Capsaicin-sensitive sensory nerves exert complex regulatory functions in the serum-transfer mouse model of autoimmune arthritis
Éva Borbély
a,b,c,1, Bálint Botz
a,b,c,1, Kata Bölcskei
a,b,c, Tibor Kenyér
a, László Kereskai
d, Tamás Kiss
b, János Szolcsányi
a,b,c,f, Erika Pintér
a,b,c,f, Janka Zsófia Csepregi
e, Attila Mócsai
e, Zsuzsanna Helyes
a,b,c,f,g,⇑aDepartment of Pharmacology and Pharmacotherapy, University of Pécs, Medical School, Pécs, Hungary
bJános Szentágothai Research Centre, Molecular Pharmacology Research Team, University of Pécs, Pécs, Hungary
cCentre for Neuroscience, University of Pécs, Medical School, Pécs, Hungary
dDepartment of Pathology, University of Pécs, Medical School, Pécs, Hungary
eDepartment of Physiology, and MTA-SE ‘‘Lendület’’ Inflammation Physiology Research Group, Semmelweis University, School of Medicine, Budapest, Hungary
fPharmInVivo Ltd, Pécs, Hungary
gMTA-PTE NAP B Pain Research Group, Hungary
a r t i c l e i n f o
Article history:
Received 23 September 2014
Received in revised form 18 November 2014 Accepted 9 December 2014
Available online 15 December 2014
Keywords:
Capsaicin-sensitive sensory nerves Pain
Inflammation Somatostatin
Matrix-metalloproteinase
a b s t r a c t
Objective:The K/BxN serum-transfer arthritis is a widely-used translational mouse model of rheumatoid arthritis, in which the immunological components have thoroughly been investigated. In contrast, little is known about the role of sensory neural factors and the complexity of neuro–immune interactions. There- fore, we analyzed the involvement of capsaicin-sensitive peptidergic sensory nerves in autoantibody- induced arthritis with integrative methodology.
Methods: Arthritogenic K/BxN or control serum was injected to non-pretreated mice or resiniferatoxin (RTX)-pretreated animals where capsaicin-sensitive nerves were inactivated. Edema, touch sensitivity, noxious heat threshold, joint function, body weight and clinical arthritis severity scores were determined repeatedly throughout two weeks. Micro-CT andin vivooptical imaging to determine matrix-metallopro- teinase (MMP) and neutrophil-derived myeloperoxidase (MPO) activities, semiquantitative histopathol- ogical scoring and radioimmunoassay to measure somatostatin in the joint homogenates were also performed.
Results: In RTX-pretreated mice, the autoantibody-induced joint swelling, arthritis severity score, MMP and MPO activities, as well as histopathological alterations were significantly greater compared to non-pretreated animals. Self-control quantification of the bone mass revealed decreased values in intact female mice, but significantly greater arthritis-induced pathological bone formation after RTX-pretreat- ment. In contrast, mechanical hyperalgesia from day 10 was smaller after inactivating capsaicin-sensitive afferents. Although thermal hyperalgesia did not develop, noxious heat threshold was significantly higher following RTX pretreatment. Somatostatin-like immunoreactivity elevated in the tibiotarsal joints in non-pretreated, which was significantly less in RTX-pretreated mice.
Conclusions: Although capsaicin-sensitive sensory nerves mediate mechanical hyperalgesia in the later phase of autoantibody-induced chronic arthritis, they play important anti-inflammatory roles at least partially through somatostatin release.
Ó2014 The Authors. Published by Elsevier Inc. This is an open access article under the CC BY license (http://creativecommons.org/licenses/by/4.0/).
1. Introduction
Rheumatoid arthritis (RA) is chronic autoimmune disease char- acterized by the destruction and deformation of the joints leading
to persistent pain, movement disability and decreased life quality.
It is a great public health problem worldwide due to its high inci- dence and prevalence, unsatisfactory therapeutic outcomes and unfavorable life expectancy (Kourilovitch et al., 2014; Jones et al., 2003). Despite promising novel drugs introduced recently in its pharmacotherapy, we still have to cope with several resistant cases and severe drug-induced adverse effects (Schett and Gravallese, 2012; Alarcón, 2000). Although our knowledge about the immuno- logical aspects of the pathophysiological mechanisms has
http://dx.doi.org/10.1016/j.bbi.2014.12.012
0889-1591/Ó2014 The Authors. Published by Elsevier Inc.
This is an open access article under the CC BY license (http://creativecommons.org/licenses/by/4.0/).
⇑Corresponding author at: Department of Pharmacology and Pharmacotherapy, University of Pécs, Medical School, Szigeti u. 12., H-7624 Pécs, Hungary. Tel.: +36 72 536 000/35591; fax: +36 72 536 218.
E-mail address:zsuzsanna.helyes@aok.pte.hu(Z. Helyes).
1 These authors made equal contributions to this work.
Contents lists available atScienceDirect
Brain, Behavior, and Immunity
j o u r n a l h o m e p a g e : w w w . e l s e v i e r . c o m / l o c a t e / y b r b i
extensively increased in the last decade, the regulatory role of sen- sory nerves and the complexity of neuro–immune interactions in this condition are still not understood (Levine et al., 2006;
Pongratz and Straub, 2010; Meinel et al., 2013; Stangenberg et al., 2014).
Capsaicin-sensitive peptidergic sensory nerves densely inner- vate the joint capsule and the synovium, which do not only mediate pain (classical afferent function), but also play an important role in inflammation via sensory neuropeptide release (efferent function).
The Transient Receptor Potential Vanilloid 1 (TRPV1) non-selective cation channel located on these nerves is activated and sensitized by a variety of exogenous irritants, such as capsaicin, and resinifera- toxin (RTX), as well as endogenous molecules like protons, bradyki- nin, prostanoids, tumor-necrosis factor-
a
, nerve growth factor, gasotransmitters or lipid peroxidase products (Yoo et al., 2014).Many of these are crucial participants of inflammatory processes in RA. As a result of activation of the capsaicin-sensitive nerve ter- minals, sensory neuropeptides are released, such as the proinflam- matory tachykinins (substance P, neurokinin A) and calcitonin-gene related peptide (CGRP) responsible for vasodilation and inflamma- tory cell recruitment (neurogenic inflammation) (Maggi, 1995;
Szolcsanyi, 1996), as well as somatostatin, which is a potent antiin- flammatory and antinociceptive agent. We have provided several lines of evidence in a variety of inflammation models that the over- all role of these fibers depends on the functional significances of the simultaneously released pro- and antiinflammatory peptides in the respective pathophysiological processes (Pintér et al., 2014). We have also shown that sensory nerve-derived somatostatin is an important endogenous inhibitor in the adjuvant-induced arthritis model of the rat (Helyes et al., 2004).
