DOKTORI (PhD) ÉRTEKEZÉS
KOLLARICSNÉ HORVÁTH MARGIT
PANNON EGYETEM GEORGIKON KAR
KESZTHELY
2019
2
PANNON EGYETEM GEORGIKON KAR
FESTETICS DOKTORI ISKOLA
Iskolavezető:
DR. ANDA ANGÉLA, DSC egyetemi tanár
Témavezető:
DR. TALLER JÁNOS, PhD egyetemi tanár
Burgonya genotípusok nitrogén hasznosítása és a nitrogén asszimiláció genetikai vizsgálata
D
OKTORI(P
hD)
ÉRTEKEZÉSKOLLARICSNÉ HORVÁTH MARGIT
KESZTHELY
2019
DOI:10.18136/PE.2019.717
3
BURGONYA GENOTÍPUSOK NITROGÉN HASZNOSÍTÁSA ÉS A NITROGÉN ASSZIMILÁCIÓ GENETIKAI VIZSGÁLATA
Az értekezés doktori (PhD) fokozat elnyerése érdekében készült a Pannon Egyetem Festetics Doktori Iskolája keretében
Növénytermesztési és kertészeti tudományok tudományágban
Írta: Kollaricsné Horváth Margit Témavezető: Dr. Taller János
Elfogadásra javaslom (igen / nem)
……….
(témavezető) Az értekezést bírálóként elfogadásra javaslom:
Bíráló neve: …... …... igen /nem
……….
(bíráló) Bíráló neve: …... …... igen /nem
……….
(bíráló) A jelölt az értekezés nyilvános vitáján …...%-ot ért el.
Keszthely, ……….
(a Bíráló Bizottság elnöke) A doktori (PhD) oklevél minősítése…...
Veszprém/Keszthely, ……….
(az EDHT elnöke)
4
Tartalomjegyzék
Rövidítések jegyzéke ... 6
1. KIVONATOK ... 7
1.1 Magyar nyelvű kivonat ... 7
1.2 Abstract ... 8
1.3 Auszug ... 9
2. BEVEZETÉS ... 10
3. IRODALMI ÁTTEKINTÉS ... 12
3.1 A burgonya jelentősége ... 12
3.2 A magyar burgonyanemesítés történeti összefoglalása ... 13
3.3 A nitrogén jelentősége a növények növekedésében és fejlődésében ... 15
3.4 Nitrogén hasznosító képesség ... 16
3.5 A nitrogén hasznosításban szerepet játszó gének ... 16
3.6 A nitrogén mérésére leggyakrabban alkalmazott agronómiai módszerek ... 21
3.7 A burgonya fajták nitrogén hasznosítási paramétereinek vizsgálata ... 25
4. CÉLKITŰZÉSEK ... 29
5. ANYAG és MÓDSZER ... 30
5.1 A vizsgált növényi anyag ... 30
5.2 Tenyészedényes kísérletek és kezelések beállítása ... 31
5.4 Mérések a betakarításkor ... 35
5.5 A nitrogén tartalom meghatározása ... 36
5.6 Molekuláris genetikai vizsgálatok ... 37
5.6.1 A nitrogén asszimilációt katalizáló enzimek génjeinek vizsgálata ... 37
5.6.2 Génexpressziós vizsgálatok ... 39
5.7 Indexek számítása ... 40
5.8 Statisztikai analízis ... 41
6. EREDMÉNYEK és KÖVETKEZTETÉSEK ... 43
6.1 A gumóról indított tenyészedényes kísérletek eredményei ... 43
6.1.1 SPAD-érték ... 43
6.1.2 Fenológiai állapotok ... 47
6.1.3 Levélterület ... 49
5
6.1.4 Gyökér-hajtás arány ... 50
6.1.5 Termés-paraméterek... 52
6.1.6 Nitrogén tartalom... 56
6.1.7 Vízpotenciál ... 64
6.1.8 Nitrát reduktáz enzim aktivitás ... 65
6.1.9 Indexek ... 68
6.2 A gumóról indított és az in vitro (szövettenyésztett) növények nitrogén hasznosítási paramétereinek összehasonlítása ... 75
6.2.1 Klorofill tartalom ... 75
6.2.2 Termés paraméterek ... 77
6.2.3 A gumók nitrogén tartalma ... 81
6.3 Molekuláris genetikai vizsgálatok eredményei ... 83
6.4 A vizsgált tulajdonságok kapcsolata és alkalmazhatóságuk nitrogén hasznosítás vizsgálatára tenyészedényes kísérletben ... 90
6.5 A vizsgált genotípusok nitrogén hasznosítási paramétereinek összefoglaló értékelése 100
7. ÖSZEFOGLALÁS és JÖVŐBENI PERSPEKTÍVÁK ... 104
8. ÚJ TUDOMÁNYOS EREDMÉNYEK ... 106
10. KÖSZÖNETNYILVÁNÍTÁS ... 108
11. MELLÉKLETEK ... 109
12. ÁBRÁK ÉS TÁBLÁZATOK JEGYZÉKE... 110
13. IRODALOMJEGYZÉK ... 113
6
Rövidítések jegyzéke
N – Nitrogén
NUE – Nitrogén hasznosító képesség (Nitrogen Use Efficiency)
NUpE – Nitrogén felvétel hatékonyságának mutatója (Nitrogen Uptake Efficiency) NUtE – A felvett nitrogén hasznosulásának mértékére utaló index (Nitrogen Utilization Efficiency)
N7,50 – 7,50 mM NO3 koncentrációjú kezelés N3,00 – 3,00 mM NO3 koncentrációjú kezelés N0,75 – 0,75 mM NO3 koncentrációjú kezelés
K2013 – gumóról indított tenyészedényes kísérlet 2013.
K2014 – gumóról indított tenyészedényes kísérlet 2014.
2014V – 2014-ben in vitro (szövettenyészetett) növényekkel beállított tenyészedényes kísérlet
HI – Harvest Index
PCR – Polimeráz láncreakció (Polymerase chain reaction)
RT-qPCR vagy qPCR – Real-time PCR, valós idejű polimeráz láncreakció (Quantitative polymerase chain reaction)
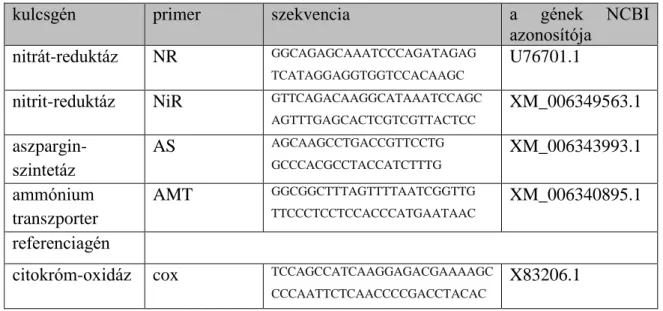
NR – nitrát-reduktáz NiR – nitrit-reduktáz
AMT – ammónium transzporter AS – aszparagin szintetáz cox – citokróm-oxidáz
WL – White Lady burgonyafajta
LSD – szignifikáns differencia (Least Significant Difference) Duncan – Duncan többszörös rang post hoc teszt
7
1. KIVONATOK
1.1 Magyar nyelvű kivonat
A műtrágyák környezetre gyakorolt káros hatásai ismertek, használatuk azonban elengedhetetlen a világ élelmiszer igényének kielégítéséhez, de ezzel egy időben törekednünk kell a környezet állapotának megóvására is. A burgonya mára a világ harmadik legfontosabb szántóföldi növénye lett, vetésterülete meghaladja a 19 millió hektárt. A burgonya genotípusok között jelentős nitrogénigénybeli és hasznosítási különbségek ismertek, jobb nitrogén hasznosító képességű burgonya fajták előállítása lehetséges és indokolt ökológiai és ökonómiai okok miatt egyaránt. A szerző a doktori munka során burgonya genotípusok nitrogén táplálkozásában lévő különbségek feltárását tűzte ki célul, hagyományos agronómiai és molekuláris genetikai módszereket egyaránt használva. A kísérletekbe öt burgonya genotípus került bevonásra, köztük négy keszthelyi nemesítésű fajtát (White Lady, Katica, Chipke, Hópehely) és egy amerikai nemesítési vonalat (S440).
A disszertáció eredményei alapján a tenyészedényes vizsgálat alkalmas a genotípusok közötti nitrogén hasznosítási különbségek kimutatására, így akár ez irányú előszelekcióra is. A legnagyobb termést mindegyik kísérletben a White Lady érte el, a Katica és a Chipke a középmezőnyben voltak, míg a Hópehely és az S440 adták a legkisebb termést. A disszertációban vizsgált genotípusok közül a legjobb nitrogén hasznosítási paraméterekkel a White Lady és Katica rendelkeznek. A keszthelyi fajták NUtE indexe kimagasló az S440-hez képest, a NUE index növeléséhez a NUpE, vagyis a nitrogén felvétel hatékonyságán lenne célszerű javítani.
A RT-qPCR vizsgálatokba a nitrogén asszimliációban szerepet játszó gének közül a nitrát reduktáz, nitrit reduktáz, aszparagin szintetáz enezimetet és az ammónium transzportereket kódoló gének expressziójának vizsgálata került bevonásra. A kísérletek során a különböző nitrogén ellátás mellett nevelt növények nitrogén táplálkozási génjeinek eltérő mértékű génexpressziót mutattak. A disszertáció a vizsgált genotípusok expressziós mintázatában lényegesen különbségeket tárt fel, a vizsgált gének expressziója és a nitrogén hasznosításban szerepet játszó tulajdonságok között szoros összefüggéseket mutatott ki.
