Pannon Egyetem, Molekuláris- és Nanotechnológiák Doktori Iskola
Hideg által indukált, fagytőrést és vernalizációt szabályozó folyamatok molekuláris hátterének
vizsgálata gabonafélékben
DOKTORI (PhD) ÉRTEKEZÉS
Vashegyi Ildikó Martonvásár, 2012
Témavezetı: Dr. Tóth Balázs
Magyar Tudományos Akadémia, Agrártudományi Kutatóközpont,
Mezıgazdasági Intézet
Hideg által indukált, fagytőrést és vernalizációt szabályozó folyamatok molekuláris hátterének vizsgálata gabonafélékben
Értekezés doktori (PhD) fokozat elnyerése érdekében
Írta:
Vashegyi Ildikó
Készült a Pannon Egyetem Molekuláris- és Nanotechnológiák Doktori Iskolája keretében.
Témavezetı: Dr. Tóth Balázs, PhD
Elfogadásra javaslom: igen / nem ………..
(aláírás)
A jelölt a doktori szigorlaton ... %-ot ért el,
Veszprém, 2010. december 21. ……….
a Szigorlati Bizottság elnöke
Az értekezést bírálóként elfogadásra javaslom:
Bíráló neve: Dr. Szabados László, DSc igen / nem
……….
(aláírás) Bíráló neve: Dr. Taller János, PhD igen / nem
……….
(aláírás)
A jelölt az értekezés nyilvános vitáján …... %-ot ért el.
Veszprém, 2012. ……….
a Bíráló Bizottság elnöke
A doktori (PhD) oklevél minısítése: …...
.………
az EDHT elnöke
TARTALOMJEGYZÉK
1. RÖVIDÍTÉSJEGYZÉK ...5
2. KIVONAT ...7
3. ABSTRACT...8
4. AUSZUG...9
5. BEVEZETÉS ÉS CÉLKITŐZÉSEK ...10
A TÉMA AKTUALITÁSA, JELENTİSÉGE...10
KÉRDÉSFELTEVÉS ÉS CÉLKITŐZÉSEK...11
6. IRODALMI ÁTTEKINTÉS ...14
A NÖVÉNYEKBEN HIDEGSTRESSZ HATÁSÁRA LEZAJLÓ FOLYAMATOK ÁTTEKINTÉSE...14
A HIDEGAKKLIMÁCIÓ ÉS A FAGYÁLLÓSÁG KIALAKULÁSÁT SZABÁLYOZÓ RENDSZER...16
A VERNALIZÁCIÓ ÉS A VIRÁGZÁS IDİZÍTÉSÉT SZABÁLYOZÓ RENDSZER KAPCSOLATA A FAGYÁLLÓSÁGGAL...20
A DIFFERENCIÁLATLAN, OSZTÓDÓ NÖVÉNYI SEJTEK ÉS SZEREPÜK A FAGYÁLLÓSÁG KIALAKÍTÁSÁBAN...23
7. ANYAGOK ÉS MÓDSZEREK ...26
AVRN1 VERNALIZÁCIÓS GÉN ÉS A CBF-COR RENDSZER KÖLCSÖNHATÁSÁNAK VIZSGÁLATA DELÉCIÓS MUTÁNS ALAKOR VONALBAN...26
A növénynevelés és az edzés körülményei ...26
Genotipizálás...27
A hajtáscsúcs fejlettségi állapotának meghatározása ...27
Real-time génexpressziós vizsgálatok (qRT-PCR) ...28
Fagyteszt ...29
Az ionkiáramlás mérése (relatív konduktancia meghatározása)...29
Az indukált fluoreszcencia intenzitásának mérése (Fv/Fm paraméter meghatározása)...30
ACBF-COR RENDSZER MŐKÖDÉSI SAJÁTOSSÁGAINAK VIZSGÁLATA DEDIFFERENCIÁLT ÁRPA SZÖVETTENYÉSZETEKBEN...31
A kallusz indukció és szövettenyésztés...31
A növénynevelés és az edzés körülményei ...31
A kalluszon végzett kísérletek beállítása és a szövettenyészetek edzése ...32
Fagyteszt kivitelezése kallusz kultúrákon ...32
Hosszú távú regenerációs teszt...33
Sejtes életképességi vizsgálat (TTC teszt)...33
Real-time génexpressziós vizsgálatok (qRT-PCR) ...33
Fitohormonok és Dicamba mennyiségi meghatározása ...34
Statisztikai elemzés ...34
8. EREDMÉNYEK ...36
AVRN1 VERNALIZÁCIÓS GÉN ÉS A CBF-COR RENDSZER KÖLCSÖNHATÁSÁNAK VIZSGÁLATA DELÉCIÓS MUTÁNS ALAKOR VONALBAN...36
ACBF-COR RENDSZER MŐKÖDÉSI SAJÁTOSSÁGAINAK VIZSGÁLATA DEDIFFERENCIÁLT ÁRPA SZÖVETTENYÉSZETEKBEN...42
9. MEGVITATÁS ÉS KÖVETKEZTETÉSEK ...52
AVRN1 VERNALIZÁCIÓS GÉN ÉS A CBF-COR RENDSZER KÖLCSÖNHATÁSÁNAK VIZSGÁLATA DELÉCIÓS MUTÁNS ALAKOR VONALBAN...52
ACBF-COR RENDSZER MŐKÖDÉSI SAJÁTOSSÁGAINAK VIZSGÁLATA DEDIFFERENCIÁLT ÁRPA SZÖVETTENYÉSZETEKBEN...57
10. ÖSSZEFOGLALÁS...63
AVRN1 VERNALIZÁCIÓS GÉN ÉS A CBF-COR RENDSZER KÖLCSÖNHATÁSÁNAK VIZSGÁLATA DELÉCIÓS MUTÁNS ALAKOR VONALBAN...63
ACBF-COR RENDSZER MŐKÖDÉSI SAJÁTOSSÁGAINAK VIZSGÁLATA DEDIFFERENCIÁLT ÁRPA SZÖVETTENYÉSZETEKBEN...64
11. ÚJ TUDOMÁNYOS EREDMÉNYEK (TÉZISEK) ...66
12. NEW RESULTS (THESES)...67
13. IRODALOMJEGYZÉK...68
14. PUBLIKÁCIÓS TEVÉKENYSÉG ...82
ADISSZERTÁCIÓBAN BEMUTATOTT TÉMÁKBAN MEGJELENT, NEMZETKÖZI TUDOMÁNYOS FOLYÓIRATOKBAN KÖZÖLT PUBLIKÁCIÓK...82
MÁS TÉMÁKBAN MEGJELENT, NEMZETKÖZI TUDOMÁNYOS FOLYÓIRATOKBAN KÖZÖLT PUBLIKÁCIÓK...82
KONFERENCIA KIADVÁNYOKBAN MEGJELENT ÖSSZEFOGLALÓK; ELİADÁSOK, POSZTEREK...82
IMPAKT FAKTORRAL NEM RENDELKEZİ FOLYÓIRATOKBAN MEGJELENT EGYÉB KÖZLEMÉNYEK...84
15. KÖSZÖNETNYILVÁNÍTÁS ...86
1. RÖVIDÍTÉSJEGYZÉK
2,4-D: 2,4-dikloro-fenoxi-ecetsav (IUPAC: (2,4-dichlorophenoxy) acetic acid)
ABA: abszcizinsav (IUPAC: (S)-5-(1-hydroxy-2,6,6-trimethyl-4-oxo-2-cyclohexen-1-yl)-3- methyl-2,4-pentanedienoic acid)
ABRE: ABA Responsive Element
ADA2: TRANSCRIPTIONAL ADAPTOR 2 AFP: Antifreeze Protein
AGLG1: AGAMOUS-LIKE GENE 1 AP1: APETALA 1
AP2/ERF: APETALA2 / Ethylene Responsive Factor APX1: Ascorbate Peroxidase 1
AREB/ABF: ABA-Responsive Element Binding Protein / Factor
BAP: 6-benzil-aminopurin (IUPAC: N-(phenylmethyl)-7H-purin-6-amine) CAL: CAULIFLOWER
CBF/DREB: C-Repeat Binding Factor / Dehydration-Responsive Element Binding Factor CBF: C-Repeat Binding Factor
CO: CONSTANS
COR: Cold-Regulated Gene
CRT/DRE: C-Repeat / Dehydration Responsive Element CYB5: CYTOCHROME B5
CYS: CYSTEINE PROTEINASE DHN: Dehydrin
Dic: Dicamba (IUPAC: 3,6-dichloro-2-methoxybenzoic acid)
DPA: dihidro-fazeinsav (IUPAC: (2Z,4E)-5-[(1R,3S,5R,8S)-3,8-dihydroxy-1,5-dimethyl-6- oxabicyclo[3.2.1]oct-8-yl]-3-methylpenta-2,4-dienoic acid)
EPS: Earlyness Per Se Gene
FR-1, -2: Frost Resistance (lókusz) 1, -2 FRU: FRUITFULL
FT: FLOWERING LOCUS T GI: GIGANTEA
HOS1: High Expression of Osmotically Responsive Genes 1
IAA: indol-3-ecetsav; auxin (IUPAC: 2-(1H-indol-3-yl) acetic acid)
IAA-Asp: IAA-aszpartát (IUPAC: 2-[[2-(1H-indol-3-yl)acetyl]amino] butanedioate) IAA-GE: IAA-glükóz észter
ICE: Inducer of CBF Expression
JA: jazmonsav (IUPAC: (1R,2R)-3-oxo-2-(2Z)-2-pentenyl-cyclopentaneacetic acid) LEA: Late Embryogenesis Abundant (Protein)
mvp: maintained vegetative phase (fenntartott vegetatív állapotú mutáns) MQ víz: MilliQ víz
MYB: Myelocytoblastosis Oncogene (transzkripciós faktor) MYBR: MYB Responsive Element
MYC: Myelocytomatosis Oncogene (transzkripciós faktor) MYCR: MYC Responsive Element
Neo-PA: neofazeinsav (IUPAC: (2Z,4E)-5-[(1S,3S,8S)-3,8-dihydroxy-1,5-dimethyl-6- oxabicyclo[3.2.1]octan-8-yl]-3-methylpenta-2,4-dienoate)
PA: fazeinsav (IUPAC: (2Z,4E)-5-[(1R,5R,8S)-8-hydroxy-1,5-dimethyl-3-oxo-6- oxabicyclo[3.2.1]octan-8-yl]-3-methylpenta-2,4-dienoic acid)
PGR: Plant Growth Regulator PHYC: PHYTOCHROME C PPD: Photoperiod Response Gene QTL: Quantitative Trait Locus
SA: szalicilsav (IUPAC: 2-hydroxybenzoic acid) SIZ1: SUMO E3 Ligase 1
TTC: 2,3,5-trifenil-tetrazólium-klorid (IUPAC: 2,3,5-triphenyl-2H-tetrazolium chloride) VRN: Vernalization Response Gene
WSOC1: Wheat Suppressor of Overexpression of CONSTANS 1 ZCCT1, -2: Zinc-Finger / CCT Domain Transcription Factor 1, -2
2. KIVONAT
Hideg által indukált, fagytőrést és vernalizációt szabályozó folyamatok molekuláris hátterének vizsgálata gabonafélékben
Deléciós mutáns alakor vonalon vizsgáltuk, hogy a VRN1 vernalizációs gén aktivitásának kiesése milyen hatással van a CBF-COR rendszer génexpressziós mőködésére, valamint ez hogyan befolyásolja a hideghatáshoz köthetı fenotípusos változásokat. Megerısítettük, hogy a VRN1 funkcionális hiánya megakadályozza a hajtáscsúcs vegetatívból generatív állapotba történı átmenetét. Megállapítottuk, hogy a CBF4, CBF9, CBF12, CBF14 és CBF17 gének, valamint a COR14b effektor gén hidegindukált aktivációjára ugyanakkor pozitív hatással van a VRN1 gén deléciója. Ezzel összhangban kimutattuk, hogy a mutáció növeli a fagyállóság mértékét és a vegetatív növényi részek ellenállóságát. A VRN1 gén tehát részt vesz annak a folyamatnak az aktiválásában, ami a vernalizáció elırehaladtával gátolja a hidegakklimáció kialakításáért felelıs szabályozó faktorokat.