The pathophysiological relevance of these peptides in humans is beyond doubt, since increased proinflammatory and decreased antiinflammatory neuropeptide levels have been demonstrated in the serum and/or synovial fluid of RA patients (Anichini et al., 1997; Larsson et al., 1991; Denko and Malemud, 2004).
Investigating rheumatoid arthritis mechanisms in animals is difficult; therefore there are many different rodent models which can more or less mimic the main symptoms of the disease (Bevaart et al., 2010; Zhang et al., 2012; Boettger et al., 2010).
The K/BxN serum-transfer arthritis is a widely-used translational mouse model of RA, it shares a lot of similarities to the human dis- ease, e.g. swelling of distal joints of all the paws with erosive syno- vitis, caused by the activation of neutrophils, macrophages, complement system which play a pivotal role in the induction and maintenance of arthritis (Kouskoff et al., 1996; Korganow et al., 1999; Fukushima et al., 2010). The immunological compo- nents of this model have thoroughly been investigated (Németh et al., 2010; Hickman-Brecks et al., 2011), but nothing is known about the role of sensory neural factors and the complexity of neuro–immune interactions. Therefore, we analyzed the involve- ment of capsaicin-sensitive peptidergic sensory nerves in autoanti- body-induced arthritis with integrative methodology after the functional impairment of these fibers with high dose RTX pretreat- ment (desensitization).
2. Material and methods 2.1. Ethics statement
Experiments were carried out according to the 1998/XXVIII Act of the Hungarian Parliament on Animal Protection and Consider- ation Decree of Scientific Procedures of Animal Experiments (243/1988), complied with the recommendations of IASP, and approved by the Ethics Committee on Animal Research of Univer- sity of Pécs (licence: BA 02/2000-2/2012).
2.2. Experimental animals
Male and female C57Bl/6 mice (10–12-week-old; 25–30 g) bred and kept in the Laboratory Animal House of the Department of Pharmacology and Pharmacotherapy of the University of Pécs at 24–25°C under a 12-h light–dark cycle were used in all studies.
Standard mouse chow and water were providedad libitum.
2.3. Resiniferatoxin pretreatment
Pretreatment with the ultrapotent TRPV1 agonist resiniferatox- in (RTX, Sigma–Aldrich; 30, 70, 100
l
g/kg s.c. on 3 consecutive days) leads to long-lasting defunctionalization of capsaicin-sensi- tive nerves (desensitization) (Szolcsanyi et al., 1990). Two weeks later the success of the pretreatment was verified by the lack of eye-wiping after capsaicin drops (50l
l, 0.1%) (Helyes et al., 2004).2.4. Induction of arthritis
Chronic arthritis of male and female C57Bl/6 mice was induced by intraperitoneal (i.p.) injection of 150–150
l
l of K/BxN serum on the days 0 and 3. Control groups of intact animals were treated with BxN (not arthritogenic/control) serum following the same protocol.2.5. Assessment of arthritis severity and paw edema
Hind paw volume was measured by plethysmometry (Ugo Basile 7140, Comerio, Italy) (Helyes et al., 2004; Szabó et al., 2005). Arthritic changes were semiquantitatively scored using a grading scale of 0–10 (0–0.5: no change, 10: maximal inflamma- tion) by evaluating edema and hyperemia (Németh et al., 2010).
Volumes and scores were assessed before serum injection and every day during the 2-week period.
2.6. Measurement of mechanical and thermal hyperalgesia
Mechanonociceptive threshold of the paw was determined by dynamic plantar aesthesiometry (Ugo Basile 37400, Comerio, Italy) before and after serum administration. Mechanical hyperalgesia was expressed as % of initial, control mechanonociceptive thresh- olds (Helyes et al., 2004; Szabó et al., 2005). The thermonociceptive threshold of the paw was determined on increasing temperature hot plate (IITC Life Sciences, Woodland Hills, CA, USA) by nocifen- sive reactions (lifting, licking, shaking) or reaching the maximum value (53°C) (Almási et al., 2003).
2.7. Assessment of joint function (grid test)
An easy and reproducible method to determine grasping ability correlating with joint function. Mice were placed on a horizontal wire-grid, then it was turned over and the latency to fall was deter- mined (Németh et al., 2010).
2.8. Measurement of arthritis induced weight loss
As a typical sign of systemic effect of arthritis, mice lost weight after serum administration. Weight measurements were per- formed daily and weight loss was expressed in % of control values.
2.9. In vivo bioluminescence imaging of myeloperoxidase-activity
Luminol bioluminescence (BLI; 5-amino-2,3-dihydro-1,4- phthalazine-dione) correlates with neutrophil myeloperoxidase activity in arthritisin vivo(Chen et al., 2004; Gross et al., 2009).
Na–luminol (150 mg/kg i.p., Sigma–Aldrich) dissolved in PBS
(20 mg/ml) was injected on days 0, 2 and 6. Images were acquired 10 min later with IVIS Lumina II (PerkinElmer, Waltham, USA; 60 s acquisition, F/Stop = 1, Binning = 8). Identical Regions of Interests (ROIs) were applied around the ankles and luminescence was expressed as total radiance (total photon flux/s).
2.10. In vivo fluorescence imaging of matrix-metalloproteinase activity
Matrix-metalloproteinase (MMP) activity was assessedin vivo on days 5 and 8 using MMPSense680 (PerkinElmer), an activatable fluorescent imaging probe for MMP-2, -3, -9 and -13 according to the manufacturer’s instructions (2 nmol/subject i.v.). Measure- ments were performed with the FMT 2000 fluorescence molecular tomography system 24 h later (PerkinElmer). Three-dimensional reconstructions of the ankles were made, isocontour ROIs were applied, and MMP was expressed as pmol fluorophore.
2.11. In vivo micro-computed tomography (micro-CT) analysis of the periarticular bone structure
The right tibiotarsal joints were repeatedly (days 0, 7, 14) scanned by SkyScan 1176in vivomicro-CT (Bruker, Kontich, Bel- gium) with 17.5
l
m voxel size. Changes of bone structure were evaluated by CT AnalyserÒ software. Standard size ROIs were applied around the periarticular tibia and fibula regions, andaround the tibiotarsal and tarsometatarsal joints. Bone volume (
l
m3) was quantified and expressed as a percentage of the total ROI volume.2.12. Histological processing and assessment of joint inflammation
Ankle joints excised on day 14 were fixed, decalcified and dehy- drated, embedded in paraffin, sectioned (3–5
l
m) (Helyes et al., 2004; Szabó et al., 2005) and stained with hematoxylin–eosin or Safranin O for detecting collagen deposition and fibroblasts. Histo- pathological changes were scored by a pathologist blinded from the study on the basis of (1) areolar tissue size and mononuclear cell infiltration, (2) synovial cell proliferation, (3) fibroblast num- ber and collagen deposition to create composite arthritis scores (between 0 and 9) (Botz et al., 2014).2.13. Determination of somatostatin-like immunoreactivity (SOM-LI) in tissue homogenates
Separate groups of mice were sacrificed in deep anesthesia on day 10 when both swelling and hyperalgesia were remarkable.
