T H E Y B E A C C U R A T E L Y D E T E R M I N E D ?
M U R R A Y D . R O S E N B E R G
Department of Zoology,
University of Minnesota, Minneapolis, Minnesota
I t is u n f o r t u n a t e t h a t t h e r a p i d accumulation of information during t h e p a s t two decades on t h e ultra-structure of cells h a s n o t led t o equally vigorous efforts to observe and measure t h e movements of m a t e r i a l s along, among, a n d by t h e peripatetic structures found in k a r y o p l a s m and cytoplasm. On t h e other h a n d , it is u n d e r s t a n d a b l e t h a t only a v e r y limited n u m b e r of investigators h a v e u n d e r t a k e n research directly applicable to such problems in view of t h e sparsity of experimental techniques and plethora of speculative models. P e r t i - nent information and d a t a p e r t a i n i n g to t h e processes a n d mechanisms of "intracellular t r a n s p o r t " are lacking a n d few a t t e m p t s h a v e been m a d e to ascertain how molecular species move within t h e cell from one specific location to another. This p a p e r a t t e m p t s to outline and categorize several subheadings of the topic, to establish p e r t i n e n t frames of reference or b o u n d a r y conditions set by experimental m e t h - ods, a n d to review some underlying physical a n d chemical mechanisms.
T o p u t t h e problem into numerical perspective the following a p - proximations m a y be useful. A mononucleate a n i m a l cell 1 5 μ in di- a m e t e r weighs roughly 2 X 1 0 ~
9
gm. P r o t e i n molecules can account for 4 χ 1 0 ~
10
g m ; if t h e cell were to divide with a generation t i m e of 2 4 hours it would h a v e to synthesize 3 0 , 0 0 0 protein molecules (of average molecule weight 1 0
5
) per second. I n other words, t h e cell must be capable of assembling in each second roughly 3 0 Χ 1 0
6 amino acids in proper sequence and in relevant locations. A t t h e same t i m e all molecular species, such as phospholipids, lipids, polysaccharides, must not only be assembled in duplicate m a n n e r , b u t also be properly located into formed elements such as m e m b r a n e s , tubules, fibrillar units, and so forth. N o a d e q u a t e estimates h a v e been m a d e of t h e t o t a l surface area of mitochondrial, ergastoplasmic, lysosomal, and other cellular m e m b r a n e s , or of t h e m a s s of microtubules or of identi- fiable filamentous structures. T h e r e is every indication, however, from
4 5
electron microscopy, cytochemical analyses, a n d isolation studies t h a t these quantities can be v a s t per cell unit. W e r e t h e assumed cell a bacterium 1 μ in diameter with a generation time of 30 minutes, it would h a v e t o synthesize roughly 1000 protein molecules per second.
On the other h a n d , giant multinucleate slime molds several centimeters in dimensions, in which nuclear division occurs almost simultaneously every 30 minutes, possibly face far greater problems of synthesis and assembly. I n general, depending upon t h e cell t y p e , t h e processes of rapid division m a y entail t h e t e m p l a t e - d e p e n d e n t synthesis of up to a half million protein molecules per second, t h e accumulation each second of m a n y millions of ions and similar small molecules, t h e enzymatic synthesis of c a r b o h y d r a t e s and fats, t h e self-duplications of nucleoproteins, a n d t h e establishment a n d m a i n t e n a n c e of s t r u c t u r a l organization on t h e molecular a n d macromolecular levels. One can readily u n d e r s t a n d w h y proliferation and specialization are almost m u t u a l l y exclusive cellular activities. I t seems as though either t h e p r e p a r a t i o n for mitosis or t h e carrying out of differentiated functions, such as secretion, contraction, a n d impulse conduction, fully occupies the synthetic c a p a c i t y and, w h a t m a y be equally i m p o r t a n t , t h e orga- nizing or structuring capabilities of certain cell t y p e s . Among problems t o be faced in future research are t h e mechanisms of intracellular t r a n s p o r t most probable, necessary, and sufficient for molecular traffic and structuring of such orders of m a g n i t u d e , a n d w h e t h e r current experimental methods provide sufficient accuracy for their determination.
An example of t h e organized circulation of molecules in protoplasm under t h e control of regulatory mechanisms, inhibitors, a n d accelera- tors h a s recently been given by Bessis [4] in his studies on hemo- globinopoiesis. Figure 1 illustrates t h e process. Glycine a n d succinic acid enter t h e cell via specific receptors in t h e cell surface. I r o n enters via pinocytosis and specific acceptors. A succinyl-coenzyme complex is formed within mitochondria. Simultaneously, ribosomes receive from t h e nuclear region messenger ribonucleic acid ( R N A ) carrying nuclear codes and t h e a c t i v a t e d amino acids bound to transfer R N A . Protein globin is synthesized a t t h e ribosomes. I n some u n k n o w n m a n n e r t h e precursors for heme are assembled within t h e cytoplasm and only interact with t h e globin in t h e vicinity of t h e Golgi a p p a r a t u s . F r o m t h e Golgi a p p a r a t u s hemoglobin is finally liberated into cytoplasm.
This is a simplified picture, one t h a t can be duplicated with respect to t h e synthesis of n u m e r o u s cell products. I t points out, however,
in conjunction with t h e figures given earlier for t h e r a t e s of synthesis, t h e immense a c t i v i t y a n d numerous w a y stations t h a t m a y be required within a single cell-type for t h e synthesis, secretion, a n d possibly d e g r a d a t i o n of a specific molecule; this process is b u t a small p a r t of t h e n o r m a l activity of t h e cell. Other examples of directed m o v e - m e n t s h a v e been d e m o n s t r a t e d by C a r o a n d P a l a d e [6] for t h e secre- tion of zymogen granules by t h e exocrine cells of t h e p a n c r e a s , by D e D u v e [12] for the formation a n d dissolution of lysosomes, a n d by Hirsch and Cohn [19] for phagocytosis by macrophages.
Before entering into a discussion of how such itineraries are e s t a b - lished, m a i n t a i n e d , halted, and regulated, one additional set of facts should be reviewed briefly, n a m e l y , t h e velocities a t which protoplasm h a s been observed to s t r e a m within a cell, t h e r a t e s a t which various cell organelles h a v e been observed to move within a cell, a n d t h e directed m o v e m e n t s of m a t e r i a l s within specific cell t y p e s . T h e o b - served r a t e s are presented in T a b l e I. T h e velocities are v e r y approxi- m a t e a n d v a r y tremendously with t h e conditions of t h e experiment, metabolic s t a t e of t h e specimen, etc. I t is a p p a r e n t t h a t t h e r a t e s of specialized m o v e m e n t s of m a t e r i a l s , fluid, a n d organelles within cells encompass a range roughly 105, a range t h a t poses considerable FIG. 1. Hypothetical paths for hemoglobin precursors in erythroblast. F r o m Bessis [4].
problems if one wishes t o singularize t h e mechanism for intracellular movement.
T h e above q u a n t i t a t i v e a n d observational d a t a are presented to p u t t h e problem in general perspective. T h e cellular traffic problem m a y concern t h e r e a r r a n g e m e n t of p e r h a p s a t h o u s a n d to a half mil- lion macromolecules per second; in some cases molecules are not homogeneously distributed t h r o u g h o u t t h e cell ; certain molecules com- bine into larger aggregates or complexes only in specific regions of
TABLE I. Cytoplasmic Flow Rates
Axoplasm 0.01
Plasmalemma ruffling and plication 0 . 0 1 - 1 Mitochondrial m o v e m e n t s 0 . 0 1 - 1 Saltatory m o v e m e n t of granules
Arbacia punctulata 0 . 6
Spisula solidissima 1-5
fish skin melanocytes 5
Protoplasmic m o v e m e n t s
Foraminifera 5
Cyphoderia ampulla 5
Diplogromia 8-10
Amoeba proteus 1-50
Rotational streaming in p l a n t cells 60
Chara braunii 60
Nitella flexilis 60
young internodal cell 10
Euphorbia marginata 0 . 7 - 1 0 Streaming in slime molds
Didymium nigripes 20
Physarum polycephalum 100-1300
t h e cell; in some cases small fluid vesicles or c o m p a r t m e n t s provide the t r a n s p o r t mechanism for molecules in solution; and t h e observed m o v e m e n t of cytoplasmic m a t r i x , organelles, or c o m p a r t m e n t s m a y v a r y from essentially a static condition to flows as rapid as 1300 μ per second.