İszi árpa merisztemoid kallusz tenyészetein vizsgáltuk, hogy sötétben nevelt, differenciálatlan sejtekben mik a CBF-COR rendszer alapvetı, tisztán hideghatáshoz köthetı mőködési sajátosságai. Ehhez kidolgoztunk egy új rendszert 'Nure' ıszi árpa speciális kallusz kultúráinak szövettenyésztésére, valamint fagytesztelésére. Kimutattuk, hogy a CBF9, CBF14 gének, valamint a COR14b effektor gén hidegindukált aktivációjának karakterisztikája hasonló csíranövények bokrosodási csomóiban, illetve kallusz tenyészetekben. A COR14b indukció szintje kalluszban alacsonyabb, de így is jelentıs mértékő volt. Ezzel összhangban megerısítettük, hogy a dedifferenciált szövettenyészetek hidegedzhetık. Megállapítottuk, hogy szintetikus auxinanalóg (Dicamba) használata kontroll körülmények között is növeli a stresszhormonok mennyiségét a kalluszokban, ezzel összhangban növeli a fagyállóság mértékét és a génexpressziós válasz erısségét, de nincs hatással a válasz megjelenésére és karakterisztikájára. Kimutattuk, hogy az inaktív abszcizinsav-katabolitok szintje hosszabb távú hidegedzés során jelentısen megemelkedett, ami korábbi abszcizinsav akkumulációra utal. A Dicamba alkalmazása esetén ez a válasz nagyobb mértékő volt, valamint a kezelés a megemelkedett fagyállóságot mutató kalluszokban a jazmonsav koncentrációját is növelte, a vizsgált endogén auxinformák hidegválaszára azonban nem volt számottevı hatással.
A fagyállóság kialakulását meghatározó CBF-COR rendszer hidegaktivációja tehát alapvetıen a differenciált sejtes állapottól, valamint a fényindukált és szisztémás jelátviteli útvonalaktól függetlenül zajlik le, ám a válasz maximális szintjének eléréséhez az említett folyamatokhoz köthetı faktorok is szükségesek. Intakt növényben a vernalizáció szabályozásában szerepet játszó gének mőködése is jelentıs hatással van a folyamatra.
3. ABSTRACT
Research on the molecular background of cold-induced processes regulating cold acclimation and vernalization requirement of cereals
Effects of loss of function of the VRN1 vernalization response gene on transcriptional activity of the CBF-COR system and phenotypical changes connected to the cold acclimation process were studied. Diploid einkorn wheat line carrying deletions encompassing VRN1 was used for the experiments. Inhibition of transition of the shoot apical meristem from vegetative to reproductive stage caused by disfunction of VRN1 gene was confirmed. However, the CBF4, CBF9, CBF12, CBF14 and CBF17 genes, as well as the COR14b effector gene showed higher cold-induced activity in homozygous mutant plants. In accordance with these findings mutation in homozygous form increased frost tolerance and cold resistance of vegetative parts of the shoot. According to this study the VRN1 activity is required for the initiation of the regulatory cascade that down-regulates the cold acclimation pathway.
Elemental cold response of the CBF-COR system was studied in dedifferentiated, meristemoid callus cultures grown in the dark at the gene expression, hormone content and freezing tolerance levels. For these purposes new protocols for tissue cultivation and testing frost tolerance were developed for winter barley line 'Nure'. The cold response of the CBF9 and CBF14 genes was very similar in the crowns of seedlings and in dark-grown callus cells, while COR14b induction was much lower than that observed in seedlings despite the similar characteristics of the response. The magnitude of cold induction of the CBF14 and COR14b genes and the accumulation of stress hormones showed Dicamba dependence in calli, which was correlated well with the higher cold acclimation ability in the presence of exogenous auxin. However, no significant effects of Dicamba were observed on the characteristics and induction of the response. At the hormonal level, two inactive catabolites of abscisic acid showed the highest level of cold response, which indicates a former abscisic acid accumulation. The increase was enhanced by Dicamba, as well as in the case of jasmonic acid. The endogenous auxin system showed only minor changes in response to cold and the effect of the exogenous analogue was marginal.
The results suggest the presence of a basal, cold-responsive activation mechanism of the CBF-COR system in dedifferentiated cells, which is independent of light-induced and systemic signals. However, these factors seem to be required for reaching the maximum level of activation. In intact plants, key regulator genes involved in the vernalization pathway play also an important role in regulation of cold acclimation process.
4. AUSZUG
Untersuchung des molekularischen Hintergrundes der Kalt-induzierten Frosttoleranz und Vernalisation regulierenden Prozesse in Getreidepflanzen
Der Einfluss des Ausfalles des VRN1 Vernalisationsgens auf die Wirkung des CBF-COR genexprimierten Systems wurde untersucht und wie es die zur Kalt verbundenen phänotypischen Alterungen beeinflusst. Es wurde bestätigt, dass der Mangel des VRN1 Gens den Durchgang der Sprossspitze aus vegetativ zu generativ behindert. Es wurde auch festgestellt, dass die VRN1 Gendeletion auf die Kalt-induzierte Aktivation der CBF4, CBF9, CBF12, CBF14 und CBF17 Gens, sowie das COR14b Effektorgen eine positive Wirkung hat.
Im Zusammenhang damit war es auch beweist, dass das Mass der Mutation die Frostbeständigkeit und den Widerstand der vegetativen Pflanzenteilen steigert. Also das VRN1 Gen nimmt Teil in der Aktivisierung des Prozesses, das bei der Vernalisation die für Kaltakklimatisation verantwortliche Faktoren verhindert.
Die zur Kaltwirkung verbundenen Eigenartigkeiten des CBF-COR Systems in undifferenzierten, im Dunkel wachsenen Zellen wurden in meristematische Zellgruppen in dem Kalluszuchten der Wintergersten untersucht. Dazu haben wir auch ein neues System für die Gewebezüchtung der Wintergerst Kulturen 'Nure' ausgearbeitet. Es wurde auch beweist, dass die Charakteristik der CBF9, CBF14 Gens, sowie die Kalt-induzierte Aktivation des COR14b Effektorgens im Halmbasen der Keimpflanzen und in Kalluszuchten ähnlich sind.
Der Induktionstand von COR14b war niedriger in calli, aber es besteht doch signifikant. Im Zusammenhang damit wurde es auch bestätigt, dass die undifferenzierten Gewebezuchten zur Kälte trainiert sein könnten. Es wurde auch festgestellt, dass der Gebrauch des syntätischen Analogon der Auxine (Diamba) unter kontrollierten Bedingungen die Menge der Stresshormone in calli auch steigerte. In Zusammenhang damit steigerte es auch die Stärke der Reaktion und die Frostbeständigkeit, aber hatte keine Wirkung auf die exogene Erscheinung und Charakteristik der Reaktion. Wir haben es auch beweist, das der Stand der inaktiven Kataboliten aus Abscisinsäure während lang dauerndes Kältetrainings signifikantisch steigert, was könnte auf frühere Abscisinsäure Akkumulation verweisen. Mit der Anwendung von Dicamba wurde diese Reaktion noch stärker, sowie die Behandlung die Konzentration der Jasmonsäure in calli mit grösserer Frostbeständigkeit steigerte, aber es hatte keine signifikante Wirkung an die Kaltreaktion der untersuchten endogen Auxineformen.