8 1.2 Abstract
The task this work was to survey the examination of nitrogen utilization parameters of potato genotypes using agronomical and genetics methods as well. In the experiment four potato cultivars of the Potato Research Centre (White Lady, Katica, Hópehely, Chipke) and the S440 American breeding line. According to our results pot experiments are applicable for detection of differences in nitrogen utilization between genotypes, including pre-selection as well. White Lady produced the highest yield among the tested potato genotypes in every experiment. Medium yield was produced by Katica and Chipke and the smallest yield was produced by Hópehely and S440. White Lady and Katica cultivars have the best nitrogen utilization parameters among the tested potato genotypes. We analyzed the expression patterns of nitrate reductase, nitrite reductase, ammonium transporter and asparagine synthase genes with RT-qPCR. The expression pattern of the examined genotypes was different. Strong correlation between then expression of the studied genes and nitrogen utilization parameters was found.
9 1.3 Auszug
Die Dissertation erzielt Aufdeckung der Unterschiede in der Nahrungsaufnahme verschiedenen Kartoffel Genotypes mit agronomischen und molekularen Methoden. In den Experimenten fünf Kartoffelsorten wurden eingenommen, ein Typ aus der Vereinigte Staaten von Amerika (S440) und vier Ungarische (White Lady, Katica, Chipke, Hópehely) Sorten. Die Gewächstopfexperimente zeigte wesentliche Unterschiede verwendbar auch für vorherige Selektion. Im allen Experimenten die höchste Erträge wurden erreicht mit White Lady und der kleinsten mit S440. Unter den Genotypen, Katica und White Lady zeigten die besten Stickstoffanwendungen. Die Expressionen von vier Genen wurden mit RT-qPCR Methoden analysiert. Die Dissertation afdeckte wesentliche Verschiedungen, und zeigte die Relation zwischen der Expression von den vier analysierten Genen und Sticksstoffanwendung.
10
2. BEVEZETÉS
Az elmúlt négy évtizedben a világ nitrogén műtrágya felhasználása a hétszeresére növekedett, ezzel párhuzamosan a mezőgazdasági termelés megduplázódott (Hirel et al., 2007). Annak ellenére, hogy a műtrágyák környezetre gyakorolt káros hatásai ismertek, használatuk elengedhetetlen a világ élelmiszer igényének kielégítéséhez. Smil (1999) szerint a N műtrágya gyártást alapját képező Haber–Bosch eljárása, 20. század egyik legnagyobb felfedezése, lehetővé tette a világ népesség közel felének rendszeres, biztonságos élelmiszer ellátását.
Előrejelzések szerint a Föld népessége 2025-re eléri a 10 milliárdot. A növekvő népesség ellátásra egyre több élelmiszert kell előállítani, amivel szükségszerűen együtt jár a mezőgazdasági termelés fokozása. Az ehhez szükséges nitrogén műtrágyák előállítása azonban energiaigényes folyamat, az előállított mennyiség és a költségek emelkedése nagyban befolyásolja a mezőgazdasági termelés gazdaságosságát. A következő évtizedekben a népesség élelmiszerigényének ellátásához növelni kell a mezőgazdaság hatékonyságát. A kihívás az, hogy ezzel egy időben a környezet állapotát is meg akarjuk óvni (Dyson, 1999).
A genotípus – nitrogén kölcsönhatás létezése számos növényfajnál bizonyított (Sanford and MacKown, 1986; Arsenault et al., 2001; Gallais and Hirel, 2004; Ahmad et al., 2008; Berry et al., 2010; Marschner, 2012). A fent említett problémákra alternatív megoldás lehet, ha a tápanyagot jobb hatásfokkal hasznosító fajtákat nemesítünk. A növények számára kijuttatott nitrogén műtrágyák hasznosulásának javításával csökkenthető a környezeti terhelés, valamint a termesztés költségei is.
A termesztett növények fajtái között a nitrogén hasznosításbeli különbségek eltérő mértékben jelentkeznek. Swain és mtsai. (2014) a NUE (Nitrogén hasznosító képesség) tulajdonságok optimalizálási lehetőségeit vizsgálták búza és burgonya esetében. A kísérletben a burgonyánál szignifikáns hatást tudtak kimutatni a fajták, a termés és más NUE tulajdonságok között. A különböző burgonya genotípusok eltérő N- igényeit számos kutató bizonyította már, fajták (Kleinkopf et al., 1981; Zebarth et al., 2004b) és vad gumó-termő fajok (Errebhi et al., 1999) tekintetében egyaránt. Ezek az eredmények azt jelzik, hogy lehetséges jobb N hasznosító képességű burgonya fajták előállítása. Ezen fajták termesztésbe vonása pedig mind ökológiai mind ökonómiai szempontból óriási előnyökkel járna.
11
Ahhoz, hogy a burgonyatermesztés során növelni tudjuk a termés mennyiségét és minőségét, optimalizálnunk kell a N ellátást is (Joern and Vitosh, 1995b). A genotípusok közötti NUE különbségek fiziológiai alapjai azonban kevéssé ismertek, annak ellenére, hogy az elmúlt évtizedekben számos fontos publikáció született ezzel kapcsolatban. Ezért rendkívül fontos, hogy megismerjük az egyes fajták N-igényét, feltárjuk a nitrogénhasznosításban meglévő genotípusos különbségeket. Így lehetőséget adva nem csak a fajtákra adaptált optimális nitrogén ellátás biztosítására (Joern and Vitosh, 1995b), de akár jobb nitrogén hasznosító képességgel rendelkező új fajták nemesítésére is.
12
3. IRODALMI ÁTTEKINTÉS
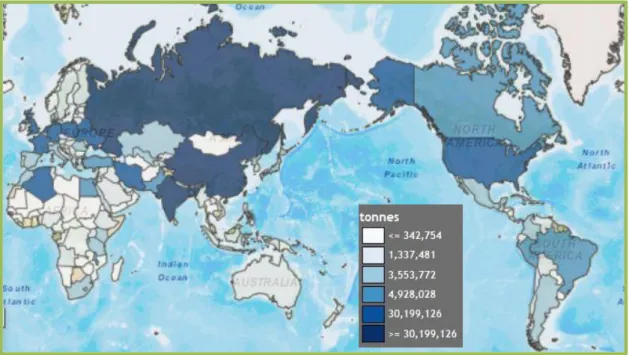
3.1 A burgonya jelentőségeA Solanaceae növénycsaládhoz – melynek a burgonya (Solanum tuberosum L.) is tagja – számos gazdaságilag fontos növényfaj tartozik, úgymint a paradicsom (Solanum lycopersicum L.), a padlizsán (Solanum melongena L.), a dohány (Nicotiana tabacum L.) vagy a paprika (Capsicum annuum L.). A burgonyát az ember körülbelül 7000-10000 évvel ezelőtt vonta termesztésbe, a mai Peru és Bolívia területén (Spooner et al., 2005). A burgonya termesztési területe a világon mára meghaladja a 19 millió hektárt (FAO, 2013), és a jelentősége egyre inkább növekszik, hiszen hektáronkénti szárazanyag és fehérje termelése meghaladhatja a legfontosabb gabonákét (Burton, 1989). A világ teljes burgonya termelése 2013-ban meghaladta a 370 millió tonnát, országonkénti megoszlását az 1. ábra mutatja be.
1. ábra: A világ burgonya termelése 2013 (tonna) (FAO, 2013)
A burgonya jelentőségét fokozzák kedvező táplálkozásélettani tulajdonságai:
változatos felhasználású, könnyen emészthető, jó biológiai értékű fehérjékben, tápanyagokban és vitaminokban gazdag táplálékunk. Ezen kedvező tulajdonságoknak köszönhető, hogy a burgonya mára – a búza és a rizs után – élelmezési szempontból a harmadik legfontosabb termesztett növénnyé lépett elő (Visser et al., 2009).
13
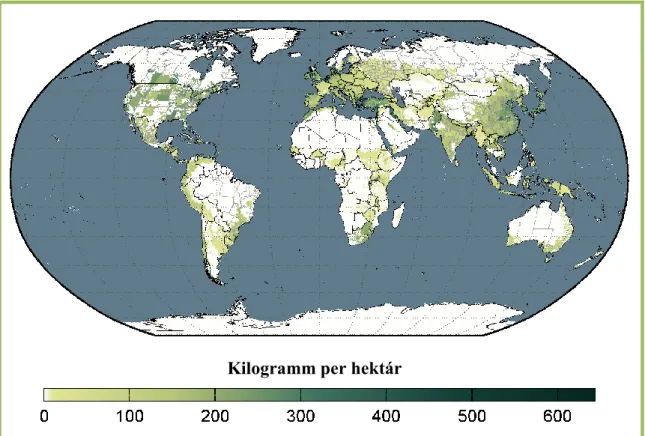
Termő területének és élelmezésbeli fontosságának megfelelően a termesztése során felhasznált nitrogén műtrágya is jelentős mennyiségű. A világ burgonya termesztési területein alkalmazott nitrogén műtrágya felhasználási rátát a 2. ábra mutatja be.
3.2 A magyar burgonyanemesítés történeti összefoglalása
Dél-Amerikában már több évezrede fogyasztják a burgonyát, azonban Európában csak a 16. század második fele óta ismerik. Megérkezéséről és a közvetítő személyéről számos elmélet kering, egyes források szerint Sir Walter Raleigh importálta Amerikából, míg más leírások Pizarro expedícióját vagy Mr. Herriotot említik (Laws, 2012). A legtöbb korabeli mű azonban Sir Francis Drakenek tulajdonítja a burgonya Európába juttatását, aki 1586-ban települt haza Virginiából.
