PANNON EGYETEM
GEORGIKON MEZ İ GAZDASÁGTUDOMÁNYI KAR
DOKTORI (PhD) ÉRTEKEZÉS
Csikászné Krizsics Anna
KESZTHELY
2008
PANNON EGYETEM
GEORGIKON MEZ İ GAZDASÁGTUDOMÁNYI KAR
Növénytermesztési és Kertészeti Tudományok Doktori Iskola Iskolavezet ı :
Dr. Gáborjányi Richard D.Sc.
egyetemi tanár, az MTA doktora
Témavezet ı k:
Dr. Debreczeni Béláné D.Sc.
egyetemi tanár, az MTA doktora Dr. Diófási Lajos D.Sc.
c. egyetemi tanár, az MTA doktora
A termés mennyiség, a termés min ı ség és a levelek tápelem tartalmának összefüggése különféle alanyokra oltott Cabernet sauvignon sz ı l ı fajtán
Készítette:
Csikászné Krizsics Anna
2008
A TERMÉS MENNYISÉG, A TERMÉS MINİSÉG ÉS A LEVELEK TÁPELEM TARTALMÁNAK ÖSSZEFÜGGÉSE KÜLÖNFÉLE ALANYOKRA OLTOTT
CABERNET SAUVIGNON SZİLİFAJTÁN Értekezés doktori (PhD) fokozat elnyerése érdekében
Írta:
Csikászné Krizsics Anna
Készült a Pannon Egyetem Növénytermesztési és Kertészeti Tudományok Doktori Iskolája keretében
Témavezetı: Dr.Debreczeni Béláné DSc
Elfogadásra javaslom (igen / nem) ………..
(aláírás) Témavezetı: Dr.Diófási Lajos DSc
Elfogadásra javaslom (igen / nem) ………
(aláírás)
A jelölt a doktori szigorlaton... % -ot ért el,
Az értekezést bírálóként elfogadásra javaslom:
Bíráló neve: …... …... igen /nem ……….
(aláírás)
Bíráló neve: …... …... igen /nem ……….
(aláírás)
A jelölt az értekezés nyilvános vitáján …...% - ot ért el.
Keszthely, ………. ……….
a Bíráló Bizottság elnöke
A doktori (PhD) oklevél minısítése…...
………
Az EDT elnöke
TARTALOMJEGYZÉK
Oldalszám
KIVONATOK 7
ABSTRACT 8
AUSZUG 8
1. BEVEZETÉS 9
2. IRODALMI ÁTTEKINTÉS 12
2.1. Szılıalanyok használatának kezdete 12
2.2. Affinitás, alany-nemes kölcsönhatás megnyilvánulási területei 14
2.3. Az alanyok gyökérrendszerének funkciója 16
2.4. Az alany-nemes kombinációk hatása 18
2.4.1. Alany-nemes kombinációk hatása a tápelemek felvételére 18 2.4.2. Alany-nemes kombinációk hatása a tápelemek megoszlására a
növényen belül 24
2.4.3. Alany-nemes kombinációk hatása a (vegetatív és generatív) terméshozamra 25 2.4.4. Alany-nemes kombinációk hatása a fürttermés-, a must- és borminıségre 27 2.5. Értékmérı paraméterekre gyakorolt tápelem hatások, korrelációk 28 2.5.1. A bogyó tápelem tartalmának változása az érés során 28
2.5.2. A must tápelem tartalma 29
2.5.3. Összefüggés a levél-, a bogyó-, a must tápelem tartalma és a termés paraméterek
között 29
2.6. Az alanyhatást módosító tényezık 31
2.6.1. Termıhely hatása, genotípus x környezet hatás 31
2.6.2. Évjárathatás 32
2.6.3. Termesztés-technológia hatása 33
2.7. A levélanalízis módszere 34
3. KÍSÉRLET HELYE, KÖRÜLMÉNYEI, ANYAGA ÉS MÓDSZERE 36
3.1. A kísérleti terület környezeti jellemzıi 36
3.1.1. A terület elhelyezkedése 36
3.1.2. A kísérleti tábla talajtani jellemzıi 37
3.1.3. A terület meteorológiai jellemzıi, talajnedvesség adatok 38
3.2. A kísérleti ültetvény jellemzése 38
3.2.1. A kísérlet beállítása 38
3.2.2. A Teleki szelekcióból származó Berlandieri x Riparia alanyok jellemzése 39 3.2.3. A Cabernet sauvignon fajta biológiai sajátosságai 40 3.3. Fenológiai vizsgálatok, a fürtátlag tömeg és a vesszısúly meghatározása 41
3.3.1. Fenológiai adatok 41
3.3.2. Fürtszám felvételezés és vesszısúly mérés 42
3.4. A fürttermés mennyiségi és minıségi vizsgálata 42
3.5. A levelek és a must tápelem tartalmának vizsgálata 43
3.6. Borok organoleptikus értékelése 44
3.7. Statisztikai értékelés 44
4. EREDMÉNYEK ÉRTÉKELÉSE, MEGÁLLAPÍTÁSOK 45
4.1. Alanyhatások értékelése 45
4.1.1. Fürttermés mennyisége 45
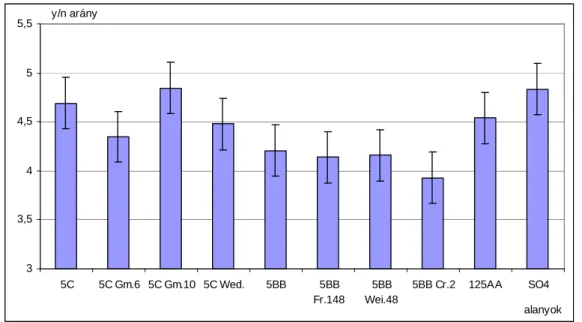
4.1.2 Vesszıtermés mennyisége, fürt- és vesszıtermés aránya 50
4.1.3. Mustvizsgálati eredmények 52
4.1.3.1. Mustfok 52
4.1.3.2. Titrálható savtartalom 55
4.1.3.3. Kémhatás (pH) 55
4.1.4. A szılılevelek tápelem tartalma 57
4.1.4.1. Nitrogén 58
4.1.4.2. Foszfor 59
4.1.4.3. Kálium 61
4.1.4.4. Kalcium 62
4.1.4.5. Magnézium 64
4.1.4.6. Mikroelemek 66
4.1.5. A must tápelem tartalma 69
4.1.5.1. Foszfor 69
4.1.5.2. Magnézium 69
4.1.5.3. Mangán 70
4.1.6. Organoleptikus borbírálat eredményei 71
4.2. Összefüggés vizsgálatok eredményei 72 4.2.1. A levelekben mért tápelem tartalmak közötti összefüggések 72 4.2.2. A must tápelem tartalmának összefüggése a levelek tápelem tartalmával,
tápelem kölcsönhatások a mustban 78
4.2.3. A termésparaméterek összefüggése a levelek tápelem tartalmával, valamint
a különbözı elemek mennyiségének egymáshoz viszonyított arányával 79
4.2.3.1. A termésmennyiséghez és mustfokhoz kapcsolódó összefüggések 79 4.2.3.2. Titrálható savtartalomhoz kapcsolódó összefüggések 87 4.2.3.3. Pathanalízis eredményei 93 4.2.4. A termés mennyiség, a termés minıség és a must tápelem tartalma közötti összefüggések 97
5. KÖVETKEZTETÉSEK ÉS JAVASLATOK 102
5.1. A vizsgált alanyok jellemzése az alany-nemes kölcsönhatásban kimutatott alanyhatások alapján 102
5.2. A tanulmányozott összefüggések rendszere 106
5.3. A kísérleti eredményekbıl levonható következtetések 108
6. ÖSSZEFOGLALÁS 110
7. TÉZISPONTOK / THESIS POINTS 115
KÖSZÖNETNYILVÁNÍTÁS 117
8. IRODALOMJEGYZÉK 118
9. FÜGGELÉK 130
9.1. Táblázatjegyzék 130
9.2. Ábrajegyzék 131
9.3. Mellékletek jegyzéke, mellékletek 133
KIVONATOK
Magyar nyelv ő kivonat
A termés mennyiség, a termés minıség és a levelek tápelem tartalmának összefüggése különféle alanyokra oltott Cabernet sauvignon szılıfajtán
A dolgozat a Berlandieri x Riparia Teleki szelekcióból származó alanyokra és alanyklónokra oltott Cabernet sauvignon (E.153 klón) ültetvény 15 éves (1990-2004) tartamkísérletének eredményeit dolgozza fel. A vizsgált alanyok: Teleki 5C és klónjai (Gm.6, Gm.10, Wed.), Teleki-Kober 5BB és klónjai (Fr.148, Wei.48, Cr.2), Teleki-Kober 125AA és Teleki-Fuhr S.O.4. A kísérlet a pécsi Szılészeti és Borászati Kutatóintézetben került beállításra 1986-ban. Az ültetvény széles sortávú, ernyımőveléső.
A dolgozat célja a fentebb felsorolt, s hazánkban általánosan alkalmazott alanyfajtákra oltott Cabernet sauvignon szılıfajta gazdasági, genetikai, élettani tulajdonságainak feltárása és értékelése az ültetvény teljes életciklusát felölelı kísérlet adatainak felhasználásával, a pécsi termıhelyen. A Szerzı vizsgálta az alanyok szerepét a Cabernet sauvignon teljesítménye és levélanalízissel követett tápanyag-hasznosítása, valamint ezek összefüggés rendszere vonatkozásában. A kitőzött célok elérése érdekében alanyonként értékelésre kerültek a következı tényezık: fürt- és vesszıtermés mennyisége, y/n arány, mustfok, titrálható savtartalom, pH, a virágzáskor és érés idején szedett levelek, valamint a must tápelem tartalma (N, P, K, Ca, Mg, Zn, B, Fe, Mn elemekre). Az alanyhatások kimutatását varianciaanalízissel, a változók közötti összefüggések vizsgálatát korrelációanalízissel, regresszió analízissel és pathanalízissel végezték.