The tibiotarsal joints were homogenized in a solution containing 20 mM KH2PO4and K2HPO4for 2 min at 24,000 rpm with Miccra D-9 Digitronic device (Art-moderne Laborteknik, Germany).
Homogenates were centrifuged for 10 min at 4000 rpm afterwards Fig. 1.Serum-induced edema and arthritis score throughout the 2 weeks experimental period. Data points represent the percentage increase of the paw volume of male (A) and female (B) mice compared to the initial control values and the absolute values of arthritis scores for male (C) and female animals (D) (n= 4–5/non-inflamed groups,n= 6–
8/arthritic groups;⁄p< 0.05,⁄⁄p< 0.01,⁄⁄⁄p< 0.001 vs. non-pretreated; two-way ANOVA + Bonferroni’s modifiedt-test).
for 15 min at 10,000 rpm and the supernatants were collected for SOM-LI determination with a specific and sensitive radioimmuno- assay (RIA) (Németh et al., 1996).
2.14. Statistical analysis
All functional, histopathological and CT results were presented and evaluated separately for male and female mice, data points represent means ± SEM. Hyperalgesia, edema and weight loss were evaluated by repeated measures two-way analysis of variance (ANOVA) + Bonferroni’s modified t-test, semiquantitative clinical and composite histopathological scores by non-parametric Krus- kal–Wallis test + Dunn’s post-test, micro-CT results by two-way
ANOVA + Dunnett and Tukey post-tests to evaluate the time- dependent self-control changes and the different groups, respec- tively. Bioluminescence and fluorescence imaging, as well as somatostatin-LI were analyzed by Student’s t-test for unpaired comparisons.p< 0.05 was considered to be significant.
3. Results
3.1. Increased joint edema after desensitization of capsaicin-sensitive sensory nerves
In non-pretreated arthritic mice an approximately 45% edema developed in both males and females, which was maintained till Fig. 2.Serum-induced hyperalgesia throughout the 2 weeks experimental period. Data points represent the percentage decrease of mechanonociceptive threshold for male (A) and female (B) mice, and the absolute values of noxious heat threshold for male (C) and female animals (D) (n= 4–5/control non-inflamed groups,n= 6–8/arthritic groups;
⁄p< 0.05,⁄⁄p< 0.01,⁄⁄⁄p< 0.001 vs. non-pretreated; two-way ANOVA + Bonferroni’s modifiedt-test).
Fig. 3.Change of neutrophil-derived myeloperoxidase activity. Panel (A) shows representative pretreatment control images, whereas panels (B and C) demonstrate inflammatory neutrophil activity on day 2 and 6 following arthritis induction, respectively. (D) Quantification of luminol bioluminescence in the diseased ankle joints. (n= 6–
8 male mice/group,###p< 0.001 vs. controls,⁄⁄⁄p< 0.001 vs. non-pretreated; Studentt-test).
the end (day 11) of the experiment. In RTX-desensitized arthritic animals this swelling was significantly higher in both genders dur- ing the whole study with a maximum of 90–95% (Fig. 1A and B).
Similarly, arthritis scores reached a maximum of 7 in mice without pretreatment and 9 in RTX-pretreated animals showing that the significant increase in paw volume was visible on all limbs between days 2 and 7–8 in male female mice, respectively (Fig. 1C and D).
3.2. Attenuated late mechanical hyperalgesia in RTX-desensitized mice
Mechanical hyperalgesia (nociceptive threshold decrease) in non-pretreated arthritic mice reached an approximately 25–30%
after 5 days, which further increased to 45% by days 10 in both male and female mice (Fig. 2A and B). Significant reduction of mechanical hyperalgesia was measured in RTX-desensitized ani-
mals from day 10. Despite the development of mechanical hyper- algesia, the noxious heat threshold was not influenced by the arthritis. However, the thermonociceptive threshold of RTX-pre- treated animals was significantly higher compared to mice without pretreatment between days 1 and 5 (Fig. 2C and D).
3.3. Similar weight loss and impaired joint function in non- desensitized and RTX-desensitized mice
Arthritis resulted in a 10–15% weight loss and 50% decrease of time spent on the grid by days 4 and 5, respectively, both in the non-pretreated and RTX-pretreated groups (Supplementary data).
3.4. Greater neutrophil-activity in desensitized mice in the acute arthritis phase
Luminol-BLI revealed a remarkable increase in neutrophil- derived MPO activity in the arthritic ankle joints of both groups, being significantly higher in RTX-pretreated mice in the early phase (day 2). This difference ceased during the later phase by day 6 (Fig. 3).
3.5. Increased MMP activity in RTX-pretreated animals
Fluorescent molecular tomography revealed that a considerable increase in MMP activity occurred in the inflamed ankle joints of arthritic animals similarly on days 5 and 8, but no signal could be detected in intact, non-inflamed mice. MMP activity signifi- cantly enhanced after functional impairment of the capsaicin-sen- sitive afferents on day 5 when the differences in swelling and arthritis severity scores of the two groups were the greatest (Fig. 4).
3.6. Altered inflammation-induced structural changes in the bone of RTX-pretreated female mice
There were considerable differences on the micro-CT scans between bone volume/total volume ration of male and female mice under normal, intact conditions, the basal bone mass in both the tibiotarsal and distal tibial regions was lower in the female group as compared to age-matched males.
RTX-desensitization alone did not induce any change in the bone mass in male animals, it evoked a moderate, but significant decrease in females. Self-control quantitative analysis of the bone structure revealed minimal, but statistically significant increase in the ankle joint of non-pretreated both males and females already on day 7 due to pathological new bone formation, which was not observed in the distal tibia. Meanwhile, in RTX-pretreated arthritic females bone mass gradually and significantly increased in the ankle and the tibia reaching a remarkable, 20% gain by day 14 compared to the initial control values of the same animals (Fig. 5).