I n discussing some of t h e mechanisms t h a t underlie intracellular t r a n s p o r t as well as t h e role of such t r a n s p o r t in cell function, it is useful initially to m a k e an artificial or a r b i t r a r y distinction between
(a) t h e movements of molecules within t h e channels of ergastoplasm or via pinocytotic vesicles, and (b) t h e movements of molecular spe- cies within and among cytoplasmic m a t r i x a n d its components. As
noted in T a b l e I I t h e first t y p e of t r a n s p o r t encompasses t h e process of ingestion a n d egestion of molecules a n d macromolecules b y m e m - brane plication—infolding and outfolding. I t concerns both p r e s u m - ably p e r m a n e n t ergastoplasm a n d short-lived, t e m p o r a r y pinocytotic and phagocytotic vesicles. I t m a y be t e r m e d "quasi-intracellular t r a n s p o r t " or " e x t r a - i n t r a c e l l u l a r t r a n s p o r t . " T h e second t y p e of t r a n s - port includes traffic from nuclear regions to cytoplasm and ribosomes, from mitochondria to other surfaces, from ribosomes to cytoplasmic m a t r i x , t h e m o v e m e n t s of organelles, contraction, expansion, a n d flow of cytoplasmic ground substance and so forth. I t concerns more or
TABLE I I . Intracellular Transport (IT)
E X T R A — I T I N T R A — I T
Pinocytosis Diffusion—random
intake of water preferential
i n t a k e of n u t r i e n t s passive
translocation facilitated
active
Cytoplasmic m o v e m e n t s streaming
organelle m o v e m e n t surface plications Nuclear rotation
less p e r m a n e n t organelles and protoplasmic ground m a t r i x . I t m a y be called " t r u e intracellular t r a n s p o r t " or " i n t r a - i n t r a c e l l u l a r t r a n s - p o r t . " M o v e m e n t between t h e e x t r a - and i n t r a - c o m p a r t m e n t s is a c - complished by t r a n s p o r t across m e m b r a n e s , m e m b r a n e fusion, m e m - b r a n e dissolution or m e m b r a n e formation.
Pinocytosis, described long ago by Lew r
is [ 3 2 ] , differs in certain animal cells and protozoa. I n tissue-cultured cells, leucocytes, ascites t u m o r cells, etc., pinocytosis is characterized by u n d u l a t i n g m o v e m e n t s of t h e cell m e m b r a n e t h a t lead to ruffles a n d folds which enclose in vesicles v a r y i n g a m o u n t s of external fluids. I n amoebae a n d some other protozoa, pinocytosis consists of t h e invagination of m e m b r a n e s leading to t h e formation of a t u b u l a r channel from whose base vesicles separate by a budding mechanism. Pinocytosis is a p p a r e n t l y a m e m - b r a n e - d e p e n d e n t process a b o u t whose initiation a n d cessation m u c h remains to be learned. T h e most efficient inducers of pinocytosis are
charged molecules such as acidic and basic amino acids a t p H 8.0, proteins in cationic states, a n d m o n o - a n d divalent cations. C a r b o h y - drates, nucleic acids, n e u t r a l amino acids, ethyl alcohol, a n d urea do not induce pinocytosis. Insulin and cortisone h a v e been said to induce pinocytosis in some m a m m a l i a n cells. C h a p m a n - A n d r e s e n [10]
has recently t a b u l a t e d the effects of these agents on several cell t y p e s . B e n n e t t [3] suggested t h a t t h e initial event is t h e adsorption or inter- action of inducer molecules a t the surface of t h e cell m e m b r a n e . I t appears, however, t h a t certain regions of t h e m e m b r a n e plicate more readily t h a n others and t h e initial event m a y consist of t h e interaction of charged molecules and specialized extended regions of the cell sur- face. These events h a v e not been a d e q u a t e l y studied nor is enough known a b o u t alterations in phospholipid lipoprotein m e m b r a n e s upon contact with various solutes. Rosenberg [43] has recently described how a monomolecular film of phospholipid and cholesterol undergoes changes in interfacial tension, depending on t h e ionic p r o d u c t of the buffer and t h e molar ratios of sterol and lipid. P r e s u m a b l y , a sufficient lowering of interfacial tension would favor the micellization of t h e monolayer. F a r more complex model studies are needed before it will be possible to d r a w direct analogies between t h e inducers of p i n o c y t o - sis and their measurable effects on model m e m b r a n e s .
T h e initial vacuoles or pinocytotic vesicles are generally 1 to 2 μ, in d i a m e t e r ; as m a n y as 100 vacuoles have been observed a t a n y one t i m e in a m o e b a e ; their average lifetime is b u t a few minutes as observed microscopically. I t is n o t clear whether vesicle formation results from flow of m e m b r a n e , or expansion, or formation of new m e m b r a n e , or simply m o v e m e n t of the old m e m b r a n e due to changes in structures subjacent to m e m b r a n e s . I n fact, different processes m a y underlie pinocytosis in different cell t y p e s . A few investigators h a v e suggested t h a t t h e pinocytotic vesicle is i n t i m a t e l y associated with the ergastoplasm and simply represents a localized herniation of t h e gap between t h e m e m b r a n e s of t h e reticulum. T h e r e is no support from actual observation t h a t such is the case.
Once formed, t h e vesicle carries into t h e cell the inducing substance and some of the fluid medium b a t h i n g t h e cell. I t s m e m b r a n e is per- meable to w a t e r and small molecules such as amino acids and glucose, as shown by t h e radioactive labeling experiments of H o l t e r [ 2 1 ] . F o r a considerable portion of its life history, the contents of the v a c - uole m a y remain inaccessible to the interior cytoplasm. T h e r e is con- siderable evidence t h a t t h e vacuoles can shrink, coalesce, or divide,
or sprout local micropinocytotic vesicles, b u t whether t h e y fully d i s a p - p e a r is not known. T h e p e r m e a b i l i t y of t h e m e m b r a n e s m a y be altered or t h e m e m b r a n e s m a y be fragmented. W h e t h e r complete digestion occurs is n o t known. T h e s t r u c t u r e of its m e m b r a n e s eventually changes such t h a t its contents can be finally interiorized a n d m a d e p a r t of t h e internal milieu. T h i s ingestion of t h e m e m b r a n e would be linked to metabolic processes. Gey [15] h a s claimed t h a t mitochon- dria m a y result from t h e fusion of shrunken pinocytotic granules with preexisting mitochondria or in de novo fashion. Other investiga- tors h a v e suggested t h a t cellular inclusion bodies such as lysosomes are end products of pinocytotic vesicles.
I n some cell t y p e s t h e contents of t h e vesicles are n o t incorporated into t h e cytoplasm b u t are k e p t exteriorized. F o r example, in t h e kidney t u b u l a r epithelium of t h e frog, t h e intestinal epithelium, t h e endothelial cells of blood capillaries, and t h e endothelium of t h e r a b b i t cornea t h e r e is an over-all directed m o v e m e n t of t h e vesicle or vacuole.
K a y e a n d P a p p a s [27] a n d K a y e et al. [28] h a v e shown t h a t for t h e corneal cells, as illustrated in Fig. 2, m a t e r i a l s are t a k e n in on one side and t r a n s l o c a t e d to another side. T h e m e m b r a n e s of these pinocytotic vesicles a p p e a r to persist. T h e l a t e r a l m a r g i n s of t h e epi- thelial cells are joined together in linear fashion by a dense connection, the t e r m i n a l bar, which m a y possibly be associated with an inner polarization or orientation of ground substance or endoplasmic reticu- lum. I n t h e case of intestinal epithelium, fat droplets are pinched off a t the base of t h e intermicrovillous space. T h e pinocytotic vacuoles travel i n w a r d t h r o u g h t h e cytoplasm a n d subsequently emerge or are extruded into t h e intercellular space a t t h e lateral m a r g i n of t h e cell basal to t h e t e r m i n a l b a r by a fusion of t h e m e m b r a n e of t h e vesicle with the external cell m e m b r a n e . This phenomenon has not been ob- served for corneal endothelial cells in frogs. I t s absence is a t t r i b u t e d t o t h e lack of t e r m i n a l bars between t h e endothelial cells of t h e frog cornea. I t is possible t h a t some of the contents of t h e vacuoles can be t a k e n into t h e cytoplasm of the cell by changes in m e m b r a n e p e r m e a b i l i t y .