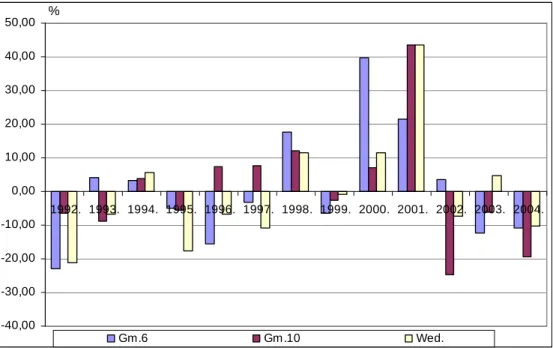
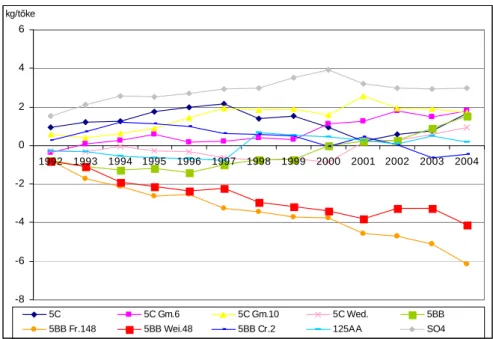
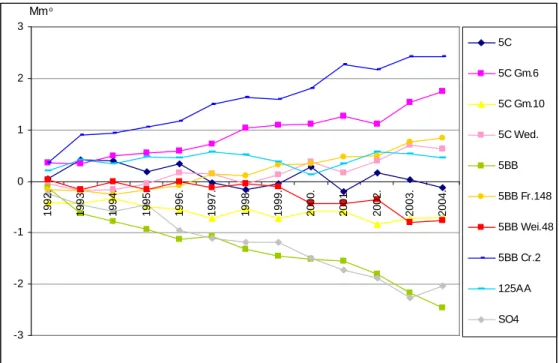
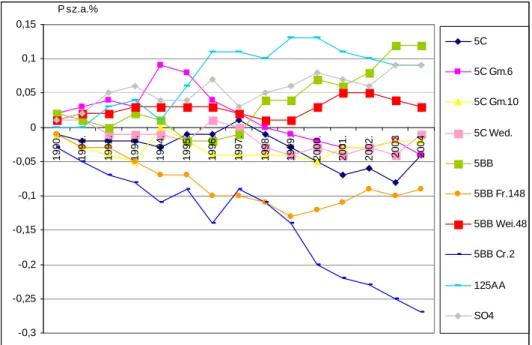
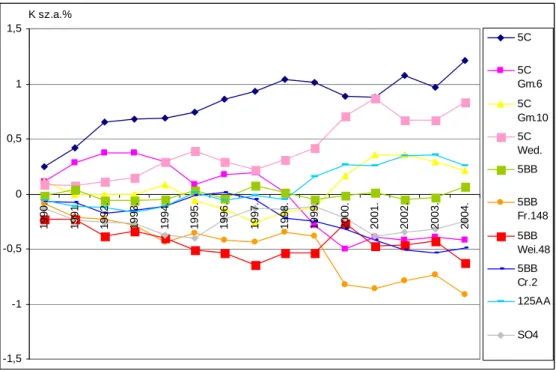
A tartamkísérletben lezajló folyamatokat és tendenciákat ’alanyhatásgörbék’ szemléltetik.
A feldolgozott eredményekbıl megállapítást nyert az adott termıhelyre vonatkozóan, hogy a Cabernet sauvignon termésprodukciója a különbözı alanyokon függ az ültetvény vizsgált életszakaszától. A Teleki 5C és Teleki-Kober 5BB alanyfajtákon lévı Cabernet tıkék rendszerint nagyobb hozamokat adtak, mint az alanyklónjaikon lévık. Az 5C és 5BB alanyfajták és klónjaik közötti különbségek a levelek tápelem tartalmára gyakorolt hatásban is jelentkeztek. A vizsgált kombinációkban és termıhelyen a tápelem tartalom alakulására a legjelentısebb alanyhatás a levelekben a magnézium, mangán és virágzáskori bórtartalom; míg a mustban a foszfor, magnézium és mangán esetében volt kimutatható. A statisztikailag igazolt összefüggésrendszer bemutatására áttekintı ábra készült. A nitrogén-bór, ill. mangán-cink elempárok közötti összefüggések a Cabernet levelekben és a mustokban is igazolódtak. Mind a kálium, mind a foszfor relatív túlsúlya vagy hiánya fontos minıségalakító tényezı, ezért a tápelem arányok optimum tartományára is készült módosító ajánlás.
Idegen nyelv ő kivonatok
ABSTRACT
The correlation between the quantity, the quality of yield and the nutrient content of leaves in Cabernet sauvignon vine variety grafted on different rootstocks
The dissertation elaborated results of 15 year-term-trial with Cabernet sauvignon vineyard grafted on Berlandieri x Riparia rootstocks. The aim of this work was to clear up the role of investigated rootstocks: on performance of Cabernet sauvignon, on utilizing nutrition – traced by leaf-analysis, and on its correlation-system.
The characterization of investigated combination in more respects, and more complete discovering of the relation-system may contribute to make the practice nutrient-management suitable for new challenge and for competitiveness. It was established that relative predominance or deficiency of potassium and phosphorus was quality forming factor, so the Author make a proposal to Cabernet sauvignon for modification of optimum range, generally used by consultants.
AUSZUG
Der Zusammenhang zwischen die Menge und die Qualität der Ernte und der Mineralstoffen Inhalt der Blätter bei der Sorte Cabernet sauvignon auf verschiedene Unterlagenreben
Die Dissertation verarbeitet die Ergebnisse eines Versuchs mit der Sorte Cabernet sauvignon, die seit 15 Jahre gelauft wurde, und die auf aus die Kreuzungen von Berlandieri x Riparia stammenden 10 verschiedene Unterlagenrebsorten und Klonen geboren ist. Das Ziel war die Erklärung die Rolle der untersuchte Unterlagenreben auf der Leistung und auf der Nährstoffverwertung – die mit Blattanalisation gemacht wurde – und mit diesem Zusammenhangsystem der Cabernet sauvignon.
Die vielseitige Charakterisierung der versuchte Unterlage-Veredlung Kombination – auf der Fläche in Pécs – und das volle Enthüllung des Relation Systems kann zu der neue Herausvorderungen in der Übung der Nährstoffwirtschafften auch entsprechen. Es wurde festgestellt, dass das relativ Dominanz oder Defizit aus Kalium und aus Phosphor spielen wichtige Rolle in der Qualität.
Deshalb machte die Autorin Modifikations-vorschläge – bei der Sorte Cabernet sauvignon – für den Optimumbereichen die in der Fachberatung angewendet werden.
1. BEVEZETÉS
„Natura parendo vincitur”
(A természetet csak neki engedelmeskedve gyızhetjük le)
Az egészséges környezet az ember lételemét, egészségének alapját jelenti. Elırelátó szemlélettel és preventív, megelızı módszereket magukba foglaló magatartásformákkal tudunk tenni az ember, a környezet, a növény egészségéért. A szelíd módszerek eszköztárának bıvítése, a meglévık kihasználása a környezetterhelés mérséklését szolgálja. Az e felismerés szellemében cselekvı integrált, környezetkímélı technológiák – a biológiai, agrotechnikai eszközök kiterjedt alkalmazásával – az agrokemikáliák korlátozott felhasználását teszik lehetıvé. A szılıtermesztés, mint intenzív kultúra esetében ennek kiemelt jelentısége van.
A szılınél a filoxéra elleni biológiai védekezésként bevezetésre került ellenálló alanyokra oltás is ezt a célt szolgálta, bár valószínőleg akkor ennek még nem tulajdonítottak jelentıséget. A biotikus eredető problémák körének bıvülésével a szılıgyökértető mellett a fonálférgekkel és Pierce betegséggel szembeni ellenálló képesség, ill. az Agrobacterium tumefaciens-el és több gombabetegséggel szembeni tolerancia megvalósulását is az alanyok teszik lehetıvé. Az alanyválasztás emellett a környezethez való alkalmazkodás lehetıségét teremtette meg. Termıhelytıl függıen elıtérbe kerülhet a mésztőrı képesség, alacsony pH értékő talajokhoz, szélsıséges nedvesség viszonyokhoz való alkalmazkodóképesség, a só- és hidegtőrés is. Az alany- nemes együttélésbıl származó tulajdonságokra gyakorolt hatás a harmadik terület, mely elsısorban az egymás közötti affinitásra, a kezdeti fejlıdés gyorsaságára, a tıke élettartamára, a növekedési erélyre, a termıképességre terjed ki. Ma is aktuális kérdés az alany-nemes kölcsönhatás vizsgálata a nemes értékmérı tulajdonságaira, a termésmennyiségre, ill. minıségi paraméterekre, a tıke fejlıdési szakaszaira, a fotoszintézis mértékére. Ezek hátterében az alany gyökerének kiterjedtsége, anatómiája, morfológiája, tápanyag felvétele, a fiziológiai tulajdonságokra (hormonok) gyakorolt hatása áll. A téma idıszerőségét napirenden tartja, hogy azonos ökológiai feltételek mellett különbözı nemes fajták nem ugyanazon alanyfajtán adják a legjobb eredményt, továbbá a vizsgált alany-nemes kölcsönhatás jelentıs mértékben módosulhat a környezeti feltételek (termıhely, évjárat) sajátosságai miatt is.