A burgonya Magyarországra az 1650-es években érkezett meg, valószínűleg német egyetemeken tanuló diákok közvetítésével (Beke, 1930). Ahogy Európa többi részén, kezdetben Magyarországon is idegenkedve fogadta a lakosság, így csak lassan terjedt.
Kilogramm per hektár
2. ábra: A burgonya termesztés nitrogén műtrágya felhasználási rátája a világon, 2000 (Mueller et al., 2012)
14
Az 1920-as évekig a hazai burgonyatermesztés a külföldről behozott „Korai rózsa” és „Korai kék” fajtákon, illetve az ezekből „szelektált” tájfajtákon alapult.
Később ipari és takarmányozási célra nemesített fajták is bekerültek az országba, úgymint az Ackersegen, Deodara, Ella, Imperator, Iduna, Küger, Mittelfrühe, Schneeflocke és a Wohlmann (Zsom, 2006).
A hazai nemesítés az 1920-as évektől indult be, az első magyar fajta a „Magyar Kincs” volt, amelyet Nyitrán nemesítettek (Mándy and Csák, 1965). A hazai nemesítés főbb állomásait a 3. ábra mutatja be.
3. ábra: A hazai fajta nemesítés főbb állomásai
•Gülbaba
•Margit , Aranyalma
•Kisvárdai rózsa
•Boldogító, Lilla
Teichmann Vilmos
•Somogyi korai
Horn Miklós és Barsy Sarolta
•Somogyi kifli
•Somogy gyöngye
Horn Miklós és Teichmann Vilmos
•Keszthelyi sárga
•Keszthelyi rózsa
Sárvári István
•Hópehely
•White LAdy
•Százszorszép
•Góliát
Horváth Sándor és Lőnhard Miklós, jelenleg Polgár Zsolt és
munkatársai folytatják
•Nyírségi korai
•Nyírségi rózsa
•Szabolcsi rózsa
Forgó Sándor
•Boró
•Réka
•Rebeka
Bukai József, Iszályné tóth Judit és Mándi
Lajosné
•Pannónia
Kruppa József
1930-as, 1940-es és 1950-es évek
1950-es évek
1960-as, 1970-es évek
1970-es évek
1970-es évektől napjainkig 1970-es évek
1980-as és 1990-es évek
1980-as évektől napjainkig
Tornyospálca és Kisvárda, nemesítés a hazai klíma viszonyokhoz
Lovászpatona, és Mariettapuszta, klónszelekciós eljárások
Kecskemét és Keszthely, vírusrezisztencia nemesítés
Keszthely, vírusrezisztencia nemesítés
vírusrezisztencia nemesítés
diploid vadfajjal előállított hibridpopulációk
vírusrezisztencia és kiváló termőképesség
15
Ma Magyarországon a legfőbb nemesítési irányok a vírusrezisztencia tulajdonságok, a termésmennyiség és minőség javítása.
3.3 A nitrogén jelentősége a növények növekedésében és fejlődésében
A nitrogén az élőszervezetek negyedik leggyakoribb eleme a szén, az oxigén és a hidrogén után. Az a tény, hogy a nitrogén egyaránt alkotóeleme az aminosavaknak, nukleotidoknak és más fontos sejtalkotó vegyületeknek, mint a klorofill vagy egyes növényi hormonok, az egyik legfontosabb elemmé teszi az élő szervezetek számára. A talajban a felvehető nitrogén optimálisnál alacsonyabb szintje a burgonya növekedésének lassulását, késleltetését eredményezi, míg a többlete nem kívánt sarj- vagy gyökérarány-növekedéshez vezet, ami negatívan hat más tápanyagok és a víz felvételére is (Marschner, 1995), nőhet a betegségekkel szembeni fogékonyság, lassulhat az érés.
Számos tanulmány született, amelyekben szoros korrelációt mutattak ki a növények nitrogén ellátottsága és hormonháztartása között, különös tekintettel a citokininre. Ezt az összefüggést már igazolták dohány (Singh et al., 1992), nagy csalán (Wagner and Beck, 1993), árpa (Samuelson and Larsson, 1993) és kukorica (Takei et al., 2001) esetében is. A citokinin serkenti a sejtosztódást, a sejtek megnyúlását és differenciálódását, a gumók képződését és a színtestek érését is, vagyis jelentős hatással van a növényi növekedésre. Garnica és mtsai. (2010) eredményei szerint a NO3-
jelenléte egyértelműen emeli az aktív citokininek (pl.: zeatin, transz-zeatin-ribóz, izopentenil adenozin) szintjét és csökkenti a kevésbé aktív citokinin formák mennyiségét (pl.: cisz-zeatin-ribózt).
A tápanyagellátás befolyással van a növények vízfelhasználására is, vagyis kedvezőbb tápanyag ellátás mellett gazdaságosabb a növény vízfelhasználása is. Víz- és tápanyaghiány esetében a növények föld feletti szerveinek fejlődése, növekedése korlátozott, mert a stressz hatás miatt aránytalanul több energiát fordítanak a gyökérfejlődésre (Debreczeni and Debreczeniné, 1983). A N-ellátásnak jelentős hatása van a gyökér-hajtás arány alakulására (Lloret et al., 1999; Shangguan et al., 2000) is. A N-hiány növeli a gyökérzet felületét, ezzel együtt a gyökér energia felhasználását is, viszont mérsékli a hajtásba irányuló N-transzportot, vagyis a hajtásnövekedés csökkenését okozza.
16 3.4 Nitrogén hasznosító képesség
Moll és mtsai. (1982) definiálták először a nitrogén hasznosító képesség (NUE - Nitrogen Use Efficiency) fogalmát. A NUE megmutatja, hogy adott növény milyen hatékonysággal hasznosítja a rendelkezésére álló nitrogént. A NUE érték a növény által megtermelt összes szerves anyag és a növény számára felvehető összes nitrogén hányadosaként is kiszámítható. A folyamat két további részre bontható, a nitrogén felvételének hatékonyságára (NUpE: Nitrogen Uptake Efficiency) és a felvett nitrogén hasznosulásának mértékére (NUtE: Nitrogen Utilization Efficiency) (Zebarth et al., 2004b).
3.5 A nitrogén hasznosításban szerepet játszó gének
A legtöbb növény nem képes a légkör nitrogénjét megkötni, hanem nitrátion (NO3-), ammóniumion (NH4+) valamint szerves vegyületek (aminosavak, amidok, karbamid) formájában képes felvenni a nitrogént a talajból. Mindkettő ion jelen van általában minden talajtípusban, azonban relatív koncentrációjuk nagyon eltérő lehet. A mérsékelt égövi, jó szerkezetű, jól szellőzött talajokban a legjellemzőbb nitrogénforma a nitrát. A savanyú, vízzel telített talajokban azonban az ammóniumionok vannak túlsúlyban. Azokban a talajokban, ahol mindkét ion jelen van, a növények elsősorban az ammóniumiont veszik fel, mivel ez redukált forma, s így elhagyhatók az energiaigényes redukciós folyamatok.
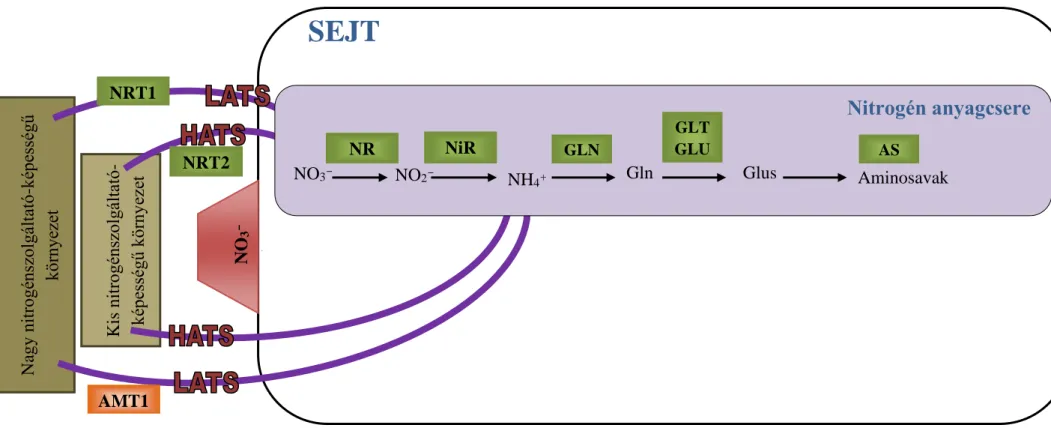
A nitrogén felvételében szerepet játszó transzportereket, enzimeket kódoló géneket már azonosították Arabidopsis-ban és rizsben (Oryza sativa L.), és egy részüket paradicsomban (Solanum lycopersicum L.) is, de burgonyában csak csekély hányadukat sikerült eddig azonosítani (Bucher and Kossmann, 2007). A növények nitrogén táplálkozásának folyamatát és felépítését a
4. ábra mutatja be.