Die Kaltaktivation des CBF-COR Systems, das die Ausbildung der Frostbeständigkeit bestimmt, verläuft unabhängig von dem differenzierten Zellenstand und der Licht-induzierten und systematischen Signal Prozessen, aber die bennante Erscheinungen und die verbundene Faktoren auch nötig sind den maximalischen Stand der Reaktion zu erreichen. Ebenso die regulierende Gene in Vernalisation haben signifikantische Wirkung in intakter Pflanzen erreicht.
5. BEVEZETÉS ÉS CÉLKITŐZÉSEK
A téma aktualitása, jelentısége
A környezet hımérsékletének rövid- és hosszabb távú változása alapvetıen befolyásolja a növények növekedését, fejlıdését, terméshozamát és földrajzi elterjedését. Az alacsony hımérséklet káros hatásaival szembeni védekezés, azok kivédése, elkerülése ezért alapvetı fontosságú a növényi szervezet túlélése és terméshozama szempontjából. A globális klímaváltozás hatására kimutathatóan gyakoribbá váltak világszerte a hımérsékleti fluktuációk, a hirtelen felmelegedést követı gyors lehőlések, ami különösen a mérsékelt égövben hátrányosan befolyásolhatja a növények megfelelı fagyállóságának kialakulását (Gu és mtsai., 2008). Ez a jelenség új kihívások elé állítja a fejlett országok mezıgazdaságát is, ezért a mővelésbe vont növényfajok különbözı fajtáinak fagyállóságát kialakító és szabályozó folyamatok mélyebb megismerése, valamint ezen információk gyakorlati hasznosítása fontos feladat.
Termesztett gabonanövényeink, így például a búza, az árpa, a kukorica és a rizs a pázsitfőfélék (Poaceae) családjába tartoznak. A pázsitfőféléknek hatalmas ökológiai jelentıségük van. Bolygónk szárazföldi területeinek 30 %-át borítják füves puszták (préri, pampa, sztyeppe, füves szavanna), ahol a pázsitfőfélékhez tartozó növényfajok az uralkodók, de a trópusoktól a sarkkörökig, a tengerparti területektıl a magashegységekig elıforduló szinte valamennyi biomban megtalálhatók a fajaik. Fontos szerepet játszanak a csupasz földfelszín beerdısüléséhez vezetı folyamatban is, a szukcessziós sor korai, talajmegkötés szempontjából kulcsfontosságú társulásai általában füves társulások, valamint a nagytestő növényevı állatok és rágcsálók elsıdleges tápnövényei.
Az emberiség szempontjából szintén igen nagy jelentıséggel bírnak a pázsitfőfélék. A Föld népessége által betakarított növényi termények éves összmennyiségének 70 %-át, az emberiség éves kalóriafogyasztásának 50 %-át adják ezek a növények. Mintegy 300 fajt hasznosítanak világszerte, ebbıl 35 fajt domesztikáltak, nemesítenek is. Termesztésük 10 000 éves múltra tekint vissza, felhasználásuk nélkül nem jöhetett volna létre a neolitikus forradalom, melynek során a vadászó-győjtögetı életmódról a letelepedett, földmővelı életmódra való átállás mérföldkı volt a szociális és kulturális evolúciónk szempontjából is.
A növénycsoport sikerének titka egyrészt a családban kifejlıdött, számos elınyös evolúciós stratégiában rejlik. Ilyen a vegetatív szaporodási módok kiaknázása, például a tarackkal, gyöktörzzsel való gyors terjedés. Jellemzı rájuk a megtermékenyítési módok sokfélesége: általában szélbeporzásúak, de képesek az önbeporzásra, valamint elıfordul körükben az apomixis jelensége is, vagyis hogy a petesejtbıl megtermékenyítés nélkül fejlıdik ki az embrió, alakul ki a mag. Elterjedésüket segíti a hatékony magszórás és különféle magterjesztı készülékek fejlesztése, valamint a gyors egyedfejlıdési ciklus (a legtöbb pázsitfőféle egynyári növény). Könnyő, de teherbíró felépítésük szintén segíti túlélésüket. Ezt a nóduszos szerkezet és a fejlett szklerenchima teszi lehetıvé. Erre a legjobb példát a bambuszok jelentik, melyek bizonyos fajai DK-Ázsiában fa méretőre megnıve akár nagyobb ligeteket, erdıket is alkothatnak.
A pázsitfőfélék sikerének másik titka, hogy biokémiai, fiziológiai védekezési mechanizmusokkal hatékonyan tudnak fellépni a káros környezeti hatások ellen. A növények alapvetıen kétféle módon tudnak alkalmazkodni a környezeti kihívásokhoz. Az egyik mód az adaptáció, a génkészlet egyedekre ható szelekciós nyomás általi optimalizálása populációs
szinten, ami evolúciós idıskálán megszerzett, elınyös tulajdonságkombinációk kialakulását, elterjedését jelenti. Trópusi területeken honos pázsitfőféléknél például a fotoszintetikus mőködés környezeti feltételekhez történı hangolásának hatékony módszere a C4-es fotoszintetikus út, valamint az ahhoz szükséges szöveti felépítés (Kranz anatómia) kialakítása.
Lényeges adaptációs jelenség az is, hogy a gabonafélék hajtáscsúcsi merisztémája, ami a földfelszín feletti, többek között a fotoszintézis fontos feladatát végzı vegetatív, valamint a generatív növényi részek létrehozásáért felelıs, vegetatív állapotban a gyökérnyaknál, a földfelszín alá rejtve található, így védelmet élvez a tőz, az elfagyás és a növényevı állatok kártételével szemben.
A másik alapvetı módszer, amivel a növények felvehetik a harcot a káros környezeti hatások ellen az egyedi alkalmazkodás. Ha egy konkrét környezeti tényezıhöz (pl. alacsony hımérséklet) történı alkalmazkodásról beszélünk, ezt a jelenséget akklimációnak, ha komplex környezeti hatásokhoz (szimultán fellépı, szuboptimális körülményekhez - pl. aszálykor megjelenı betegségekhez, vízhiányhoz és hıstresszhez) együttesen történı alkalmazkodásról beszélünk, akkor akklimatizációnak hívjuk. A télállóság tehát akklimatizáció eredménye: azt a tulajdonsághalmazt értjük alatta, melynek segítségével a növények képesek túlélni a téli, kedvezıtlen idıszak káros hatásait (jégképzıdés, elfagyás, oxigénhiány, vízhiány, betegségek). Ennek egyik komponense a fagyállóság, ami a hidegakklimáció következménye, és a növény fagypont alatti hımérsékleten való túlélését biztosítja. Természetesen ez is komplex genetikai, biokémai, fiziológiai változások összességét jelenti, hiszen magának az alacsony hımérsékletnek is számos, különbözı biológiai szinteken jelentkezı hatása van. Az akklimáció során a növények a genetikai állományuk által lehetıvé tett kereteken belül változtatják meg úgy az anyagcseréjüket, hogy az - például védıanyagok termelésével - segítse ıket a védekezésben. Ez alapvetıen sejt szintő alkalmazkodást jelent. A másik, egyed szintő alkalmazkodási módszer, hogy szisztémás jelek hatására a növény az egyedfejlıdését, szaporodási ciklusát igazítja úgy, hogy a lehetı legellenállóbb életszakaszban vészelhesse át a kedvezıtlen idıszakot. Ennek érdekében például egyes növények csak meghatározott idıtartamú hideghatást követıen virágoznak (ezt nevezik vernalizációs igénynek). A növények a vernalizáció következtében, a generatív állapotba történı átmenetet követıen azonban elveszítik korábban megszerzett fagyállóságukat (Fowler és mtsai., 1996a; Fowler és Limin, 2004; Limin és Fowler, 2006). Ez alapján, valamint annak a ténynek az ismeretében, hogy mind a vernalizáció, mind a hidegakklimáció adott idıtartamú hideg periódus hatására következik be, feltételezhetı, hogy a két rendszer között kölcsönös regulációs kapcsolat van, ám ennek a kapcsolatnak a részletei még nem ismertek teljesen.
Kérdésfeltevés és célkitőzések
A különbözı biokémiai, fiziológiai mőködések hatékonyságának optimalizálása, valamint azok idızítésének és intenzitásának környezeti feltételekhez történı igazítása érdekében a növények tehát összehangolják a különbözı környezeti jelekre, valamint az ugyanarra a külsı stimulusra adott, de különbözı célú válaszaikat. Mivel ezt az összetett folyamatot egy bonyolult kölcsönhatásokból felépülı szabályozó rendszer irányítja, annak mőködését a jelenleg rendelkezésünkre álló eszközökkel, módszerekkel csak részleteiben tudjuk szakszerően vizsgálni, s a teljes folyamatra csak a részeredmények alapján kialakított elméleti modellek engednek rálátást. Az ilyen típusú kutatómunka során viszont nagyon lényeges, hogy több aspektusból is tanulmányozzuk a jelenséget. Munkánk során a meglévı ismereteink ellenırzése, kiegészítése céljából két különbözı megközelítésbıl vizsgáltuk a hideg által indukált, fagytőrést és vernalizációt szabályozó folyamatok molekuláris hátterét.