A tápanyag-gazdálkodási rendszer megújításában az alanyoknak is szerepük lehet. Az alanyok eltérı tápanyag-hasznosítási (felvétel, transzport) képességének felhasználásával mérsékelhetı a mőtrágyák alkalmazása, ami alkalmazásuk káros következményeinek elmaradását is eredményezi. Kiemelten a talajsavanyodást és a tápelem antagonizmusokat. A termıhelyhez és a nemes fajtához jól társított alany a gazdaságos és minıségi borszılıtermesztésnek a záloga. A körültekintı alanyválasztás – minden külön befektetés nélkül – képes jól hasznosítani a talaj adottságokat - vagyis mérséklıdnek a költségek. Másrészt ma ugyan egyre kevésbé termelési cél a jövedelemnek a termés mennyiség növelésével való biztosítása, viszont a gazdasági hasznot növeli,
ha az ültetvény élettartama nı, hosszabb ideig képes teljes értékő termés produkálására. Ebben jelentıs szerepet tölt be a tıkekondíció, ami a tıke alanyoktól függı sajátossága. A hosszú élettartam forrása emellett az egyensúlyi helyzetek (generatív-vegetatív, harmonikus tápanyagellátás) fenntartásában, helyreállításában van. A Cabernet sauvignon a térség ajánlott szılıfajtája, helyi adaptációjának jobb megismerése az eredményesebb termesztést segítheti elı.
A világ kiemelkedı szılıtermesztı államaiban - a helyi viszonyokhoz alkalmazkodva - sok alanyfajtával dolgoznak. Magyarországon viszont a termesztett fajták nagy száma jellemzı, ugyanakkor kevés alanyfajta. A filoxéravészt követıen leginkább a Riparia portalis terjedt el, az 50-es években az oltványok 40%-a volt ezen az alanyon. Az 1950-es években végzett alanykísérletek eredményei rámutattak arra, hogy a Riparia portalis értékét túlbecsülték, s helyét mindinkább átvették a Berlandieri x Riparia Teleki hibridek és azok szelektált klónjai. A 70-es évekig 12 alanyfajta szerepelt a termesztésre javasolt fajták között, majd uralkodó váltak az Teleki- Kober 5BB, Teleki 5C és Teleki 8B fajták. Az alanyválasztásnál meghatározó szempont volt, hogy erıteljes növekedéső és kiváló regenerálódó képességő alanyfajtákra van szükség a nagy termıfelület kialakításához. A több termıhelyen is - különbözı nemes fajtákkal - végzett kísérletek rámutattak az árnyaltabb alanyhasználatban lévı lehetıségekre. Az elmúlt 20-30 évben újabb alanyfajták kerültek be az üzemi kísérletekbe: Teleki 4A S.O.4, T.K. 125AA, Ruggeri 140, Chasselas x Berlandieri 41B stb.
Az alany- és nemesfajta közötti affinitás jelentısége, valamint a szılırekonstrukció szükségességének érezhetı szakmai kényszere miatt az 1980-as évek elején a pécsi Szılészeti és Borászati Kutatóintézetben is megtervezték a témában beállításra kerülı kísérleteket. Dr.Diófási Lajos vezetésével 1986-ban négy termıhelyen (Pécsett, Villányban, Siklóson és Szekszárdon) négy nemes fajtával (Olasz rizling, Rajnai rizling, Kékfrankos, Cabernet sauvignon) négy alanyfajtával és három-három alanyklónnal kerültek eltelepítésre ültetvények. A kísérletbe vont alanyok a következık voltak: Teleki 5C és klónjai (Gm.6, Gm.10, Wed.), Teleki-Kober 5BB és klónjai (Fr.148, Wei.48, Cr.2), Teleki-Kober 125AA, Teleki-Fuhr S.O.4. Valamennyi alany Berlandieri x Riparia keresztezésbıl származik, s eredetüket tekintve Teleki Zsigmond és fia Sándor révén a Villányi szelekciós mőhelyhez kapcsolódnak. A további szelekciók révén világfajtákká vált alanyok kísérletbe állításakor a villányi kötıdés a környezethez való alkalmazkodás tekintetében különösen szerencsés momentum. Az évek során e kísérletek aktuális eredményeirıl számos elıadásban, poszteren és OTKA jelentésben (1999) számoltunk be.
Az elmúlt 20 év sok tekintetben új kérdéseket vetett fel a témában – elsısorban a termelési cél tekintetében történt jelentıs változás -, így e tartamkísérlet eredményeinek célzott feldolgozása és a 15 éves tapasztalatok összegzése új információkkal szolgálhat. A dolgozatban csak a pécsi termıhely és a Cabernet sauvignon fajta eredményeinek feldolgozására került sor. Az alanyhatásokat módosító és a kiértékelés bizonytalanságát növelı tényezıket igyekeztünk kiszőrni
szempontra kell tekintettel lenni, hiszen az ültetvényszerkezet kialakítása, az alkalmazott termesztés-technológiai mőveletek is meghatározóak. Az elvégzett munka eredményessége függvényében ez a kiértékelési módszer a többi termıhelyre és fajtára is adaptálható. A vizsgálatba vont fajták genetikai háttere miatt a vizsgált paraméterek tekintetében nem lehetnek nagy különbségek, viszont az alany fajtakörön belüli eligazodást – elsısorban a relatív különbségeket – csak egy fajtacsaládra leszőkített vizsgálatban lehet igazán kimutatni.
A dolgozatban célul tőztük ki a hazánkban általánosan alkalmazott Berlandieri x Riparia eredető, Teleki szelekcióhoz köthetı alanyfajtákra és az azokból származó klónokra oltott Cabernet sauvignon szılıfajta gazdasági, genetikai, élettani tulajdonságainak feltárását és értékelését az ültetvény teljes életciklusát felölelı kísérlet keretében a pécsi termıhelyen. A cél elérése érdekében elvégzett feladatokat három csoportba sorolhatjuk:
Értékeltük az alany-nemes kombinációk hatását a tápelemek felvételére, a termés mennyiségére és minıségi mutatóira.
Az értékmérı paraméterek és a levelek, a must tápelem tartalma közötti összefüggéseket vizsgáltuk, hogy a termelési céloknak, a környezetkímélı termesztésnek leginkább megfelelı alany-nemes kombináció kiválasztását megalapozó információkhoz jussunk.
Vizsgáltuk és értékeltük az 5C és 5BB alanyklónok teljesítményét és tápelem hasznosítási sajátosságait a pécsi termıhelyen.
A bonyolult kapcsolatrendszer - amit az egyes évjáratok szélsıségei is jelentısen módosítanak - szinte lehetetlenné teszi az eligazodást a gyakorlatban. A dolgozatban a 15 éves tartamkísérlet eredményeinek feldolgozása több megközelítésben történt, hogy ebbe az összefüggésrendszerbe bepillantást nyerhessünk – döntéseink jobb megalapozása érdekében. A feltárt összefüggések korlátozás nélkül nem általánosíthatók, mivel mind a nemes fajta, mind a termıhely jelentıs módosító tényezık az alanyhatások érvényesülésénél.
A pécsi termıhely ökológiai adottságai a szılıtermesztés szempontjából kedvezıek. A terület környezeti érzékenysége jelentıs és az idıjárási szélsıségek növekedésével fokozódik. A terület vízmegtartó képessége gyenge, az aszályra érzékeny, s ez a tápelemek felvehetısége szempontjából is limitáló tényezı. Az alanyok széles köre lehetıvé teszi, hogy e problémát is kezelni lehessen, bár minden kritériumnak valószínőleg egy alany sem tud megfelelni. A Szılészeti és Borászati Kutatóintézet az alanyok aszálytőrésének tekintetében is igyekszik minél szélesebb körő információt adni, melyhez a Pécsett 1999-ben eltelepítésre került közepes sortávolságú, kisebb terheléső ültetvény nyújt majd remélhetıleg értékes információkat.
2. IRODALMI ÁTTEKINTÉS
2.1. Sz ı l ı alanyok használatának kezdete
Az alanyhasználatból és alany-nemes kölcsönhatásból adódó szılıtermesztési kérdésekkel Európa a filoxéravészt követıen szembesült. A szılıgyökértetőt (Daktulosphaira vitifoliae FITCH) Amerikából szılı szaporítóanyaggal hurcolták be. Károsítását Dél- Franciaországban 1858-ban, Magyarországon 1872-ben Pozsonyban és 1875-ben Pancsován észlelték elıször. A károsító Franciaországban 1,5 millió ha szılıt irtott ki, 10 év alatt nálunk a terméshozam a felére csökkent (LEHOCZKY és REICHART, 1968).
A szılıgyökértető leküzdésében áttörést a filoxéra ellenálló észak-amerikai fajok egymással, majd Vitis vinifera-val való keresztezéses nemesítése hozott. A legfontosabb rezisztenciaforrásokat a Vitis riparia MICHAUX, Vitis rupestris SCHEELE és a Vitis berlandieri PLANCH fajokban találták meg, mivel az alanyfajtákkal szemben támasztott követelményeknek leginkább ezek feleltek meg. E tulajdonságok a következık: filoxéra tőrıképesség, alkalmazkodás az európai éghajlati és talajviszonyokhoz (adaptáció), forradási képesség az európai szılıfajtákkal (affinitás), és dugványgyökeresedési képesség (EINSET és PRATT, 1975 in KOCSIS et al.,2000, BOUBALS, 1976, VERCESI, 1987).
