Néhány szuperoxid dizmutáz és kataláz enzimmodell vizsgálata
DOKTORI (Ph.D.) ÉRTEKEZÉS
Készítette:
Kripli Balázs okleveles vegyész
Témavezető:
Dr. Speier Gábor egyetemi tanár
PANNON EGYETEM
KÉMIAI ÉS KÖRNYEZETTUDOMÁNYI DOKTORI ISKOLA
Veszprém
2011
Témavezető: Dr. Speier Gábor
Elfogadásra javaslom (igen/nem)
...
(aláírás) A jelölt a doktori szigorlaton ... %-ot ért el.
Veszprém, ...
...
Szigorlati Bizottság elnöke Az értekezést bírálóként elfogadásra javaslom:
Bíráló neve: ... igen/nem
...
(aláírás) Bíráló neve: ... igen/nem
...
(aláírás) A jelölt az értekezés nyilvános vitáján ... %-ot ért el.
Veszprém, ...
...
Bíráló Bizottság elnöke A doktori (PhD) oklevél minősítése: ...
...
EDT elnöke
Néhány szuperoxid dizmutáz és kataláz enzimmodell vizsgálata
Készítette: Kripli Balázs
Mérnöki Kar, Kémia Intézet, Szerves Kémia Intézeti Tanszék
Témavezető: Dr. Speier Gábor
A természetes szuperoxid dizmutáz (SOD)-enzimek hiányos működését, ennek következ- tében a szuperoxid gyök-anion kumulálódását, számos betegség (pl. AIDS, rák, gyulladásos betegségek stb.) egyik kiváltó okaként tartják számon. Ennek eredményeképpen az utóbbi évtizedben jelentős érdeklődés irányult mesterséges helyettesítők előállítására és vizsgálatára.
A természetes SOD-enzimek mintájára Fe, Mn, Cu/Zn és Ni átmenetifém-tartalmú modelleket egyaránt előállítottak és teszteltek. Az eredmények alapján úgy tűnik, hogy a SOD-katalízis során a fém redoxi átmenete fontos szerepet játszik. Az ismeretek alapján azok a vegyületek mutatnak SOD utánzó aktivitást, melyeknek redoxi potenciálja a O2/O2·–
és O2·–
/H2O2 átmeneteknek megfelelő -0,16 V < E1/2 (vs. NHE) < +0,89 V határértékek közé esik pH
= 7 esetén (1a).
O2 (1a)
2 + 2 H+ H2O2+ O2 [SOD]
Az alkalmazott izoindolin-tartalmú ligandumokkal (HL1-HL7) előállított Mn(II)- Fe(II)- Ni(II)- és Cu(II)-komplexeket spektroszkópiai módszerekkel jellemeztük. A SOD-utánzó méréseket kompetitív kinetikai módszerrel NBT (nitroblue tetrazolium) és citokróm c(III) reagens jelenlétében is elvégeztük. Az élő szervezetekben felhalmozódó hidrogén-peroxid bomlási folyamatai a sejtfalra nézve káros, reaktív oxigén származékokhoz (szuperoxid, hidroxilgyök stb.) vezethetnek. Ennek kiküszöbölésére az élő szervezetekben ún. kataláz enzimek szolgálnak, amelyek a hidrogén-peroxid vízzé és dioxigénné történő reakcióját katalizálják (1b).
2H2O2 kataláz 2H2O + O2 (1b)
Kataláz modellként előállítottuk a [MnII(HL1)]Cl2 összetételű komplexet. Szerkezetét spektroszkópiai (IR, UV-Vis, ESR) mérésekkel, illetve röntgendiffrakciós méréssel igazoltuk. Vizsgáltuk az előállított komplex kataláz aktivitását és megállapítottuk, hogy az előállított komplex katalizálja a H2O2 vízzé, illetve dioxigénné való dizmutációját, tehát
Malfunction of superoxide dismutase (SOD) enzymes may lead to cumulation of superoxide radical anion that is held as the reason for several diseases (AIDS, cancer, inflammations). This commonality provides the opportunity to control the diseases by using synthetic SOD mimics that can suppress the superoxide concentration to a safe level. Artificial scavengers with various metal ions (Mn, Fe, Ni, Cu) have been prepared and tested. Based on the current knowledge, those transition metal complexes show SOD scavenger activity that have a metal-associated redox potential between the redox potentials of the two steps of the spontaneous superoxide dismutation process: the O2/O2·–
at -0.16 V and the O2·–/H2O2 at +0.89 V vs. NHE at pH = 7 value (1a).
O2 (1a)
2 + 2 H+ H2O2+ O2 [SOD]
With the syntetized isoindoline-based ligands (HL1-HL7) the Fe(II),- Mn(II),- Ni(II)-, Co(II)- and Cu(II)-complexes have been prepared and characterized by various spectroscopic methods. SOD-like activity tests have been made with competitive kinetic methods in aqueous HEPES buffer using NBT (nitroblue tetrazolium) or cytochrome c(III) reagents according to the standard McCord–Fridovich method. Catalases are enzymes that protect cells from deleterious effects of hydrogen peroxide, a by product of respiration, by disproportionating H2O2 into water and dioxygen (1b).
2H2O2 Catalase 2H2O + O2 (1b)
As a catalase model compound we prepared the [MnII(HL1)]Cl2 complex, which has been fully characterized by (IR, UV-Vis, ESR and X-ray diffraction analyses). The complex is an active catalyst in the dismutation of H2O2 to water and dioxygen therefore we are can consider the eximaned reaction to be the functional model of catalase.