Gabonafélék vernalizációs és hidegakklimációs rendszerének mőködésével kapcsolatban viszonylag széleskörő a tudásunk, azonban e két, hideghatáshoz köthetı, ám különbözı eredıjő folyamat kapcsolatáról, a bennük szerepet játszó szabályozó faktorok közti kölcsönhatásokról már kevesebb információ áll rendelkezésünkre. Célunk tehát az volt, hogy speciális genetikai anyagon végzett vizsgálatokkal teszteljük a gabonafélék hidegstresszre adott molekuláris válaszreakcióiról kialakított modellek, hipotézisek helyességét, valamint kiegészítsük a hidegakklimációs és vernalizációs folyamatok szabályozásában szerepet játszó transzkripciós faktorok közti kölcsönhatásokról meglévı tudásunkat. Ennek érdekében tesztkísérleteknek vetettük alá az alakor (Triticum monococcum L.) MVP2 vonalat (mvp – maintained vegetative phase: fenntartott vegetatív állapotú mutáns), ami a VRN1 merisztéma identifikációs faktor génjét érintı deléciót tartalmaz. A kísérleti alany kiválasztását az indokolta, hogy a Triticum nemzetségbe tartozó, diploid kromoszómakészlető alakor (Triticum monococcum, L.; 2n=2x=14; genom: AmAm) és a nagy gazdasági jelentıségő, de bonyolultabb genomfelépítéső, allohexaploid kenyérbúza (Triticum aestivum, L.; 2n=6x=42;
genom: AABBDD) A kromoszómaszerelvénye, mely a fagyállóságot befolyásoló legfontosabb szabályozó géneket hordozza, ugyanattól a diploid ıstıl származik. Az alakor növények vizsgálata során kapott eredményekbıl tehát kenyérbúzára vonatkoztatva is fontos következtetéseket vonhatunk le a fagytőrés kialakulásának szabályozásában szerepet játszó molekuláris genetikai folyamatokról. A kísérletek során azt vizsgálatuk, hogy a VRN1 vernalizációs gén aktivitásának kiesése milyen hatással van a hidegakklimációs folyamatok szabályozásában kulcsszerepet játszó CBF-COR rendszer mőködésére, valamint hogy a vernalizációs gént érintı mutáció hogyan befolyásolja a hideghatáshoz köthetı (vernalizációs és akklimációs folyamatokkal kapcsolatos) fenotípusos változásokat. Ennek érdekében a következı feladatok elvégzését tőztük ki célul:
• Genotipizálás segítségével MVP2 mutáns vonal szegregáló populációjából származó növények közül a mutációt homozigóta recesszív formában hordozó (mvp/mvp) egyedek elkülönítése a vad allélt legalább egy kópiában hordozó (Mvp/-) egyedektıl.
• A genotipizálás alapján kialakított csoportokban a hajtáscsúcs fejlettségi állapotának meghatározása, valamint a VRN1 gén aktivitásának ellenırzése az elınevelés, illetve az edzés során.
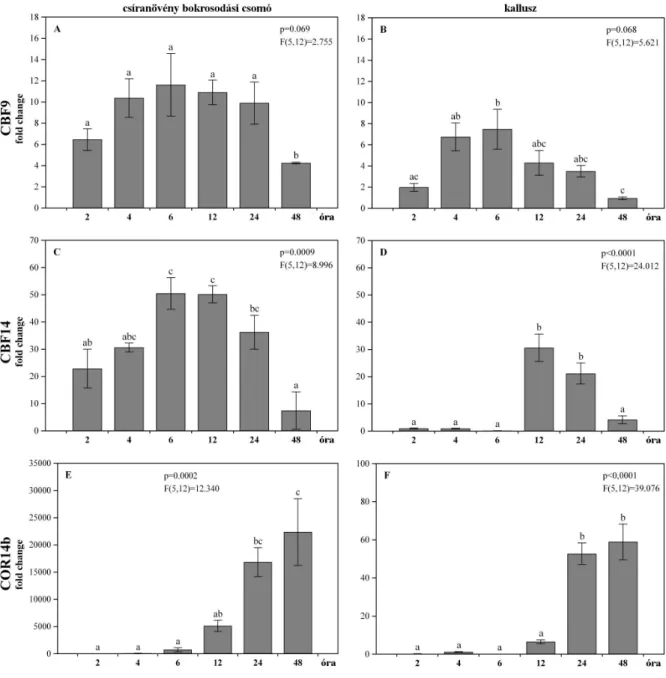
• A két csoportban az FR-2 lókuszhoz kapcsolható CBF gének, valamint a COR14b gén hidegindukált aktivációjának, illetve utóbbi, késıbb indukálódó effektor gén esetében annak idıbeli változásának vizsgálata.
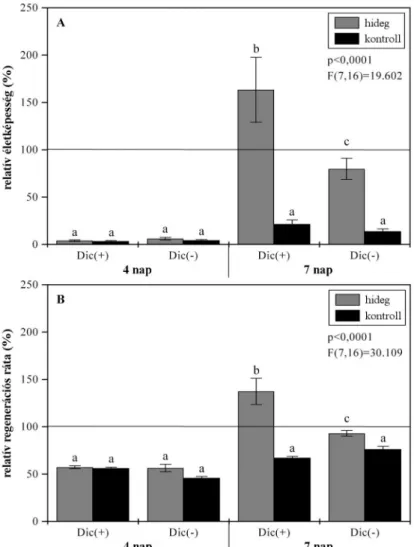
• Fagytesztek kivitelezésével az akklimációs folyamatok következtében kialakuló fagyállóság mértékének meghatározása a különbözı genetikai hátterő csoportokban.
• A növények általános fiziológiai állapotát érintı, hideg hatására bekövetkezı membránkárosodások következményeinek monitorozása az elınevelés, az edzés, valamint a fagyteszt során a levelek ionkiáramlásának, valamint a fotoszintetikus elektrontranszportlánc mőködési hatékonyságának mérésével.
A hidegindukált jelátviteli folyamatokkal kapcsolatos tudásunk hiányosságai miatt viszonylag kevés információ áll rendelkezésünkre azzal kapcsolatban is, hogy gabonafélékben a hidegstressz hatására meginduló akklimációs folyamatok szabályozásának mik az általános, tisztán hideghatáshoz köthetı molekuláris alapjai. Különösen fontos ez a kérdés a növény fagyállóságának legfontosabb komponensét, a regenerációs képességet meghatározó, elsısorban a hajtáscsúcsban elhelyezkedı, differenciálatlan osztódó szövetek esetében. Annak érdekében, hogy hozzájárulhassunk e terület felderítéséhez, a hidegakklimációval foglalkozó kutatások során általánosan használt intakt növényi rendszerektıl eltérı, speciális kísérleti
objektumon, a szisztémás és fényindukált jelátviteli kölcsönhatások kizárásával vizsgáltuk a CBF-COR rendszer alapvetı mőködési sajátosságait. Ehhez fagytőrı ıszi árpa (Hordeum vulgare L. cv. Nure; 2n=2x=14; genom: HH) embriogén kallusz kultúrák dedifferenciált, totipotens merisztemoid sejtjeinek tenyészeteit használtuk fel. A kísérleti alany kiválasztását egyrészt annak diploid genomfelépítése indokolta, ami egyszerőbbé teszi a vizsgált folyamatok genetikai hátterének felderítését. Emiatt az árpát - mint a Triticeae nemzetségcsoportba tartozó, termesztett gabonafélék közül a kenyérbúza legközelebbi rokonát – a távolabbi rokon Brachypodium distachyon helyett gyakran használják a gabonafélék modellnövényeként molekuláris genetikai vizsgálatoknál. Használatát az is indokolta, hogy szövettenyészeti rendszerekben jól fenntartható, könnyen kezelhetı faj.
Munkánk során a transzkripciós faktor gének expressziós profiljának meghatározása mellett célunk volt a hidegakklimáció fenotípusos változásokban megnyilvánuló jeleinek vizsgálata is: az esetlegesen kialakuló fagyállóság mértékének, valamint az akklimáció folyamatát befolyásoló, illetve azt indikáló endogén hormonok koncentrációjában bekövetkezı változások meghatározása. Ennek érdekében a következı feladatok elvégzését tőztük ki célul:
• İszi árpa dedifferenciált kallusz tenyészetébıl kialakított merisztemoid kultúrák szövettenyésztési rendszerének kidolgozása.
• Az elıkísérletek alapján kiválaszott CBF gének és a COR14b effektor gén expressziós profiljának felvétele hidegedzés során; az expressziós kinetika összehasonlítása intakt növényi és dedifferenciált szövettenyészeti rendszerekben.
• Rövidebb és hosszabb ideig tartó hidegkezelés hatásának vizsgálata a fent meghatározott mintázatra a szövettenyésztés során használt, szintetikus auxinanalóg alkalmazásának függvényében.
• Fagytesztek beállítása; a speciális objektum egyes fagytőrési komponenseinek (sejtes életképesség, tömegnövekedési ráta) méréséhez kellı módszertani beállítások elvégzése.
A tenyészetek hidegakklimáció hatására kialakuló fagytőrésének meghatározása a szövettenyésztés során használt, szintetikus auxinanalóg alkalmazásának függvényében.
• A tenyészetekben található endogén auxin, annak legfontosabb származékai, valamint a hideghatásra adott válaszreakciók kialakításában szerepet játszó stresszhormonok edzés hatására bekövetkezı koncentrációváltozásának vizsgálata a szövettenyésztés során használt, szintetikus auxinanalóg alkalmazásának függvényében.