Az ideális alannyal szemben támasztott követelményeket KOCSIS (1996) így összegzi:
erıteljes növekedésőnek kell lennie, hidegtőrınek, a különbözı gombabetegségekkel és rovarok kártételével szemben ellenállónak. Nagy vesszıhozamot kell produkálnia. A vesszınek nagy mennyiségő gyökeret kell képeznie, a nemes fajtával az összeoltást követıen egységet kell alkotnia. Gyökérzetének rezisztensnek kell lennie a filoxéra, a nematódák és egyéb talajlakó kártevıkkel szemben, s mindeközben a talajok tulajdonságaihoz is alkalmazkodnia kell, úgymint a magas mésztartalomhoz, sótartalomhoz, vagy szárazsághoz.
Az alanyokkal szemben támasztott követelmények tehát három fı területen fogalmazódtak meg, és fokozatosan bıvültek (/nek) is. Az alapterület a filoxéra (majd a rasszai) elleni ellenálló képesség, továbbá a fonálférgek, gyökérgolyva és egyéb károsítókkal szembeni tolerancia megléte. Az adott termıhely talaj-és klíma sajátosságaihoz, a környezethez való adaptáció is egyre újabb követelményeket támaszt az alanyokkal szemben a környezeti és termesztési feltételek módosulásával, ill. az új termıhelyek bevonásával. A filoxéravész után hazánkban elsısorban a mész indukálta klorózis tolerancia volt a legfontosabb tényezı, ma mindinkább elıtérbe kerül az aszálykérdés is. A világ szılıtermı területeit nézve viszont az alacsony pH, a szikes talaj, túl vizes talaj, hideg tolerancia is fontos tényezı (SANJUN GU, 2003). Az alany-nemes együttélésbıl származó tulajdonságokra gyakorolt hatás kiterjed többek között az egymás közötti affinitásra, a kezdeti fejlıdés gyorsaságára, a tıke élettartamára, a növekedési erélyre, a termıképességre, a bor minıségére, a tápelemek felvételére, a nemes fiziológiájára (CSEPREGI és ZILAI, 1988).
E harmadik területen belül is új gazdasági – társadalmi igények jelentkeztek, ill. kerültek elıtérbe (pl. környezetkímélı termesztés), melyek révén a korábbi értékítélet módosítására is sor kerülhet.
A kiindulási anyagok jellemzıi:
V.riparia: Észak-Amerika mérsékelt övezetének öntés talajairól származik. Sekélyen gyökeresedik,
rostos erısen elágazó gyökeret nevel. A talaj iránt nagyon igényes. Termékeny, mélyrétegő, humuszos, nem túl száraz, de nem is túl nedves talajt kíván. Nagy a mészérzékenysége. Igen korán fakad, és a ráoltott nemes fajtát is ilyen irányban befolyásolja. Filoxéra tőrıképessége igen nagy (JABOREK, 1988). Származása alapján a hideg klímát jól tőri. Kis növekedési erélye és korai érése miatt genetikailag a minıségi termesztést szolgálja (MORTON, 1994).
V.rupestris: Észak –Amerika déli részének mérsékelten meleg klímáján honos. Itt száraz homok- és
köves talajokon vadon fordul elı. Mélyen gyökeresedik. Kevés, elágazó húsos gyökeret nevel, szárazságtőrı. A talaj iránt nem nagyon igényes. Nagyobb a mésztőrı képessége, mint a Ripariáé.
Késın fakad. A ráoltott fajta érése elhúzódik. Filoxéra tőrıképessége igen jó (JABOREK, 1988).
V. berlandieri: Hazája Közép-Texas forró, száraz klímájú vidéke. Vadon a száraz, forró, nagy
mésztartalmú talajokon fordul elı. Gyökeresedı képessége rossz. A talajjal szemben igénytelen.
Igen nagy a mésztőrı képessége. Korán fakad és a ráoltott fajta korai érését segíti elı. Filoxéra tőrıképessége igen jó (JABOREK, 1988, COUSINS, 2005).
A Berlandieri x Riparia keresztezések célja a Riparia korai érésének és jó gyökeresedésének összekapcsolása a Berlandieri mésztőrı képességével (JABOREK, 1988).
Ezek a keresztezések eléggé erıteljes növekedésőek, de kevésbé, mint a Berlandieri x Rupestris keresztezések, melyek a meleg övezetek alanyai lehetnek (MORTON, 1994). Müller Thurgau elsıként hívta fel a figyelmet az amerikai és az eurázsiai fajok örökítı anyagainak kombinálására, mert az abból kapott hibrideknél várható jó adaptivitás (HAJDU és BAKONYI, 2006).
A filoxéravész idején tevékenykedett Franciaországban Alexis Millardet, Marques Charles de Gaset, Georges Couderc, Viktor Ganzin, Franz Richter, Olaszországban Antonió Ruggeri, Frederico Paulsen, akik kiváló, még ma is termesztésben lévı alanyfajtákat állítottak elı (GALET, 1988 in BAKONYI és KOCSIS, 2004).
„A filoxéravész záróköve szılıtermesztésünk ezeréves történetének. Számos új termesztési eljárás bevezetését tette szükségessé (oltványtermesztés, vegyi védekezés stb.), megnyitotta az utat a telepítés és termesztéstechnika korszerősítéséhez, a fajtaszerkezet és választék rendezéséhez” (KOZMA, 1966). Hazánk sajátos éghajlati - és talajviszonyainak megfelelı alanyok nemesítésén Horváth Géza, Szilágyi János és Teleki Zsigmond dolgozott az 1800-as években.
Teleki Zsigmond a Franciaországból hozatott alanymagokból magoncokat nevelt. Ezekrıl kiderült, hogy természetes keresztezıdés útján létrejött hibridek. A magoncokat szelektálta, majd tíz csoportba rendezte. Teleki Zsigmond Berlandieri x Riparia 5A néven három típusnak, Berlandieri x
Riparia 8B jelzéssel 5 típusnak a keverékét forgalmazta, ezzel lehetıséget teremtett a csoportokban található fajták különválasztására, szelektálására (KOCSIS és BAKONYI, 2000).
A Teleki fajták elsı nagy szelektálója Ausztriában Kober volt. İ az erıs növekedésőeket dupla nagy betőkkel jelölte (AA, BB). Kober is még fajtakeveréket adott tovább, csupán új jelzéssel, így lett a Teleki 5A-ból Kober 5BB (KOCSIS és BAKONYI, 2000, BAKONYI és KOCSIS, 2004, HAJDU és BAKONYI, 2006). WALKER és LIU (1995) izoenzim analízissel (hét enzimrendszer felhasználásával), GUERRA és MEREDITH (1995) RFLP analízissel állapították meg az 5A és 5BB fajták azonosságát.
Teleki Sándor az 1920-as években szelektálta ugyancsak a Teleki 5A-ból a Teleki 5C fajtát.
Kober szelektálta a 125AA fajtát a Teleki 7B csoportból.
Fuhr 1912-ben Oppenheimbe vitte a Teleki 4A fajtacsoportot, ebbıl szelektálta ki az SO4-t.
Külföldön a nagyon megbecsült Teleki alanyfajtákat jóval korábban klónozták, Magyarországon az alanyfajták szelekciója csak az 1960-as években kezdıdött.
Wädenswil-ben a svájci Szılészeti Kutató Állomáson a Teleki 5C-bıl szelektálták az 5C Wed.
klónfajtát, míg Geisenheim-ben a Gm.6 és Gm.10 klónokat.
Freiburgban a Teleki-Kober 5BB-bıl szelektálták a Fr.148-as klónt, a Weinsbergi Szılészeti Kutató Állomáson pedig a Wei.48-as klónt.
Ambrosi Romániában a Craciunal Blaj-i Kutató Állomáson szelektálta az 5BB-bıl a Cr.2 klónt (BAKONYI és KOCSIS, 2004).
Teleki Zsigmond munkássága világhírnevet vívott ki magának, hiszen a világ kötött talajú szılıinek 60-70%-a Teleki fajtacsoportból származó alanyfajtákra oltott ültetvény (BAKONYI et al., 1997 in KOCSIS és BAKONYI, 2000). Ennek magyarázata vitalitásuk, filoxérával szembeni toleranciájuk, mésztőrésük, jó affinitásuk és széles ökovalenciájuk (HAJDU és BAKONYI, 2006).
2.2. Affinitás, alany-nemes kölcsönhatás megnyilvánulási területei
Az affinitás az oltási komponensek közötti biológiai (morfológiai, biokémiai, fiziológiai) rokonságon alapuló szöveti egyesülésre és kölcsönös aktív együttélésre alkalmasság. Az együttélés a kölcsönös alkalmazkodás révén valósul meg (KOZMA, 1993). HEGEDŐS (1960) véleménye szerint kétféle affinitás van, amelyet egymástól el kell határolni: oltási affinitás, ami megismételt oltási kísérlettel eldönthetı és együttélési (valódi) affinitás. Utóbbi csak legalább 30 éven át tartó megfigyeléssel dönthetı el – ennek élettani, mégpedig anyagcsere forgalomban gyökerezı okai vannak. Együttélı képesség (affinitás) = kompatibilitás + adaptáció. Ez alatt nemcsak az alany és nemes kölcsönös kapcsolatát, kompatibilitását, hanem az ökológiai tényezık módosító hatására történı együttes reakcióképességet, adaptációt is értik (EIFERT et al., 1965).