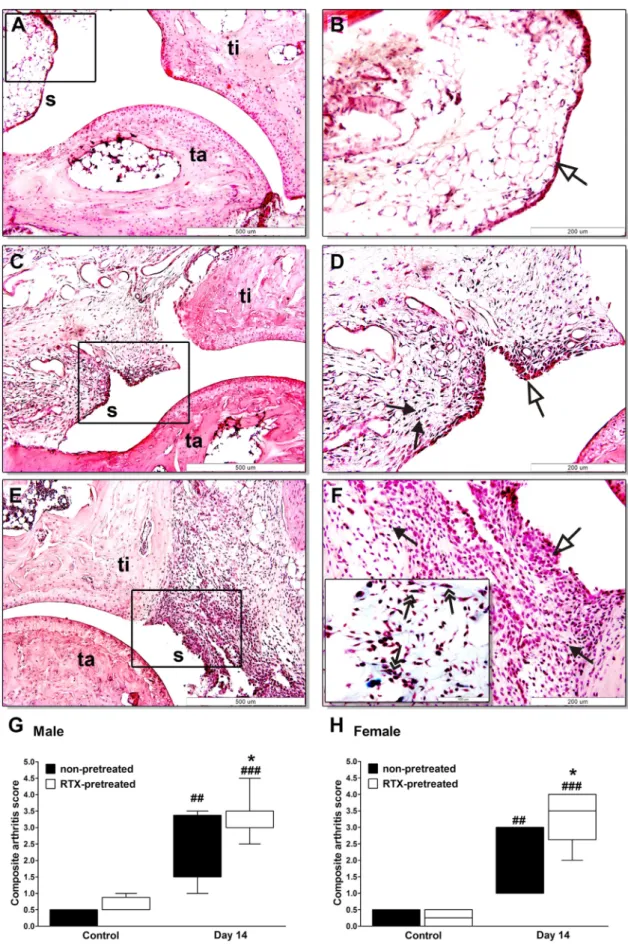
3.7. More severe arthritic histopathological alterations after RTX pretreatment
There was no histopathological difference between the intact joints of untreated and RTX-pretreated animals in either males or females (Fig. 6A and B). In non-pretreated arthritic mice character- istic chronic arthritic changes developed by day 14, such as syno- vial hyperplasia with a minimal mononuclear infiltration, moderate fibroblast formation and collagen deposition (Fig. 6C and D). In desensitized arthritic animals these changes were more pronounced with significantly greater synovial swelling, higher number of fibroblasts and more collagen (Fig. 6E and F). Semiquan- titative scoring of these parameters showed remarkable worsening Fig. 4.Matrix-metalloproteinase activity in the diseased hind limbs. (A) Represen-
tative pretreatment control images, (B and C) demonstrate inflammatory matrix- metalloproteinase activity on day 5 and 8 following arthritis induction. (D) Quantification the amount of fluorophore in the inflamed ankle joints (n= 3–4 male mice/group,⁄p< 0.05 vs. non-pretreated, + indicates that the probe was tested in intact mice in a self-control manner before the induction of the inflammation, but remained below the detection threshold; Studentt-test).
effect of RTX pretreatment on these characteristic histopatholo- gical features in both sexes (Fig. 6G and H).
3.8. RTX desensitization decreases arthritis-induced elevation of somatostatin-LI in the tissue homogenates
Somatostatin-LI significantly increased to 75.54 ± 3.07 fmol/g wet tissue in the arthritic paws of non-pretreated mice compared to their intact controls (25.19 ± 1.53 fmol/g wet tissue), while in RTX-desensitized arthritic animals its inflammation-induced elevation was significantly smaller, from 28.43 ± 1.19 to 62.39 ± 2.58 fmol/g wet tissue (p= 0.0059; Student’s t-test for unpaired comparisons).
4. Discussion
We provided here the first evidence that capsaicin-sensitive peptidergic sensory nerves play an important and complex regula- tory role in a primarily autoimmune arthritis model of the mouse.
Inactivation of these fibers results in significantly more severe characteristics of arthritis, such as increased swelling, MMP-activ- ities and ROS production, inflammatory cell accumulation and his- topathological alterations, but despite the enhanced inflammation decreased late mechanical hyperalgesia (Table 1.).
Peptidergic afferents densely innervate the synovium and the joint capsule and are involved in the pathophysiology of RA through the release of sensory neuropeptides and consequent modulation of cytokine production (Konttinen et al., 2006). Increasing evidence suggests that modulating the function of these nerves might open new perspectives in arthritis therapy (Helyes et al., 2004; Szabó et al., 2005). The present results are perfectly supported by our rev- olutionary findings obtained in the adjuvant arthritis rat model 10 years ago, when we described a potent protective function of capsaicin-sensitive afferents via somatostatin release (Helyes et al., 2004). However, that time we had no experimental tools to have a deeper insight into the underlying mechanisms. Our optical in vivoimaging methods provide a great opportunity to investigate the cellular components of the arthritic process and provide direct evidence for the importance of sensory–immune interactions. Acti- vation of the capsaicin-sensitive afferents inhibits both MPO and MMP activities, decreases leukocyte activity, and interestingly, in
females even attenuates pathological new bone formation.
Although there was no difference between male and female mice in any inflammatory parameters, our unique finding obtained by quantification of the self-control micro-CT scans is that there was in fact a decreased bone mass in females compared to age-matched males. Furthermore, in females inactivation of the capsaicin-sensi- tive afferents resulted in basically decreased bone volume, but the arthritis-induced pathological bone formation was more severe.
These results are supported by recent data showing remarkably lower BV/TV morphological parameter and higher histopathological osteophyte score in old female C57Bl/6 mice compared to the age- matched males (Cai et al., 2014). Additionally, the same BV/TV parameter determined in the human radius also provided similar results; this value was lower in all age groups of women compared to men (Vanderschueren et al., 2014). Since unlike in female mice, in RTX-pretreated males we could not detect a significantly increased pathological bone formation as compared to the non-pretreated animals, it can be suggested that the androgens might have a pro- tective role on the bones particularly under inflammatory condi- tions. It is clear that sex steroids are important influencing factors in osteoclast/osteocyte/chondrocyte functions and that in men not only the bone mineral density, but also the bone structure (bone length, width and rigidity) differs from these in women (Vanderschueren et al., 2014). However, the precise mechanisms of peptidergic sensory nerve activation and bone turnover regula- tion are still unclear, because there are very few data about the importance of afferents/TRP channels in chondrocyte–osteoclast–
osteoblast functions. Desensitization itself is reported to affect only the nerve-endings and not the non-neural TRPV1 channels (Czikora et al., 2013; Kun et al., 2012; Bíró et al., 1998), but influencing the endovanilloid/endocannabinoid system in chondro- and osteocytes can be involved in bone formation and resorption, although these data are available only on osteoclasts from osteoporosis patients (Rossi et al., 2011).