Untersuchungen über Superoxide Dismutase und Katalase Models Von: Balázs Kripli
Fakultät von Ingenierwesen, Institute für Chemie
Doktorvater: Dr. Gábor Speier
Die fehlhafte Wirksamkeit der Superoxid Dismutase Enzyme erhöht die Konzentration der Superoxide wird erhöht und nach allgemeinen Meinungen ist das die Ursache für verschieden Krankheiten wie z.B. AIDS, Krebs und Entzündungen. Aus diesem Grund wurden Fe(II)-, Mn(II)- und Ni(II)-haltige Modelle als künstliche Enzyme hergestellt und getestet. Diese Untersuchungen ergaben, dass die Redox-Eigenschaften von den entsprechenden Metallen eine wichtige Rolle spielt. Es scheint so, dass diese SOD-Modelle zeigen gute Aktivität, die die Halbpotentiale zwischen den von O2/O2·–
und O2·–
/H2O2 fallen -0,16 V < E1/2 (vs. NHE) <
+0,89 V bei pH = 7 (1a).
O2 (1a)
2 + 2 H+ H2O2+ O2 [SOD]
Wir haben verschiedene isoindolinhaltige Fe(II)-, Mn(II)-, Co(II)-, Ni(II)- und Cu(II)-Komp- lexe mit den Liganden (HL1-HL7) hergestellt und ihre Struktur spektroskopisch festgestellt.
Die SOD-Aktivität Messungen wurden mit der Hilfe Von kompetitiven kinetischen Methoden in der Anwesenheit von NBT (Nitroblau tetrazolium) und citokrom c(III) Reagent ausgeführt.
2H2O2 Katalase 2H2O + O2 (1b)
Die Ansammlung von Hydrogen-Peroxide in den lebenden Organizmen führt infolge seiner Zersetzung zu reaktiven Derivaten (Superoxide, Hydroxyl-Radikal, usw.) die die Zellwand beschädigen. Um das zu verhindern wird Hydrogen-Peroxide durch den Enzym Katalase zu Wasser und Sauerstoff umgewandelt (1b). [MnII(HL1)]Cl2 wurde hergestellt, seine Struktur bestimmt und seine Katalase-Aktivität untersucht. Es hat sich ausgestellt, dass der Komplex H2O2 zersetzt und dabei entstehen Wasser und Sauerstoff.
1.6. Kobalttartalmú szuperoxid dizmutáz enzimek 12
1.7. Nikkeltartalmú szuperoxid dizmutáz enzimek 13
1.8. Réztartalmú szuperoxid dizmutáz enzimek 14
1.9. SOD-utánzó vegyületek 16
1.9.1. Szuperoxid gyök-anion reakciója SOD-utánzó vegyületekkel NBT reagens
jelenlétében 19
1.9.2. Szuperoxid gyök-anion reakciója SOD-utánzó vegyületekkel citokróm c(III)
reagens jelenlétében 22
2.0. Kataláz enzimek 24
2.1. Mangántartalmú kataláz enzimek 25
2.2. Ligandumszintézis 28
2. Célkitűzések 30
3. Eredmények és értékelésük 31
3.1. A ligandumok szintézise és szerkezetük azonosítása 31 3.2. MnII(Ln)2 összetételű SOD-utánzó modellvegyületek előállítása és jellemzése 36 3.3. FeII(Ln)2 összetételű SOD-utánzó modellvegyületek előállítása és jellemzése 39 3.4. CoII(Ln)2 összetételű SOD-utánzó modellvegyületek előállítása és jellemzése 44 3.5. NiII(Ln)2 összetételű SOD-utánzó modellvegyületek előállítása és jellemzése 47 3.6. CuII(Ln)2 összetételű SOD-utánzó modellvegyületek előállítása és jellemzése 51 3.7. CuII(HLn)Cl2 és CuII(Ln)Cl SOD-utánzó modellvegyületek előállítása és jellemzése 56 3.8. MnII(Ln)2 modellvegyületek SOD-utánzó aktivitása és redoxi tulajdonságainak vizsgálata 62 3.9. FeII(Ln)2 modellvegyületek SOD-utánzó aktivitása és redoxi tulajdonságainak vizsgálata 68 4.0. CoII(Ln)2 modellvegyületek SOD-utánzó aktivitása és redoxi tulajdonságainak
vizsgálata 72
4.1. NiII(Ln)2 modellvegyületek SOD-utánzó aktivitása és redoxi tulajdonságainak
vizsgálata 75
4.2. CuII(Ln)2 modellvegyületek SOD-utánzó aktivitása és redoxi tulajdonságainak
vizsgálata 76
tulajdonságainak vizsgálata 78
4.4. Kataláz modellek 79
4.5. A [MnII(HL1)]Cl2 összetételű modellvegyület előállítása és jellemzése 80 4.6. A [MnII(HL1)]Cl2 modellvegyület kataláz aktivitása 82
5. Összefoglalás 87
6. Kísérleti rész 89
7. Irodalomjegyzék 102
n-BuOH normál butil-alkohol
FT-IR Fourier-transzformációs infravörös spektroszkópia
KBr kálium-bromid
UV-Vis ultraibolya-látható spektroszkópia
ANu nukleofil addíció
Vk kezdeti sebesség
DMSO dimetil-szulfoxid
NBT 2,2’-bisz(4-nitrofenil)-5,5’-difenil-3,3’-(3,3’-dimetoxi-4,4’- difenilén) ditetrazolium klorid (nitroblue tetrazolium) ε moláris abszorpciós együttható (M-1cm-1)
HL1 1,3-bisz(2’-benzimidazolil-imino)-izoindolin
HL2 1,3-bisz(N-metil-2’-benzimidazolil-imino)-izoindolin HL3 1,3-bisz(2’-tiazolil-imino)-izoindolin
HL4 1,3-bisz (2’-piridil-imino)-izoindolin
HL5 1,3-bisz(3’-metil-2’-piridil-imino)-izoindolin HL6 1,3-bisz(4’-metil-2’-piridil-imino)-izoindolin HL7 1,3-bisz(2’-benztiazolil-imino)-izoindolin
NMR mágneses magrezonancia spektroszkópia
Cis cisztein
Glu glutaminsav
Asp aszparaginsav
Arg arginin
His hisztidin
Tyr tirozin
Trp triptofán
Gln glutamin
Lys lizin
Ser szerin
ESR elektron spin rezonancia
EXAFS finomszerkezetű abszorpciós élközeli spektroszkópia
Et3N trietil-amin
n-Bu4NClO4 n-tetra-butil-ammónium perklorát
NHE normál hidrogén elektród
SCE telített kalomel elektród
vizsgálatok felvételeit és a Radiokémia Tanszéknek (Pannon Egyetem) a ciklikus voltametria mérések lehetőségét..