HIDALGO és CANDELA (1973) szerint az oltott tıkék erıteljessége is lehet affinitási mutató.
TANDONNET et al. (2005) adatai alátámasztják a hipotézist, az alany-nemes között a genetikai komponensek kölcsönhatásáról, ami a talajtermékenységtıl függetlenül jelentkezik.
RIVES (1971) szerint ez a kölcsönhatás megnyilvánul, pl. a gyökerekbe való szárazanyag allokációban. A kölcsönhatáson belül mindkét biotípus szerepet játszhat.
Az alany és nemes funkcionális viszonyának megértéséhez segítséget nyújt DANIEL (1908 in KOZMA, 1993) funkcióképesség elmélete. A szılıoltványnál az alany rendeltetése az abszorpció, az alany által felvett vízben oldott tápanyagok fogyasztója pedig elsısorban a nemes rész. Jó affinitás és forradás esetén az abszorpciós képesség és a fogyasztási képesség azonos mértékő (KOZMA, 1993). Ha a forradás, az affinitás és az ökológiai feltételek kedvezıtlenek, az elégtelen ellátáshoz vezet, kiegyensúlyozatlansághoz és gátolja a fejlıdést (HIDALGO és CANDELA, 1973).
Ugyancsak HIDALGO és CANDELA (1973) állapították meg, hogy az alanyok jobban párologtatnak, mint a termı szılık, ill. az oltott szılık jobban, mint a nem oltottak. Az amerikai alanyfajták több klorofillt tartalmaznak, mint a Vitis vinifera fajták. A xantofill és karotin mennyisége arányos a klorofilléval, s ennek révén a szárazanyag, a levélzet és a termés mennyisége magasabb az oltott tıkéknél, mint a Vitis vinifera fajtáké saját gyökéren. Az oltott tıkék enzimatikus tevékenysége is módosul.
MISURENKO és SZAMBORSZKIJ (1971) véleménye szerint a Berlandieri x Riparia keresztezésbıl származó alanyokra oltott európai fajták nagyobb hozama és növekedése a transzspiráció és légzés intenzitás növekedésével kapcsolatos. BICA et al. (2000), valamint VASCONCELOS et al. (2005) magasabb transzspirációs és fotoszintetikus arányt figyeltek meg, pl. az 5BB-nél. VASCONCELOS et al. (2005) vizsgálatai szerint azok az alanyok, melyeknek magasabb a szárazság toleranciája, pl. 125AA – általában jobb fotoszintetikus teljesítménnyel rendelkeznek. A légzés intenzitása nemcsak a ráoltott fajtától függ, hanem az alany és ráoltott fajta kompatibilitásától is, tehát az oltási komponensek fiziológiai kompatibilitásának egyik mutatójaként fogható fel.
A hormonvizsgálatok szerint a citokininek fakadáskor befolyásoltak az alany által, kötıdéskor viszont már nem. KELLER (2005b) kísérletében a legnagyobb termést adó tıkék alanya (Ramsey – V.champini) a legtöbb citokinint termelte, és a legkisebb termésprodukcióhoz kötödı alany, az 5BB, a legkevesebb citokinint. Emellett a magas citokinin koncentráció párhuzamba vonható a koraibb fakadással, növeli a növekedési erélyt és a virágok számát.
Az oltványok növekedési és fejlıdési ritmusa is eltérhet a saját gyökerő növényekétıl.
Az oltványszılık hamarabb fordulnak termıre (ZSUKOV et al., 1973). Az oltványban megváltozik a saját gyökerő összetevık jellemzı ásványi tápanyagforgalma is. Az alany és nemes kölcsönhatás megnyilvánul a termékenységi koefficiens alakulásában, a terméshozamban és a termés minıségében (KOZMA, 1993).
Az alanyok hatását a nemesre a következıképpen összegzik STRIEGLER és HOWELL (1991): Az alanynak közvetlen, elsıdleges, ill. közvetett hatása van a nemesre. Az elsıdleges hatások a gyökérrendszer szerepébıl adódnak: vízfelvétel, tápanyag felvétel és transzlokáció, a növényi növekedés anyagainak szintézise és metabolizmusa, szénhidrát és aminosav tartalékok képzése – mindez növeli a hideggel szembeni rezisztenciát.
A közvetett hatás a tıke méret növekedésének módosítása, a termesztéstechnológia révén érvényesül.
2.3. Az alanyok gyökérrendszerének funkciója
A gyökerek szerkezeti támaszt nyújtanak, ill. a víz és tápanyag adszorpció szervei (MULLINS et al., 1992 in SMART et al., 2006). Itt folyik a növekedéshez szükséges szerves anyagok szintézise, a szénhidrátok (VERCESI, 1987, WHEELER és PICKERING, 2003) és nitrogén tartalmú anyagok raktározása – ami fontos a vegetáció indulásakor (WOLPERT et al., 2005).
A víz- és tápanyagok hasznosításában a Vitis fajon belül az alanyfajták és hibridek gyökérrendszerének mérete, horizontális és vertikális eloszlása fontos tényezı (VERCESI, 1987). A gyökérsőrőség kulcstényezı lehet a nemes növekedésére gyakorolt alanyhatásban (SWANEPOEL és SOUTHEY, 1989, WILLIAMS és SMITH, 1991) inkább, mint a gyökér mélység (SMART et al. 2006). MORLAT és JACQUET (1993) szoros összefüggést találtak a gyökér mennyiség és a föld feletti növekedési erély (vigor) között, amit BATES (2005) azzal magyaráz, hogy a nyugalmi idıszakban a gyökerek szénhidrát- és tápanyagraktárként mőködnek.
A gyökérszerkezet ugyan a genetikai sajátosságok következménye, de emellett a fizikai és kémiai talajadottságok, a talajmővelés módja és a talaj növény-egészségügyi helyzete mutat befolyást a szılı földalatti viselkedésére (VERCESI, 1987, SWANEPOEL és SOUTHEY, 1989, WILLIAMS és SMITH, 1991, SOUTHEY és ARCHER 1988, in SWANEPOEL és SOUTHEY, 1989). MORLAT és JACQUET (1993), valamint WHEELER és PICKERING (2003) emellett a felvehetı vízellátottság, vízintenzitás, penetrométeres talajellenállás fontosságát hangsúlyozzák.
ZSUKOV et al. (1973), valamint DAULTA és CHAUHAN (1980 in SWANEPOEL és SOUTHEY, 1989) is bizonyították, hogy nem csak az alany, de a nemes is hat a maximális gyökérkoncentráció elhelyezkedésére és a teljes gyökérrendszer méretére.
A növény föld feletti része közvetlenül befolyásolhatja a gyökér adszorpciót, különösen a vízfelvételt. A víz felhasználás hatékonyságának növelése – a nem szikes terülteken – elsısorban a nemestıl és kevésbé az alanytól függ (VIRGONA et al., 2003).
A transzspirációs aktivitás határozza meg a tömegáramlást is, vagyis azt az ionmennyiséget, ami a gyökerek felületén adszorbeálódhat. A gyökérszövetek növekedését az áthelyezıdött fotoszintetizátumok metabolikus hasznosítása determinálja. Nem kizártak ezen kívül
a hormonok esetleges áthelyezıdései a föld feletti részekbıl a gyökerekbe- ezek „feed back”
szabályozást gyakorolnak az ionok adszorpciójára (VERCESI, 1987).
A növekedés a szılı tápelem felvételének „pacemakere”, így a vízhiány okozta növekedés gátlás csökkenti a tápelem ellátottságot. A levél magas nitrát tartalma csökkenti a gyökér: vesszı arányt. A víz és/vagy N bıséges adagolása érzékenyebbé teszi a növényeket a stresszre, amikor egyik vagy másik tényezı hiányzik. A víz hiánya a vegetatív növekedésre nagyobb, mint a generatívra, de ez nem minden fenológiai stádiumra igaz. A víz és tápelemek hiánya csökkent termést eredményez különösen, ha a hiány a szezon elején jelentkezik (KELLER, 2005a). USSAHATANONTA et al. (1996) szerint is a vízstressz hatása nagyobb megfelelı tápanyag ellátás mellett, mint hiánynál (akár 10 nappal is kitolja az érést). Nincs általános válasz a stresszre, de a válasz függ a kiváltó tényezı milyenségétıl. Az alacsony tápanyagtartalom kívánatos a talajokban ott, ahol a többi tényezı erıs növekedésre serkent.
CARBONNEAU (1985) eredményei arra utalnak, hogy a gyökérrendszer adszorpciós kapacitása az elsıdleges a szárazsághoz való alkalmazkodásban.
A szárazsággal szemben legellenállóbb alanyok csoportja: 110R, 140Ru, 44-53 Ellenálló alanyok: P1103, 196-17, P1447, SO4, 99R, 7383
Kevésbé ellenállók: 3309, 420A, Fercal, 5BB, 41B, Rupestris du Lot Érzékeny: Riparia Gloire, Yuga, valamint nagyon érzékeny: a Vialla.
A vízdeficites növényekben a prolin felhalmozódása figyelhetı meg, melynek mértéke a szárazságtőrésre is utal (DEBRECZENI és DEBRECZENINÉ, 1983).