6. IRODALMI ÁTTEKINTÉS
A növényekben hidegstressz hatására lezajló folyamatok áttekintése
A növényeket természetes környezetükben kétféle alacsony hımérsékleti stressz érheti: a molekuláris biológiai folyamatok mőködési hımérséklete (~25 °C) szempontjából szuboptimális, de fagypont feletti hımérsékletnek (<20 °C), illetve a fagypont alatti hımérsékletnek (<0 °C) való kitettség. Utóbbi az inter- és intracelluláris jégképzıdés, valamint a membránok és fehérjék funkcióromlással járó szerkezeti átalakulása miatt komoly károkat okozhat a növényi szervezetben, valamint a talajvíz megfagyásának elıidézésével gátolja a víz- és tápanyagfelvételt. Ezeken a folyamatokon keresztül ozmotikus és oxidatív stresszt, dehidratációt is okozhat a növényben, emellett rontja a fotoszintetikus mőködés és a transpiráció hatásfokát. Azonban a külsı hımérséklet csökkenésének érzékelése lehetıvé teszi olyan molekuláris védekezési mechanizmusok aktiválódását még a visszafordíthatatlan károkat okozó, fagypont alatti hımérséklet elérése elıtt, melyek segítségével a növény anyagcseréjének és fejlıdési folyamatainak átprogramozásával felkészülhet az alacsony hımérsékleti stressz hatásainak kivédésére, kikerülésére. Ez a folyamat a hidegakklimáció. A fejlıdési ritmus, a vegetatívból a generatív állapotba történı átmenet hımérsékleti tényezıktıl függı idızítését gabonafélékben pedig a vernalizáció folyamata garantálja, melynek során bizonyos növényfajokban, genotípusokban a virágzás csak hosszabb-rövidebb hideg periódus hatására következik be, biztosítva ezzel, hogy a hidegre rendkívül érzékeny generatív forma ne a kedvezıtlen idıszak alatt alakuljon ki.
Az elsıdleges jelet az alacsony hımérséklet által aktivált jelátviteli folyamatban valószínőleg a sejtmembrán fizikai tulajdonságainak hideg által okozott megváltozásai szolgáltatják (a membránfluiditás csökken, a membrán rigidebbé válik, ami bizonyos enzimek aktivitásának megváltozását okozza, valamint közvetlenül is befolyásolja a fehérjék konformációját). A sejtmembránban lezajló fizikai változások jelét érzékelı receptormolekula még mindig ismeretlen, feltehetıen a membránokhoz kapcsolódó hisztidin-kinázok vagy kalcium csatornák látják el ezt a feladatot (Monroy és Dhindsa, 1995). Ezen kívül a jégkristályok képzıdése ozmotikus és az esetleges sejtkárosodás miatt oxidatív stresszt is elıidéz a növényben, ami szintén szignálként is funkcionáló molekulák (pl. reaktív oxigéngyökök) felszabadulását eredményezi (összefoglalva: Van Breusegem és mtsai., 2001;
Foyer és Noctor, 2009).
A másodlagos, immár kémiai jelet a hideg-stressz molekuláris hatásai miatt képzıdı, illetve felszabaduló reaktív oxigéngyökök és szignál lipidek (pl. foszfatidsav), valamint a Ca2+-csatornák mőködésének megváltozása miatt bekövetkezı intracelluáris Ca2+- koncentráció-emelkedés alkotja. Ezen kívül stressz-hormonok (pl. abszcizinsav, jazmonsav) is felszabadulnak, amik egyrészt szintén jeltovábbító szerepet töltenek be, másrészt az intercelluláris térbe és a szállítórendszerbe lépve segítik a jel – biotikus stresszekkel szembeni rezisztencia kialakulásában már felismert jelentıségő – szisztémás elterjedését (összefoglalva:
Pieterse és van Loon, 1999; Shah, 2009). A jel felerısítését a Ca2+ belsı raktárakból (endoplazmatikus retikulumból), illetve az extracelluláris térbıl a citoszolikus térbe történı áramlását segítı folyamatok (Ca-burst kialakítása), valamint a kináz kaszkádok aktivációja segíti (Kacperska, 2004; Chinnusamy és mtsai., 2004). Ezek a változások transzkripciós faktorok aktiválódását eredményezik a sejtmagban, melyek további, szabályozó szerepet betöltı kulcsgéneket kapcsolnak be, melyek mőködése a génexpressziós mintázat megváltozását eredményezi. Ez de novo fehérjeszintézis, illetve poszt-transzkripciós és poszt- transzlációs fehérjemódosítások révén a fehérjemőködés megváltozását, enzimek
aktiválódását, inaktiválódását okozza, végeredményében a metabolom, a sejt anyagcseréjének átrendezıdését eredményezi.
Alapvetıen két csoportba lehet osztani a növényben hideg hatására lezajló molekuláris eseményeket: egyrészt a fagyállóság növelését célzó változásokra, másrészt az egyedfejlıdési ciklus stádiumainak idızítését szabályozó változásokra. Mindkettı a növény túlélését és a szaporodás hatékonyságának növelését segíti elı. A fagyállóság növelését, a hideg-stressz hatásainak kivédését krioprotektánsok (pl. poliszacharidok, prolin, AFP (Antifreeze Protein)) felhalmozódása, az antioxidáns és ozmoregulációs rendszer aktiválása, a membrán lipidösszetételének fluiditást növelı megváltozása és a fehérjék szerkezetét védı hısokkproteinek, LEA (Late Embryogenesis Abundant) fehérjék, chaperonok akkumulációja segíti elı. A generatív szaporító szervek kialakulásának idızítését a hajtáscsúcsban a vernalizációs rendszeren keresztül, a sejtosztódás és sejtdifferenciáció szabályozásán át hajtja végre a növény. A hormonális rendszer megváltozása mindkét folyamatban szerepet játszik:
egyrészt a fotoszintetikus aktivitást is befolyásoló, szisztémás jelátviteli rendszer részeként (abszcizinsav, jazmonsav), másrészt a sejtosztódást, sejtmegnyúlást és -differenciálódást szabályozó hatásával a növekedés, fejlıdés ütemét a megváltozott környezeti adottságoknak megfelelıen alakítja, a hideg hatására aktiválódó, szintén igen energiaigényes védekezési folyamatokkal összehangolja.
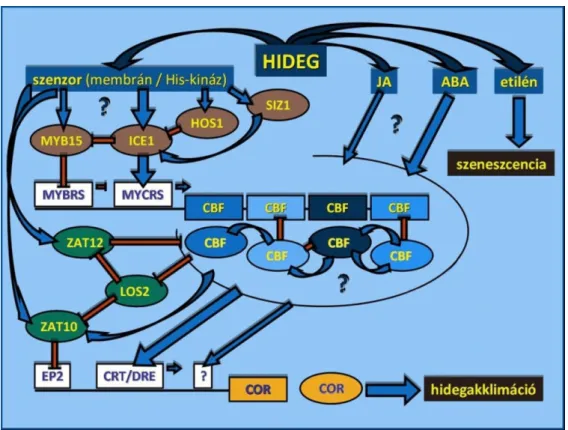
(A növényekben hideg hatására aktiválódó, általános molekuláris folyamatokat az 1. Ábra foglalja össze.)
1. Ábra Az alacsony hımérsékleti stressz hatására aktiválódó molekuláris folyamatok kölcsönhatási sémája. (A kérdıjelekkel jelölt folyamatok, kölcsönhatások még nem teljesen ismertek, feltérképezésük jelenleg is folyik.) (A Szerzı ábrája)
Ám a fagyállóság növelését és a virágzás ütemezését szabályozó mechanizmusok nem csak egymással állnak folyamatos, nagyon összetett kölcsönhatásban, hanem egyéb, az anyagcsere aktivitását befolyásoló környezeti tényezıket érzékelı, illetve azok szabályozása alatt álló
rendszerekkel is. Ilyen a fotoperiodizmus és a cirkadián óra rendszere, a vízháztartás és a tápanyagfelvétel szabályozó rendszere, a fényintenzitásra és fényminıségre reagáló rendszer, valamint a biotikus stresszek és a sebzıdés hatására aktiválódó rendszer. Tehát a növény ténylegesen mutatott megjelenési formáját, a fenotípust a genom és a környezeti tényezık között folyamatosan mőködı, komplex interakciók „konklúziója” alakítja ki. A ténylegesen megfigyelhetı fenotípusos tulajdonságok, például a növény növekedési üteme, a vegetatívból a generatív állapotba történı átmenet idızítése, az elért fagytőrés mértéke mind ezen folyamatok kölcsönhatásainak az eredıi.
A hidegakklimáció és a fagyállóság kialakulását szabályozó rendszer
A mérsékelt éghajlati övben a termesztett, ıszi gabonafélék télállósága és megfelelı terméshozama szempontjából nélkülözhetetlen az optimális fagyállóság elérése. A növény genetikai háttere által lehetıvé tett, maximális fagyállóság a hidegakklimációs folyamatok következtében alakul ki (Takahashi és Yasuda, 1971; Levitt, 1980). Ennek hiányában a gabonanövények már -3 °C és -5 °C közötti hımérséklet hatására kifagynak, míg a genotípustól függı maximális fagyállóságot elérve akár -20 és -30 °C közötti fagyot is túlélhetnek (Limin és Fowler, 1985). A megszerzett fagyállóság mértéke függ a hidegakklimáció idıtartamától (Fowler és mtsai., 1999a), valamint a hımérséklettıl: 10 °C és 0 °C között fordított arányosság figyelhetı meg az akklimáció mértéke és a kezelési hımérséklet között (Paulsen, 1968; Gusta és mtsai., 1982a).