A PTR (peptide transporter family), vagy a POT (proton dependent oligopeptide transporter family) géncsalád nagy jelentőséggel bír baktériumok, gombák és emlősök nitrogén asszimilációjában, amelyek a nitrogént rövid peptidek formájában veszik fel a környezetből (Steiner et al., 1995). Ezzel ellentétben a növények NO3- formában veszik fel a N-t a talajból (Crawford, 1995), amelyért az NRT1 és NRT2 géncsaládok által
17
kódolt, főleg gyökérsejtekben expresszálódó transzporterek felelősek (Tsay and Hsu, 2011). Az NRT1 géncsalád nitrát transzportereinek filogenetikai vizsgálata során beigazolódott, hogy a PTR génekhez tartoznak, ezért újabban az NPF (NRT1/PTR family) elnevezést is használják (Léran et al., 2014; Parker and Newstead, 2014;
Wittgenstein et al., 2014).
A magasabb rendű növények nitrát felvételében négy különböző transzporter rendszer játszik szerepet. A nagy affinitású transzporter rendszernek, a HATS-nak (high-affinity transport system) – amelyet az NRT2 géncsalád kódol (Orsel et al., 2002) – két típusa van: a cHATS (constitutive high-affinity) és iHATS (nitrate-inducible high- affinity). Az NRT1 géncsalád által kódolt a kis affinitású transzporter rendszer, a LATS (low-affinity transport system), amelynek szintén két típusa ismert, a cLATS (constitutive low-affinity) és az iLATS (nitrate-inducible low-affinity) (Glass et al., 2002; Glass, 2003; Okamoto et al., 2006).
Az NRT2 géncsalád által kódolt HATS akkor aktiválódik a növényekben, ha a környezet N-szolgáltató képessége kicsi. Ekkor először a cHATS lép működésbe, mely egy lassabb nitrogén felvételt eredményez, ezzel szemben az iHATS egy sokkal gyorsabb felvételt biztosít a növény számára. Az NRT1 által kódolt LATS-ot a jó nitrogén ellátottságú környezet aktiválja (Williams and Miller, 2001).
18
4. ábra: A nitrát és citokinin szintézis szabályozó-hálózatának modellje Sakakibara és mtsai. (2006) kutatásai alapján
AS: aszparagin szintetáz, AMT: ammónium transzporter, GLN: glutamin szintáz, GLT: NADH-függő glutamát szintáz, GLU: ferredoxin-függő glutamát szintáz, NR: nitrát reduktáz, NiR: nitrit reduktáz, NRT: nitrát transzporter
SEJT
NO3- receptor
Nagy nitrogénszolgáltató-képességű környezet Kis nitrogénszolgáltató- képességű környezet NRT1
NRT2
AMT1
NO3- NO2-
NH4+ Gln Glus
NiR GLN
N
GLT GLU
Aminosavak
Nitrogén anyagcsere
NR AS
19
A gyökerekben lévő NRT gének működésén és a gyökér sejtek plazma membránján levő kis és nagy affinitású transzporterek aktivitásán kívül a gyökérzet mérete és szerkezete is befolyásolja a nitrogén felvétel hatékonyságát (Li et al., 2015;
Zhang et al., 2015), így ez a három tényező együtt határozza meg azt (Léran et al., 2015; Liu et al., 2015).
A nitrát reduktáz (NR) a N asszimiláció első génje, amely a nitrátot nitritté redukáló nitrát reduktáz enzimet kódolja. Jelenlegi ismereteink szerint a nitrát reduktáz gén a citoplazmában expresszálódik, működését számos endogén és környezeti tényező befolyásolja, úgymint a rendelkezésre álló nitrát, továbbá a fény, a cukorok és a víz (Meyer and Stitt, 2001).
A nitrát reduktáció a N asszimiláció intenzíven kutatott területe. A nitrát reduktáz gént már sikerült klónozni, és működését (Kruse et al., 2002; Djennane et al., 2004; Lillo et al., 2004; Lea et al., 2006; Zhao et al., 2013) különböző expressziós kísérletekben vizsgálni. Quilleré és mtsai. (1994) szintén dohányon figyelték meg, hogy a nitrát reduktáz gén felül expresszálásával csökkenhet az N akkumuláció. Harris és mtsai. (2000) burgonyában vizsgálták a NR gén expresszióját különböző N ellátású kezelésekben. Eredményeik azt mutatták, hogy az alacsony N szolgáltató környezetben a NR csak a hajtásokban és levelekben expresszálódik, de magas N szolgáltató környezetben a sztolóban és a gumókban is aktív.
A N asszimiláció második szakasza a nitrit ammóniává redukálása, amit a nitrit reduktáz enzim génje (NiR) kódol. A NiR génexpressziója az eddig ismert adatok szerint növényfajonként eltérő lehet. A közönséges dohány (Nicotiana tabacum L.) esetén négy különböző homológja ismert: a nii1, nii2, nii3 és nii4 (Kronenberger et al., 1993).
Szintén Nicotiana tabacum vizsgálata során bizonyították, hogy a nii gének gyökérben és levélben is expresszálódhatnak, sőt a különböző homológok kifejeződése szervspecifikus. A nii1 és nii3 a levelekben, míg a nii2 és nii4 a gyökérben fejeződik ki nagyobb mértékben (Kato et al., 2004). A nitrit redukció a plasztiszokban – gyökérben is – megy végbe, a nitrát reduktáz működéséhez hasonlóan a nitrát mennyisége és a fény intenzitása szabályozza (Crété et al., 1997). A nitrát reduktáz gén sötétben inaktívvá válik vagy legalábbis csökken az expresszió szintje (Lillo et al., 2004), ezzel szemben a NiR kifejeződésének szintje bizonyos mértékig stabil marad sötét periódusok után is (Li and Oaks, 1995). Kyaing és mtsai. (2012) bizonyították, hogy a nitrit reduktáz túlműködése növeli a NR és NiR mennyiségét, csökkenti a sejtek nitrit tartalmát, de nem volt hatással az oldható fehérje koncentrációra és a növények növekedésére.
20
A növények képesek nitrogént felvenni nitráton kívül ammónia formájában is.
Ebben az esetben is szerepet kap a HATS és a LATS transzporter rendszer. Az ammónium felvételét kódoló géncsaládok az AMT1 és AMT2. Az ammónium transzportereket kódoló AMT géneket számos növényfajban azonosították már. Az AMT1 géncsalád öt tagját azonosították Arabidopsis-ban (Ninnemann et al., 1994;
Loqué and Wirén, 2004), hármat paradicsomban (Wirén et al., 2000; Lea and Azevedo, 2006), de ezen kívül ismertek AMT gének rizsből (Suenaga et al., 2003) és repcéből (Brassica napus L.) (Pearson et al., 2002) is. Utóbbi növénynél Babourina és mtsai.
(2007) bizonyították a nitrát jelentős szabályozó hatását, elsősorban az alacsony affinitású AMT transzporterre.
Az ammónium ion tovább alakulását a glutamin szintáz katalizálja, majd glutamát szintáz segítségével két glutamát képződik. amelyből az egyik bekapcsolódik az aminosavak szintézisébe. Az aszparagin szintézis magába foglalja az amid csoportok transzportját, a glutamin aszpartáttá alakulását, a folyamatot pedig a glutamin-függő aszparagin szintetáz (AS) katalizálja. Az elmúlt két évtizedben számos AS gént azonosítottak különböző növényfajban. A spárga (Asparagus officinalis L.) és a rizs esetében egyet-egyet, számos hüvelyes növényből két AS gént, a napraforgóból és Arabidopsis-bón hármat, míg kukoricában (Zea mays L.) négy AS gént izoláltak. A gének pontos funkciója és működése még kevéssé ismert, de az eddigi kutatási eredmények alapján az AS gének expresszióját főleg a fény és növény szöveteinek szén- nitrogén aránya szabályozza (Herrera-Rodríguez et al., 2002). Brears és mtsai. (1993) kísérleteikben túlexpresszáltatták az AS gént dohányban, ami 10-100 szoros aszparagin koncentráció növekedéshez vezetett és kis mértékű, de nem szigifikáns vegetatív növekedéshez. Az ASN1 aszparagin szintetáz gén felül expresszálódásának következményeként Lam és mtsai (2003) a magok fehérje tartalmának növekedését is megfigyelték Arabidopsis-ban.
21
3.6 A nitrogén mérésére leggyakrabban alkalmazott agronómiai módszerek
3.6.1 A burgonyagumó termésének és termés-paramétereinek mérése
A N gumótömegre gyakorolt hatását (White et al., 1974a; Belanger et al., 2000a;
Rodrigues et al., 2005a; Jatav et al., 2013) és a különböző genotípusok lényeges eltérő N-igényét (Kleinkopf et al., 1981; Westermann et al., 1994; Arsenault et al., 2001; Alva et al., 2002) már számos szerző leírta. Ezen kutatások eredményei megmutatták, hogy szoros korreláció van a gumó termés mennyisége és a NUE között. Zebarth és mtsai.
(2008) szerint a nagy terméshozamra való szelektálással egyben indirekt módon magas NUE-ra is szelektálunk.
A növekvő N műtrágya adagok kijuttatásával a termés mennyisége is növekszik (Saluzzo et al., 1999; Zabihi-e-Mahmoodabad et al., 2011; Zheng et al., 2015), de ha ez elér egy szintet, onnantól drasztikus termésdepressziót figyeltek meg. Joern and Vitosh (1995b) öntözött szántóföldi kísérletben (Michigan, USA) vizsgálta a N bevitel hatását a termésre, ‘Russet Burbank’ fajtánál. Kísérleteik során ezt a kritikus értéket 112 kg N ha−1-ban határozták meg. Feibert és mtsai. (1998) négy fajtánál állapította meg a legnagyobb hozamot adó N-határértéket. ‘Russet Burbank’ esetén 135, ‘Fontier Russet’
és ‘Ranger Russet’-nél 200, ‘Shepody’ esetén pedig 270 kg ha−1 N értéket állapítottak meg, szintén öntözött szántóföldi kísérlet keretében (Oregon, USA). Zheng és mtsai.