A szárazság ellen az egyes alanyok különbözı stratégiákat alkalmaznak. Némelyek csökkentik a lombfelületet (Riparia Gloire, Börner, 44-53M, 3309C), mások alacsonyabb szárazság stressz küszöbbel rendelkeznek (140Ru, 99R, 5C), míg néhány erıteljes növekedéső alany (1103-P, 125AA, 5BB, 1616C) képes fenntartani a stresszmentes vízállapotot (SAMPAIO és VASCONCELOS, 2005).
A gyökerek jobban fejlıdnek, ha a talaj jól ellátott mindenekelıtt nitrogénnel és foszforral. Az ásványi elemek eloszlása a talajszelvényben nem egyenletes, a foszfor és kálium, fıleg az agyagos talajokban a felszíni rétegekben található, míg a kalcium és magnézium a mélyebbekben (VERCESI, 1987). A talajhımérséklet növekedésével fokozódik a szerves foszfor ásványosodása, ill. száraz években az oldható kálium tartalom megnövekszik a talajban. Az idıjárás legfontosabb hatása az oldható tápanyagok kimosódásában mutatkozik meg (DEBRECZENI és DEBRECZENINÉ, 1983).
Nemcsak a talaj hat a gyökerek fejlıdésére a gyökérszerkezetek is képesek befolyásolni a talaj tulajdonságait a közvetlen szomszédságukban (poliszacharid természető mézgák, savak és enzimek kiválasztásával). Továbbá a talaj mikrobaflórája is, elımozdítja a tápelemek szabaddá válását és ennek révén új elemek jelenlétét a gyökérfelületeken.
Különösen fontosnak tőnik a szılıalanyok citromsav és huminsav kibocsátási képessége, pl. a vas
adszorbeálására. A kationok (NH4, K, Ca, Mg, Fe, Zn, Cu, Mn) felvételét akadályozza a talajkolloidokon való konzisztens megkötıdésük (amelyek negatív felületi töltésőek), addig az anionok (NO3, PO4, BO3) kisebb mértékben visszatartottak (VERCESI, 1987).
VERCESI (1987) a víz és tápelemek adszorpciója kapcsán megállapítja, hogy az számos tényezıtıl függ, amelyek magukra a gyökérszervekre (morfológiai és fiziológiai jellemzık), a talajra (a víz és tápelemek hozzáférhetısége, hımérséklet stb.), valamint a föld feletti részre (fotoszintetizáló- és transzspirációs képesség) terjednek ki. Ez a három tényezı többé-kevésbé erısen egymásra hat és nagyon konzinsztens módon befolyásolhatják ıket az éghajlat, az alkalmazott termesztés-technika és az esetleges kórokozók.
2.4. Alany-nemes kombinációk hatása
2.4.1. Alany-nemes kombinációk hatása a tápelemek felvételére
A témával kapcsolatos szakirodalom egységes abban, hogy az oltványtıke tápelem ellátottsága mind az alany-, mind a nemesfajta által befolyásolt. Ugyanakkor az együttélésben egyes szerzık a nemesfajták szerepét hangsúlyozzák, míg mások az alanyfajták jelentıségét emelik ki. DELAS és POUGET (1988 in AVENANT et al., 1997), valamint BERGMANN (1979) véleménye szerint a nemes tápelem státusza az alany, ill. a gyökér (MIKLÓS et al., 1985) tápelem adszorpciós képességének eredménye, és a nemes transzportáló képességéé. Egy olyan fajta, amely az adott elembıl kevesebbet halmoz fel, mindig alacsonyabb értéket mutat, mint más fajták – akkor is, ha olyan alanyra van oltva, mely ebbıl az elembıl nagy mennyiséget vesz fel. Az erıs tıkék jobban ellátottak tápelemekkel, mint a gyengék – de kérdés, hogy ez ok vagy következmény (DELAS és POUGET, 1979). BOVAY és GALLAY (1956, in CSEPREGI és ZILAI, 1961) svájci kutatók 13 alanyfajtának a Chasselas-ra gyakorolt hatását vizsgálták levélelemzéssel. Szerintük az egyes alanyfajták azonos környezeti viszonyok között a rájuk oltott fajta táplálkozását jelentısen befolyásolják.
Makroelemek
A csoportba tartozó elemeket (N, P, K, Ca, Mg) nem az adott tápelem élettani jelentısége alapján, hanem a növények számára szükséges mennyiség alapján tekintjük makroelemeknek.
A tápelemek élettani szerepe és biokémiai tulajdonságai alapján az elemek négy csoportba sorolhatók (DEBRECZENI B.-né in FÜLEKY Eds., 1999). Az elsı csoportba tartozó C, H, O, N és S a növényeket felépítı legfontosabb szerves vegyületek szerkezeti elemei. A második csoportba sorolt P, B és Si elemek ionos formában a talajoldatból kerülnek felvételre. A szerves alkoholos csoportokkal észtereket képeznek, amelyek részt vesznek az elektron átviteli reakciókban. A harmadik csoport elemei: K, Na, Mg, Ca, Mn és Cl. Enzimekre nem specifikus hatásúak. Részt vesznek a növények vízháztartásának szabályozási folyamataiban, az ozmoregulációban, könnyen
helyettesíthetik egymást. A 4. csoport elemei: Fe, Cu, Zn és Mo – az elektrontranszportban is közremőködnek, fıleg a vegyértékváltók.
Nitrogén
A nitrogén elsıdleges alkotója a következı fontos növényi összetevıknek: fehérjék, enzimek, nukleinsavak, klorofill és vitaminok. A szılınövényben a N létfontosságú a szılı növekedéséhez és fenntartásához, a termés minıségéhez, s ennek következtében a must és bor minıségéhez is. A N módosítja a növény összetételét, sokkal inkább, mint más ásványi elem.
Amennyiben a nitrogén ellátás nem megfelelı: a növény növekedése gyenge, az idısebb levelek korai elöregedése következik be, limitált a termırügyek kialakulása, és csökken a termés mennyisége. A hozzáférhetı N alacsony szintje hat a termés minıségére is, de a túlzott ellátottság is gond (KELLER, 2003). A szılınövény nitrogén állapota függ a genetikai tényezıtıl, a környezeti feltételektıl és az alkalmazott termesztéstechnológiától (ROUBELAKIS-ANGELAKIS és KLIEWER, 1992).
KOBLET et al. (1996) Müller Thurgau fajtán vizsgálták az alanyhatást nitrogéntrágyázással, ill. a nélkül. A trágyázatlan kezelésben a legnagyobb fotoszintetikus arány az 5BB- hez kötıdött. A trágyázatlan kezelésben a legalacsonyabb arányt az SO4 képviseli, míg a trágyázottban a legmagasabbat. KELLER et al. (2001a) szintén a Müller Thurgau fajtát vizsgálták a következı alanyokon: 5BB, 5C, 8B, SO4, Couderc 3309, Ruggeri 140. Az alanyok hatása érvényesült a talaj nitrogén tartalmára adott nemes reakcióra, így a levél klorofill tartalom és a levél gázcsere alakulására, ugyanakkor a nemes nitrogén ellátottsági állapotában az alanyhatás nem eredményezett szignifikáns különbséget, bár az irodalomban vannak ezzel ellentétes tapasztalatok is. Az 5BB alanyon a legnagyobb a levelek klorofill tartalma. Általában a nagyobb növekedési erélyő alanyok magasabb N értékeket eredményeznek a nemesben. Ebben a kísérletben ezt kevésbé tapasztalták, de a Berlandieri x Riparia keresztezések magasabb fotoszintézis értéket eredményeztek, mint a többi. A Berlandieri x Riparia alanyok úgy tőnik az alacsony nitrogénszolgáltatású talajokon is jól szerepelnek, kivéve az SO4-et – mely alacsony N tartalom mellett kevésbé teljesít, de jól reagál a N adagokra, ahogy azt KOBLET et al. (1996) is megfigyelték. Az alanycsaládon belüli hasonlóságok mellett az eltérések lehetıségére utal COUSINS (2005).
SARIČ et al. (1977) szerint a nitrogén felvételt és tartalmat a nemes jobban befolyásolja, mint az alany.
Foszfor
A foszfát az energia metabolizmus fı eleme, és a membrán szintézisé. Fontos szerepet játszik a fotoszintézisben, légzésben, számos enzim regulációjában, így a foszfor éhség csökkenti
a fotoszintézist (KELLER, 2005a). A szılı foszfor hiánya gátolja a fürtök, virágok kialakulását és fenntartását – ennek eredménye a csökkent termés.
Az alanyok különböznek abban a képességükben, hogy a talajból mennyire hasznosítják a foszfort, a gyökérbıl a vesszıbe való transzlokációban, és abban, ahogy a nemes foszfor hasznosítására hatnak (GRANT és MATTHEWS, 1996 in GRANT, 1999).
Több kísérletben nagy különbségeket találtak a levelek foszfor tartalmában, bár azok igen alacsony értékek, de úgy tőnik, hogy a foszfor szintekre a nemesnek van nagyobb hatása (KOCSIS et al., 2001, SARIČ et al., 1977).
Kálium
A kálium számos enzim aktivátora, a protein szintézisben, a víz regulációban van szerepe, szerves savak semlegesítıje, a kation - anion egyensúlyban vesz részt (BALOGH és TÓTHNÉ, 2000). FARDOSSI (1991) szerint a K a mustok sav, ill. pH, valamint a szín és a minıség alakításában vesz részt. A cukortartalom és bogyóméret alakításában a csemegeszılıknél a kálium fontos elem.
Az alanyfajták hatását a kálium akkumulációra és a K státuszára a következıkben összegezhetjük:
- Az alanyfajták között különbség van az elemek, így a K adszorpciója tekintetében.
