoid in frontal view, rounded at the posterior end & tapered
at the anterior end, in lateral view the spores are lemon-shaped, 14.3(13.5–15) x 7.03(6–7.8) x
Pangasius
hypophthalmus Cultured K. Terengganu, Terengganu
29
4.6(4–5); PC 6.42(6–7.5) x 1.68(1.4–2)
13
Myxobolus omari Székely et al., 2009a
Muscle cells
SP unique, larger dimensions in width than in length, frontal view-bluntly ellipsoidal, 7.9(7.2- 8.8) x 12(11-13.9) x 5.9(5.2-6.6); 2 nearly equal or somewhat differing polar capsules PC 5.9(4.4-6.6) x 4.7(4.0-5.3)
Pangasius
hypophthalmus Cultured K. Terengganu, Terengganu
14
Myxobolus leptobarbi Székely et al., 2009a
Muscle cells
SP oval frontal view, sharply tapering at the anterior poles, in
sutural view they were spindle-shaped, sharply tapering at the anterior end, 16(14.8-17) x 8.9(8.4-9.6) x 5.1(5.5-6.5); Larger PC 10.5(9.9- 11.5) x 3(2.3-3.6), smaller PC 9.9(8.8-10.6) x 3(2.3-3.6)
Leptobarbus
hoevenii Cultured Machang, Kelantan
15
Myxobolus terengganuensis Székely et al., 2009b
Muscle
SP ellipsoidal in frontal view, in sutural view- elongated ellipsoidal shape, 12.7(12-13.4) x 7.4(6.7-8.3) x 5(4.8-5.2); Larger PC 6.8(6.2-7.3) x 3.2(2.9-3.4), smaller PC 5(4.4-5.5) x 2.3(2.2- 2.6)
Osteochilus hasselti Wild Tasik Kenyir, Terengganu
16
Myxobolus tasikkenyirensis Székely et al.,
Muscle
SP oval in frontal view, tapering at the anterior poles, in sutural view-elongated ovals, tapering at both ends, 12.8(11.8-13.8) x 9.2(8.3-9.9) x
Osteochilus vittatus Wild Tasik Kenyir, Terengganu
30
2009b 6(5.5-6.5); PC 6.4(5.9-7.2) x 3(2.7-3.4)
17
Myxobolus csabai Székely et al., 2009b
Kidney
SP oval in frontal view, tapering at the anterior poles, in sutural
view, spindle-shaped, 12.9(12.1-14) x 8.6(8.2- 9.6) x 5(4.7-5.6); Larger PC 6.1(6.4-6.8) x 3.8(3.7-4.0), smaller PC 3.6(3.3-4.8) x 2.3(2.0- 2.9)
Osteochilus hasselti Wild Tasik Kenyir, Terengganu
18
Myxobolus osteochii Székely et al., 2009b
Kidney
SP oval in frontal view, tapering at the anterior poles, in sutural
view-lemon shaped, 13(12-14) x 8.7(8-9) x 5(4.5-5.5); PC 6.6(6-7) x 2.4(2-3)
Osteochilus hasselti Wild Tasik Kenyir, Terengganu
19
Myxobolus dykovae Székely et al., 2009b
Gill lamellae
SP oval both in frontal and sutural view, tapering at the anterior poles, 12(11-12.7) x 6.2(5.6-6.7) x 5.8(5.7-6.0); PC 6.0(5.9-6.1) x 2.1(2.0-2.3)
Barbonymus
schwanenfeldii Wild Tasik Kenyir, Terengganu
31 2.6 Myxospores Characterization Methods
The classical way of estimating the relationship between species is to compare their morphological characters. Taxonomy is still based largely on morphology. The increasingly available molecular information, such as nucleotide or amino acid sequences, and restriction fragment length polymorphisms (RFLPs), can also be used to infer phylogenetic relationships. In this study, we try to enhance our knowledge on the myxosporean on the bases of spores morphology and molecular identification. Through molecular biological data, phylogenetic assessment on identified-specimens could be performed.
2.6.1 Morphological Examination
Most myxozoans morphological-classification works in favor using the guidelines by Lom et Arthur (1989). Nevertheless, several works in recent years contributed other aids on myxozoan description (Lom etDyková 2006, Yokoyama et al. 2012). On the other hand, morphological characterization and differential diagnoses were most likely conducted on specimen to specimen bases, thus comparison may follow literature available for that particular specimen (Fig. 2, 3).
2.6.2 Genetic Information
The use of molecular data for inferring phylogenetic trees has now gained considerable interest among biologists of different disciplines, and it is often used in addition to morphological data to study relationships in further detail. When two sequences are compared, one can always calculate the percentage similarity by counting the number of identical nucleotides or amino acids, relative to the length of the sequence.
According to Hallett et al. (2002), while they were working on myxozoans, partial 18S rDNA analysis could greatly facilitated the process of species classification and clarification, and those authors recommend such an approach be incorporated into all taxonomic descriptions.
32
Figure 2. Diagrams of bivalvulid (A: frontal view, B: side view) and multivalvulid (C & E, top view, D:
side view) myxosporean spores. PC: polar capsule, SP: sporoplasm, SV: shell valve, SL: sutural line, L:
spore length, W: spore width, T: spore thickness, PCL: polar capsule length, PCW: polar capsule width (Adopted from Yokoyama et al. 2012).