(2015) a ‘Kexin 1’ and ‘Shepody’ fajtáknál 225–270 kg ha−1 közötti N mennyiségben határozta meg a kritikus értéket, évjárattól függően. Lin és mtsai. (2004) tenyészedényes kísérletben, ‘Atica’ fajtánál erős korrelációt mértek a növekvő N- adagok és a termés között, viszont 1,9 g N / edény mennyiség felett termésdepressziót figyeltek meg. A N-adagok kritikus szintje genotípusonként eltérő és számos tényező befolyásolhatja, úgymint az évjárathatás (Arsenault and Malone, 1999; Zebarth et al., 2006; Haase et al., 2007; Ruza et al., 2013; Ospina et al., 2014), vagy a tápanyag kijuttatás módja (adagok, időzítés) (MacKerron et al., 1993; Chowdhury et al., 2002;
Gianquinto et al., 2004).
Fontos a gumó alak és a méret is, hiszen a termesztési célnak megfelelő változik az ezekkel a tulajdonságokkal szemben támasztott elvárás. Belanger és mtsai. (2002) a tápanyagutánpótlás és az öntözés gumó paraméterekre gyakorolt együttes hatását vizsgálták ‘Shepody’ és ‘Russet Burbank’ fajtákon, szántóföldi kísérletben (Kanada).
22
Eredményeik azt mutatták, hogy az emelkedő N-adagok hatására növekedett az átlagos gumótömeg. A növekedés aránya magasabb volt az öntözött kísérletben az öntözés nélkülihez képest, de jelentős különbségeket találtak a genotípusok között is. A
‘Shepody’ fajtánál szignifikánsan magasabb átlagos gumótömeget mértek, mint a
‘Russet Burbank’ fajtánál, az öntözés pedig tovább növelte ezt a különbséget.
Ismert a fajták közötti eltérés a gumószámban, Belanger és mtsai. (2002) szántóföldi körülmények közt vizsgáltak két fajtát, a ‘Russet Burbank’ esetében 8,7; a
‘Shepody’ fajtánál pedig 4,8 gumó növény−1 mennyiséget regisztráltak. Bár több kutató leírta, hogy a N megnöveli a gumók számát (Belanger et al., 2000a; Rostami et al., 2013), olyan kísérleteket is publikáltak, ahol nem tudták a N gumó szám növelő hatását szignifikánsan kimutatni (Morena et al., 1994; Zebarth and Rosen, 2007). Ennek valószínűleg az lehet az oka, hogy számos tényező, úgy mint a hőmérséklet, a talaj állapot, a különböző kórokozók vagy a gumók ültetéskori fiziológiai állapota a N ellátással együtt vagy külön-külön is hatással vannak a gumóképződésre.
A gumószám és az átlagos gumótömeg esetében egyaránt, a termés mennyiséghez hasonlóan megfigyelhető, hogy a növekvő N-adagokkal együtt növekszik ezek mennyisége is, de csak egy kritikus N-szint eléréséig. Ezt a szintet meghaladó N- adagok esetén mindkét paraméternél csökkenést figyeltek meg (Kleinhenz and Bennett, 1992; Kumar et al., 2007; Saeidi et al., 2009).
A burgonya – más termesztett növényekhez hasonlóan – képes a számára szükségesnél több N felvenni, ez azonban a burgonyatermesztők számára nem kívánatos tulajdonság. Ez a többlet költséget és terméskiesést okozó „luxusfogyasztás” csak úgy kerülhető el, ha ismerjük a termesztett burgonya fajtáink számára optimális N-adagot.
3.6.2 A klorofill tartalom indirekt mérése
A növények klorofill tartalma a N-ellátottság egyik legfontosabb mutatója. A klorofill tartalom meghatározására gyakran analitikai módszereket alkalmaznak, amely minden esetben a kísérleti növények roncsolása szükséges. Ezek a mérések idő és költségigényesek is. A hordozható klorofillmérő eszközök megjelenése alternatív megoldást jelent, kiküszöböli az analitikai módszerek hátrányait. Jelenleg két hasonló eszköz terjedt el a SPAD 502-Soil–Plant Analyses Development (Minolta, Japán) és a Hydro N-tester vagy HNt (Yara, Oslo, Norvégia) klorofill méter (Goffart et al., 2008;
Lemaire et al., 2008). Mindkét műszer a növények roncsolása nélkül, a levelek zöld
23
színének intenzitása alapján, 650 nm hullámhosszú fényt kibocsátva, annak elnyelését méri az eszköz (Goffart et al., 2008). A két optikai eszköz közül a SPAD (Minolta, Japán) az elterjedtebb, számos növényfajnál alkalmazzák a N-állapot jellmézésre (Peng et al., 1996; Feibo et al., 1998; Liu et al., 2012). Vos és Bom (1993) vizsgálta először az eszköz alkalmazhatóságát burgonyánál, és erős korrelációt találtak a burgonya analitikai módszerekkel mért klorofill tartalma és a SPAD eszköz mérési eredményi közt. Azóta számos kísérletben alkalmazták az eszközt burgonya N-ellátottságának meghatározásához szántóföldi, tenyészedényes és in vitro kultúrákban egyaránt (Minotti et al., 1994; Gianquinto et al., 2004; Jongschaap and Booij, 2004; Mauromicale et al., 2006; Wu et al., 2007; Busato et al., 2010; Giletto and Echeverría, 2013; Zheng et al., 2015). A tanulmányok legtöbbjében a SPAD értékek lényeges különbségeket mutatnak a N-ellátás, a genotípus és a növények kora függvényében.
3.6.3. A növények nitrogén tartalmának meghatározása
A növényi szövetek N tartalmának meghatározására számos módszert ismerünk, tudományos cikkek és könyvfejezet széles skálája foglalkozik a témával. Ezek közül a legrészletesebbek Haase és mtsai (2000) munkája, amiben az ismert és elfogadott invazív módszereket ismerteti. Booij és mtsai (2000) összegyűjtötték a növény roncsolása nélkül elvégezhető vizsgálatokat, Goffart és mtsai (2008) szemlecikke pedig az összes eddig ismert N-állapot meghatározásra alkalmas módszert hasonlítja össze.
Az invazív, teljes N tartalom meghatározására alkalmas analitikai módszerek közül a legáltalánosabban alkalmazott a Kjeldahl (Bradstreet, 1965) és a Dumas módszer (Kirsten and Hesselius, 1983). Ezek hátránya azonban, hogy laboratóriumi felszerelést és viszonylag sok időt igényelnek. Ezzel szemben a levélnyél nitrát koncentrációjának mérése (petiole nitrate concentration test) egy speciális műszernek köszönhetően helyben a szántóföldön is elvégezhető (Goffart et al., 2000; Haase et al., 2000; Zebarth and Rosen, 2007).
A N-tartalmat meghatározhatjuk egész növény, levelek, szár, hajtás (levelek és szár együtt) valamint a gumó vizsgálatával. Nagyszámú publikációt ismerünk, amelyek eredményei alapján megállapíthatjuk, hogy a burgonya szerveinek N tartalmát a N- kezelések szignifikánsan befolyásolják (Haase et al., 2007; Hu et al., 2014). A nitrogén adagok mértékével egyenes arányosságban nő a N tartalom is (Biemond and Vos, 1992;
Belanger et al., 2002; Zebarth et al., 2004b).
24 3.6.4. A vízpotenciál mérése
A víz a növényekben mindig a negatívabb vízpotenciálú hely felé mozog, ugyanez igaz a talaj – növény – légkör rendszerben is. A vízpotenciált energia / térfogat, vagyis nyomás (MPa) egységekben adjuk meg. A vízpotenciál mérését elsősorban szárazság-stressz mérésére, illetve a szárazságtűrés kimutatására használják. A tápanyagellátás befolyásolja a növények vízfelhasználást is, az ebből adódó különbségeket a vízpotenciál mérésével kimutathatjuk. A mérésére több módszert is használnak, a leggyakrabban alkalmazott műszer a Scholander-féle készülék (Scholander bomba). Az eszköz működésének lényege, hogy a vizsgált növény levele egy zárt térbe kerül, míg a vágási felület a műszeren kívül helyezkedik el. A zárt tér nyomását folyamatosan emeljük, így a benne lévő növényi részből a nedv a vágási felület felé nyomódik. Amikor a nedvcsepp megjelenik a vágási felületen, akkor kell leolvasni a gépen a nyomás értéket, amely megegyezik a levágott növényi szerv vízpotenciáljával (Wei et al., 2000).
5. ábra: A Scholander-féle készülék működése
25 3.6.5. A nitrát reduktáz enzim meghatározása
Az 1970-es években elterjedt nézet volt, hogy a N asszimiláció első lépése, a nitrát nitritté redukálása a folyamat fő limitáló tényezője, ezért nemesítési programokat indítottak magas nitrát reduktáz aktivitású kukorica genotípusok szelektálására, amiről úgy gondolták, együtt jár a nagyobb terméssel is (Andrews et al., 2004). Az bizonyított, hogy kukoricánál létezik NR-aktivitás beli különbség a genotípusok között és a tulajdonság örökölhető (Hirel et al., 2001). A magas nitrát reduktáz aktivitás azonban maga után vonta a levél alacsonyabb nitrát koncentrációját és az alacsonyabb SLA-t (specific leaf area; levél területe / levél száraz tömege) (Sherrard et al., 1986).