- A levélnyél és/vagy must kémiai összetételében különbségek vannak: különbözı nemes fajták között- azonos alanyon, ill. azonos nemesnél különbözı alanyokon.
- A kálium adszorpciós és akkumulációs képesség számos alanyfajtánál negatív összefüggést mutat a magnézium adszorpciós képességgel.
- Az alanyfajta hat a must pH-ra - a nemes K akkumulációjára gyakorolt hatásán keresztül.
- A különbözı alanyfajták kálium akkumulációs képességében lévı különbségek genetikai eredetüknek tulajdoníthatóak (AVENANT et al., 1997, RUHL, 1989, 2000).
A kálium és bór felvételében az oltványok hatékonyabbak, mint a saját gyökéren lévı szılıtıkék (CANDOLFI-VASCONCELOS et al., 1997).
BRANCADORO et al. (1995) 20 alanyon és saját gyökéren vizsgálták a Croatina fajtát.
Minden alany szignifikánsan hatott a levelek és a must K tartalmára. A legmagasabb szinteket a 44- 53 M és SO4 alany mutatta, a legalacsonyabbakat a 140Ru, 420A és az 1202C.
Az SO4 alany révén elért magasabb K tartalomról számol be POLYÁK et al. (1986) is. A tápoldat (N,P,K) tápelem koncentrációjának és a növény tápelem tartalmának a növekedése csökkentette a vesszık kalluszképzı képességét – leginkább a K koncentráció növekedése révén.
A talaj magas K tartalma feltételezi a termés magas K szintjét is. Az alanyok hatnak az oltványszılık kálium tartalmára (SARIČ et al., 1977, FARDOSSI et al., 1990), de ez nem minden esetben igazolható (WOOD és PARISH, 2003). DELAS és POUGET (1979) pl. a kálium tartalom tekintetében a ráoltott fajta szerepét tartja meghatározónak a kombinációban.
Kalcium és magnézium
MIKLÓS (1994) feltételezése szerint a levél Ca2+ tartalom, ill. a K+/ Ca2+ arány összefüggésben lehet a mésztőréssel. A jó mésztőrı nemes- és alanyfajták intenzív kalcium akkumulációval reagáltak a megnövekedett külsı kalcium ionkoncentrációra. Ez a védekezési mechanizmus hasonlít a sótőrı növények nátriumion levélbeli akkumulációjához.
CABANNE és DONÉCHE (2003) szerint a kalcium tartalom kapcsolatban van a betegség ellenállósággal, pl. a bogyóban a magas kalcium tartalom növeli a szürkerothadás ellenállóságot.
A bogyók kalcium tartalma függ biológiai (fajta, alany), talaj (felvehetı kation és víztartalom), és klimatikus tényezıktıl. Fajtától és évjárattól függıen módosul a kalcium tartalom.
A tıke érzékenysége a magnézium hiányra függ az alanytól, ugyanígy a kocsányszáradással szembeni érzékenysége is, ami évente eltérı erısségő lehet. A kocsány száradás azokon az alanyokon volt a legnagyobb mértékő, amelyeknek a levélnyele a legkevesebb magnéziumot tartalmazta. DELAS és POUGET (1979) kísérletei alapján az SO4 alany magnéziumtrágyázásra nem nagyon változtatja a levél ásványi összetételét, sem a kocsányszáradásra való hajlamát.
SARIČ et al. (1977) úgy vélik, hogy a levél és vesszı Mg tartalmát a nemes inkább meghatározza, mint az alany, a kalciumnál viszont fordított a helyzet.
Cink
MARSCHNER és SCHROPP (1977) 143A, M.G., 125 AA, 26 G., 5C, 5BB, 8B alanyokkal állítottak be tenyészedényes kísérletet. A cink hiányos talajban a növekvı foszfor trágya adagok mellett a fiatal levelek Zn tartalma minden alanyon csökkent. A vesszınövekedés extrém csökkenése és a Zn hiánytünetek megjelenése leginkább az 5BB és 8B alanyokra volt jellemzı. A vízkultúrás kísérletben, ezzel szemben, a növekvı foszfor adagok nem vezettek a Zn tartalmak csökkenéséhez. Egyik alanyon sem jelentkeztek hiánytünetek, de a foszfor tartalmak a levelekben nagyobb mértékben nıttek, mint a tenyészedényes kísérletben. A P/Zn arány itt 1000 fölötti volt, cink hiánytünetek megjelenése nélkül. Véleményük szerint kedvezıtlen környezeti feltételek mellett, amikor a gyökérnövekedés gátolt az alacsony fényintenzitás miatt, az 5BB és 8B alanyfajták érzékennyé válhatnak a cink hiányára.
Több tápelem együttes vizsgálata
Számos szerzı számol be az alanyok hatásáról az oltványtıkék tápelem állapotának vizsgálata kapcsán. Az egyes kísérletek eredményei azonban rendszerint nem vethetık össze, mivel a tápanyag ellátottságot befolyásoló körülmények és a vizsgálati módszerek is eltérıek. Az
eredmények reprodukálhatósága a szerint is eltérı, hogy vízkultúrás, tenyészedényes vagy szabadföldi kísérletek eredményeirıl számolnak be.
FARDOSSI et al. (1995) vízkultúrában vizsgáltak 10 különbözı alanyon lévı (5BB, 5C, 125AA, 8B, 26G, Fercal, 1G, 725 P, 1103P, Riparia Sirbu) Zöld veltelini fajtát. A levél ásványi elem tartalmát - a K, Ca, Mg, P, Fe, Zn és Mn elemekét - befolyásolta az alany.
DELAS (1979) kísérletében SO4 alanyon a levelek foszforban, Mg-ban és fıleg Ca-ban gazdagabbak voltak, mint 3309C vagy Riparia alanyon. A saját gyökéren lévı Merlot az SO4-hez hasonlóan jó tápelem ellátottságot mutatott. FARDOSSI et al. (1990) viszont egyrügyes dugványok vizsgálatakor azt tapasztalta, hogy az SO4 a legalacsonyabb Ca, P és Mg felvételt eredményezte.
FARDOSSI et al. (1991) Zöld veltelini szılıfajtán (tenyészedényes kísérletben) az 5BB, 5C és SO4 alanyok összevetése kapcsán a következı eredményekrıl számol be:
K-tartalom: Az 5BB esetében a legmagasabb, majd 5C és SO4 alanyokon közel azonos érték.
Ca és Mg: Az 5BB alanyon a legmagasabb, de minimális a különbség a vizsgált alanyok között.
P, Zn, Mn: Az 5BB-nél a legmagasabb, a másik két alany között nincs számottevı eltérés.
Fe: 5BB, SO4, 5C a sorrend, de mindegyik magas érték.
TARDÁGUILA et al. (1995), valamint KOCSIS et al., (2001) statisztikailag igazolt különbséget találtak a vizsgált alanyok között a levelek N, P, K, Ca és Mg szintjeinél.
CONRADIE (1983) eredményei szerint az oltványok levelében a legkisebb különbség az alanyok szerint a N tartalomban mutatkozott. Ugyanakkor véleménye szerint, bár úgy tőnik, hogy az alanyok nem befolyásolják úgy a N felvételét (amit a levél koncentrációval mérünk), mint a többi makroelemét, mégis a különbözı alanyok eltérı N hasznosítása lehet annak az oka, hogy azonos feltételek között a különbözı alanyok teljesítménye eltérı. A vizsgált alanyok: 99R, 101-14 Mgt, 140Ru voltak – homok kultúrában, tenyészedényben, a mintákat zsendüléskor értékelték. A gyökér és hajtásfejlıdés is a 99R-en volt a leggyengébb, ennek ellenére a növények ugyanannyi nitrogént adszorbeáltak.
KOCSIS et al. (2001) kísérletei alapján a tápelem felvételre az alany-nemes kölcsönhatásnak volt a legnagyobb hatása, továbbá a száraz körülmények szintén erısen befolyásolták az elemfelvételt, a normál évjáratú évekhez képest.
FARDOSSI et al. (1993a, b) az 5BB, 5C, 140Ru, 3309C, Fercal alanyok és klónok szabadföldi vizsgálatakor értékelték a Tramini fajta levéllemezének tápelem tartalmát 1991-1992-ben (a fürtzónából vett mintákon). Az 1992-es extrém száraz év szignifikáns csökkenést eredményezett a K, P, Fe, Zn és Mn elem tartalomban 1991-hez képest, megfelelı talajellátottság mellett. Az öregedı levelekben kedvezı volt a Ca és Mg akkumuláció. Ez diszharmóniát eredményezett a tápelem arányoknál, fıleg a Ca-K viszonyánál.
POLYÁK et al. (1986), valamint POLYÁK (1992) az SO4 rossz kalluszképzését is a diszharmonikus tápelem ellátottság következményének tartják. Azt tapasztalták, hogy minél
magasabb volt az alany levelének és vesszıjének K tartalma, annál kevesebb volt annak Ca és Mg tartalma.
A tápelemek felvételére gyakorolt alany sajátosságokról összegzı áttekintést kapunk VERCESI (1987) munkájából:
Riparia x Rupestris: Ca és P felvétele gyenge, a K-t jól hasznosítja
Berl. x Rupestris: általában jó tápelem felvétel jellemzi, fokozott K, viszont gyenge Mg hasznosítás Vinifera x Berlandieri: K és Mg ellátottság jó, de a Ca gyenge
K hiánnyal szembeni érzékenység: a 110R nagyon érzékeny, az SO4 és 5BB közepesen, a Rupestris du Lot rezisztens
Mg hiánnyal szembeni érzékenység: az SO4 nagyon érzékeny, a 125AA, 5BB és 140R közepesen, a Rup.du Lot rezisztens.