Figure 3. Diagram of actinosporean spores. A: Triactinomyxon, B & C: Aurantiactinomyxon, D & E:
Neoactinomyxum, F & G: Tetractinomyxon.B, D & F: top views , C, E & G: side view. SB: spore body, LSB: length of spore body; WSB: width of spore body; S: style; LS: length of style; WS: width of style;
CP: caudal process; LCP: length of caudal process (regardless of curvature). LSCP: largest span of between the tips of the caudal processes; PC: polar capsule; DSB: diameter of spherical spore body (Adopted from Yokoyama et al. 2012)
33
There are parallel efforts in Europe, USA, and Japan to maintain public databases with all published nucleic acid sequences. One of the databases is the GenBank, which was operated and maintained at the NCBI (National Center for Biotechnology Information) in Bethesda, Maryland, USA. The sequences that are submitted since the last release are available as “daily updates”. Upon submission, authors or identifier (ID) will be given the accession number (AC) of their sequence isolate/s. The AC remains constant between releases and there exists an agreement between the managers of the three databases to give the same AC to the same sequence.
2.6.3 Phylogenetic Analysis
Phylogenetics has become a standard and increasingly powerful tool in evolutionary biology. Through understanding of molecular evolution and the molecular data collected, phylogenetics has now impinged upon almost every aspect of biological research (Littlewood, 2004). Prior to genetic information obtained, phylogenetic analysis could be conducted. Phylogenetic analysis establishes the relationships between genes or gene fragments, by inferring the common history of the genes or gene fragments. To achieve this, it is essential that homologous sites be compared with each other (positional homology). For this reason, the homologous sequences under investigation are aligned such that homologous sites form columns in the alignment. Obtaining the correct alignment for closely related species can be done manually. The more distantly related the sequences, the less straightforward it is to find the best alignment. Therefore, alignments are usually constructed with specific software packages that implement particular algorithms (details in Chapter 3: Materials & Methods).
34 CHAPTER III
MATERIALS & METHODS
3.1 Malaysian Myxosporean Collections
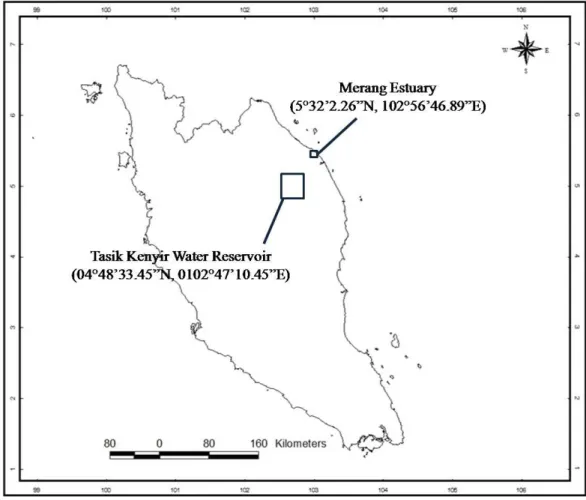
Samplings were conducted in freshwater and estuarine biotopes near Kuala Terengganu, Terengganu. Fish were collected with gill nets in the Tasik Kenyir Water Reservoir (04°48’33.45”N, 0102°47’10.45”E) and Merang Estuary (5°32’2.26”N, 102°56’46.89”E). Kenyir Lake freshwater reservoir is a forested upper water catchment area which drains into the main channel of the Terengganu River that meanders across the coastal lowland region before flowing into the South China Sea (Sultan et Shazili, 2010).
On the other hand, Merang estuary located in the north eastern coastline of Kuala Terengganu comprises of minimal mangrove line and mudflat area towards the shore.
Intertidal sandy mud beaches occur on both sides of the river mouth with open water channels. Small mangrove creeks presence throughout the estuary and this system experienced a maximum tidal range of about 0.2-1.8m.
Specimens of the fishes were transported to the laboratory of the Institute of Tropical Aquaculture (AKUATROP), University Malaysia Terengganu (UMT), and maintained in an aerated aquarium. Fish were freshly pithed, dissected and examined within two days post capture. Examinations for the presence of myxosporeans were carried out using a Leica EZ4 dissecting microscope and a Leica DM750 compound microscope at a different magnification appropriate for the analysis of parasites. The main organs of infection such as gills, fins, as well as muscle were examined thoroughly. Other organs, such as kidneys, gall bladder, and intestine, were also examined under a microscope. Infected organ/s with myxozoan plasmodia were observed with a Nikon model MM-800 microscope.
After examinations of hosts, plasmodia were carefully removed from tissues with a fine needle and dissected on a slide. Fresh preparations of ruptured plasmodia containing the spores were further studied by using a Nikon Model Eclipse 80i advanced light
35
microscope. Plasmodia in filaments, lamellae or in the gill arches were differentiated according to Molnár (2002) into epithelial, vascular, muscular and chondroid locations.
Thirty spores were measured according to the guidelines of Lom et Arthur (1989). Spores were preserved in 80% ethanol for molecular biology analysis. Voucher sample of myxosporean spores were deposited in the collection of the Zoological Department of the Hungarian Natural History Museum of Budapest, Hungary. Measurements and drawings of spores were compared to available literature reported from same hosts as well as to species that were congeners to on the phylogenetic tree.
Figure 4: Locations of myxozoa (myxospores) sampling in Malaysia