T h a t intracellular t r a n s p o r t by pinocytotic vesicles is a normal, physiological process was shown b y F a r q u h a r a n d P a l a d e [14] for t h e renal glomerulus. Similarly, t h e significance of pinocytosis in t h e secretion of zymogen in p a n c r e a t i c exocrine cells has been described by C a r o and P a l a d e [6, 7 ] . T h i s process of t h e t r a n s l o c a t i o n or m o v e - ment of m a t e r i a l s t h r o u g h channels of t h e endoplasmic reticulum, or
by pinocytotic vesicles wherein m a t e r i a l s m a y be acted upon chemi- cally and m a y or m a y not be m a d e p a r t of cytoplasmic m a t r i x , a p p e a r s to be a major mechanism for intracellular t r a n s p o r t . I t not only p r o - vides for t h e p a r t i a l l y to t o t a l l y directed m o v e m e n t of large and small molecules b u t also for t h e quasi-exteriorization of certain cyto- plasmic components a w a y from t h e outer cell borders.
Descemets
R A B B I T
Anterior chamber Endoth.
° Q ° / q O . O
Stroma
' . * S O U R C E -
Epithelium
S O U R C E
C : I '.
Anterior chamber
Descemets
F R O G
Anterior chamber u Endoth. Θ
> ·
s —
© ο Stroma.
SOURCE.
ι· ι ιο ι ωΙ©
Α
Ψ
!S5 Ι Ι Ι I Epith
FIG. 2. Translocation through endothelial cells of cornea. F r o m Kaye and P a p p a s [27] and K a y e et al [28].
T h e mechanisms underlying this form of t r a n s p o r t still escape us. T h e sources of energy a n d their transduction for t h e formation of vacuoles, for pinching t h e m off and moving t h e m are still specula- tive. Questions to be answered a r e : H o w can m a t e r i a l s be moved within t h e endoplasmic reticulum and how do pinocytotic vacuoles move a b o u t ? Are m e m b r a n e s being constantly disintegrated a n d r e - formed or are t h e y sliding? W h a t is t h e m a n n e r of their expansions,
plication, and contractions, a n d w h y are these plications induced b y charged ions a n d molecules? H o w i n t i m a t e l y are their activities deter- mined b y t h e physical properties of enclosed m a t r i x ? H o w i m p o r t a n t are diffusion mechanisms in channels of t h e endoplasmic r e t i c u l u m ? Are t h e motive forces localized a t t h e m e m b r a n e s which in some cases h a v e been shown to h a v e adenosinetriphosphatase ( A T P a s e ) a c t i v i t y or localized in associated contractile fibers? D o motive forces reside within the contents of vacuoles? Are t h e driving forces within t h e cytoplasm ? W h y does t h e pinocytotic process differ in a n i m a l cells and protozoa?
M a n y more detailed serial observations by light a n d electron microscopy on t h e directed or r a n d o m m o v e m e n t s of pinocytotic vesi- cles are necessary. F o r t u n a t e l y , q u a n t i t a t i v e d a t a can be obtained with considerable a c c u r a c y using c u r r e n t techniques. F o u r experimen- t a l approaches h a v e been developed. T h e first consists of direct counts
of the n u m b e r of channels for pinocytosis t h a t develop in amoebae following initiation by inducers. C h a n n e l formation can be determined as a function of time, t e m p e r a t u r e , p H , etc. This method has limited usefulness for t r a n s p o r t studies. T h e second technique m a k e s use of radioactively tagged molecules which are incorporated by pinocytosis.
E x a m p l e s a r e 1 4
C - l a b e l e d glucose, 2 2
N a C l , N a 8 2
B r , 3 5
S - l a b e l e d m e t h i o - nine, a n d
1 3 1
I - l a b e l e d serum a l b u m i n . Some of these molecules a r e also inducers of pinocytosis. R a d i o a u t o g r a p h i c experiments show con- siderable promise not only for determining t h e mechanisms associated with pinocytosis b u t also for following t h e m o v e m e n t s of vesicles within cells. I t should be possible to measure t h e r a t e a n d direction of m o v e m e n t s of individual vesicles in large n u m b e r s of cells to assess t h e common elements characteristic of this form of intracellular t r a n s - port. I n view of t h e relatively large size of t h e vesicle d u r i n g most of its life cycle, it can be localized with considerable precision. Q u a n - t i z a t i o n of t h e ingested molecular population undergoing t r a n s p o r t presents a far more difficult problem which h a s n o t been a d e q u a t e l y assessed.
A third highly useful experimental method involves t h e use of fluorescent compounds such as fluorescein-labeled γ-globulin or p l a s m a albumin. This method was initiated b y H o l t e r a n d M a r s h a l l [22]
and h a s even been used for s h o r t - t e r m studies in vivo of t h e progress of pinocytotic vesicles.
T h e fourth related experimental approach is t h e s t u d y of t h e u p - t a k e of basic dyes which are inducers of pinocytosis. E x a m p l e s a r e
acridine orange a n d Alcian blue (copper p h t h a l o c y a n i n e ) . I n general it is difficult to follow t h e course of these dyes since t h e compounds m a y form preferential associations or readily diffuse t h r o u g h m e m - branes. Other investigators h a v e tried t o circumvent these difficulties b y inducing t h e pinocytosis of enzymes or iron d e x t r a n a n d t h e n localizing these compounds by histochemical stains.
Results to d a t e h a v e been divergent and no common p a t t e r n of intracellular t r a n s p o r t by pinocytosis has y e t emerged. M a j o r discrep- ancies are expected in view of t h e difficulty in s t u d y i n g large n u m b e r s of cells in similar physiological states a n d in view of t h e m a r k e d sensitivity of t h e pinocytotic mechanism to physical a n d chemical alterations in environmental conditions. As these environmental de- pendencies are recognized a n d brought under control, there is consid- erable hope t h a t the combined use of radioactive and fluorescent tracers, a n d interference, phase, a n d electron microscopy for t h e s t u d y of individual cells a n d large synchronized populations will m a k e it possible to localize vesicles w
T
ith great accuracy and to determine, p e r h a p s less accurately, t h e direction and r a t e of their movements t h r o u g h o u t the cell. I n this m a n n e r q u a n t i t a t i v e d a t a on u p t a k e and intracellular t r a n s p o r t by pinocytosis will be obtained.
T h e t r a n s p o r t a n d m o v e m e n t of m a t e r i a l s within cytoplasm, t h e i n t r a - i n t r a c e l l u l a r t r a n s p o r t , is even less well understood t h a n p i n o - cytotic mechanisms. T h e experimental difficulties a n d h a z a r d s are far greater. I t is arduous and often not possible to trace the movements of macromolecules by serial electron microscopy, whereas pinocytotic vesicles are sufficiently identifiable to be followed readily by electron microscopy, fluorescence microscopy, a u t o r a d i o g r a p h y , and so forth.
As discussed earlier, t h e problems in molecular traffic control m a y involve from a few t h o u s a n d to m a n y millions of macromolecular units per second, depending on t h e cell t y p e , within a m a t r i x exhibiting flows ranging from essentially static conditions to hundreds of microns per second. R e g a r d i n g t h e m o v e m e n t s of m a t e r i a l s within t h e c y t o - plasmic m a t r i x , two interrelated subheadings m u s t be considered in
detail. T h e first is t h e passive diffusion along both r a n d o m a n d prefer- ential p a t h w a y s . T h e second is the convective t r a n s p o r t of m a t e r i a l s associated with v a r i a b l e or directed cytoplasmic m o v e m e n t s or flows, and t h e m o v e m e n t of organelles.