It is well-known that in the early stage of the inflammatory reac- tion in this model, the activation of neutrophils is a predominant component (Bevaart et al., 2010) and MPO is the major constituent of neutrophil azurophilic granules. Moreover, it is recently shown that in patients with active rheumatoid arthritis a very high concen- tration of MPO can be detected and it positively correlates with IgM levels (Wang et al., 2014). Similarly to what we found in this arthri- tis model, elevated MPO-levels in RTX-desensitized mice were pre- Fig. 5.Bone structural changes in the inflamed region. (A) Representative micro-CT images of the same mice, in intact state and on day 14. (B and C) Bone volume/total volume ratio in the ankle joint, expressed as raw data and as percentage of the initial self-controls. (D and E) Bone volume/total volume ratio in the distal tibia, expressed as raw data and as percentage of the initial self-controls (n= 6/group,#p< 0.05,##p< 0.01,###p< 0.001 vs. controls,⁄⁄p < 0.01,⁄⁄⁄p < 0.001 vs. non-pretreated; two-way ANOVA + Dunnett and Tukey post-tests).
Fig. 6.Histopathological changes of the ankle joints. Panels (A and B) show representative histopathological pictures of an intact tibiotarsal joint (ti: tibia, ta: tarsus, s:
synovium), panels (C and D) demonstrate the joint structure of a non-pretreated mouse on day 14 after arthritogenic serum administration, panels (E and F) show the significantly pronounced arthritic changes of RTX-pretreated animals. (G and H) Semiquantitative histopathological scoring on the basis of synovial enlargement (white arrows), inflammatory cell accumulation (black arrows), fibroblast formation with collagen deposition (two headed arrows). Box plots represent the composite scores for male and female animals (n= 4–5/control non-inflamed groups,n= 6–8/arthritic groups;##p< 0.01,###p< 0.001 vs. controls,⁄p< 0.05 vs. non-pretreated; Kruskal–Wallis followed by Dunn’s post-test).
viously detected in LPS-induced acute airway inflammation (Elekes et al., 2007). Our MMP results are also in good correlation with spo- radic earlier evidence demonstrating that TRPV1 receptor activation results in decreased MMP-9-secretion (Tauber et al., 2012). Both MPO and MMP are important, but not the exclusive participants of this complex inflammatory process, and their correlation with either edema formation, pain or bone pathophysiology is only indi- rect. Although we do not have data obtained from later phase of the model, the activation of these enzymes leads to a variety of patho- physiological alterations including the recruitment of different inflammatory cells, as well as activation of other enzymes (e.g.
NOS;Arnhold and Flemmig, 2010), which can result in a propaga- tion of the inflammatory processes or even deformation of bones, despite lower MPO or MMP levels.
To understand the mechanisms involved in the inhibitory action of capsaicin-sensitive afferents, somatostatin was determined in the tibiotarsal joint homogenates. Somatostatin and its receptors (sst1–5) are widely distributed throughout the body and show a prominent expression in the sensory nerve endings (Pintér et al., 2006), but they are also present on immune cells suggesting its important regulatory function in inflammatory diseases (Pintér et al., 2014). Somatostatin ameliorates RA symptoms not only in murine models (Imhof et al., 2011), but also as a chronic intra-artic- ular treatment in humans (Fioravanti et al., 1995; Paran et al., 2001).
We found that it was increased in the arthritic joints, but signifi- cantly decreased after RTX-pretreatment. Therefore, its inflamma- tion-evoked elevation is likely to be derived from the capsaicin- sensitive fibers, and be involved the protective actions of these nerves. The MMP-activity increase in desensitized mice is also con- sistent with these results, since somatostatin was reported to reduce MMP-1, -2 and -9 mRNA expression and MMP-1 production by synovial cells of RA patients (Takeba et al., 1997).
Intriguingly, in contrast to our earlier findings in RTX-pretreated rats in the adjuvant arthritis model (Helyes et al., 2004), the increased inflammation was not accompanied by a proportionally enhanced mechanical hyperalgesia. In the late phase, when inflam- matory signs were attenuated, but mechanical hyperalgesia was still present, it was even significantly milder in RTX-pretreated mice. Compared to the greater severity of inflammation, mechani- cal hyperalgesia is clearly smaller in the RTX-pretreated group.
Therefore, capsaicin-sensitive nerves might participate in arthritic mechanical hyperalgesia during the whole process, but the differ- ence was only manifest when the degree of inflammation was equal in both groups. Capsaicin-sensitive nociceptors are polymodal, i.e.
activated by noxious heat, chemical and mechanical stimuli, and their role is provenin vivo in thermonociception (Almási et al.,
2003; Cavanaugh et al., 2009; Mishra and Hoon, 2010; Danigo et al., 2014; Bölcskei et al., 2010). In contrast, mechanonociceptive thresholds were not different in desensitized mice if von Frey fila- ments were used (Cavanaugh et al., 2009; Mishra and Hoon, 2010), but increased mechanical thresholds were found with the Randall–Selitto pressure test (Danigo et al., 2014). It is likely, that the latter method, as well as our aesthesiometer activated a differ- ent mechanonociceptive neurone subset, which explains the differ- ence after RTX pretreatment. Furthermore, recent data showed that in the chronic phase of the K/BxN arthritis, when swelling and hyperemia disappeared, neuropathic pain developed. It was only relieved by gabapentin, a typical adjuvant analgesic for neuropathy (Christianson et al., 2010). In heat and mechanical hyperalgesia TRPV1 channels are the most important participants (Sousa- Valente et al., 2014; Hulse et al., 2014). In agreement with these results we detected a significantly attenuated mechanical hyperal- gesia in RTX-pretreated animals only in the late phase (from day 10). This strongly suggests that TRPV1 channels play a pivotal role in pain mediation, as described in neuropathy (Brito et al., 2014) and arthritis models (Fernandes et al., 2011; Kelly et al., 2013), but the time course-dependent involvement of these channels has never been proven. Thermal hyperalgesia is neither characteristic in K/BxN arthritis, nor in RA patients (Edwards et al., 2009; Leffler et al., 2002). However, we found a significant difference in the RTX-pretreated group during the 1st week, which is probably due to the well-known heat threshold increasing effect of RTX (Almási et al., 2003), but arthritis did not alter thermosensitivity.
Based on all these data capsaicin-sensitive peptidergic sensory nerves and sensory–immune interactions are important regulators of immune-mediated arthritis. Their activation inhibits the charac- teristic arthritis symptoms (edema, inflammatory cell activation and functions) at least partially through somatostatin release, but despite this potent anti-inflammatory role, they mediate the later pain response. These results are in agreement with our previous results obtained from LPS-induced pneumonitis, where the inacti- vation of the capsaicin-sensitive nerves leads to greater inflamma- tion (cell accumulation, edema), but decreased bronchial hyperreactivity (Elekes et al., 2007). In contrast, it has very recently been published that both RTX-desensitization and TRPV1 receptor deficiency ameliorate the clinical severity in the IL-23-mediated, T-cell dependent psoriasiform dermatitis model (Riol-Blanco et al., 2014).
It can be concluded that neuropeptide-containing sensory nerves exert a complex regulatory function in inflammatory condi- tions, and the overall effect of their activation depend on the tis- sues and the pathophysiological mechanisms of the disease.