Továbbá a Szerves Kémia Intézeti Tanszék minden munkatársának bármiféle segítségét, amivel munkámat támogatták.
Mindennél jobban köszönöm családom végtelen bíztatását és bizalmát.
Veszprém, 2011. október 15.
Kripli Balázs
Bevezetés
Az élő szervezetekben lezajló biokémiai folyamatok többsége összetett, gyors reakciók sorozata. A reakciók sebességét és specifikusságát az enzimek alakítják. Az élő szervezetek harmonikusan működő összetett rendszerek. A nem megfelelő működésük során kialakuló zavarok betegségeket eredményezhetnek, és ez sok esetben éppen a biokatalizátorok hiányos, vagy túlzott működésének következménye. Felépítésüket te- kintve az enzimek nagy része a fehérjéken kívül fémet is tartalmaz, ezeket nevezik metalloenzimeknek. Számos esetben a fémek is részt vesznek az aktív centrum kialakításában, ami teljes mértékben meghatározza az enzim specifikusságát. Működési mechanizmusuk részleteinek tisztázásához azonban összetett – nem csupán egy tudo- mányágat magába foglaló – vizsgálatsorozatra van szükség.
Az analitikai technikák, mint például a röntgendiffrakció, ICP, AAS fejlődése, valamint az elektronmikroszkópok felbontásának javulása több lehetőséget biztosít számunkra az anyagok pontos szerkezetének megismerésében. A biológiai ismeretek fontossága a testben lezajló enzimreakciókról és azok körülményeiről, lényeges mindenféle gyógyászattal kapcsolatos tevékenységgel, kutatással kapcsolatban. Az egyes folyamatok pontos megértéséhez azonban többre van szükség. Így alakulhatott ki évtizedekkel ezelőtt egy új, interdiszciplináris tudományterület, a bioszervetlen kémia.
Az enzimek nagy méretük és összetettségük révén nehezen kezelhető vegyületek.
Tisztításuk, izolálásuk nem kevésbé körülményes munka. Szerkezetüket vagy funkciójukat alapul véve ún. bioutánzó vegyületeket hozhatunk létre, amelyek fel- építésükben sokkal egyszerűbbek a megfelelő enzimnél. Enzimutánzó tulajdonságuk abból fakad, hogy azonos folyamatokat katalizálnak, többnyire nagyságrendekkel kisebb sebességgel. Mivel az elsődleges cél a folyamat lépésről-lépésre történő leírása, így az utóbbi jellemző kulcsfontosságú, hiszen a lassú reakciók jobban nyomon követhetőek. A modellvegyületek reakcióinak pontos feltérképezésével tehát, egy lépéssel közelebb kerülhetünk a valós enzimmechanizmus megértéséhez.
Az enzimek egyik nagy és jelentős csoportját képezik az oxidoreduktázok, amelyek reakcióiban elektron, vagy oxigénatom transzfer történik a kölcsönhatásba lépő molekulák között. Ebbe a csoportba tartoznak az értekezés témáját képező szuperoxid- dizmutázok és katalázok is. Ezek bemutatása, valamint funkciójuknak pontosabb megismerése modellvegyületeik leírásán, jellemzésén keresztül valósítható meg.
metabolizmusnak, vagy anyagcserének nevezzük (1. ábra).
Energia
TERMÉK TÁPANYAG Metabolizmus
lebontás f elépítés
Enzimatikus reakciók SZUBSZTRÁTUM
1. ábra Enzimreakciók szerepe az élő szervezetekben.
Enyhe körülmények között az egyes kémiai átalakulások nem, vagy csak csekély mértékben játszódnak le, mivel a reakciók aktiválási energiája általában nagy. Ezzel szemben a tapasztalatok azt mutatják, hogy a metabolizmus részfolyamatai élő szervezetekben gyorsan mennek végbe, ami nagyszámú enzim jelenlétének köszönhető.
Ez alapján az enzimek olyan biokatalizátoroknak tekinthetők, amelyek a reakciók aktiválási energiáját lecsökkentve, lehetővé teszik azok gyors lejátszódását a megfelelő biológiai környezetben. Az enzimek aminosavakból épülnek fel, tehát a fehérjék családjába tartoznak. Molekulatömegük 12000-50000 Dalton között lehet [1].
Amennyiben csak aminosavak alkotják, egyszerű fehérjékről beszélünk (proteinek, apoenzimek), ha nem fehérje természetű részt is tartalmaznak (prosztétikus csoport, koenzim), akkor összetett fehérjékről (proteidek, holoenzimek) van szó. Egy, vagy több speciális, ún. aktív helyet tartalmaznak, melyek az enzimfunkcióért felelősek, itt játszódik
le egy adott reakció katalízise. Jellemző rájuk továbbá, hogy csak egy adott típusú reakciót gyorsítanak [2].
A katalizált reakciók szempontjából 6 fő csoportot különböztetünk meg:
I. Hidrolázok: Hidrolízis reakciókért felelős fehérjék, peptidek kötésének, poliszacharidok glikozidkötésének, zsírok, foszfátok észterkötésének hasításáért felelnek.
II. Oxidoreduktázok: Redoxireakciók lejátszódásáért felelős fémtartalmú enzimek, működésük során elektronok vagy oxigénatom kerül át egyik molekuláról a másikra.
III. Transzferázok: Meghatározott atomcsoport átvitelét végzik egyik molekuláról a másikra, pl: -NH2, -CO csoport.
IV. Izomerázok: Különböző átrendeződéses reakciók katalízisét elősegítő fehérjék.
V. Liázok: A szubsztrátum adott csoportját távolítják el eliminációs reakciók során.
VI. Ligázok: Két molekula összekapcsolódásáért felelnek ezek alapján lehetnek: C-C, C-N és C-O kötést katalizáló ligázok.