A levél nitrát reduktáz aktivitása és a termés mennyisége között negatív irányú korrelációt mutattak ki kukoricán és búzán (Triticum aestivum L.) (Hirel et al., 2001;
Masclaux et al., 2001). Ezek alapján a nitrát reduktáz aktivitás szintje nem limitálja a N asszimilációt, a termés mennyiséget elsősorban a N ellátottság határozza meg (Andrews et al., 2004).
3.7 A burgonya fajták nitrogén hasznosítási paramétereinek vizsgálata
A különböző burgonya genotípusok eltérő N igényeit már számos kutató bizonyította, nemesített fajták (Kleinkopf et al., 1981; Zebarth et al., 2004b) és vad gumó-termő fajok (Errebhi et al., 1999) tekintetében egyaránt. A jobb N hasznosító képességű burgonya fajták előállítása lehetséges, ez mind ökológiai mind ökonómiai okok miatt fontos törekvés. Ennek az első lépése a termesztett fajták és a nemesítésben használt vonalak nitrogén hasznosító képességének megismerése. A nitrogén hasznosítás vizsgálatára leggyakrabban alkalmazott agronómiai módszereket a 3.5 fejezetben ismertettük.
Egyre több cikk foglalkozik a nitrogén hasznosítással kapcsolatos tulajdonságok elemzésével, már az összes fontos termesztett növényfajnál elkezdték felmérni NUE tulajdonságok genotípusos különbségeit (kukorica: (Gallais and Hirel, 2004; Hirel et al., 2007), olajrepce: (Berry et al., 2010), rizs: (Cheng et al., 2011), búza (Gaju et al., 2011)), mind agronómiai mind molekuláris genetikai módszerekkel. Bár a burgonyánál is ismertek a genotípusos különbségek, a jobb NUE tulajdonságokra való szelektálás még nem terjedt el a burgonyanemesítés gyakorlatában, sőt a nemesítési anyagok és
26
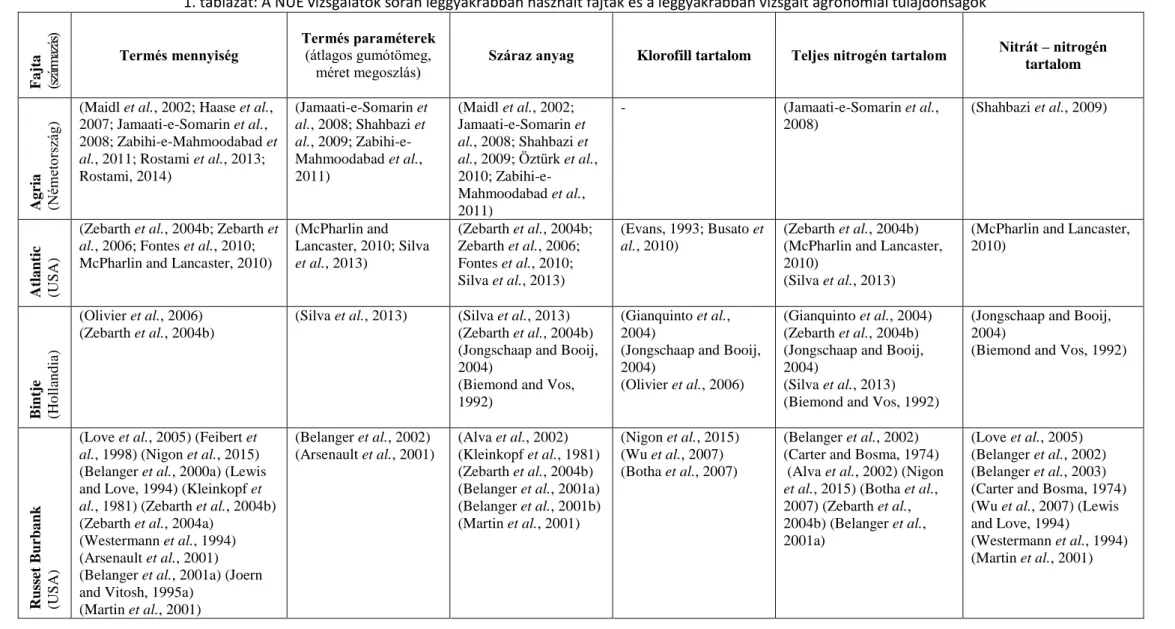
fajták ez irányú széleskörű felmérése sem történt meg. A burgonya fajták NUE tulajdonságaival foglalkozó cikkek ugyanazzal 15-20 fajtával dolgoznak, holott a világon termesztésben levő fajták száma ennél lényegesen nagyobb. A NUE tulajdonságok tekintetében legjobban kutatott fajtákat az 1. táblázat mutatja be. Ezek az:
’Agria’, ’Atlntic’, ’Bintje’, ’Russet Burbank’, ’Shepody’ és a ’Spunta’. A nitrogénellátás termésre gyakorolt hatása jól dokumentált (White et al., 1974b; Belanger et al., 2000b; Rodrigues et al., 2005b; Jatav et al., 2013), nem meglepő tehát, hogy a NUE vizsgálatok során a leggyakrabban vizsgált tulajdonság a termés mennyiség, melyet főleg szántóföldi körülmények közt vizsgáltak. A termésparaméterek: az átlagos gumótömeg és a méretmegoszlás vonatkozásában lényegesen kevesebb publikáció foglalkozik, habár a termesztési célnak megfelelően változnak az ezen tulajdonságokkal szemben támasztott elvárások. A száraz anyag tartalommal kapcsolatban a termésmennyiséghez hasonlóan nagyszámú publikáció született, a N ellátás gumók száraz anyag tartalomára gyakorolt hatása régóta ismert (Biemond and Vos, 1992;
Zebarth et al., 2006; Haase et al., 2007). A klorofill tartalom, a teljes nitrogén tartalom és a nitrát – nitrogén tartalom NUE vonatkozásai szintén jól kutatottak (kivéve az
’Agria’ fajtát). Az irodalmi adatok alapján a jobb nitrogén ellátás magasabb klorofill tartalmat (SPAD értéket) eredményez. Minotti és mtsai (1994) az ‘Allegheny’ és
‘Castile’ fajtáknál, míg Busato és mtsai (2010) az ‘Atlantic’, ‘Agata’, ‘Monalisa’ és
‘Asterix’ fajtáknál mértek klorofill tartalom növekedést az emelkedő N adagok hatására.
A burgonya N tartalom mérésére során a legtöbb irodalmi adat gumóra vonatkozik, viszont vizsgálatok alátámasztották, hogy a levelekben, lombban mért N tartalom is alkalmas a burgonya N állapotának meghatározására (Rodrigues et al., 2005a).
A hazánkban termesztett fajták nitrogén hasznosító képességéről alig, vagy semmilyen adattal nem rendelkezünk.