Acknowledgments
The research infrastructure was supported by SROP-4.2.2.A-11/
1/KONV-2012-0024, the MTA–National Brain Research Programme B (888819, Hungary, Zs. Helyes). Bálint Botz was supported by the TÁMOP 4.2.4. A/2-11-1-2012-0001 ‘‘National Excellence Program’’
of the European Union and the State of Hungary co-financed by the European Social Fund. This work was supported by the European Research Council – Belgium (Starting Independent Investi- gator Award No. 206283 to Attila Mócsai), and the Wellcome Trust – United Kingdom (International Senior Research Fellowship No. 087782 to Attila Mócsai).
The authors are grateful to Anikó Perkecz for her expert help in histological processing, to Teréz Bagoly for the somatostatin mea- surements and to Ágnes Kemény for editing Fig. 5. We thank Diane Mathis and Christophe Benoist for the KRN transgene-positive mice. The present scientific contribution is dedicated to the 650th anniversary of the foundation of the University of Pécs, Hungary.
Table 1
Summary of functional, morphological and immunological alterations in RTX- pretreated mice compared to the non-pretreated animals. Edema formation, arthritis score, neutrophil activity, matrix-metalloproteinase activity and histopathological changes were significantly aggravated, while paw somatostatin level and mechanical hyperalgesia were significantly attenuated in RTX-pretreated animals. Noxious heat threshold, weight loss and joint function did not differ between the two groups.
Arthritis parameters Effect of RTX-pretreatment
on arthritis changes
Paw edema "
Arthritis score "
Neutrophil-activity "
Matrix-metalloproteinase activity "
Pathological bone formation (in females) "
Histopathological changes "
Mechanical hyperalgesia ;
Paw somatostatin level ;
Heat threshold –
Joint function –
Weight loss –
Appendix A. Supplementary data
Serum-induced weight loss and impaired joint function: data points represent the percentage change of weight loss of male (A) and female (B) mice compared to the initial control values and the absolute values of time spent on the grid for male (C) and female animals (D) (n= 4–5/control groups,n= 6–8/arthritic groups; two-way ANOVA + Bonferroni’s modifiedt-test).
Supplementary data associated with this article can be found, in the online version, athttp://dx.doi.org/10.1016/j.bbi.2014.12.012.
References
Alarcón, G.S., 2000. Methotrexate use in rheumatoid arthritis. A Clinician’s perspective. Immunopharmacology 47, 259–271.
Almási, R., Pethö, G., Bölcskei, K., Szolcsányi, J., 2003. Effect of resiniferatoxin on the noxious heat threshold temperature in the rat: a novel heat allodynia model sensitive to analgesics. Br. J. Pharmacol. 139, 49–58.
Anichini, M., Cesaretti, S., Lepori, M., Maddali, Bongi S., Maresca, M., Zoppi, M., 1997.
Substance P in the serum of patients with rheumatoid arthritis. Rev. Rhum.
Engl. Ed. 64, 18–21.
Arnhold, J., Flemmig, J., 2010. Human myeloperoxidase in innate and acquired immunity. Arch. Biochem. Biophys. 500, 92–106. http://dx.doi.org/10.1016/
j.abb.2010.04.008.
Bevaart, L., Vervoordeldonk, M.J., Tak, P.P., 2010. Evaluation of therapeutic targets in animal models of arthritis: how does it relate to rheumatoid arthritis? Arthritis Rheum. 62, 2192–2205.
Bíró, T., Maurer, M., Modarres, S., Lewin, N.E., Brodie, C., Acs, G., Paus, R., Blumberg, P.M., 1998. Characterization of functional vanilloid receptors expressed by mast cells. Blood 91, 1332–1340.
Boettger, M.K., Weber, K., Gajda, M., Bräuer, R., Schaible, H.G., 2010. Spinally applied ketamine or morphine attenuate peripheral inflammation and hyperalgesia in acute and chronic phases of experimental arthritis. Brain Behav. Immun. 24, 474–485.http://dx.doi.org/10.1016/j.bbi.2009.12.002.
Bölcskei, K., Tékus, V., Dézsi, L., Szolcsányi, J., Petho, G., 2010. Antinociceptive desensitizing actions of TRPV1 receptor agonists capsaicin, resiniferatoxin and N-oleoyldopamine as measured by determination of the noxious heat and cold thresholds in the rat. Eur. J. Pain 14, 480–486.
Botz, B., Bölcskei, K., Kereskai, L., Kovács, M., Németh, T., Szigeti, K., Horváth, I., Máthé, D., Kovács, N., Hashimoto, H., Regl}odi, D., Szolcsányi, J., Pintér, E., Mócsai, A., Helyes, Z., 2014. Differential regulatory role of pituitary adenylate-cyclase activating polypeptide in the serum-transfer-induced arthritis model. Arthritis Rheumatol. 66, 2739–2750.http://dx.doi.org/10.1002/art.38772.
Brito, R., Sheth, S., Mukherjea, D., Rybak, L.P., Ramkumar, V., 2014. TRPV1: a potential drug target for treating various diseases. Cells 3, 517–545.
Cai, A., Hutchison, E., Hudson, J., Kawashima, Y., Komori, N., Singh, A., Brush, R.S., Anderson, R.E., Sonntag, W.E., Matsumoto, H., Griffin, T.M., 2014. Metabolic enrichment of omega-3 polyunsaturated fatty acids does not reduce the onset of idiopathic knee osteoarthritis in mice. Osteoarthritis Cartilage 22, 1301–
1309.http://dx.doi.org/10.1016/j.joca.2014.06.033.
Cavanaugh, D.J., Lee, H., Lo, L., Shields, S.D., Zylka, M.J., Basbaum, A.I., Anderson, D.J., 2009. Distinct subsets of unmyelinated primary sensory fibers mediate behavioural responses to noxious thermal and mechanical stimuli. Proc. Natl.
Acad. Sci. U.S.A. 106, 9075–9080.http://dx.doi.org/10.1073/pnas.0901507106.
Chen, W.T., Tung, C.H., Weissleder, R., 2004. Imaging reactive oxygen species in arthritis. Mol. Imag. 3, 159–162.
Christianson, C.A., Corr, M., Firestein, G.S., Mobargha, A., Yaksh, T.L., Svensson, C.I., 2010. Characterization of the acute and persistent pain state present in K/BxN serum transfer arthritis. Pain 151, 394–403.
Czikora, Á., Rutkai, I., Pásztor, E.T., Szalai, A., Pórszász, R., Boczán, J., Édes, I., Papp, Z., Tóth, A., 2013. Different desensitization patterns for sensory and vascular TRPV1 populations in the rat: expression, localization and functional consequences.
PLoS ONE 8, e78184.http://dx.doi.org/10.1371/journal.pone.0078184.