Az enzimek az átalakítandó vegyületre nézve is szelektívek. Az adott enzimre nézve változást szenvedő vegyületet szubsztrátumnak nevezzük. A szelektivitást az enzimek (E) aktív helyének sztérikus és elektronikus sajátságai biztosítják. Ezen a helyen kötődik meg és aktiválódik a szubsztrátum (S). A reakció lejátszódását követően a termékek (P) távozik az aktív helyről (2. ábra). Az eddig ismert enzimek közel egyharmada fémiono- kat tartalmaz, amelyek többféle módon kötődhetnek a fehérjéhez, és többféle funkciót is elláthatnak. Ezeket két csoportra osztjuk: metalloenzimekre és fémionok által aktivált enzimekre. A metalloenzimekben a fémion az enzimmolekulába beépült alkotórész, a fémion és a fehérje sztöchiometrikus aránya meghatározott érték. Amennyiben a fémiont kiszakítjuk a metalloenzimből, az enzim elveszíti aktivitását. A fémionok által aktivált enzimek esetében egyensúly áll fenn a fémion és az enzim, továbbá a fémion aktiválta enzim között. Ennél az enzimcsoportnál a fémet egyszerű kémiai módszerekkel el lehet választani a fehérjétől anélkül, hogy aktivítását teljesen elveszítené.
2. ábra Az enzimkatalízis körfolyamata.
1.2. Enzimmodellek
A legtöbb enzim nehezen hozzáférhető és kevés vizsgálatra nyújt lehetőséget, mivel tiszta formában való elkülönítésük nehéz feladat. Pontos hatásmechanizmusuk felderítése részletes kinetikai vizsgálatokat igényelne, amelyhez több és pontosabb mérésre lenne szükség. Az enzimek vizsgálata során a szerkezet és a működés a két legfontosabb paraméter, amelyek segítik a megismerésüket. Ezek szorosan összefüggő sajátságok, hiszen az enzim funkcióját elsősorban az aktív centrum szerkezete, elektronikus és sztérikus viszonyai befolyásolják. A szerkezet és a funkció megismerésének érdekében kettős célú enzimmodellek (3. ábra) megalkotása szükséges, melyekkel az enzimvizsgálatok nehézségei kiküszöbölhetők.
Szerkezet
Metalloenzim Fémkomplex
Funkció
Szerkezeti modellek
Funkcionális modellek
Stabilis köztitermék
Instabilis köztitermék
Reakció mechanizmus
Katalitikus reakció
3. ábra Enzimmodellek és szerepük.
A szerkezeti modellek:
A legfontosabb elvárás velük szemben az, hogy geometriai és elektronikus tulajdonságaikban minél jobban hasonlítsanak az enzim aktív helyéhez. Metalloenzimek esetében, a nagyméretű fehérjemolekulát szerves ligandumokkal helyettesítik, ebből kifolyólag standard körülmények között spektroszkópiailag vizsgálhatók. A kialakuló komplex általában túl stabilis, hogy az enzimatikushoz hasonló reakciót katalizálja.
A funkcionális modellek:
Esetükben a leglényegesebb, hogy minél nagyobb szelektivitás mellett képezzék az enzimreakció termékeit, tehát az enzim funkcióját utánozzák. Ez lehetséges úgy, hogy csak a modellkomplexet visszük reakcióba, máskor a modellt katalizátorként alkalmazzuk a szubsztrátum megfelelő reakciójában. Ezek után a mechanizmus felderíthető kinetikai vizsgálatok alkalmazásával. A SOD-, és kataláz-utánzó modellek esetében a funkcionális modellek előállítása volt a cél.
1.3. Szuperoxid dizmutáz enzimek
Az anyagcsere során az élő szervezetben reaktív oxigén származékok (ROS) képződnek, ezek a következők: hidroxil gyök (OH•), szuperoxid gyök-anion (O2•-), nitrogén-monoxid (NO•) és peroxil (RO2•) gyökök. A peroxinitrit (ONOO-), a hipoklórossav (HOCl), a hidrogén-peroxid (H2O2), valamint a szingulett dioxigén (1O2) és az ózon (O3) nem szabad gyökök, de könnyen szabad gyökös reakcióhoz vezetnek a szervezetben [3]. Az oxidatív stressz az egyensúly megbomlását jelenti a reaktív oxigén vegyületek és az antioxidánsok hatása között. Ez a folyamat a sejtek károsodását és pusztulását okozhatja, ami betegségekhez vezet [4]. A szuperoxid gyök-anion a dioxigén molekula egyelektronos redukált formája viszonylag szelektív reaktivitással. Enzimrendszerek okozzák létrejöttét autooxidációs reakciókban, azonban nem enzimatikus elektron transzferrel is kialakulhat, miközben a molekuláris oxigén redukálódik. Vizes oldatokban, a szuperoxid gyök-anion oxidálhatja az aszkorbinsavat, továbbá képes redukálni vaskomplexeket, mint a citokróm c(III)-t és a Fe3+ EDTA-t [5]. Oxidációval, redukcióval és diszproporcióval azonban semlegesíthető. A szuperoxid dizmutáz (SOD) a szuperoxid gyök-anion vízzé és hidrogén-peroxiddá való átalakulását katalizálja (1-3) [6]. A szuperoxid gyök-anion önmagában is képes diszproporcionálódásra, ennek sebessége (k ~
4 -1 -1
M(n+1)++ 2 H++ O2 M(n+1)+ H2O2 (2) O2
2 + 2 H+ H2O2+ O2 (3)