Undoubtedly, diffusion as a r a n d o m collision process is an effec- tive a n d principal mechanism for distributing r e a c t a n t s a n d their products within fluid systems. As a physical mechanism, it is ever
present a n d cannot be disregarded. I t is relatively r a p i d although n o t necessarily t h e m o s t efficient mechanism for assembling subcom- ponents of molecular aggregates since large n u m b e r s are required for r a n d o m collisions. F o r small molecules a n d ions, t h e diffusion coeffi- cients in gels differ little from those in homogeneous liquids a n d these molecules are readily available from t h e environmental pool. One would expect r a p i d mixing of such molecules within cells provided no barrier were interposed. I t seems unlikely t h a t reaction times in- volving these small molecules are diffusion limited. F o r larger mole- cules t h e situation a p p e a r s to differ. Such molecules m a y n o t be r e a d - ily available to cells from t h e external environment. T h e y h a v e often been observed to h a v e inhomogeneous distributions within cells ; their n u m b e r is far more limited; t h e y act a t specific sites; a n d t h e y are more easily restrained or blocked in their m o v e m e n t s within a cell by m e m b r a n e s a n d other lamellar s t r u c t u r e s . Diffusion coefficients
for proteins such as egg albumin in aqueous solution are of the order of 10~
6 c m
2
/ s e c U n d e r such conditions these protein molecules, under a concentration gradient, can move distances of 5 μ in 0.04 seconds.
W i t h i n t h e cytoplasm, however, there is no reasonable estimate of the diffusion coefficient for macromolecules. I n view of t h e multiplicity of ergastoplasm, microtubules, fibrils, formed organelles, gel-like r e - gions, multiple surfaces for adsorption, all of which are subject to v a r i a t i o n s with t i m e a n d act as barriers to t h e m o v e m e n t of large molecules, it is difficult to see how t h e p a r a m e t e r diffusion c o n s t a n t is meaningful over appreciable distances. Over short distances for example between n e a r b y organelles, r a n d o m diffusion can be exceed- ingly rapid, b u t over larger distances it seems t h a t t h e over-all m o v e - m e n t s of cytoplasm, t h e convections of bulk liquid, t h e mechanical m o v e m e n t s a n d g y r a t i o n s of inclusions, a n d preferential p a t h w a y s for molecular traffic m a y be i m p o r t a n t m e a n s for mixing and t r a n s - port. Some synthetic steps m a y a c t u a l l y become diffusion-limited.
A few cases of diffusion-controlled enzyme reactions h a v e been r e - ported. F o r example, Strother a n d A c k e r m a n [47] h a v e shown t h a t t h e reaction of t h e h e m e - t y p e enzyme, catalase, with hydrogen perox- ide is diffusion-controlled in solutions whose viscosities are several times t h a t of w a t e r . U n f o r t u n a t e l y little additional information con- cerning such mechanisms is available. W i t h i n cells, t h e situation is far more complex. T h e m e m b r a n o u s or fibrous n e t w o r k m i g h t b a r t h e r a n d o m diffusion of some macromolecules or inherently provide directed, facilitated, channeled, or preferential p a t h w a y s for the m o v e -
merits of other macromolecules. L a m e l l a r or fibrous units might impose a vector character to t r a n s p o r t and furnish selective p a t h w a y s for specific molecular species. A small a m o u n t of evidence for t h e presence of such preferential p a t h w a y s is being accumulated. Grover [16] has observed t h a t ions moving within tendons are confined p r i m a r i l y to t h e interface between t h e collagen fiber a n d its surrounding m a t r i x r a t h e r t h a n being distributed t h r o u g h o u t t h e bulk m a t r i x . These ions aggre- gate along t h e polypeptide a t t h e position of t h e polar amino acids.
T r a n s p o r t of ions along t h e direction of t h e collagen fibers is roughly two to three times more r a p i d t h a n t r a n s p o r t normal to t h e fiber. P a t h - w a y s such as these point out t h e possibility t h a t selective p a t h s m a y be involved in intracellular t r a n s p o r t . A m a t h e m a t i c a l analysis of diffu- sion in the presence of sinks or t r a p s has been given by H e r m a n s [18]
a n d Dishon et al. [ 1 3 ] .
I t is a p p a r e n t t h a t t h e relative significances of r a n d o m versus directed t r a n s p o r t by diffusive processes h a v e not been a d e q u a t e l y analyzed or studied experimentally. I n highly structured systems the analysis of diffusive mechanisms becomes more complex and the most applicable analyses in this regard are now being carried out by chemi- cal engineers working on t h e properties of structured continua, t h a t is, systems whose states depend n o t only on stress, t e m p e r a t u r e , com- position, charge, etc., b u t also on t h e average molecular configuration and interaction. D a h l e r a n d Scriven [11] have recently initiated a systematic m a t h e m a t i c a l analysis of these structured continua assum- ing certain aggregative properties of macroscopic groupings. T h e r e is every indication t h a t approaches such as these will be applicable to t h e problem of intracellular t r a n s p o r t . T h e problem is further com- plicated by coupling between diffusive and connective processes and by the presence of reactive sites.
As discussed above, convective t r a n s p o r t by mass movement, v a r i - ously referred to as cytoplasmic or protoplasmic streaming, appears to p l a y a role in several cell t y p e s . I n small cells such as bacteria t h e distances involved and t h e barriers to diffusion m a y n o t be exces- sive such t h a t r a n d o m collision processes suffice for most t r a n s p o r t of chemical r e a c t a n t s or metabolic products. I n other cells, especially elongate mononucleate ones such as nerve cells or p l a n t cells like Acetabularia, or cells whose cytoplasm is composed of multiple b a r -
riers to diffusion, mass t r a n s p o r t m a y be significant.
F o u r interrelated components of cytoplasm h a v e b y now been iden- tified which can generate stress distributions t h a t can lead to convec-
tive flow, in addition to diffusive and other coupled mechanisms. These are the ill-defined ground substance, membranous or interfacial re- gions, microtubular or columnar elements, and microfibrillar units.
A t present there is no reason to assume t h a t the specialized driving mechanism for convective transport is limited to any one of these components. Rather, all m a y play interdependent roles of varying significance in different cell types. Indeed, the idea t h a t several driving mechanisms underlie transport phenomena seems more consonant with the striking differences in rate, direction, location, and character of the cytoplasmic movements described earlier for several types of cells.
TABLE I I I . Intracellular Transport Mechanisms MECHANOCHEMICAL TRANSDUCTIONS
Sol-gel transformations—contractile fibrils and fibers, cooperative phenomena, torsional screw mechanisms, reversible syntheses, polymerizations, and crys- tallizations
Microtubular dynamics—undulations, sliding mechanisms, contractions, crystallizations
Interfacial dynamics—molecular mechanics of surfactants, metaphase changes, fluid pumps, fluctuations within membranes, fluctuations and undulations of membranes
ELECTROMECHANICAL TRANSDUCTIONS
electrokinesis—cataphoresis, electroosmosis, streaming potentials électrostriction
ELECTROMECHANOCHEMICAL TRANSDUCTIONS COLLISION PROCESSES
random diffusion
selective sources and sinks preferential pathways
The problem is essentially one of mechanochemistry where altera- tions in chemical potential lead to reversible contractions and expan- sions which are coordinated and directed or polarized, t h a t is, a driving force is considered to be developed through a reversible transduction of chemical energy among mechanical and possibly electrical forms.
Several such mechanisms are tabulated in Table I I I . These driving forces are in turn coupled in various ways to cytoplasmic flows.
Perhaps the most familiar physical change leading to contraction is t h a t of sol-gel transformation, a mechanism t h a t Marsland [35]
and others have long favored as a cause of protoplasmic stream- ing. The cytoplasm is pictured as a potentially contractile gel-like structure in which changes in fluidity and volume result from forcible
foldings o f protei n molecule s wit h maintenanc e o f intermolecula r link - ages. Cytoplas m i s viewe d a s a ge l o f typ e I I , namely , on e fo r whic h contraction i s accompanie d b y th e desorptio n o f heat , t h a t is , a n exergonic process . Solatio n i s impose d b y increasin g pressur e an d b y decreasing temperature . I t i s accompanie d b y a decreas e i n volum e (negative volum e change) , wherea s gelatio n i s a n endergoni c proces s and i s accompanie d b y a n increas e i n volum e (positiv e volum e change). Energ y fo r formin g an d sustainin g an d possibl y contractin g the ge l structur e i s supplie d b y adenosin e triphosphat e ( A T P ) . Th e principal uncertaint y involve s th e contractio n o f th e ge l b y A T P hydrolysis fo r th e accomplishmen t o f mechanica l work .