Danigo, A., Magy, L., Richard, L., Sturtz, F., Funalot, B., Demiot, C., 2014. A reversible functional sensory neuropathy model. Neurosci. Lett. 571, 39–44.
Denko, C.W., Malemud, C.J., 2004. The serum growth hormone to somatostatin ratio is skewed upward in rheumatoid arthritis patients. Front. Biosci. 9, 1660–1664.
Edwards, R.R., Wasan, A.D., Bingham 3rd, C.O., Bathon, J., Haythornthwaite, J.A., Smith, M.T., Page, G.G., 2009. Enhanced reactivity to pain in patients with rheumatoid arthritis. Arthritis Res. Ther. 11, R61.http://dx.doi.org/10.1186/
ar2684.
Elekes, K., Helyes, Z., Németh, J., Sándor, K., Pozsgai, G., Kereskai, L., Börzsei, R., Pintér, E., Szabó, A., Szolcsányi, J., 2007. Role of capsaicin-sensitive afferents and sensory neuropeptides in endotoxin-induced airway inflammation and consequent bronchial hyperreactivity in the mouse. Regul. Pept. 141, 44–54.
Fernandes, E.S., Russell, F.A., Spina, D., McDougall, J.J., Graepel, R., Gentry, C., Staniland, A.A., Mountford, D.M., Keeble, J.E., Malcangio, M., Bevan, S., Brain, S.D., 2011. A distinct role for transient receptor potential ankyrin 1, in addition to transient receptor potential vanilloid 1, in tumor necrosis factora-induced
inflammatory hyperalgesia and Freund’s complete adjuvant-induced monarthritis. Arthritis Rheum. 63, 819–829. http://dx.doi.org/10.1002/
art.30150.
Fioravanti, A., Govoni, M., La Montagna, G., Perpignano, G., Tirri, G., Trotta, F., Bogliolo, A., Ciocci, A., Mauceri, M.T., Marcolongo, R., 1995. Somatostatin 14 and joint inflammation: evidence for intraarticular efficacy of prolonged administration in rheumatoid arthritis. Drugs Exp. Clin. Res. 21, 97–103.
Fukushima, A., Boyle, D.L., Corr, M., Firestein, G.S., 2010. Kinetic analysis of synovial signalling and gene expression in animal models of arthritis. Ann. Rheum. Dis.
69, 918–923.
Gross, S., Gammon, S.T., Moss, B.L., Rauch, D., Harding, J., Heinecke, J.W., Ratner, L., Piwnica-Worms, D., 2009. Bioluminescence imaging of myeloperoxidase activity in vivo. Nat. Med. 15, 455–461.
Helyes, Z., Szabó, A., Németh, J., Jakab, B., Pintér, E., Bánvölgyi, A., Kereskai, L., Kéri, G., Szolcsányi, J., 2004. Antiinflammatory and analgesic effect of somatostatin released from capsaicin-sensitive sensory nerve terminals in Freund’s adjuvant- induced chronic arthritis model of the rat. Arthritis Rheum. 50, 1677–1685.
Hickman-Brecks, C.L., Racz, J.L., Meyer, D.M., LaBranche, T.P., Allen, P.M., 2011. Th17 cells can provide B cell help in autoantibody induced arthritis. J. Autoimmun.
36, 65–75.
Hulse, R.P., Beazley-Long, N., Hua, J., Kennedy, H., Prager, J., Bevan, H., Qiu, Y., Fernandes, E.S., Gammons, M.V., Ballmer-Hofer, K., Gittenberger de Groot, A.C., Churchill, A.J., Harper, S.J., Brain, S.D., Bates, D.O., Donaldson, L.F., 2014.
Regulation of alternative VEGF-A mRNA splicing is a therapeutic target for analgesia. Neurobiol. Dis. 71C, 245–259. http://dx.doi.org/10.1016/
j.nbd.2014.08.012.
Imhof, A.K., Glück, L., Gajda, M., Lupp, A., Bräuer, R., Schaible, H.G., Schulz, S., 2011.
Differential antiinflammatory and antinociceptive effects of the somatostatin analogs octreotide and pasireotide in a mouse model of immune-mediated arthritis. Arthritis Rheum. 63, 2352–2362.
Jones, G., Halbert, J., Crotty, M., Shanahan, E.M., Batterham, M., Ahern, M., 2003. The effect of treatment on radiological progression in rheumatoid arthritis: a systematic review of randomized placebo-controlled trials. Rheumatology (Oxford) 42, 6–13.
Kelly, S., Chapman, R.J., Woodhams, S., Sagar, D.R., Turner, J., Burston, J.J., Bullock, C., Paton, K., Huang, J., Wong, A., McWilliams, D.F., Okine, B.N., Barrett, D.A., Hathway, G.J., Walsh, D.A., Chapman, V., 2013. Increased function of pronociceptive TRPV1 at the level of the joint in a rat model of osteoarthritis pain. Ann. Rheum. Dis.http://dx.doi.org/10.1136/annrheumdis-2013-203413.
Konttinen, Y.T., Tiainen, V.M., Gomez-Barrena, E., Hukkanen, M., Salo, J., 2006.
Innervation of the joint and role of neuropeptides. Ann. N. Y. Acad. Sci. 1069, 149–154.
Korganow, A.S., Ji, H., Mangialaio, S., Duchatelle, V., Pelanda, R., Martin, T., Degott, C., Kikutani, H., Rajewsky, K., Pasquali, J.L., Benoist, C., Mathis, D., 1999. From systemic T cell self-reactivity to organ-specific autoimmune disease via immunoglobulins. Immunity 10, 451–461.
Kourilovitch, M., Galarza-Maldonado, C., Ortiz-Prado, E., 2014. Diagnosis and classification of rheumatoid arthritis. J. Autoimmun. 48–49, 26–30.
Kouskoff, V., Korganow, A.S., Duchatelle, V., Degott, C., Benoist, C., Mathis, D., 1996.
Organ-specific disease provoked by systemic autoimmunity. Cell 87, 811–822.
Kun, J., Helyes, Z., Perkecz, A., Bán, Á., Polgár, B., Szolcsányi, J., Pintér, E., 2012. Effect of surgical and chemical sensory denervation on non-neural expression of the transient receptor potential vanilloid 1 (TRPV1) receptors in the rat. J. Mol.
Neurosci. 48, 795–803.
Larsson, J., Ekblom, A., Henriksson, K., Lundeberg, T., Theodorsson, E., 1991.
Concentration of substance P, neurokinin A, calcitonin generelated peptide, neuropeptide Y and vasoactive intestinal polypeptide in synovial fluid from knee joints in patients suffering from rheumatoid arthritis. Scand. J. Rheumatol.
20, 326–335.