27
1. táblázat: A NUE vizsgálatok során leggyakrabban használt fajták és a leggyakrabban vizsgált agronómiai tulajdonságok
Fajta (származás)
Termés mennyiség
Termés paraméterek (átlagos gumótömeg,
méret megoszlás) Száraz anyag Klorofill tartalom Teljes nitrogén tartalom Nitrát – nitrogén tartalom
Agria (Németország)
(Maidl et al., 2002; Haase et al., 2007; Jamaati-e-Somarin et al., 2008; Zabihi-e-Mahmoodabad et al., 2011; Rostami et al., 2013;
Rostami, 2014)
(Jamaati-e-Somarin et al., 2008; Shahbazi et al., 2009; Zabihi-e- Mahmoodabad et al., 2011)
(Maidl et al., 2002;
Jamaati-e-Somarin et al., 2008; Shahbazi et al., 2009; Öztürk et al., 2010; Zabihi-e-
Mahmoodabad et al., 2011)
- (Jamaati-e-Somarin et al.,
2008)
(Shahbazi et al., 2009)
Atlantic (USA)
(Zebarth et al., 2004b; Zebarth et al., 2006; Fontes et al., 2010;
McPharlin and Lancaster, 2010)
(McPharlin and Lancaster, 2010; Silva et al., 2013)
(Zebarth et al., 2004b;
Zebarth et al., 2006;
Fontes et al., 2010;
Silva et al., 2013)
(Evans, 1993; Busato et al., 2010)
(Zebarth et al., 2004b) (McPharlin and Lancaster, 2010)
(Silva et al., 2013)
(McPharlin and Lancaster, 2010)
Bintje (Hollandia)
(Olivier et al., 2006) (Zebarth et al., 2004b)
(Silva et al., 2013) (Silva et al., 2013) (Zebarth et al., 2004b) (Jongschaap and Booij, 2004)
(Biemond and Vos, 1992)
(Gianquinto et al., 2004)
(Jongschaap and Booij, 2004)
(Olivier et al., 2006)
(Gianquinto et al., 2004) (Zebarth et al., 2004b) (Jongschaap and Booij, 2004)
(Silva et al., 2013) (Biemond and Vos, 1992)
(Jongschaap and Booij, 2004)
(Biemond and Vos, 1992)
Russet Burbank (USA)
(Love et al., 2005) (Feibert et al., 1998) (Nigon et al., 2015) (Belanger et al., 2000a) (Lewis and Love, 1994) (Kleinkopf et al., 1981) (Zebarth et al., 2004b) (Zebarth et al., 2004a)
(Westermann et al., 1994) (Arsenault et al., 2001) (Belanger et al., 2001a) (Joern and Vitosh, 1995a)
(Martin et al., 2001)
(Belanger et al., 2002) (Arsenault et al., 2001)
(Alva et al., 2002) (Kleinkopf et al., 1981) (Zebarth et al., 2004b) (Belanger et al., 2001a) (Belanger et al., 2001b) (Martin et al., 2001)
(Nigon et al., 2015) (Wu et al., 2007) (Botha et al., 2007)
(Belanger et al., 2002) (Carter and Bosma, 1974) (Alva et al., 2002) (Nigon et al., 2015) (Botha et al., 2007) (Zebarth et al., 2004b) (Belanger et al., 2001a)
(Love et al., 2005) (Belanger et al., 2002) (Belanger et al., 2003) (Carter and Bosma, 1974) (Wu et al., 2007) (Lewis and Love, 1994)
(Westermann et al., 1994) (Martin et al., 2001)
28
Fajta (származás)
Termés mennyiség
Termés paraméterek (átlagos gumótömeg,
méret megoszlás) Száraz anyag Klorofill tartalom Teljes nitrogén tartalom Nitrát – nitrogén tartalom
Shepody (Kanada)
(Feibert et al., 1998) (Belanger et al., 2002) (Hu et al., 2014) (Zheng et al., 2015) (Belanger et al., 2000a) (Zebarth et al., 2011) (Zebarth et al., 2004b) (Arsenault et al., 2001) (Belanger et al., 2001b)
(Hu et al., 2014) (Zebarth et al., 2011) (Zebarth et al., 2004b) (Li et al., 2010)
(Zheng et al., 2015) (Zebarth et al., 2011) (Botha et al., 2007) (Li et al., 2010)
(Belanger et al., 2002) (Hu et al., 2014) (Zebarth et al., 2011) (Botha et al., 2007) (Zebarth et al., 2004b) (Belanger et al., 2001a)
(Belanger et al., 2003) (Belanger et al., 2002) (Zebarth et al., 2011) (Li et al., 2010)
Spunta (Hollandia)
(Darwish et al., 2003) (Mohammad et al., 1999) (Tadesse et al., 2001)
(Mohammad et al., 1999)
(Tadesse et al., 2001)
(Darwish et al., 2003) (Mohammad et al., 1999)
(Saluzzo et al., 1999) (Tadesse et al., 2001)
(Mauromicale et al., 2006)
(Evans, 1993)
(Darwish et al., 2003) (Saluzzo et al., 1999)
-
29
4. CÉLKITŰZÉSEK
A hazánkban termesztett burgonya fajták nitrogén hasznosító képességéről kevés adattal rendelkezünk. Kutatásaink során keszthelyi nemesítésű burgonya fajták és nemesítési vonal nitrogén hasznosításában meglévő különbségeinek feltárását tűztük ki célul.
Kísérleteinket szabályozott környezetben, tenyészedényben nevelt növényekkel végeztük. Az agronómiai szempontból fontos mutatók vizsgálata mellett molekuláris genetikai módszereket használtunk.
1. A keszthelyi burgonya fajták nitrogén hasznosítási paramétereinek részletes vizsgálata, valamint nitrogén hasznosítási indexek értékelése tenyészedényes kísérletekben.
2. A tenyészedényes kísérletben mért paraméterek alkalmazhatóságának értékelése burgonya nemesítés során a nitrogén hasznosítási különbségek feltárására, fenotipizálásra és előszelekcióra.
3. Gumóról indított, illetve in vitro szövettenyészett növények nitrogén hasznosítással összefüggő tulajdonságainak összehasonlító értékelése.
4. A nitrát reduktáz, a nitrit reduktáz, az aszparagin szintetáz enzimet és az ammónium transzportereket kódoló gének vizsgálata, továbbá alkalmazási lehetőségeik a jobb nitrogén hasznosítású genotípusok szelektálására.
30
5. ANYAG és MÓDSZER
5.1 A vizsgált növényi anyagA kísérletekhez öt burgonya genotípust választottunk ki (2. táblázat), köztük négy keszthelyi nemesítésű fajtát (White Lady, Katica, Chipke, Hópehely) és egy amerikai nemesítési vonalat (S440). A fajták kiválasztása Hoffmann és mtsai. (2013) korábbi szántóföldi kísérletének eredményei, illetve a Burgonya Kutatási Központ fajtáinak figyelembe vételével történt. Az S440 vonal kísérletbe vonásának oka, hogy a Burgonya Kutatási Központ és a Növénytudományi és Biotechnológiai Tanszék Biotechnológia Csoportjának együttműködésével elkészült egy részleges kapcsoltsági térkép a White Lady-ben és az S440-ben is.
2. táblázat: A kísérletbe bevont genotípusok tulajdonságai (Fotók: Kollaricsné H.M.)
White Lady Katica Chipke Hópehely S440
Nemesítés helye
Keszthely, Magyarország
Keszthely, Magyarország
Keszthely, Magyarország
Keszthely, Magyarország
Wisconsin University,
USA Tenyészidő Középkorai Középkorai Középkorai Középkorai Középkorai
Gumó jellemzők
Világossárga héj és hús, kerek-ovál alak
Fényes rózsa héj, sárga hús,
szép ovális alak
Sárga héj, sárga hús, szép
szabályos alak
Sárga héj, fehér hús, kerek-ovál alak
Sárga héj, sárga hús
Lombozat jellemzői
Közepesen erős szár, világoszöld nagy levelek
Erős szár, magas, közepes
méretű sötétzöld
levelek
Erős szár, középmagas,
nagy, középzöld
levelek
Kevés vastag szár, középmagas, nagy és ovális középzöld
levelek
Közepesen erős szár, sötétzöld levelek
Termesztési cél
Étkezési burgonya,
alkalmas organikus termesztésre
Étkezési burgonya
Elsősorban feldolgozóipari
célfajta
Étkezési burgonya
Elsősorban feldolgozóipari
célfajta
31
5.2 Tenyészedényes kísérletek és kezelések beállítása
A tenyészedényes kísérleteket a Burgonyakutatási Központ temperálható, természetes fénnyel megvilágított üvegházában állítottuk be 2013-ban és 2014-ben.
A kísérletet 2013. március 18-án, illetve 2014. április 18-án indítottuk, 2013-ban gumóról, 2014-ben gumóról és in vitro növényekről. A kísérletek időtartama alatt az üvegházak napi átlaghőmérséklete 20 és 26 °C között mozgott.
Az ültetőközeg minden tenyészedényes kísérletben tápanyagmentes kvarchomok
#30 (600 µm) volt. A kísérletek során három liter űrtartalmú tenyészedényeket használtunk, melyekbe 1 darab – 2013-ban átlagosan 43,8 g (kivéve: S440 56,9 g) 2014-ben 35,2 g tömegű – gumót, illetve 4 darab in vitro növényt ültettünk. Az in vitro szaporított növények ültetését követően az akklimatizáció sikeressége érdekében a tenyészedényeket 3 napra polietilén tasakokkal takartuk, majd 7 nap múlva minden tenyészedényben 3 darabra csökkentettük a hajtások számát.
A kísérleteket kezelésenként négy ismétléssel (6. ábra), randomizált elrendezésben valósítottuk meg.
6. ábra: Tenyészedényes kísérlet 2014. (Fotó: Kollaricsné H.M.)
A növényeket 3 hetes korukig egységesen 50%-os Hoagland tápoldattal (Hoagland and Syder, 1933) öntöztük. Ezt követően kezdtük a kezeléseket, amelyek
32
időtartama alatt a növényeket három különböző nitrát koncentrációjú Hoagland tápoldattal öntöztük 42 (2013) illetve 28 (2014) napon keresztül.
A koncentrációk kiválasztásához a Li és mtsai (2010) munkája szolgált alapul, amely a burgonya nitrogén ellátottságát tanulmányozta a génexpresszió oldaláról. A kutatás során 0,00; 0,75 és 7,50 mM nitrogén koncentrációt alkalmaztak a növények neveléséhez. A saját kísérleteink irodalommal való összehasonlíthatósága miatt célszerűnek találtuk, hogy támaszkodjunk az általuk alkalmazottakra, különösen, hogy ezeknél a nitrogén koncentrációknál kimutathatók voltak szignifikáns génexpressziós különbségek. Alapos átgondolás után a 0,00-ás nitrogén koncentrációt elvetettük. Úgy ítéltük meg, hogy az általunk tervezett, a cikktől eltérő növénynevelési módszer és a cikkhez képest lényegesen hosszabb kísérlet indokolttá teszi ezt. Úgy gondoltuk több információhoz juthatunk, ha úgy árnyaljuk a módszerünket, hogy a 0,75 és 7,50 mM-os érték között felveszünk egy köztes koncentrációt, így esett a választásunk a 3,00 mM-os nitrogén koncentrációra.
A 7,50 mM koncentrációjú, 50%-os alap Hoagland tápoldat bőséges nitrogénellátást biztosított, közepes nitrogénellátásnak felel meg a 3,00 mM-ra, és alacsony ellátásnak a 0,75 mM NO3 koncentrációra módosított Hoagland tápoldat. A módosított Hoagland tápoldatok törzsoldatának makro elem összetételét az 3. táblázat mutatja be.
3. táblázat: A kezelésekhez alkalmazott módosított Hoagland tápoldatok makro törzsoldatának vegyszer összetétele.