Az akklimáció során a süllyedı, de még fagypont feletti hımérséklet hatására gyors jelátviteli útvonalak aktiválódnak a növényben, melyek transzkripciós faktorok és egyéb, hideg által szabályozott kifejezıdéső gének aktivációjához vezetnek, amik megnövelt fagyállósági szintet eredményeznek (Thomashow, 1999). Ez az összetett válasz Arabidopsis modellnövényen végzett kutatások alapján alapvetıen két rendszerre osztható attól függıen, hogy az abszcizinsav (ABA) részt vesz –e a jelátviteli folyamatokban, vagy sem (Gilmour és Thomashow, 1991; Shinozaki és Yamaguchi-Shinozaki, 2000; Shinozaki és mtsai., 2003). A legfontosabb szerepet ebben a folyamatban az ICE-CBF-COR rendszernek tulajdonítják (ICE:
Inducer of CBF Expression, CBF: C-Repeat Binding Factor, COR: Cold-Regulated Gene), amit az abszcizinsavtól független jelátviteli út részének gondolnak, és megfigyelték, hogy hideg hatására azonnali, gyors választ produkál (Chinnusamy és mtsai., 2003).
Az elsı lókusz, amit gabonafélékben a fagyállóság kialakításával kapcsolatban azonosítottak, az 5A kromoszómán elhelyezkedı FR-1 (Frost Resistance 1) lókusz volt (Sutka és Snape, 1989; Sutka és mtsai., 1999; Galiba és mtsai., 1995). Kobayashi és munkatársai (2005) azt feltételezik, hogy a Vrn-A1 allélhoz szorosan kapcsolt Fr-A1 allél játsza a fı szerepet a CBF-ek által közvetített COR/LEA aktivációban. Ám mivel ez a lókusz a legtöbb tanulmányban a VRN1 génnel közösen szegregál (Roberts, 1990; Hayes és mtsai., 1993;
Galiba és mtsai., 1995; Francia és mtsai., 2004; Galiba és mtsai, 2009), az sem kizárható, hogy az FR-1 lókuszon nem független gén van jelen, hanem csupán a VRN1 gén pleiotróp hatását mutatták ki a vizsgálatok.
A fagyállóság különbözı búza és árpa genotípusok között megfigyelhetı, természetes variációjának háttereként térképezett másik lókusz az FR-2 lókusz. Ez szintén az 5-ös homeológ csoport kromoszómáinak hosszúkarján helyezkedik el, a VRN1 géntıl proximális irányban kb. 30 cM távolságra. Ez a lókusz a rizs 5 tagú klaszteréhez képest a közelebbi rokon búzában és árpában a CBF gének 11-15 tagú klaszterét foglalja magában (Vágújfalvi és mtsai., 2003; Francia és mtsai., 2004, 2007; Miller és mtsai., 2006; Skinner és mtsai., 2006, Knox és mtsai., 2010). A klaszteren belül bizonyos CBF-ek csak erre a csoportra jellemzık
(Badawi és mtsai., 2007). Árpa és diploid búza (alakor) összehasonlításával megállapították, hogy a gének sorrendje a klaszteren belül konzervált (Miller és mtsai., 2006), tehát ez a génstruktúra a csoport közös evolúciós vívmánya, ami a szabályozó gének számának növelésével segíthette nagyon eltérı éghajlatú területek kolonizálását. Ez a szorosan kapcsolt régió több búza és árpa térképezési populáció vizsgálata alapján a fagyállóság mértékét meghatározó fı QTL-ként (Quantitative Trait Locus) azonosítható (Vágújfalvi és mtsai., 2003; Francia és mtsai., 2004, 2007; Båga és mtsai., 2007). A rendszer hideg hatására bekövetkezı, gyors, kontrollált aktivációja egyértelmő kapcsolatba hozható a fagyállóság emelkedésével (Kobayashi és mtsai., 2005; Kume és mtsai., 2005; Vágújfalvi és mtsai., 2005). A CBF gének szerepét részletesen Arabidopsis thaliana modellnövényben vizsgálták (Jaglo-Ottosen és mtsai., 1998; Chinnusamy és mtsai., 2003), valamint a szekvencia- homológiák alapján hasonló géneket azonosítottak árpában is (Choi és mtsai., 2002; Skinner és mtsai., 2005). Árpában 17 különbözı CBF transzkripciós faktort fedeztek fel (Skinner és mtsai., 2005; Francia és mtsai., 2007), több közülük már pár órás hidegkezelés hatására is bekövetkezı aktivációt mutatott (Jaglo és mtsai., 2001; Shen és mtsai., 2003; Stockinger és mtsai., 2007). Bár a pázsitfőfélék összehasonlító térképezési adatai alapján a gabonafélék genomjai között nagymértékő makrohomológia figyelhetı meg (Gale és Devos, 1998), s azonosítottak is általánosan elkülöníthetı CBF klasztereket (Miller és mtsai., 2006), azonban az azonosított CBF-ek egyedi szerepe még mindig nem bizonyított. A különbözı fajokban (Jaglo és mtsai., 2001; Campoli és mtsai., 2009), valamint egy fajon belül is a különbözı genotípusokban (Grossi és mtsai., 1998; Kume és mtsai., 2005; Stockinger és mtsai., 2007) ugyanis különbözı mértékő az aktivációjuk, illetve más expressziós profilt mutatnak.
Mőködésük bonyolult rendszerét tovább árnyalja, hogy bizonyos CBF gének alacsony szinten stresszmentes körülmények között is expresszálódnak, valamint expressziójuk a generatív átmenetet követıen is megfigyelhetı, ami a géntermék lassú, egyenletes akkumulációjához, s egy fajra jellemzı, alapszintő hidegtőrés kialakulásához vezethet. Ebbıl következhet, hogy – bár fagytőrésük jelentısen kisebb a vegetatív állapotban lévı, hidegedzett, ıszi genotípusú növényekéhez képest – a tavaszi genotípusú, valamint a virágzó növények is rendelkeznek bizonyos mértékő hidegtőréssel. (Ezeknek a géneknek a szabályozásában például a fajra jellemzı fejlıdési ütemet meghatározó koraiság gének játszhatnak szerepet.) Más CBF gének csak stressz hatására, de akkor gyorsan, fokozott mértékben szólalnak meg, majd aktivitásuk lecseng, s ha a hideghatás továbbra is fennáll, akkor alacsonyabb szinten, diurnális ciklus szerint expresszálódnak (Vágújfalvi és mtsai., 2005; Stockinger és mtsai., 2007; Badawi és mtsai., 2007). Utóbbiak szabályozzák valószínőleg a hidegakklimációs folyamatok révén szerzett fagyállóság kialakulását.
Maguk a CBF transzkripciós faktorok a CBF regulonhoz tartozó gének upstream szabályozó régióiban található CRT/DRE (C-Repeat / Dehydration Responsive Elements) cisz-ható elemekhez kapcsolódva serkentik a gének kifejezıdését (Baker és mtsai., 1994;
Stockinger és mtsai., 1997; Thomashow, 1999). Megfigyelték, hogy Arabidopsisban a hideg hatására megváltozott aktivitást mutató gének mintegy 20 %-a áll közvetve vagy közvetlenül a CBF transzkripciós faktorok szabályozása alatt (Vogel és mtsai., 2005). A CBF faktorok direkt célpontjai a COR (Cold Regulated), valamint a fagyás miatt bekövetkezı vízvesztés káros hatásai ellen védı DHN (dehidrin) géncsalád tagjai (Danyluk és mtsai., 1994, 1998;
Jaglo-Ottosen et al, 1998; Choi és mtsai., 1999; Dal Bosco és mtsai., 2003). (Utóbbiakat bizonyos publikációkban nem is különítik el önálló csoportként, hanem a LEA fehérjék dehidrin csoportjához tartozó COR fehérjeként nevezik meg ıket.) A Triticeae csoportban a COR gének expressziója egyértelmő korrelációt mutat a fagyállóság mértékével: a tesztek során fagyállóbbnak bizonyuló genotípusokban bizonyos COR gének termékei nagyobb mennyiségben akkumulálódtak, mint az alacsonyabb fagyállóságot mutató genotípusokban (Crosatti és mtsai., 1996; Fowler és mtsai., 1996a; Limin és mtsai., 1997; Danyluk és mtsai.,
1994, 1998; NDong és mtsai., 2002). Árpában a fagyállóság növekedése kapcsolatban áll azzal, hogy a fagyállóbb genotípusban a COR fehérje akkumulációja már magasabb hımérsékleten megkezdıdik (Rizza és mtsai., 1994). Gabonafélékben a CBF választ az Arabidopsis COR15a génjével homológ COR14b gén expressziójának emelkedése követi (Vágújfalvi és mtsai., 2000; Stockinger és mtsai., 2007). A COR15a génrıl Arabidopsisban kimutatták, hogy a gén polipeptid terméke a membránok károsodásának fı okát, a lipidek lamellárisból hexagonális II fázisba történı átmenetét akadályozza meg. Bár maga a gén a nukleuszban kódolt, de fehérjeterméke a kloroplasztisz sztrómájában lokalizált, valószínőleg a fotoszintetikus folyamatokban szerepet játszó membránok elfagyást követıen, a fénykitettség következtében kialakuló fotodegradációval szembeni védelme a feladata (Crosatti és mtsai., 1995, 1999; Rapacz és mtsai., 2008). Összességében a COR gének CBF gének mőködéséhez köthetı aktiválódása gabonafélékben nevezhetı a hidegakklimáció folyamatának megkezdıdését jelzı, leglényegesebb mozzanatnak is (Jaglo-Ottosen és mtsai., 1998).