The term s so l an d ge l refe r t o measurabl e change s i n th e consis - tency o r fluidit y o f cytoplasm . I n thi s sens e the y revea l littl e abou t the macromolecula r alterations , th e distributio n o f contractil e o r shearing materials , th e localization s o f thei r actions , th e fibrillar , reticular, an d tubula r organizations , an d thei r permanenc e o r ephem - eral o r dynami c qualities , an d th e function s o f membranou s o r lamel - lar element s withi n cytoplasm . Severa l investigator s hav e recentl y reported o n th e isolatio n o f substance s t h a t underg o structura l change s under th e influenc e o f A T P . Loew y [33] , Ts' o et al. [50 ] an d mor e recently Nakajim a [37 ] hav e prepare d fro m myxomycet e plasmodi a (Physarum polycephalum) differen t contractil e protei n fraction s which reac t specificall y wit h A T P in vitro. Thes e contractil e protein s behave somewha t lik e th e contractil e muscl e proteins , actomyosin . Wohlfarth-Bottermann [51 ] ha s describe d a pleomorphis m o f groun d plasm i n amoeb a an d myxomycète s includin g a threadlik e reticulum , presumably o f contractil e materials . H a y a s h i [17] , an d K a m i y a an d Kuroda [24 ] hav e show n t h a t th e motiv e forc e fo r rotationa l stream - ing i n th e p l a n t cells , Char a an d Nitella, i s a n activ e shearin g forc e generated a t th e boundar y betwee n th e cortica l ge l an d endoplasm . Recently Jarosc h [23 ] describe d fo r Chara, an d K a m i y a an d Kurod a
[24] describe d fo r Nitella th e presenc e o f movin g fibril s oriente d i n organized fashio n o n th e inne r surfac e o f th e cortica l ge l layer . M a n y undulating polygon s o f thes e fibril s hav e bee n observe d i n drop s o f cytoplasm isolate d fro m thes e plan t cell s wit h rate s o f propagatio n of th e orde r o f 9-10/x/sec. Hoffmann-Berlin g [20 ] ha s show n t h a t contractile protein s sensitiv e t o A T P an d i n way s simila r t o thos e in th e mor e primitiv e cel l type s ar e presen t i n fibroblasts .
The mod e o f actio n o f filament s suc h a s thes e i n cel l movemen t and cytoplasmi c flo w ha s bee n th e subjec t o f muc h speculation .
Jarosch [23] has proposed t h a t torsional forces within protein helices give rise to alterations in helical pitch a n d t h a t previously r a n d o m l y a r r a n g e d helices m a y associate into bundles of superhelices. T h u s , he places protoplasmic m o v e m e n t on a screw-mechanical basis, where small changes in pitch can lead to considerable displacements. T h e mechanism is broadly applicable to m a n y forms of m o v e m e n t . Other similar mechanisms m a y p l a y i m p o r t a n t roles. F o r example, changes in the cross-bonding of helical polymers or in t h e configuration of protein molecules can cause contraction as can t h e more familiar r a t c h e t a n d sliding mechanisms proposed for contraction of muscle.
Other forms of m o v e m e n t m a y depend on less similar mechanisms involving t h e synthesis and polymerization of macromolecules into filaments and fibrils. T h e work of K a t c h a l s k y [25] on t h e m e c h a n o - chemistry of fibers a p p e a r s p e r t i n e n t in this regard. H e has demon- s t r a t e d t h a t the stretching or contraction of collagen fibers can be measured as functions of the changes in chemical potential of t h e system. T h e conversion between mechanical a n d chemical energy was found to pass through extremely s h a r p m a x i m a , depending on t h e concentration of t h e salt solution surrounding t h e fiber. M a x i m a oc- curred a t t h e melting point of t h e collagen, as though t h e fiber were undergoing m a x i m u m changes in length by melting and recrystallization.
This phenomenon of reversible melting a n d recrystallization has not been a d e q u a t e l y studied, especially in relation to biological sys- tems. I n a liquid polymer, which can be looked upon as a mass of writhing, a r t i c u l a t i n g fibrils, crystallization consists of t h e alignment of fibrils alongside one another in orderly a r r a y . A p p a r e n t l y t h e t r a n - sition between the crystalline a n d noncrystalline phases can be m a d e quite r a p i d l y (in less t h a n 10 seconds in t h e case of collagen) and as a cooperative process could lead to considerable m o v e m e n t of sur- rounding ground substance. Crystallization can also result either in the formation of long fibrillar units or t h e formation of lamellar or cylindrical micelles if an interface or similar imperfection is present.
Surface loci a t m e m b r a n e s can provide large n u m b e r s of nucleating sites to initiate transcrystallization, t h a t is, t h e formation of lamellar micelles, whereas s t r a i n imperfections within t h e cytoplasmic m a t r i x can encourage t h e formation of cylindrical micelles. T h e r e is a small a m o u n t of evidence t h a t crystallization of some macromolecular units m a y be associated with surface movements. T a y l o r a n d R o b b i n s [48]
have described microspikes or microprojections extending from the
surfaces of cells. These projections are roughly 1000 Â in diameter, form very rapidly, can be several microns in length, can have very sharp bends, fold upwardly to merge once again with the cell surface, and at times can be shown by electron microscopy to have a crystal- line-like core. Whether cytoplasmic filaments evidence the working of similar processes is not known. For the present, we must keep in mind t h a t there m a y be semicrystalline, transient structures within cells in general, which m a y be important in causing movements of cytoplasm.
Thus, some of the questions to be faced in future investigations are as follows. Are contractions localized solely or in p a r t to fibrillar components within cytoplasm? How can these components be identi- fied and isolated? In w h a t manner are they formed and associated from a cytoplasmic molecular pool or molecular reticulum? How per- manent are such structures or is their existence only temporary? W h a t are their dynamic properties? In what ways do they contract or undulate so as to transmit coordinated and reversible motions to the cytoplasmic matrix, or do they take p a r t in the totality of matrix move- ment? D o they cause movements of matrix by contracting or expand- ing as linear macromolecules, by distortion of linkages and torsion of the helix, by undulating wave motion, by polymerization and crys- tallization? W h a t is their relative importance and possible interrela- tion with other formed organelles and membranes within cytoplasm t h a t m a y play significant roles in cytoplasmic mixing and flow?
Several investigators [ 2 , 3 0 ] , have recently observed and described formed organelles t h a t are in some ways similar to and possibly re- lated to the filamentous units described above; they m a y also provide the motile drive for cytoplasmic movements. These structures have been termed, for the moment, microtubules. They are 200 to 270 Â in diameter and have been observed by electron microscopy to be present in m a n y animal and plant cells subjacent and tangential to the outer cell membrane. These "tubular" structures m a y be up to several microns in length and appear to have an inner core of different consistency from their outer walls. In a sense the word tubule m a y be misleading since the inner core m a y be highly structured.
There are numerous other instances in which cytoplasmic tubules which m a y or m a y not be related are found. Among these are the spindle fibers of mitosis in the giant amoeba (Pelomyxa carolinensis), the spindle fibers of dividing cells of sea urchin embryos, the filamen- ture of cilium and flagellum, the marginal band of nucleated red blood
cells, t h e developing spermatid, t h e cores of microspikes, certain n e u r o - t u b u l a r structures, a n d so forth.
L e d b e t t e r a n d P o r t e r [29] h a v e ascribed t o cortical microtubules the development a n d orientation of displacement forces. W h e t h e r flow of cytoplasm is a p r o d u c t of u n d u l a t i n g motions of these tubules or a result of interactions a t t h e surface of s t a t i o n a r y tubules cannot be decided. W e do not know whether microtubules bring a b o u t c y t o - plasmic flows b y processes similar to those described above for fila- ments, n a m e l y , by undulation, or linear contraction, or torsional changes in pitch, or reversible crystallizations, or by a p u m p i n g action.
However, all these proposed mechanisms require detailed inquiry.
T h e r e is some p r e l i m i n a r y evidence from L e d b e t t e r a n d P o r t e r [30]
t h a t microtubules m a y consist of cylindrically a r r a n g e d longitudinal a r r a y s of filaments, but from a s t r u c t u r a l a n d biochemical viewpoint little is y e t known. T h e r e have been suggestions t h a t these organelles m a y be continuous a n d stable hoops a r o u n d t h e cells, subjacent t o t h e cortex, b u t more evidence is needed to s u b s t a n t i a t e these proposals.