Hoagland tápoldat makro törzsoldat
összetétele (1 liter, pH= 5,8)
7,50 mM NO3
koncentráció
3,00 mM NO3
koncentráció
0,75 mM NO3
koncentráció
KNO3 5,055 g 0 g 0 g
Ca(NO3)2 · 4 H2O 11,81 g 7,086 g 1,7715 g
MgSO4 · 7 H2O 4,932 g 4,932 g 4,932 g
KH2PO4 1,361 g 1,361 g 1,361 g
CaSO4 · 2 H2O 0 g 3,444 g 7,319 g
K2SO4 0 g 4,357 g 4,357 g
33
A kísérletek folyamán végig minden tenyészedénybe azonos mennyiségű folyadékot juttattunk. Hetente átlagosan 500 ml alap, illetve a kezeléshez módosított Hoagland tápoldatot öntöztünk, amit a növények hőmérséklettől függő igényének megfelelően ioncserélt vízzel egészítettünk ki. A kísérletek során kijuttatott tápoldatok és N pontos mennyiségét a 4. táblázat tartalmazza.
4. táblázat: A kísérletek során tenyészedényenként kijuttatott módosított Hoagland tápoldat és összes kijuttatott nitrogén mennyisége
2013 2014
Kijuttatott tápoldat
(ml) összes kijuttatott N tartalom (g)
Kijuttatott
tápoldat (ml) összes kijuttatott N tartalom (g)
Előnevelés Kezelés Előnevelés Kezelés
N7,50
2800 3200
0,630607
2600 2000
0,483466
N3,00 0,428836 0,357359
N0,75 0,327931 0,294293
5.3 Mérések és mintavételek a kezelés időtartama alatt
5.3.1 A klorofill-tartalom mérése
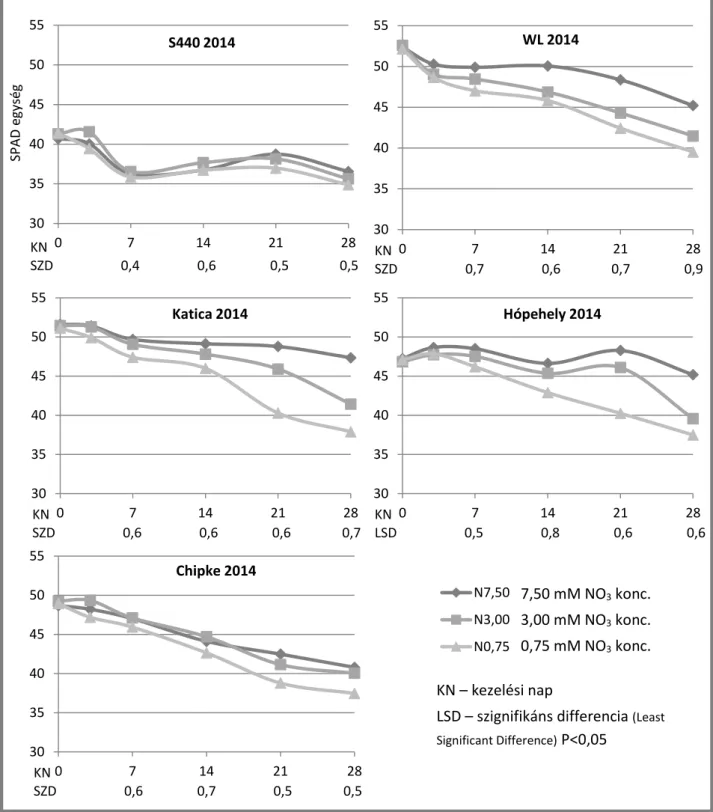
A kezelést megelőző (0.) napon, a kezelés 1., 3. és 7. napján, majd ezt követően hétnaponta klorofill mérést végeztünk, melyhez a roncsolás nélküli mérést lehetővé tevő SPAD-502-es (Konica Minolta, Osaka, Japán) hordozható klorofill-métert használtunk.
A méréseket mindig azonos napszakban, délelőtt 9 és 11 óra között végeztük. Minden mintavételi napon növényenként 10 mérést (ismétlést) végeztünk a levél színén, a fő ereket elkerülve (Jongschaap and Booij, 2004). A méréseket Dobos és mtsai. (2011) eredményei alapján a burgonya középső lombszintjén végeztük, mert ennek a lombszintnek az átlagos SPAD értéke megegyezik a teljes lombozat SPAD értékének átlagával.
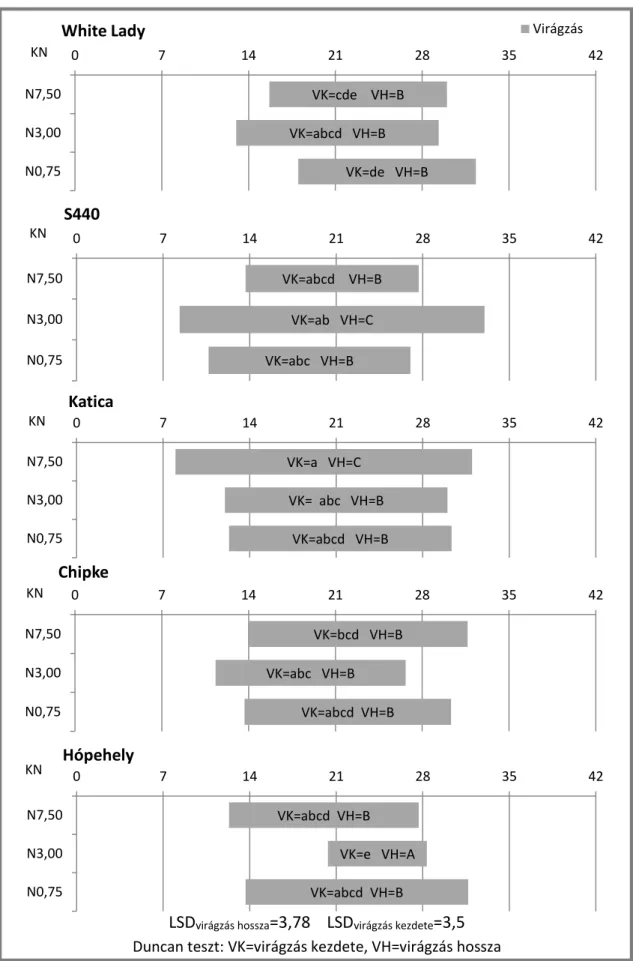
5.3.2 Fenológiai megfigyelések
A tenyészedényes kísérletek időtartama alatt feljegyeztük a növények (3 növény / genotípus) fenológiai állapot változásait. Dokumentáltuk a növényenkénti első bimbó megjelenését, az első virág kinyílását és a virágzás végét is.
34
5.3.3 Mintavétel molekuláris genetikai vizsgálatokhoz
A kezelést megelőző (0.) napon, a kezelés megkezdése utáni 1., 3. és 7. napon, majd ezt követően hétnaponta, a klorofill mérést követően levélmintákat gyűjtöttünk molekuláris genetikai vizsgálatokhoz. Három növényről, növényenként kettő darab levélkét szedtünk, egy teljesen kifejlődött összetett levélről, alufólia tasakba tettük és azonnal folyékony nitrogénben lefagyasztottuk azokat. A mintagyűjtő eszközöket minden növényt követően alkohollal és papírtörlővel tisztítottuk. A mintákat -70 °C-on tároltuk feldolgozásig.
5.3.4 Nitrát-reduktáz enzim aktivitás mérése
A méréseket a 2014. évi kísérletben, három alkalommal végeztük el, a 2.
(04.24.), a 15. (05.07.) és a 22. (05.14.) kezelési napokon. A mintavételek mindig délelőtt 10 és 11 óra között történek, igazodva a növények diurnális ritmusához. A genotípusonkénti azonos kezeléshez tartozó ismétlésekből (3 növényből) kevert mintát készítettünk, ezeket párhuzamosan 2 ismétlésben dolgoztuk fel, majd méréskor 3-3 technikai ismétlést végeztünk.
A vizsgálathoz levélmintákat gyűjtöttünk, melyeket folyamatosan jégen tartottunk. A kísérlethez a levéllemezeket azonos nagyságú, ép vágási felületű darabokra kell vágni (Jaworski, 1971), ezért egy papírlukasztó segítségével egyforma levélkorongokat készítettünk belőlük. Analitikai pontossággal 100 mg mennyiségű levéldarabkát mértünk ki és egy kémcsőbe tettük, amit folyamatosan jégen tároltuk.
Mintánként 5 ml assay keveréket (30 mM KNO3 66 mM Na-K- foszfát-pufferben oldva (pH 7,4-7,5), mely izopropanolra nézve 5 v/v%-os) adtunk hozzá (Jaworski, 1971; Min et al., 1998), majd a kémcsöveket parafilmmel lezártuk. A kémcsöveket 30 °C-os vízfürdőben, 60 percen keresztül sötétben inkubáltuk, közben 15 percenként a csöveket vortexeltük (Brunetti and Hageman, 1976; Min et al., 1998). Az inkubálási idő letelte után a kémcsöveket 5 percre forrásban lévő vízbe merítettük (Min et al., 1998) majd 10 percig szobahőmérsékleten hűtöttük. Ezt követően a kémcsövekből 2-2 ml folyadékot az új kémcsövekbe pipettáztunk és hozzáadtuk a Griess-Ilosvay reagenst, amely 1 %-os szulfanilamid és 0,02 %-os NED (N-1 naftil-etiléndiamin-dihidrogénklorid) oldatokból állt (Jaworski, 1971; Syrett, 1973; Kaiser and Lewis, 1984). A kémcsöveket parafilmmel lezártuk és 30 percre szobahőmérsékleten állni hagytuk, hogy