A bemutatott rendszer a sejtek anyagcseréjét alapvetıen befolyásoló hatása, túlélésüket meghatározó szerepe miatt sokrétő szabályozás alatt áll. Legfontosabb szabályozó eleme a MYC (Myelocytomatosis Oncogene) típusú ICE transzkripciós faktor, ami a CBF gének promóterében megtalálható MYCR (MYC Responsive Element) elemekhez kapcsolódva, valamint a MYBR (MYB Responsive Element) elemekhez kapcsolódó MYB15 (Myelocytoblastosis Oncogene) represszort gátolva serkenti a CBF gének expresszióját (Badawi és mtsai., 2008; Chinnusamy és mtsai., 2003). A CBF gének mellett az ICE a hidegakklimáció folyamatában szerepet játszó egyéb szabályozó faktorok, például a ZAT12 represszor expresszióját is befolyásolja (Benedict és mtsai., 2006). A ZAT12 faktorról bebizonyították, hogy reaktív oxigénszármazékok képzıdésével járó stresszhatás esetén például a citoszolikus APX1 (Ascorbate Peroxidase 1) gén expressziójának megindításához szükséges (Rizhsky és mtsai., 2004). Maga az ICE1 gén folyamatosan expresszálódik, alacsony hımérséklet általi aktivációja a fehérjetermék térszerkezeti változásán keresztül történik, ami ezért rendkívül gyors választ tesz lehetıvé (Thomashow és mtsai., 2001;
Chinnusamy és mtsai., 2003). Az ICE fehérje aktivitását a HOS1 (High Expression of Osmotically Responsive Genes 1) és SIZ1 faktorok befolyásolják, poszt-transzlációs módosítások révén. A HOS1 gén olyan ubiquitin E3 ligázt kódol, ami az ICE fehérjét proteolízis általi degradáció útjára tereli (Lee és mtsai., 2001), a SIZ1 gén terméke, egy SUMO E3 ligáz viszont az ICE1 fehérje szumoilációjának segítségével megvédi a fehérjét az ubiquitin által közvetített proteoszómás degradációtól (Miura és mtsai., 2007). Miura és mtsai. (2007) kimutatták azt is, hogy ez a folyamat elengedhetetlen az ICE1 hideg hatására bekövetkezı, génexpressziót érintı regulációs hatásainak kifejezıdéséhez. A CBF transzkripció az ICE1 aktiváló hatása mellett negatív visszacsatolási kontroll alatt áll egyes CBF géntermékek (Arabidopsisban pl. a CBF2) (Novillo és mtsai., 2004), valamint egyéb géntermékek (pl. a már említett ZAT12 és ZAT10 transzkripciós faktorok) által (Guo és mtsai., 2002; Vogel és mtsai., 2005). Poszt-transzkripciós szabályozó mechanizmusok (pre- mRNS splicing, kis RNS-ek által befolyásolt mRNS export és degradáció), a megvilágítottság napi ritmusa, a fotoperiodizmus (Hayes és mtsai., 1997; Limin és Fowler, 2006; Stockinger és mtsai., 2007; Lindlöf, 2010) és diurnális ciklusok (Badawi és mtsai., 2007) szintén befolyásolják az expressziójukat, valamint a hidegakklimációs folyamatok kialakulását (Gong és mtsai., 2002; Xiong és mtsai., 2002; Mastrangelo és mtsai., 2005; Palusa és mtsai., 2007).
A COR14b gén expressziója a hideghatás és feltételezhetıen a CBF géntermékeken keresztül az ICE aktivitása mellett további faktorok által is befolyásolt. Lokalizációjából fakadóan ezek közül a legfontosabb a fény minısége, a vörös és a kék fény, valamint ma még ismeretlen, kloroplasztiszhoz köthetı faktorok (Crosatti és mtsai., 1999; Dal Bosco és mtsai., 2003, Franklin és Whitelam, 2007).
(Az ICE-CBF-COR rendszert és szabályozásuk fıbb komponenseinek kölcsönhatását a 2.
Ábra foglalja össze.)
2. Ábra Az ICE-CBF-COR rendszer legfontosabb szabályozó elemeinek mőködési sémája. (A kérdıjelekkel jelölt folyamatok, kölcsönhatások még nem teljesen ismertek, feltérképezésük jelenleg is folyik.) (A Szerzı ábrája)
Azonban a hideghatás által indukált génexpressziós változások nem csupán a CBF regulonhoz köthetık (Fowler és Thomashow, 2002; Yamaguchi-Shinozaki és Shinozaki, 2005). Ismert, hogy exogén abszcizinsav kezeléssel hideghatás hiányában is kialakítható fagyállóság a növényekben (Chen és Gusta, 1983). Ezt megerısítendı ABA-deficiens mutánsok vizsgálata során kimutatták, hogy fagyállóságuk abszcizinsavas kezeléssel a vad növényekhez hasonlóan hideghatástól függetlenül kialakítható (Koornneef és mtsai., 1982; Heino és mtsai., 1990;
Mäntylä és mtsai., 1995). A másik fontos jelátviteli útvonal, ami szerepet játszik a hidegakklimáció folyamatában tehát abszcizinsav által közvetített. Ennek az útvonalnak az aktivációját viszont nem közvetlenül a hideghatás, inkább a jégképzıdéssel párhuzamosan jelentkezı dehidratáció váltja ki. Ennek megfelelıen nem csupán hidegstresszel szembeni védekezésben van szerepe, hanem a szárazság, hı, ozmotikus és oxidatív stresszhatásokkal szembeni toleranciát is fokozza a mőködése (Kim és mtsai., 2004). Ez a válasz a CBF-ek által közvetítettnél lassabb, és egyrészt a bZIP családba tartozó AREB/ABF (ABA-Responsive Element Binding Protein / Factor) fehérjék (Choi és mtsai., 2000), másrészt MYB és MYC transzkripciós faktorok bevonásával (Abe és mtsai., 1997) játszódik le, melyek a válaszgének szabályozó régióiban megtalálható ABRE (ABA Responsive Element), illetve MYBR és MYCR cisz-ható elemekhez kötıdve fejtik ki hatásukat (Leung és Giraudat, 1998). Ezekrıl a transzkripciós faktorokról kimutatták, hogy aktivációjukhoz abszcizinsavtól függı szignálra, valószínőleg protein-kinázok által közvetített foszforilációra van szükség (Yoshida és mtsai., 2002; Johnson és mtsai., 2002).
Az abszcizinsav által közvetített és az általa nem befolyásolt útvonal természetesen nem független egymástól, bonyolult kölcsönhatási kapcsolat áll fenn a kettı között (Xiong és mtsai., 1999; Chinnusamy és mtsai., 2004). Ezt az is megerısíti, hogy az abiotikus stresszhatásokra megváltozott expresszióval reagáló gének promoterében az ABRE és a CRT/DRE elemek gyakran egyszerre fordulnak elı (Heino és Palva, 2003; Knight és mtsai., 2004), a cisz-ható elemek között mőködı kölcsönhatást szintén kimutatták (Narusaka és mtsai., 2003).
Az abszcizinsav mellett más növényi hormonok is szerepet játszanak a hidegakklimáció folyamatának szabályozásában, mint például a jazmonsav (Ding és mtsai., 2002) és a szalicilsav (Janda és mtsai., 1999; Shah, 2003). Ezeknek a növényi hormonoknak is kimutatták a kölcsönhatását a már tárgyalt útvonalakkal. Arabidopsisban például felfedezték, hogy az abszcizinsav által indukált génexpressziós változásokat szabályozó AtMYC2 transzkripciós faktor a jazmonsav által szabályozott védekezési válaszok kialakításában is szerepet játszik (Abe és mtsai., 2003).
A vernalizáció és a virágzás idızítését szabályozó rendszer kapcsolata a fagyállósággal
A mérsékelt égövi gabonafélék ıszi genotípusai általában nagyobb terméshozamot garantálnak, mivel az ısszel elvetett és kicsírázó, megerısödı növény tavasszal intenzívebb növekedésre képes a még csak akkor elvetett tavaszi fajtákhoz képest. Kalászolásuk ezért hamarabb következik be, elkerülve ezzel a különösen a szemfeltöltıdés idıszakát tekintve kritikus, aszályos nyári periódust. Az említett stádiumban fellépı vízhiány és magas hımérséklet ugyanis elsısorban az endospermium sejtszámának csökkenése (Nicolas et al, 1985; Kobata és mtsai. 1992), valamint a keményítı-bioszintézis enzimeinek gátlása miatt (Hurkman et al 2003; Gibson és Paulsen 1999) jelentısen csökkenti a hozamot. Az ısszel kicsírázó gabonanövényeknek azonban el kell viselniük a téli idıjárás káros hatásait, amikkel szemben a már kihajtott növény érzékenyebb, mint például a nyugalmi állapotban lévı mag.
Tavasszal, a vegetatívból a generatív stádiumba történı átlépés után különösen érzékennyé válnak a növények az alacsony hımérsékleti stresszre. Ennek porzósterilitás, a fejlıdı mag károsodása, valamint kalászkaelhalás lehet a következménye (Reinheimer és mtsai., 2004). A generatív állapotban fagy hatására fellépı, erıteljesebb károsodást valószínőleg egyrészt a reproduktív szervek magasabb érzékenysége, másrészt az anyagcsere átrendezıdése okozza, melynek során a sikeres szaporodás érdekében a generatív növényi részek, valamint az azokat szolgáló tápanyag- és energiaraktározó szövetek (pl. a magban lévı endospermium) felépítését végzı, energiaigényes folyamatok elsıbbséget élveznek a védıanyagok termelésével szemben. Az ıszi gabonaféléknek ezért a virágzás idızítését meghatározó vernalizációs igényük van, tehát a hajtáscsúcs vegetatívból reproduktív állapotba történı átmenetéhez elengedhetetlen, hogy a növény bizonyos idıtartamú, elızetes hideghatásnak legyen kitéve. Ez biztosítja, hogy a növény nem fejleszt a fagy által különösen veszélyeztetett, érzékeny generatív szerveket a várhatóan bekövetkezı, hosszabb ideig fennálló téli idıszak elıtt. Ebben az idıszakban ugyanis a vegetatív növényi részek optimális fagyállóságának fenntartásával a növény életben maradásának biztosítása a fı feladat. A vernalizáció és a hidegakklimáció folyamatainak összehangolása tehát kulcsfontosságú a növény túlélése és hatékony szaporodása szempontjából.