Elsőként McCord és Fridovich számolt be a SOD enzimek enzimatikus aktivitásának felfedezéséről 1968-ban [6,8]. Mann és Keilin [9] 30 évvel korábban már izoláltak egy proteint szarvasmarhák véréből és májából mint ismeretlen funkcióval bíró réztartalmú proteint. A protein több nevet is kapott: eritrokuprein, hepatokuprein, citokuprein. A szuperoxidot, a SOD enzimek szubsztrátumát Linus Pauling fedezte fel az 1930-as években [10]. Pauling nem tudta, hogy a gyök biológiailag is képes keletkezni, és
hogy számos betegség okozója. Knowles kimutatta 1969-ben [11], hogy a xantin oxidáz nevű enzim képes szuperoxidot produkálni. McCord és Fridovich pedig bebizonyította, hogy a Mann és Keilin által izolált réz-protein képes katalitikusan megszüntetni Pauling szabad gyökét. Huber [12] az 1960-as években izolálta ugyanezt a proteint szarvasmarhák májából. Ezt a proteint Orgoteinnek nevezték el. Hogy hogyan kapcsolódik ez a felfedezés a SOD aktivitáshoz, az már Bernard Babior [13] felfedezése volt 1973-ban, aki megállapította, hogy a fagocitáló neutrofilek nagy mennyiségű szuperoxid gyököt produkálnak, amit ő bakteriális folyamatnak tartott. Később kiderült, hogy a szuperoxid gyulladásos betegséget okozhat. A további kutatások pedig elvezettek ahhoz a felismeréshez, hogy számos betegség kialakulása is ezzel a gyökkel hozható összefüggésbe (iszkémia, reperfúzió, cukorbetegség, metasztázis, angiogenézis, Parkinson-kór és a rák). Ez onnan eredeztethető, hogy a gyök túltermelődése lipid per- oxilációhoz, protein oxidációhoz és a DNS károsodásához vezethet [14,15]. A megelőzésben kulcsfontosságú szerep jut a SOD enzimeknek. A szuperoxid dizmutáz enzimek gyógyszerként való alkalmazása során felmerült a kérdés, vajon hatásuk pozitív vagy negatív-e a szervezetre nézve. A TBARS-kísérletek azt bizonyították (TBARS = Thiobarbituric Acid Reactive Substances; Tiobarbitursav reaktív szubsztanciák. Ezek
olyan kísérletileg meghatározott értékek, amelyek az oxidatív stressz folyamatát jellemzik. Az orvosi diagnosztikában gyakran alkalmazott kísérletek, például a lipid peroxiláció vizsgálata során is. Alkalmazásuk egy pontig nagyon hatékony, ezen a ponton túl azonban súlyosbítja a betegséget a növekvő lipid peroxilációval együtt [16]. Az optimális SOD koncentráció és a káros koncentráció közti különbség hatszoros (4. ábra).
A szuperoxid gyök kezdeményezni és megszüntetni is képes a lipid peroxilációt. A maximális gyógyulás akkor érhető el, ha a lipid peroxiláció minimális.
4. ábra A SOD enzim működésének optimuma a lipid peroxiláció során [16].
A SOD enzimek többfajta szerkezetét különböztették meg a fém kofaktortól függően. A réz- és cinktartalmú SOD enzimek (Cu/Zn-SOD) általánosságban az eukarióta sejtek citoplazmájában és néhány növény kloroplasztiszában találhatók meg, de az emberi szervezetben is számos képviselőjük ismert, mint a későbbiekben ismertetendő Cu/Zn- tartalmú analóg is. A nikkeltartalmú SOD enzimeket (Ni-SOD) néhány baktériumban találták meg először. A vas és mangántartalmú SOD enzimek (Fe-, Mn-SOD) pedig gyakoriak a prokariótákban és az eukarióta sejtek mitokondriumában, azaz az alacsonyabb szerveződésű élőlények szervezetében. Az utóbbi osztály vas- vagy mangán- iont tartalmaz fém kofaktorként és nagyon hasonló szekvenciával és szerkezettel rendelkeznek, míg a réz/cink- és a nikkeltartalmú enzimek szerkezete ettől jelentősen eltér, ezért ezeket a következő fejezetekben külön tárgyaljuk.
% Funkció helyreállítása O
TBARS (nmol/mg protein) ■
[SOD] (mg/l)
5. ábra A tetramer szerkezetű Mn-SOD enzim (balra) és a mangán ionok elhelyezkedése a dimer szerkezetű szuperoxid dizmutázban (jobbra).
A Mn-SOD szerkezetét tekintve alegységekből épül fel, minden alegységet két domén képez: egy túlnyomórészt α-helikális, N-terminális domén és egy kevert α/β-szerkezeteket tartalmazó C-terminális domén. A fémet megkötő hely a két domén érintkezési helyén található a fehérje belső részében, valamint az N-terminális (His26, His81) és a C- terminális (Asp167, His171) régiók ligandumaiból jön létre. A homodimer szerkezet tengelyes szimmetriával rendelkezik és egy kiterjedt hidrofób felülettel stabilizálódik az alegységek érintkezésénél. Az alegységek közti kapcsolat 2 csoporton keresztül jön létre (Glu170, Tyr174), melyek egy ‘dupla híd’ alakú motívumot képeznek hidrogén kötések révén a komplementer alegységgel. A homotetramer szerkezet esetében az alegységek közti kölcsönhatások sokkal bonyolultabbak [5,6].
6. ábra Az Escherichia coli-ban található Mn-SOD aktív helyének szerkezete [7].