F r o m functional a n d d y n a m i c viewpoints even less is known, and the previously m a d e comments concerning t h e p e r m a n e n c e a n d actions of fibers and filaments are equally applicable and p e r t i n e n t to microtubules.
T h e roles of m e m b r a n e s in intracellular t r a n s p o r t h a s been s u b - jected to much controversy. I n recent y e a r s t h e r e h a s been a t e n - dency to assume t h a t t h e fibrous and now possibly t u b u l a r reticulum of cytoplasm, in preference to m e m b r a n o u s elements, provides driving forces for cytoplasmic flows. Yet, t h e observable movements of m i t o - chondria, pinocytotic vesicles, inclusion bodies, lysosomal particles, etc., and the d y n a m i c v a r i a t i o n s in ergastoplasm all suggest t h a t m o - tility a n d driving mechanisms m a y arise from m e m b r a n o u s systems.
M e m b r a n o u s elements and adjacent macromolecular coats can serve several i m p o r t a n t functions concerning intracellular t r a n s p o r t . F i r s t , t h e y can provide selective, preferential p a t h w a y s for passive diffusion or active m o v e m e n t of certain molecules, as m a y t u b u l a r a n d filamen- tous structures. Second, t h e y can act as sources a n d sinks for certain macromolecules via adsorption and desorption. T h i r d , t h e y can p r o - vide selected regions for t h e enhanced or reduced catalysis of certain reactions. F o u r t h , in interactions with other structures t h e y can in t u r n affect molecular a r r a n g e m e n t s within t h e cytoplasm. Fifth, t h e y can affect a n d possibly limit diffusive mechanisms by bringing a b o u t convective flows in t h e bulk liquid. Sixth, t h e y m a y through m e c h a n o -
chemical t r a n s d u c t i o n of energy provide motive forces for surface movements associated with pinocytosis a n d phagocytosis, for c y t o p l a s - mic flows, a n d for t h e t r a n s l a t o r y a n d r o t a t i o n a l m o v e m e n t s of m e m - branous organelles. This energy t r a n s d u c t i o n will differ a m o n g differ- ently structured a n d composed m e m b r a n o u s units.
K a v a n a u [26] has recently suggested t h a t m e t a p h a s e changes in the u l t r a s t r u c t u r e of t h e m e m b r a n e s lead to t h e forceful removal of w a t e r molecules in certain regions. T h e p u m p i n g o u t of low-molecu- lar-weight ions and hydrions is assumed to provide a propulsive force which can bring a b o u t m o v e m e n t of t h e m e m b r a n e . W h e t h e r such transitions are significant is not known, b u t t h e proposal w a r r a n t s investigation. L u c y a n d G l a u e r t [ 3 4 ] , on t h e basis of electron micro- scope studies of various mixtures of lecithin, cholesterol, and saponin, have suggested t h a t small globular micelles as well as bimolecular leaflets m a y function as building blocks of m e m b r a n o u s units, a n d t h a t regions of t h e m e m b r a n e m a y undergo transitions between these
forms.
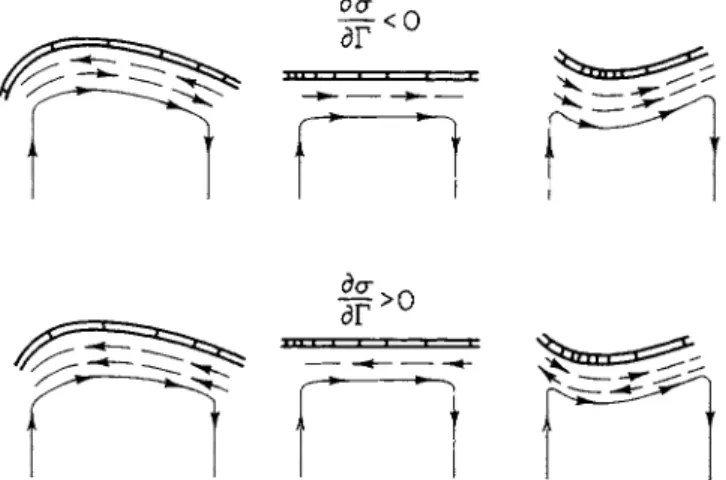
I t is well known t h a t changes in chemical potential a t certain regions of an interface can result in mechanical movement of t h e interface. T h e phenomena are variously categorized as P l a t e a u - G i b b s - M a r a n g o n i effects of "interfacial d y n a m i c s . " T h e y consist of two related surface effects. T h e first is m o v e m e n t in a fluid interface due to local v a r i a t i o n s of interfacial tension which in t u r n result from differences in composition, t e m p e r a t u r e , or electric charge or pressure. T h e second is m o v e m e n t of t h e interface. Differences in interfacial tension then arise from differential deformations, t h e exten- sion or contraction of t h e fluid interface. These surface effects are illustrated in Fig. 3. T h e model system is pictured as a mobile inter- facial region separating two liquid phases. Several interdependent and interacting processes are assumed to be t a k i n g place. Among these processes are localized areas of t h e interface where certain chemical reactions, transformations, selective adsorption, or desorption are oc-
curring. These heterogeneous reactions result in differences in inter- facial tensions which in t u r n lead to surface t a n g e n t i a l stresses and movements of m a t e r i a l s within the interface. T h e r a t e s of these p r o - cesses are controlled by the relative rates of diffusion of r e a c t a n t s to the interfacial region, chemical and physical reaction rates, and the r a t e of movement within the interface. I n addition, changes in shape of the interface are associated with t h e development of n o r m a l stress components (capillary pressure) and gradients in concentration
of t h e surface-active m a t e r i a l s as interfacial regions a r e compacted and expanded. C o n c u r r e n t alterations in t h e t a n g e n t i a l a n d n o r m a l stress components will result. T h e vectorial form of these stresses will depend on w h e t h e r t h e surface-active m a t e r i a l s t e n d t o decrease or increase interfacial tensions. M o v e m e n t s within a n d of t h e inter- facial region will result in m o v e m e n t s of t h e adjacent, bulk liquid.
T h e movements of m a t e r i a l s to t h e interfacial region will be governed by two processes: molecular diffusion due to concentration differences and convective t r a n s p o r t by t h e moving bulk solution. T h e a u g m e n t a - tion or diminution of interfacial tensions by s u r f a c t a n t s together with
FIG. 3. Interfacial dynamics, surface tension driven flows.
t h e process of eonveetive-diffusion can amplify, stabilize, or d a m p e n movements within a n d of the interfacial region. These effects are currently being subject to rigorous m a t h e m a t i c a l a n d experimental analysis. Excellent studies and reviews h a v e been provided by Scriven
[45] a n d Levich [ 3 1 ] . T h e y can be viewed as a form of m e c h a n o - chemistry similar to t h a t described by K a t c h a l s k y for fibers, where an alteration in chemical potential results in mechanical motions, a n d vice versa. F o r surfaces, t h e principal p a r a m e t e r s t h a t determine t h e t r a n s d u c t i o n between chemical a n d mechanical energy are t h e reaction r a t e affecting interfacial tension, t h e interfacial tension, a n d the surface d i l a t a t i o n a l viscosity. E n e r g y dissipation in this region will depend upon t h e surface shear viscosity.
W h e t h e r similar processes t a k e place a t biological interfaces is
not known. Models of biological interfaces must obviously always be viewed with critical skepticism. The organization of materials at the interfaces of surfaces of cells is far more complex. However, macromolecular components of biological interfaces can provide local- ized sites for chemical reactions or transformations, or in their inter- actions with the polar end groups of the phospholipids and other molecular constituents of the membranous leaflet can be selectively
adsorbed and desorbed, or structural changes m a y occur in the molecu- lar organization of the interface. Local variations in interfacial tension will arise from such changes in composition, concentration, or struc- ture. Additional factors can cause similar disturbances in the physical properties of the interface. These include changes in electric charge distribution at and in the vicinity of the interface, variation in surface temperature and hydrogen ion concentration, ion formation and neu- tralization, energetic sources, configurational changes of the mem- brane, the presence of contractile proteins in the vicinity of the inter- face or membrane, nonuniform flow of the bulk fluid, and so forth.