A gabonafélék virágzási ideje három különbözı géncsalád egymástól függetlenül öröklıdı génjeinek kontrollja alatt áll. Ezek a vernalizációs igényt meghatározó gének (VRN) (Trevaskis és mtsai., 2003; Danyluk és mtsai., 2003), a nappalhossz-érzékenységet befolyásoló fotoperiodizmus válaszgének (PPD – Photoperiod Response Gene) (Turner és
mtsai., 2005; Beales és mtsai., 2007) és a koraiság gének (EPS – Earlyness Per Se Gene) (összefoglalva: Snape és mtsai., 2001). A vernalizációs igényt meghatározó gének biztosítják a vegetatívból a generatív állapotba történı átmenet idızítését, annak összehangolását a hidegakklimációs folyamatokkal. Az ide tartozó öt génlókusz (Vrn-A1 – 5A, Vrn-1; Vrn-B1 – 5B, Vrn-2; Vrn-D1 – 5D, Vrn-3; Vrn-B4 – 7B, Vrn-5 és egy ismeretlen kromoszomális lokalizációjú gén) határozza meg az adott genotípus ıszi vagy tavaszi jellegét (Law és mtsai., 1991). A fotoperiodizmus válaszgének a nappalhosszra adott választ szabályozzák. A nappalhosszra érzékeny fajták fejlıdése ısszel, a nappalok rövidülésével lelassul, tavasszal pedig csak akkor kezdenek intenzívebb növekedésbe, amikor a napi megvilágítottság elér egy bizonyos idıtartamot, azt követıen azonban fejlıdésük felgyorsul. A koraiság gének a környezeti hatásoktól függetlenül határozzák meg a fejlıdés alapütemét.
A növények a vernalizáció következtében, a generatív állapotba történı átmenetet követıen elveszítik korábban megszerzett fagyállóságukat (Fowler és mtsai., 1996a, 1996b;
Fowler és Limin, 2004; Limin és Fowler, 2006). Limin és Fowler (2006) feltételezése szerint a hidegakklimációs képesség csökkentésében alapvetı szerepe van a vernalizáció során aktiválódó fı válaszgénnek, a VRN1 génnek. Ez a gén az Arabidopsisban leírt AP1 (APETALA 1) merisztéma identifikációs faktorhoz hasonló, CArG motifhoz kötıdı MADS box fehérjecsaládba tartozó terméket kódol (bár Arabidopsisban az AP1 gén nem része a vernalizációs rendszernek) (Danyluk és mtsai., 2003; Trevaskis és mtsai., 2003; Yan és mtsai., 2003). A géntermék egy kritikus koncentrációt elérve beindítja a hajtáscsúcsban a merisztéma generatív növényi szervek létrehozása felé mutató átalakulását, a reproduktív állapotba történı átlépést, meghatározó szerepet töltve be a folyamatban (Shitsukawa és mtsai., 2007).
A VRN1 gén aktivitása és a fagyállóság közötti kapcsolat vizsgálata során megfigyelték, hogy míg az ıszi genotípusok vernalizáció elıtt nagyon kis mértékben expresszálják a VRN1 gént, addig a fagyérzékeny tavasziakban az expresszió mértéke folyamatosan magas. Mivel a virágzás mindkét genotípusnál akkor következik be, amikor a géntermék koncentrációja elér egy küszöbszintet, ebbıl következik, hogy tavaszi genotípusoknál ez hideghatástól függetlenül, bizonyos fejlettségi állapotot elérve végbemegy, az ısziekben azonban csak a vernalizációt követıen alakul ki (Loukoianov és mtsai., 2005).
Ezt megerısítik korábbi, kromoszóma szubsztitúciós búza vonalakon végzett megfigyelések is, melyek szerint az 5-ös homeológ csoport kromoszómáin található gének (amik közé a VRN1 gén is tartozik) gyakorolják a legnagyobb hatást a fagytőrésre (Roberts, 1986; Danyluk és mtsai., 2003). Ezzel összhangban kimutatták, hogy az ıszi és tavaszi növekedési forma kialakításában a VRN-A1 lókuszon elhelyezkedı alléloknak van a legnagyobb szerepe (Snape és mtsai., 2001). Ez a jelenség a VRN1 gén szekvenciájában megfigyelhetı különbségekre vezethetı vissza. Búzában és árpában megállapították, hogy a domináns tavaszi jelleget a gén promóterének egy jól meghatározott területét érintı, kisebb deléciók, valamint az elsı intronjában elıforduló, nagyobb inszerciók és deléciók alakítják ki (Yan és mtsai., 2004b; Fu és mtsai., 2005; Cockram és mtsai., 2007; Pidal és mtsai., 2009). Hexaploid búzában egyetlen domináns tavaszi allél jelenléte a három kromoszómaszerelvény bármelyikén elegendı a tavaszi fenotípus kialakításához (Stelmakh, 1987). A VRN1 alléloknak a fagyállóság kialakítására gyakorolt hatását támasztja alá az a vizsgálat is, melyben kimutatták, hogy a nagyobb fagyállósági potenciállal bíró ıszi genotípusokban, melyekben tehát a VRN1 gén recesszív allélja található meg, bizonyos CBF gének nagyobb mértékben expresszálódnak, mint a domináns allélt hordozó tavaszi genotípusokban. Azonban ıszi genotípusokban vernalizációt követıen, amikor a VRN1 fehérje koncentrációja megnı, a CBF gének expressziója a kontroll növények szintjére esik vissza (Stockinger és mtsai., 2007).
A VRN gének aktivitását a vernalizáció mellett azonban a fotoperiodizmus is befolyásolja (Dubcovsky és mtsai., 2006). Fotoperiodizmusra érzékeny genetikai háttérrel
vizsgálva a VRN1 gén expresszióját kimutatható, hogy a hosszú nappalos megvilágítás növeli, míg a rövid nappalos megvilágítás csökkenti a géntermék felhalmozódásának mértékét, így elnyújtja a vegetatív fázist. A VRN1 gén expressziójában bekövetkezı csökkenés rövid nappalon (késı ısszel, télen), illetve a generatív állapotba történı átmenetkor viszont növeli a fagyállóság mértékét (Fowler és mtsai., 2001; Limin és Fowler, 2006), ami szintén a két rendszer közötti szoros kapcsolatra utal.
A VRN1 gén mőködését tehát alapvetıen a vernalizációs hideghatás és a fotoperiodizmus szabályozza, ám más VRN gének is hatással vannak az expressziójára.
Összetett kölcsönhatási rendszerük mőködésének magyarázatára a kísérleti eredmények alapján különbözı modellek születettek, ám még mindig nincs egységes álláspont ezek helyességét illetıen. Az egyik alternatív hipotézis szerint a VRN1 transzkripciós faktor a VRN3 gén aktivitását serkenti, ám a VRN2 direkt gátló hatása alatt áll (Shimada és mtsai., 2009), valószínőbb azonban, hogy a következıkben ismertetett, Distelfeld és munkatársai (2009a, 2009b), valamint Trevaskis és munkatársai (2007) által ajánlott megközelítés alapján megalkotott modell jobban szemlélteti a valós kapcsolatokat: Az Arabidopsis FT (FLOWERING LOCUS T) génjével – ami szisztémás virágzásmegindító jelként (florigén) funkcionál – homológ, hosszú nappalos megvilágítás által aktivált VRN3 RAF kináz inhibítor-szerő molekula serkentı hatással van a VRN1 gén mőködésére (Yan és mtsai., 2006;
Li és Dubcovsky, 2008). Ennek valószínőleg tavasszal van szerepe, amikor a nappalhossz növekedésének hatására „rásegít” a VRN1 gén vernalizáció-okozta aktivációjára, hogy a VRN1 faktor mennyisége elérhesse a generatív átmenethez kellı küszöbszintet. A VRN3 gén aktivitását nem vernalizált növényben azonban gátolják a szintén hosszú nappalon aktiválódó, CCT domaint tartalmazó, Zn-ujj fehérjéket kódoló VRN2 lókusz termékei, a ZCCT1 és a ZCCT2 (Zink-finger/CCT Domain Transcription Factor), melyeknek Arabidopsisban nincsenek egyértelmően azonosítható homológjaik. Ezek a faktorok tehát a virágzás represszoraiként funkcionálnak. A VRN2 gén azonban vernalizáció következtében, a VRN1 gén aktivitásának emelkedésével, illetve a nappalok rövidülésével inaktiválódik (Yan és mtsai., 2004a; Loukoianov és mtsai., 2005; Trevaskis és mtsai., 2003). A VRN2 gén mőködésének ez alapján az lehet a szerepe, hogy nyár végén, kora ısszel, amikor a nappalok még hosszabbak, ám a növény a közelgı hideg periódus elıtt áll, meggátolja a VRN1 gén VRN3 transzkipt általi, korai aktivációját, így azt a késıbbi vernalizációs hideghatáshoz köti.
A VRN2 gén tehát a vernalizációs igény kialakításáért felelıs. A vernalizáció folyamata közben, a nappalok rövidülésével azonban inaktiválódik, lehetıvé téve tavasszal a VRN1 gén direkt és indirekt (VRN3 által közvetített), pozitív visszacsatoláson alapuló, gyors aktivációját. Ezt bizonyítja, hogy a VRN2 gén deléciója ıszi VRN1 allélok jelenlétében is recesszív tavaszi növekedési formát eredményez (Yan és mtsai., 2004a).
(A VRN rendszer mőködésének, szabályozásának összefoglalása a 3. Ábrán látható.)