A fémkötő helyet tekintve jól látható, hogy a fémion 4 aminosavhoz és egy oldószer molekulához (H2O vagy HO- a fémion oxidációs állapotától függően) koordinálódik létrehozva egy torzult trigonális bipiramisos szerkezetet (6. ábra). Az aktív hely tulaj- donságait döntően a belső koordinációs szféra határozza meg [6,8], de a külső koor- dinációs szférának is nagy szerepe van a komplex felépítésében és katalitikus funkciójában. A koordinált oldószer molekula hidrogénkötést létesít egy külsőszférás csoporttal (Gln146 az E. coli Mn-SOD-ban), így megváltoztatja a fémcentrum reak- tivitását. Ez a csoport a legfontosabb szerkezeti determináns a fém specificitás szempontjából [9-11]. Azokban az enzimekben, ahol a mangán felelős az aktivitásért, ez a csoport a C-terminális doménből ered [12,13]. A szuperoxid egy elektrosztatikus tölcséren keresztül éri el az aktív centrumot, melynek szűk bejárata van, csupán a kis méretű ionok számára biztosítva a bejutást [18]. A működésük mechanizmusát a következő (4-7) egyenletekben tüntettem fel, ahol a SOD a fehérjerészt reprezentálja [19]:
Mn2+(H2O)SOD + O2
O2 (4)
Mn3+(OH-)SOD + + H+ k1 Mn2+(H2O)SOD+O2 + H+ k2
Mn3+(OH-)SOD + H2O2 (5) Mn2+(H2O)SOD+O2 k3
Mn3+(O22-)SOD + H2O (6) Mn3+(O22-)SOD + H++ H2O k4
Mn3+(OH-)SOD + H2O2 (7)
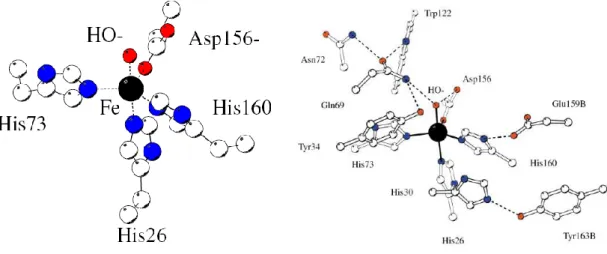
hasonlóan a Mn-SOD-okhoz dimer és tetramer szerkezetben is előfordulhatnak, mind az alacsonyabb, mind pedig a magasabb sejtes szerveződésű élőlényekben. Az Escherichia coli-ban található dimer Fe-SOD monomer egységei 22 kDa molekulatömegűek, amelyeknek dimer és monomer háromdimenziós röntgendiffrakciós felvételeit a 7. ábrán tüntettem fel [20, 21, 22-24, 26-31]. A monomer részeket két domén alkotja, mindegyikhez kapcsolódik még két ligandum és egy aktív centrum, amely a fémiont tartalmazza. Az N-terminális domén két hosszú hélixre különül el, egy rövidebbre és egy könnyebben variálhatóra, mint a másik rész (1-80 terjedő részek). Az első hélix tartalmaz egy hurkot amely behatárolja a Tyr34, His30 és a His26 aktív centrum körüli orientációját. A His73 ligandum a második hosszú hélixből kapcsolódik az első doménhez [27]. A linker megközelítőleg 10 módosítható aminosavból áll a két doménben és a C-terminális egységben (90-190 részek), továbbá α/β-szerkezeteket tartalmaz 3-3 β- lemezes és 4-4 α-hélixet mindkét oldalon. Az Asp156 és a His160 a C-terminális domén ligandumai, amelyek egy hurokkal kapcsolódnak az aktív centrumhoz. Az ötödik ligandum az oldószer molekula, amely oxidációs állapottól függően OH- (Fe3+) vagy H2O (Fe2+ esetén) [31]. A domének közötti térrész túlnyomóan hidrofób [27]. Az alegységek közötti rész nagyon rögzített és szimmetriafüggő elektrosztatikus csatornák gondoskodnak a szubsztrátum aktív centrumhoz jutásához. A nagyobb méretű oldószermolekulák hozzáférhetetlenek az aktív hely számára, ugyanis a kapcsolódó külső aminosav-egységek, mint Trp77, Tyr34 és His30 megakadályozzák ezt (8. ábra). Ezek az egységek a másodlagos koordinációs szférát képezik a monomer egységben. Tehát ezen a tölcséren keresztül tudnak csak a szubsztrátum molekulák (amelyek méretüknél fogva képesek odajutni) az aktív helyhez érkezni [27]. Most az iménti szerkezet működésének leírásával lehetőségünk nyílott a metalloenzimek általános bemutatására is. A kapcsolódó domének, alegységek, és a köztük lévő térrész, továbbá az őket összetartó erők, mind az enzimekre jellemző igen nagy szelektivítást és nagy katalitikus hatást szolgálják. A fémionok körüli geometriák hozzávetőleg trigonális bipiramisos szerkezettel jelle-
mezhetőek, ahol axiális pozícióban a His26 aminosav és a koordinált oldószermolekula találhatók. Ekvatoriális pozícióban pedig a His73, His160 és Asp156 aminosav egységek kapcsolódnak. A koordinált oldószermolekula biztosítja a tartós kiterjedt kapcsolatot hidrogénkötések révén a Gln69, Tyr34, Trp122, Asn72, és Asp156 továbbá a His30 és Tyr163B aminosav-részletek között [27,32]. Ez az aktív helyhez közeli térrész túlnyomóan aromás szerkezetű aminosavakból áll. Ezek elegendő védelmet nyújtanak a relatíve hosszú életű szabad gyökök ellen az aktív centrum számára.
7. ábra Az Escherichia coli-ban található dimer Fe-SOD háromdimenziós szerkezete és egyik monomer egységének szerkezete.
8. ábra Az Escherichia coli -ban található dimer Fe-SOD aktív centrumának és másodlagos koordinációs szférájának szerkezete.
léteznek. A Cu/Zn-SOD-okban a cinket helyettesítve fordulhat elő, mint azt a 10. ábra is szemlélteti [33]. Az aktív centrumról a 10. ábra alapján elmondható, hogy azonos koor- dinációs szférával jellemezhető, mint a következő fejezetben részletesebben tárgyalandó réz-cink szuperoxid dizmutázok. Vizsgálataink során arra kerestük a választ, hogy a szintetikusan előállított Co(II)-komplexeink, mint enzimmodellek, rendelkeznek-e önál- lóan, réz(II)-ion jelenléte nélkül is SOD-utánzó aktivitással, ami azért lényeges, mert később látni fogjuk, hogy a réz-centrum lesz a felelős a katalitikus funkciók ellátásáért a Cu/Zn-SOD enzimben. A kobalt-tartalmú modellvegyületeink vizsgálatával kívántunk hozzájárulni a szuperoxid dizmutázok már amúgy sem csekély kutatási területének bővítéséhez. A működésük bővebb kifejtése a réz-tartalmú szuperoxid dizmutázok fejezetben történik majd, amely azonos az ebbe a csoportba tartozó vegyületekével.