Leffler, A.S., Kosek, E., Lerndal, T., Nordmark, B., Hansson, P., 2002. Somatosensory perception and function of diffuse noxious inhibitory controls (DNIC) in patients suffering from rheumatoid arthritis. Eur. J. Pain 6, 161–176.
Levine, J.D., Khasar, S.G., Green, P.G., 2006. Neurogenic inflammation and arthritis.
Ann. N. Y. Acad. Sci. 1069, 155–167.
Maggi, C.A., 1995. Tachykinins and calcitonin gene-related peptide (CGRP) as cotransmitters released from peripheral endings of sensory nerves. Prog.
Neurobiol. 45, 1–98.
Meinel, T., Pongratz, G., Rauch, L., Straub, R.H., 2013. Neuronala1/2-adrenergic stimulation of IFN-c, IL-6, and CXCL-1 in murine spleen in late experimental arthritis. Brain Behav. Immun. 33, 80–89. http://dx.doi.org/10.1016/
j.bbi.2013.06.003.
Mishra, S.K., Hoon, M.A., 2010. Ablation of TrpV1 neurons reveals their selective role in thermal pain sensation. Mol. Cell. Neurosci. 43, 157–163.
Németh, J., Helyes, Z., Görcs, T., Gardi, J., Pintér, E., Szolcsányi, J., 1996. Development of somatostatin radioimmunoassay for the measurement of plasma and tissue contents of hormone. Acta Physiol. Hung. 84, 313–315.
Németh, T., Futosi, K., Hably, C., Brouns, M.R., Jakob, S.M., Kovács, M., Kertész, Z., Walzog, B., Settleman, J., Mócsai, A., 2010. Neutrophil functions and autoimmune arthritis in the absence of p190RhoGAP: generation and analysis of a novel null mutation in mice. J. Immunol. 185, 3064–3075.
Paran, D., Elkayam, O., Mayo, A., Paran, H., Amit, M., Yaron, M., Caspi, D., 2001. A pilot study of a long acting somatostatin analogue for the treatment of refractory rheumatoid arthritis. Ann. Rheum. Dis. 60, 888–891.
Pintér, E., Helyes, Z., Szolcsányi, J., 2006. Inhibitory effect of somatostatin on inflammation and nociception. Pharmacol. Ther. 112, 440–456.
Pintér, E., Pozsgai, G., Hajna, Z., Helyes, Z., Szolcsányi, J., 2014. Neuropeptide receptors as potential drug targets in the treatment of inflammatory conditions.
Br. J. Clin. Pharmacol. 77, 5–20.
Pongratz, G., Straub, R.H., 2010. The B cell, arthritis, and the sympathetic nervous system. Brain Behav. Immun. 24, 186–192. http://dx.doi.org/10.1016/
j.bbi.2009.07.002.
Riol-Blanco, L., Ordovas-Montanes, J., Perro, M., Naval, E., Thiriot, A., Alvarez, D., Paust, S., Wood, J.N., von Andrian, U.H., 2014. Nociceptive sensory neurons drive interleukin-23-mediated psoriasiform skin inflammation. Nature 510, 157–161.
Rossi, F., Bellini, G., Luongo, L., Torella, M., Mancusi, S., De Petrocellis, L., Petrosino, S., Siniscalco, D., Orlando, P., Scafuro, M., Colacurci, N., Perrotta, S., Nobili, B., Di Marzo, V., Maione, S., 2011. The endovanilloid/endocannabinoid system: a new potential target for osteoporosis therapy. Bone 48, 997–1007.http://dx.doi.org/
10.1016/j.bone.2011.01.001.
Schett, G., Gravallese, E., 2012. Bone erosion in rheumatoid arthritis: mechanisms, diagnosis and treatment. Nat. Rev. Rheumatol. 8, 656–664.
Sousa-Valente, J., Andreou, A.P., Urban, L., Nagy, I., 2014. Transient receptor potential ion channels in primary sensory neurons as targets for novel analgesics. Br. J. Pharmacol. 171, 2508–2527.
Stangenberg, L., Burzyn, D., Binstadt, B.A., Weissleder, R., Mahmood, U., Benoist, C., Mathis, D., 2014. Denervation protects limbs from inflammatory arthritis via an impact on the microvasculature. Proc. Natl. Acad. Sci. U.S.A. 111, 11419–11424.
Szabó, Á., Helyes, Z., Sándor, K., Bite, A., Pintér, E., Németh, J., Bánvölgyi, A., Bölcskei, K., Elekes, K., Szolcsányi, J., 2005. Role of TRPV1 receptors in adjuvant-induced chronic arthritis: in vivo study using gene-deficient mice. J. Pharmacol. Exp.
Ther. 314, 111–119.
Szolcsanyi, J., 1996. Capsaicin-sensitive sensory nerve terminals with local and systemic efferent functions: facts and scopes of an unorthodox neuroregulatory mechanism. Prog. Brain Res. 113, 343–359.
Szolcsanyi, J., Szallasi, A., Szallasi, Z., Joo, F., Blumberg, P.M., 1990. Resiniferatoxin:
an ultrapotent selective modulator of capsaicin-sensitive primary afferent neurons. J. Pharmacol. Exp. Ther. 255, 923–928.
Takeba, Y., Suzuki, N., Takeno, M., Asai, T., Tsuboi, S., Hoshino, T., Sakane, T., 1997.
Modulation of synovial cell function by somatostatin in patients with rheumatoid arthritis. Arthritis Rheum. 40, 2128–2138.
Tauber, S., Paulsen, K., Wolf, S., Synwoldt, P., Pahl, A., Schneider-Stock, R., Ullrich, O., 2012. Regulation of MMP-9 by a WIN-binding site in the monocyte-macrophage system independent from cannabinoid receptors. PLoS ONE 7, e48272.
Vanderschueren, D., Laurent, M.R., Claessens, F., Gielen, E., Lagerquist, M.K., Vandenput, L., Börjesson, A.E., Ohlsson, C., 2014. Sex steroid actions in male bone. Endocr. Rev. 9, er20141024.
Wang, W., Jian, Z., Guo, J., Ning, X., 2014. Increased levels of serum myeloperoxidase in patients with active rheumatoid arthritis. Life Sci. S0024–3205 (14), 00761–
769.http://dx.doi.org/10.1016/j.lfs.2014.09.012.
Yoo, S., Lim, J.Y., Hwang, S.W., 2014. Sensory TRP channel interactions with endogenous lipids and their biological outcomes. Molecules 19, 4708–4744.
Zhang, X., Lan, N., Bach, P., Nordstokke, D., Yu, W., Ellis, L., Meadows, G.G., Weinberg, J., 2012. Prenatal alcohol exposure alters the course and severity of adjuvant- induced arthritis in female rats. Brain Behav. Immun. 26, 439–450.http://
dx.doi.org/10.1016/j.bbi.2011.11.005.