This model consists of a network of closely interacting variables.
Interdependent transductions such as these provide an inherent degree of self-regulation. The energy exchanges are in a steady-state form of equilibrium. Small deviations or perturbations in any process or in the materials involved can be compensated for. For example, an excessive change in interfacial tension in one region, due to the pres- ence of a large pool of reactants, m a y be reduced by convective forces t h a t reduce the diffusion of reactants to t h a t region. I n this manner, the interfacial structure m a y have an intrinsic regulatory capacity which m a y be of importance in biological systems.
Undoubtedly, other physical and chemical processes can operate in intracellular transport. Among these are electrical field gradients which result in électrostriction, electrosmosis, electrophoresis, coopera- tive flows, flow with distributed chemical reactions, etc. These pro- cesses can lead to flows and movements of particles, inclusion bodies, surfaces, organelles, and liquids, and can markedly influence diffusion and protoplasmic streaming. The analysis and investigation of these complex phenomena can be carried out only if numerous simplifying assumptions are made. Some inquiries along these lines are being conducted by chemical engineers who face similar transport prob- lems. Whether they will be pertinent to intracellular transport is not known, but biologists interested in transport will have to follow these developments.
T h e m o v e m e n t s of organelles and t h e s t r e a m i n g of cytoplasmic m a t r i x a p p e a r to be interrelated phenomenon. W h e t h e r driving forces are supplied p r i m a r i l y b y changes in consistency a n d h y d r a t i o n of m a t r i x or by fibrillar, t u b u l a r , or surface drives is not clear. I n some cells, one t y p e of driving force for convective flow including organelle m o v e m e n t m a y be more significant t h a n another. T h e r e is for t h e m o m e n t no evidence t h a t one m e c h a n i s m of mechanochemical t r a n s - duction is preferable.
F r o m an a n a l y t i c a l point of view there has been extensive work on separate a n d coupled diffusive a n d convective mechanisms t h a t underlie t r a n s p o r t . As s t a t e d earlier, these studies h a v e been m a d e by investigators concerned with flow in complex chemical processes.
T h e y require determination of s t a t e variables such as chemical poten- tial, mechanical stress a n d macroscopic p a r a m e t e r s such as diffusion
and viscosity coefficients.
F r o m a biological point of view we face a dual problem. W h a t are needed a r e : (a) p e r t i n e n t model systems for studying diffusion a n d convection in structured systems in which m e a s u r a b l e chemical reactions t a k e place, and (b) more refined techniques for determining flux gradients of certain molecular species within cells. Hopefully, r e - cent t h e r m o d y n a m i c approaches to e v a l u a t e t h e stability of m a c r o - molecular associations in t e r m s of w e a k bonding look sufficiently promising for providing some of t h e baseline d a t a necessary for d y - namic studies.
Concerning t h e first need, t h e model m e m b r a n e s described by Mueller et al. [ 3 6 ] , T h o m p s o n [ 4 9 ] , Rosenberg [ 4 2 ] , a n d others serve as initial models for t h e investigation of interfacial d y n a m i c s , m e m - b r a n e stability, m e t a p h a s e change, energy t r a n s d u c t i o n s , a n d t r a n s p o r t t a n g e n t i a l and n o r m a l to interfaces. M u c h more d a t a are required a b o u t diffusion a n d t r a n s p o r t in polymer gels, especially those close to a s t a t e of crystallization. T h e mechanochemistry of model filaments of proteins a n d other macromolecules needs experimental s t u d y . W i t h t h e current development of techniques for t h e isolation of fibrils, microtubules, and certain m e m b r a n o u s units from cells, n a t u r a l phos- pholipids, proteins, mucopolysaccharides can be used for t h e model- ing-type experiments by reconstituting respective units. I t would be useful a t this point t o r e a c t i v a t e t h e classic studies on coacervates, a n d liquid crystals, especially model systems involving several phases where certain chemical reactions are t a k i n g place.
Concerning the second need to develop more refined techniques
for localization of macromolecules, several problems arise. R a d i o a u - t o g r a p h y combined with electron microscopy a p p e a r s t o be t h e most
sensitive method for t h e localization of certain molecular species com- bined with q u a n t i t a t i v e m e a s u r e m e n t of t h e n u m b e r of molecules present. E r r o r s in q u a n t i t a t i o n arise from specimen geometry in rela- tion to the emulsion, density of t h e specimen, t h e energy spectrum of t h e ^-emission, a n d t h e thickness a n d sensitivity of t h e emulsion.
Low-energy β - r a y s emitted by t r i t i u m offer t h e best resolution in t h e sense t h a t a one-to-one m a p p i n g of silver grains is desirable.
Other difficulties arise from t h e fact t h a t current techniques of speci- men p r e p a r a t i o n wash out water-soluble compounds and a n y associ- a t e d tracers. E r r o r s in localization arise for similar reasons. F o r opti- m u m precision t h e specimen section m u s t be less t h a n 1000 À to reduce self-absorption and crystal size in the emulsion and developed grain size must be kept as small as possible. Pelc [40], Caro [5], Perry [41], and Salpeter and Bachmann [44] discuss these problems in great detail. The general conclusion is t h a t resolution in space is approximately 1000 Â using the best current emulsions. The diffi-
culty in reducing this error now stems from the geometry of the sec- tion. For the identification and localization of molecular groupings of various sizes separated by large distances, the technique has much to offer. For the determination of fluxes or concentration gradients in certain cell types t h a t are well synchronized, the technique is prom- ising. For the localization of individual molecules being transported in a closely packed traffic column the resolution is still inadequate.
Finally, one additional warning relates to the combined use of electron microscopy with any technique involving the uptake of molecules.
The permeability of the membrane m a y be altered with fixatives.
The combined use of radioautography, electron microscopy, and cytochemistry provides additional flexibility in the localization of cer- tain molecular species but no greater resolution. Ostrowski et al. [39]
have used tritium-labeled diisofluorophosphate, a covalently bound inhibitor of esterases, to determine the number of molecules of acetyl- cholinesterase localized in certain regions. B a r r n e t t and Tice [1] and Novikoff [38] among m a n y others have applied the techniques of enzyme histochemistry to electron microscopy. By using aldehyde fixa- tive, Barrnett has been able to preserve oxidative enzymes. Novikoff
[38] and his associates for several years have been able to localize with histochemical stains acid phosphatase at the electron microscope level. Numerous similar applications are discussed in the review by
B a r r n e t t and Tice [1] a n d Seligman [ 4 6 ] . T h e precision of this t e c h - nique for localization of certain molecules is subject t o controversy.
Questions arise as to t h e degree to which histochemical fixation a n d retention of recognizable fine s t r u c t u r e are compatible. T h e r e q u i r e - m e n t of cofactors for t h e chemical reaction a n d migration of t h e final p r o d u c t due to diffusion leads to errors in localization. I n c u b a t i o n procedures can distort fine s t r u c t u r e . I n view of difficulties such as these it is not easy to estimate the accuracy of this techniques which varies with the method of fixation for electron microscopy a n d t h e histochemical procedure. T h e recent use of frozen-dried sections offer considerable promise for more accurate localizations and possible q u a n t i t a t i v e densitometric m e a s u r e m e n t of enzyme a c t i v i t y .
Ultramicrospectrographic procedures h a v e been developed for sev- eral y e a r s by workers such as Caspersson [8] a n d Chance [ 9 ] . These methods offer great a d v a n t a g e s in t h a t t h e cell can be examined in vitro for wavelengths in t h e visible region. Cumbersome difficulties arise due to inhomogeneities in the biological m a t e r i a l . Optical resolu- tion is generally slightly less t h a n 1 μ, a major d i s a d v a n t a g e in t r a n s - port studies. T h e use of ultraviolet and X - r a y microspectography requires fixed specimens a n d a v e r y great technical effort will be
required to yield resolutions t h a t will be useful for t h e s t u d y of i n t r a - cellular t r a n s p o r t mechanisms.