10. ábra A Cu/Co-SOD-ok aktív centruma és működése [33].
1.7. Nikkeltartalmú szuperoxid dizmutáz enzimek
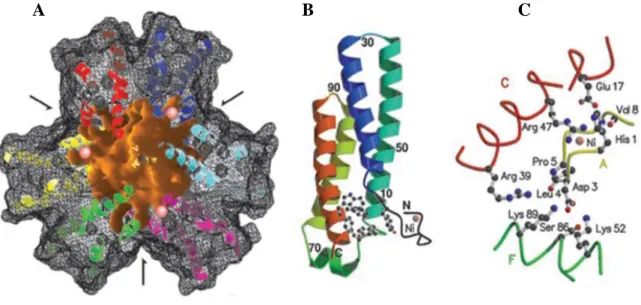
A mangán- és vastartalmú analógokon keresztül már részletesen bemutattam a SOD- enzimek működését és főbb jellemzőit. Történt ez azon megfontolás alapján, mert ez a két csoport a legtöbbet tanulmányozott a szuperoxid dizmutázok közül. Itt is egy baktériumból a Streptomyces seoulensis-ból elkülönített példán keresztül mutatnám be a nikkeltartalmú SOD-enzimek szerkezetét és fontosabb tulajdonságaikat (11. ábra) [34]. A működési mechanizmusukat a (10-11) egyenletekkel tüntettem fel. Ennek a csoportnak a tagjait csak nemrégiben sikerült azonosítani.
11. ábra A Streptomyces seoulensis baktériumból elkülönített hexamer szerkezetű, négy α-hélix alegységgel rendelkező Ni-SOD röntgendiffrakciós felvétele és aktív centruma.
Az oldószer számára megközelíthető Ni-SOD felületét látjuk térben a három szimmetriatengely feltüntetésével. A protein külső részét feketével tüntettük fel, a belső szférát naranccsal, az egyes alegységeket különböző színekkel, a Ni(II)-ionok pedig rózsazsínnel láthatóak az (A) ábrán. A nyilak a 3 db digir szimmetriatengelyt mutatják és a csatornák bejáratát, amelyeken keresztül az oldószer molekulák bejuthatnak a belső szférába. Az átmenetifémet tartalmazó alegységet a (B) ábrán láthatjuk. Az N-terminális doménben a Ni(II)-ion kissé kinyúlik a négy α-hélix által övezett egységből. Az aktív centrumot a (C) ábrán tüntettem fel. Az összekötő egységek között (az A ábrán sárgával jelölt alegység) másodrendű kölcsönhatások lépnek fel, amelyek az egész komplexumot összetartják. A His1, a Glu17 és az Arg47 háromszög hidrogénkötésekkel kapcsolódnak egymáshoz és az C-hélixhez (az ábrán pirossal jelölve). Az A lánc (sárga rész) oxigén
A B C
Ni2+-SOD + 2 H++ O2 k2 Ni3+-SOD + H2O2 (11) 1.8. Réztartalmú szuperoxid dizmutáz enzimek
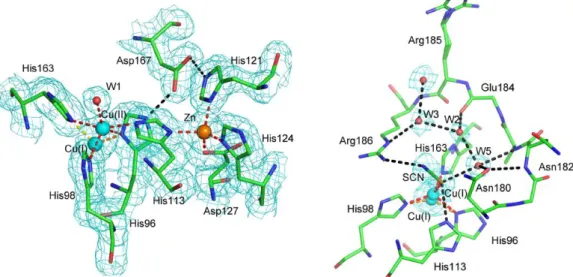
Ahogy az eddigi szuperoxid dizmutázok esetében, úgy itt is egy példán keresztül mutatnám be az analóg réztartalmú enzimek szerkezetét és főbb vonásaikat. A működési mechanizmusukat a (12-13) egyenletekben tüntettem fel [35], amely megegyezik a fentebb tárgyalt kobalttartalmú rendszer esetében leírtakkal. A réztartalmú szuperoxid dizmutázok hasonlóan az eddig tárgyalt enzimekhez dimer és tetramer szerkezetben is előfordulhatnak. Ami megkülönbözteti a többi rendszertől őket, az hogy az aktív centrumuk két központi iont is tartalmaznak és hogy az emberi szervezetben is gyakoribbak, mint korábban ismertetett társaik, ezért felépítésüket is a humán SOD3 tetramer enzimen keresztül fogom bemutatni [36]. Bár a cink szerepe még nem tisztázott, az egyértelműen kiderül az irodalomból, hogy a katalitikus funkciók ellátásáért a rézcentrum a felelős. A cinket helyettesítheti más átmenetifém is. Ha a röntgenfel- vételeket szemléljük, akkor láthatjuk hogy a 12. és 13. ábrán az előbbi állításunk igazo- lódik, a két különböző központ jól megkülönböztethető. Antonyuk és munkatársai azt is megállapították, hogy az aktív centrumban His113, His121, His124 és Asp127 egységek találhatók. Egy 8 Å széles csatorna vezet a réz(II)-ion oldali proteinegységek felületéhez.
Az egyik alegység mindegyik dimer esetében (ezek a B és C alegységek), tiocianát-aniont tartalmaz. A másik alegységek pedig koordinált vízmolekulát tartalmaznak a Cu-oldali övezetben. A His113 aminosav-egység imidazolgyűrűje hídligandumként funkcionál a réz- és cink-ionok között. A különálló dimer egységekben a réz(II)-ion ötös koordinációs számmal jellemezhető. Ha a SCN--ion kénatomján keresztül koordinálódik a rézhez, akkor azt redukálja, a hídligandum felhasad, az így kialakuló Cu(I)-ion már csak hármas koordinációs számú lesz. A csekély redukált állapotú réz(I)-ion jelenlétét az aktív centrumban a röntgenvizsgálat is bizonyítja. A réz-His113 kötéstávolság is bizonyítja a redukált állapot kialakulását, mert a kezdeti ~ 3,1 Å érték megváltozik, és lecsökken 2,3
Å-re, amely szintén igazolja a Cu(I)-SOD létrejöttét [37-39,40]. Tehát ez bizonyíték a rézcentrum kettős oxidációs állapotára, attól függően, hogy a tiocianát-ion koordinált vagy nem koordinált állapotban van a fémközpontot tekintve.