Megállapíthatjuk, hogy az alanyoknak tulajdonított tápelem felvétel valójában többnyire az alany-nemes kombináció sajátossága, s így ellentmondásossá válhat egy-egy alany megítélése az irodalom alapján a tápelem felvétel tekintetében. Szabadföldi viszonyok között a termıhelyi adottságok és az ökológiai viszonyok jelentıs hatása miatt az alanyhatás is módosul. A vízkultúrás kísérletek megállapításai pontosabb információt nyújtanak az alany tápelem felvételérıl, viszont a gyakorlatban érvényesülésük bizonytalan, a környezet x alanyhatás ilyen módon nem mérhetı.
A növényben a tápelemek nem önmagukban, hanem egymással kölcsönhatásban fejtik ki hatásukat. SKINNER és MATTHEWS (1990) megállapítják, hogy a talaj alacsony foszfor koncentrációja mellett a fotoszintézis mérétke a levelek magnézium koncentrációjának függvénye.
Kísérleteik eredményei azt mutatják, hogy a magnézium transzlokációja a gyökerekbıl a szılıvesszıbe függ a gyökerek foszfor ellátottságától, valamint a foszfor adagolás a magnézium transzlokációját érzékenyebben érinti, mint a felvételt. A magnézium felvételét erısen gátolják más kationok, pl. a káliumion. Foszfor hiányos, magnéziummal megfelelıen ellátott talajon a P-Mg összefüggés fennállása elegendı ahhoz, hogy magnézium hiány tünetek lépjenek fel és a fotoszintézis magnézium limitált legyen.
A bór esszenciális elem az ATP pumpa mőködéséhez, így a B hiánya erısen csökkenti a gyökerek nitrát felvételét (CAMACHO-CRISTÓBAL és GONZÁLEZ-FONTES, 1999 in KELLER, 2005a). A visszafogott N felvétel a bórhiányos tıkéknél alacsony levél N tartalmat eredményez és alacsony cukor és keményítı akkumulációt a levelekben. Ugyanakkor a bıséges nitrogén ellátás csökkentheti a foszfor felvehetıségét (KELLER, 2005a).
Pozitív kapcsolat van a K/N+P és az alany életképessége között. A klorózis a Ca+P mennyiségével kifejezetten szoros összefüggésben van. Minél nagyobb az említett két elem mennyisége, annál nagyobb a kockázat különösen, ha a nemes is klorózisra érzékeny, mint pl. a Cabernet sauvignon fajta is (KISS, 1987).
2.4.2. Alany-nemes kombinációk hatása a tápelemek megoszlására a növényen belül TARDÁGUILA et al. (1995) az alany indukálta növekedési erélynek tulajdonítja, hogy néhány alany elsısorban a vegetatív részek tápelem koncentrációjára hat, míg mások a reproduktívakra. Kísérletükben az alany befolyásolta a N koncentrációt a fiatal hajtásban, a levelekben és a levélnyélben, de a fürtökben és a fásodott vesszıben nem.
WILLIAMS és SMITH (1991) szerint viszont az alanyoknak minimális a hatása az egyes szervek N és K koncentrációjára. Ugyanakkor a gyökerek alacsony nitrogén tartalma a fürt alacsony N tartalmát eredményezte.
WERMELINGER és KOBLET (1990) vizsgálatai alapján a szárazanyagnak nagyobb része kerül át a vesszıbe, mint a levélbe, de több N található a levelekben, mint a vesszıben. A szárazanyag és a nitrogén mennyiség a termésben közel azonos arányban található.
A vizsgálat idején a fürtben akkumulálódott N a korábban a gyökerekben és idıs fás részekben asszimilált N-bıl származik (KELLER, 2005a).
CONRADIE (1983) kiszámította, hogy a zsendülés elıtti 4 hétben a fürtök fejlıdéséhez szükséges N-nek mintegy ¾ része származik korábban asszimilált N készletekbıl.
WOLPERT et al. (2005) megállapították, hogy az alany-nemes kombinációk virágzáskor jelentısen eltérı K koncentrációt mutatnak, és a K újraelosztása lényeges szerepet játszik a K ellátásban, hasonlóan a nitrogénhez. A kálium adszorpció tavasszal fakadástól-virágzásig, kritikusabb idıszak lehet a K beszerzésben, míg a retranszlokáció a szövetekbıl az érı termésbe elsıdleges forrást jelent az érés idején.
A K felhasználás tekintetében verseny van az egyes szervek között, de éréskor a termés az elsıdleges felhasználó. Az elöregedı (érett) levelek kipumpálják az oldott tápelemeket, így a K-t.
Amennyiben adott idıben a termés a legnagyobb felhasználó, akkor az válik a legnagyobb K raktárrá (WOOD és PARISH, 2003, CONRADIE, 1981, SMART, 1985).
Az alany hatását a K eloszlására magyarázhatja, pl. a vegetációs idıszak alatt a K felvétel eltérı aránya. A hajtásvég növekedésének intenzitása és perzisztenciája képes versenyezni a fürttel a káliumért, a levélöregedés szabályozása révén a kálium transzlokációja a levelekbıl a fürtbe történik (FAILLA et al., 1990).
WILLIAMS és BISCAY (1991) Cabernet sauvignon-t vizsgáltak 5C alanyon, szabadföldön.
Vizsgálataik alapján a fürt K tartalmának növekedése – az érés másfél hónapja alatt - 75%-ban a föld feletti szervekbıl történı K átáramlás eredménye.
A kálium megoszlása a levél és fürt között függ a fajtától, a tıke termésszintjétıl, a szüreti idıponttól, a talaj K és vízszolgáltatásától (FAUST, 1980 in FAILLA et al., 1990).
KLEIN et al. (2000) a kálium és foszfor relatív csökkenését a levélnyelekben és a levéllemezben virágzástól érésig jó indikátornak tartják e két tápelem állapota, státusza
A fejlıdés fenológiai állapotától függ a kálium tartalom. Ennek kapcsán WOLPERT et al. (2005) megállapítják, hogy a teljes K szint általában szignifikánsan csökken a virágzás és érés között minden általuk vizsgált fajtán (Chardonnay, Cabernet sauvignon, Zinfandel), a csökkenés mértéke alanyfüggı. A Vitis berlandieri genetikai háttérrel rendelkezı alanyok virágzáskor alacsonyabb K szinteket mutatnak a levélnyélben, de ezek a különbségek nem nyilvánvalóak zsendüléskor és éréskor.
RÜHL (2000) véleménye szerint a vesszık különbözı K akkumulációs arányáért felelıs mechanizmus helye a gyökér. Ez is arra utal, hogy a raktározott K allokációja végbemegy a szükségleteknek megfelelıen a vesszıkbe, a kiegyensúlyozott tıke eltérı választ adhat, mint a kiegyensúlyozatlan (WOOD és PARISH, 2003).
2.4.3. Alany-nemes kombinációk hatása a (vegetatív és generatív) terméshozamra NUZZO és MATTHEWS (2006) úgy ítélik meg, hogy nem kellıen tisztázott az alanyok szerepe a reproduktív fejlıdésben. Az eredmények értékelését nehezíti, hogy a kísérletek eltérı körülmények (öntözés, trágyázás) között zajlanak. Az alanyok elsısorban az oltványtıke növekedési erélyét, továbbá a termés mennyiségét befolyásolják. Kevésbé hatnak a must minıségi paramétereire – a cukortartalomra és a titrálható savtartalomra (MAY et al., 1973, BLAHA, 1975, DOBROVODA, 1975, GAPRINDASVILI et al., 1980, GAPRINDASVILI, 1982, CIRAMI et al., 1984, CRESCIMANNO et al., 1984, OCHABA, 1987, 1988, AVENANT et al., 1997). Az alany közvetlenül hat a gyökérfunkcióra, a tıkeméretre is (BOGONI et al., 1996).
ZAHAROVA (1956 in KOZMA és POLYÁK, 1973) kísérletei igazolták, hogy az oltványszılık nagyobb terméseket adnak, mint a saját gyökerőek. KOZMA és POLYÁK (1973) tenyészedényes kísérletei szerint az oltványtıkék három éves átlagban kétszer annyi termést adtak, mint a saját gyökerőek. A terméseltérések okaként szolgál a saját gyökerő tıkék kisebb kezdeti fejlıdése, gyöngébb kondíciója és kisebb terhelhetısége (KOZMA és POLYÁK, 1965, KOZMA et al., 1969).
Az alany hatása a különbözı nemes fajták teljesítményére nagyon változó. Egyetlen fajta nem nyilvánítható általánosan jó alanynak minden nemeshez és minden talajra (DOBROVODA, 1975). Az adott fajtának és környezeti viszonyoknak legjobban megfelelı alanyfajta megválasztása fontos termesztési kérdés, s végsı soron a termesztés gazdaságosságára van hatással (CSEPREGI és ZILAI, 1961).
A területegységre esı termésmennyiség minıségromlás nélküli növeléséhez, elsı teendı az alanyfajta helyes megválasztása. EIFERT et al. (1965) megállapították, hogy az oltványtıke kondíciójának, élettartamának és termésmennyiségének szabályozásában elsıdleges szerepe az alanyfajtának van. McCARTHY és CIRAMI (1990) szerint az alanyok származása befolyásolja a nemes fürttermés mennyiségét és a vesszısúlyt.