Considering t h e precision of current experimental methods, t h e study of t h e m o v e m e n t s of vesicles a n d inclusion organelles, whether m e m b r a n o u s , t u b u l a r , or filamentous, promises greater success with regard to intracellular t r a n s p o r t . F a r more refined techniques m u s t be developed to measure molecular distributions, flow p a t t e r n s , and reaction r a t e s in living cells for determining t h e p a t t e r n s a n d m e c h a - nisms of their t r a n s p o r t . C o m p u t e r - t y p e r a p i d mass-screening devices will be necessary to scan the m a n y t h o u s a n d s of electron micrographs t h a t form a composite of a single cell. T h e m a g n i t u d e of this problem is further amplified since it is necessary to analyze large n u m b e r s of cells synchronized in growth b u t fixed a t different times. A t t h e m o m e n t it seems t h a t significant information can be obtained more readily from t h e s t u d y of molecular t r a n s p o r t in p e r t i n e n t model sys- tems. These systems h a v e an added a d v a n t a g e in t h a t experimental m e a s u r e m e n t a n d theoretical analysis can supplement one another.
T h e significance of these problems should n o t be u n d e r e s t i m a t e d or shunned in view of their complexity. A n u n d e r s t a n d i n g of intracel- lular t r a n s p o r t is basic to an assay of t h e concept t h a t t h e cell func-
tions as a s e l f - s t r u c t u r i n g u n i t . I n t r a c e l l u l a r t r a n s p o r t m e c h a n i s m s a r e as y e t u n d e t e r m i n e d c o n c o m i t a n t s of m e t a b o l i c a c t i v i t y a n d changes in s t r u c t u r a l o r g a n i z a t i o n . T h e e x t e n t t o which associated m o l e c u l a r traffic in t u r n influences, r e g u l a t e s or is n e c e s s a r y for m e t a - bolic a c t i v i t y a n d s t r u c t u r a l o r g a n i z a t i o n also r e m a i n s to be answered.
REFERENCES
1. Barrnett, R. J., and Tice, L. W., in "Histochemistry a n d Cytochemistry"
(R. Wegman, ed.), p . 139. Macmillan, New York, 1963.
2. Behnke, 0 . , / . Ultrastruct. Res. 1 1 , 139 (1964).
3. Bennett, H . S., J. Biophys. Biochem. Cytol. 2 , Suppl. 99 (1956).
4. Bessis, M., " H a r v e y Lectures," Series 58, p . 125. Academic Press, New York, 1963.
5. Caro, L. G., J. Cell Biol. 1 5 , 189 (1962).
6. Caro, L. G., and Palade, G. E., Compt. Rend. Soc. Biol. 1 5 5 , 1750 (1961).
7. Caro, L. G., and Palade, G. E., J. Cell Biol. 2 0 , 473 (1964).
8. Caspersson, T., Exp. Cell Res. Suppl. 4 , 1 (1957).
9. Chance, B., Science 1 2 0 , 767 (1954).
10. Chapman-Andresen, C , in " M e t h o d s in Cell Physiology" ( D . M . Prescott, ed.), Vol. I, p. 277. Academic Press, New York, 1964.
11. Dahler, J. S. a n d Scriven, L. E., Proc. Roy Soc. (London) Ser. A 2 7 5 , 5 0 4 (1963).
12. D e D u v e , C , in "Subcellular Particles" ( T . Hayashi, e d . ) , p . 128. Ronald Press, New York, 1959.
13. Dishon, M.. Grover, Ν . B., and Weiss, G. H., J. Theoret. Biol. (1966) in press.
14. Farquhar, M . G., and Palade, G. E., J. Cell Biol. 1 3 , 55 (1962).
15. Gey, G. O., " H a r v e y Lectures," Series 50, p . 154. Academic Press, New York, 1956.
16. Grover, Ν . B., Biophys, J. 6 , 71 (1966)
17. Hayashi, T., in "Primitive Motile Systems in Cell Biology" (R. D . Allen and N . Kamiya, eds.), p . 19. Academic Press, New York, 1964.
18. Hermans, J. J., Colloid Chem. 3 , 387 (1947).
19. Hirsch, J. G., and Cohn, Ζ. Α., J. Exp. Med. 1 1 2 , 1005 (1960).
20. Hoffmann-Berling, H., Biochim. Biophys. Acta 1 9 , 453 (1956).
21. Holter, H., in "Biological Structure and Function" ( T . W. Goodwin and O. Lindberg, eds.), Vol. I , p . 605. Academic Press, New York, 1961.
22. Holter, H., and Marshall, J. M . Jr., Compt. Rend. Trav. Lab. Carlsberg (Ser. Chim.) 2 9 , 7 (1954).
23. Jarosch, R., in "Primitive Motile Systems in Cell Biology" (R. D . Allen and N . K a m i y a , eds.), p . 599. Academic Press, N e w York, 1964.
24. Kamiya, N., a n d Kuroda, K., Proc. Japan Acad. 3 3 , 201 (1957).
25. Katchalsky, Α., personal communication.
26. K a v a n a u , J . L., Develop. Biol. 7 , 22 (1963).
27. Kaye, G. I., and Pappas, G. D., J. Cell Biol. 1 2 , 457 (1962).
28. Kaye, G. I., Pappas, G. D., Donn, Α., and Mallet, H., J. Cell Biol. 1 2 , 481 (1962).
29. Ledbetter, M . C., and Porter, K. R., J. Cell Biol. 1 9 , 239 (1963).
30. Ledbetter, M . C , and Porter, K. R., Science 1 4 4 , 872 (1964).
31. Levich, V. G., "Physicochemical Hydrodynomics." Prentice-Hall, Englewood Cliffs, New Jersey, 1962.
32. Lewis, W. H., Bull. John Hopkins Hosp. 4 9 , 17 (1931).
33. Loewy, A. G., J. Cellular Comp. Physiol. 4 0 , 127 (1952).
34. Lucy, J. Α., and Glauert, Α., J. Mol. Biol. 8 , 727 (1964).
35. Marsland, D . Α., Intern. Rev. Cytol. 5 , 199 (1956).
36. Mueller, P., Rudin, D . O., Tien, H . T., and Westcott, W. C , in "Recent Progress in Surface Science" (J. F . Danielli, K. G. A. Parkhurst, A. C.
Riddiford, eds.), Vol. I, p. 379. Academic Press, New York, 1964.
37. Nakajima, H., Protoplasma 5 2 , 413 (1960).
38. Novikoff, A. B., in "Analytical Cytology" (R. C. Mellors, ed.), p. 69.
McGraw-Hill, New York, 1959.
39. Ostrowski, K., Barnard, Ε . Α., Stocka, Ζ., and Darzynkiewicz, Ζ., Exp. Cell Res. 3 1 , 89 (1963).
40. Pelc, S. R., Exp. Cell Res. Suppl. 4 , 231 (1957).
41. Perry, R. P., in " M e t h o d s in Cell Physiology" (D. M. Prescott, ed.), Vol.
I, Academic Press, New York, 1964.
42. Rosenberg, M. D., Biophys. J. 5 , 66A (1965).
43. Rosenberg, M. D., Frascati Symposium on Membranes (1965) in press.
44. Salpeter, M . M., and Bachmann, L., J. Cell Biol. 2 2 , 469 (1964).
45. Scriven, L. E., Chem. Eng. Sci. 1 2 , 98 (1960).
46. Seligman, A. M., in "Histochemistry and Cytochemistry" (R. Wegmann, ed.), p. 203. Macmillan, New York, 1963.
47. Strother, G. K., and Ackerman, E., Biochim. Biophys. Acta 4 7 , 317 (1961).
48. Taylor, A. C , and Robbins, E., Develop. Biol. 7, 660 (1963).
49. Thompson, T. E., in "Cellular Membranes in D e v e l o p m e n t " (M. Locke, ed.), p. 83. Academic Press, New York, 1964.
50. Ts'o, P . 0 . P., Eggman, L., and Vinograd, J., Biochim. Biophys. Acta 2 5 , 532 (1957).
51. Wohlfarth-Bottermann, Κ. E., in "Primitive Motile Systems in Cell Biology"
(R. D . Allen and N . Kamiya, eds.), p. 79. Academic Press, New York, 1964.

![FIG. 2. Translocation through endothelial cells of cornea. F r o m Kaye and P a p p a s [27] and K a y e et al [28]](https://thumb-eu.123doks.com/thumbv2/9dokorg/1138374.81147/8.648.96.556.271.682/fig-translocation-endothelial-cells-cornea-f-kaye-p.webp)