12. ábra A tetramer szerkezettel rendelkező emberi SOD3 Cu/Zn-SOD röntgendiffrakciós felvétele és a dimerizáció folyamata [36].
13. ábra Az emberi SOD3 tetramer Cu/Zn-SOD monomer egységeinek aktív centruma.
bal oldalon az A és D, jobb oldalon a C és D alegység felépítése látható.
A Cu-SOD-ok működési mechanizmusának általános egyenlete (12-13).
fékezze a nem kívánt melléktermékek túltermelődését, például kontrollálni és korlátozni az ártalmas anyagok koncentrációját. A szervezetben a dioxigén tetemes része az egyelektronos redukciójával képződő szuperoxid gyök-anionon keresztül metabo- lizálódik. Normális körülmények között az egészséges egyedekben a gyökök feldúsulását a SOD enzimek fékezik, melyek a mitokondriumban, sejtplazmában és a sejten kívüli térben vannak. Ez a káros, dioxigéntől származó szabad gyök bizonyítottan a reperfúziós betegségek közvetítője, valamint az azt követő akut miokardiális infarktusé és a sztróké, továbbá kimutatták, hogy társítható a gyulladást okozó folyamatok fejlődésével, mint például az artritisz és fő szerepet játszik számos neurológiai zavar megjelenésében, mint például a Parkinson-kór [41]. A kis molekulatömegű katalizátorok, melyek utánozzák a természetes enzimek funkcióját, használhatók számos betegség megelőzésére és gyógyítására, ahol az eredeti enzimek nem működnek. A szintetikus enzimek sok olyan betegség esetében használhatók, ahol a nem kívánt, toxikus, metabolikus melléktermékek túltermelése szerepet játszik a betegség kialakulásában és folytatásában. A SOD-utánzó vegyületeknek több előnyük is van a természetes enzimekkel szemben, például a sejtek közötti tér megközelítésének képessége, nagyobb áthatolóképesség a membránokon, hosszabb élettartam a vérben (az emberi enzimek rövid ideig stabilisak). Számos fém ismeretes, melyek komplexei katalizálják a szuperoxid gyök-anion bomlási folyamatát hidrogén-peroxiddá és dioxigénné (Cu, Mn, Fe, Ni). Az eddigi tanulmányok főként mangán- és vaskomplexekre irányultak, amelyek alacsony molekulatömegű SOD-utánzó enzimmodellekként hasznosíthatók [42], ugyanis a vegyületekben alkalmazott fém tulajdonságai nagyon fontosak, hiszen a katalizátor lebomlásával szabad fémion keletkezik, amely pedig toxikus hatású lehet. Gyógyszerként főként a mangán központú SOD-utánzókat alkalmazzák, mivel kevésbé mérgező hatásúak. A réz- és vasionok toxikusabbak, és elősegítik a Fenton-reakciót hidrogén-peroxiddal, amiből hidroxil gyökök keletkeznek. Fontos tehát, hogy az alkalmazott vegyület megfelelő stabilitású legyen, hogy az esetleges toxikus fémion ne tudjon felszabadulni. A megoldást olyan ligandumokkal képzett komplexek jelentik, melyek elősegítik a redox reakciót,
ugyanakkor elegendő stabilitással rendelkeznek a SOD-utánzó reakciók vizsgálatához. A helyettesített penta-aza-ciklusokkal [43] és a salen-származékokkal [44] képzett vegyületek eljutottak a klinikai tesztekig. A ligandumokat és az ezekkel képzett SOD- utánzó aktivitást mutató komplex vegyületeket a 14. ábrán összegeztük. Az ábrán feltüntettük az egyes komplexek IC50 értékeit, melyek a komplexek azon koncentrációját jelölik, amely 50 %-kal csökkenti a közvetett módszer [45] során alkalmazott indikátor vegyület és a szuperoxid gyök-anion között végbemenő redoxi reakció sebességét. Több esetben bizonyították, hogy egy vegyület akkor rendelkezik megfelelő SOD utánzó aktivitással, ha redoxi átmenetének féllépcső potenciálja az O2/O2·– és a O2·–/H2O2 átmeneteknek megfelelő -0,16 V < E1/2 (vs. NHE) < 0,89 V értékek közé esik pH = 7 esetén [46].
Ligandum IC50 Redoxi átmenet Ref.
X
OH NH
NH NH
HO
X
OH X
5,50 Mn(III)/Mn(II)
vagy Mn(IV)/Mn(III) [47]
N N
HB N N N
N
0,75 Mn(III)/Mn(II) [48]
N
NH N HN N
NH
N 4,30 Mn(III)/Mn(II) [49]
MeO2C
N N O
N O N
2,93 Mn(III)/Mn(II) [51]
14. ábra Mn-SOD-utánzó vegyületek.
A salenH2 (bisz(szalicilidén-etilén-diamin)) és a makrociklusos SOD utánzó vegyületeken kívül említést érdemelnek a metalloporfirin vegyületek is [51]. Ezek közül néhány SOD- utánzó vegyületként alkalmazott képviselőt a 15. ábrán tüntettünk fel a funkciós cso- portjaikkal együtt.
15. ábra Mezo-porfirin SOD-utánzó vegyületek szerkezete (TBAP = tetrakisz(4-ben- zoesav)porfirin, TM-4-PyP = tetrakisz-(N-metil-4-piridil)porfirin, OBTM-4-PyP = β-
oktabromo-tetrakisz(N-metil-4-piridil)porfirin és TM-2-Pyp = tetrakisz(N-metil-2-piri- dil)porfirin) [52].
16. ábra Mangántartalmú salen komplex (felül) és oligo(etilén-glikol) származékainak (alul) szerkezete.
A közelmúltban végzett kísérletek eredményeképpen elkészítették a mangántartalmú bisz- (3-metoxiszalicilidén)-1,2-etiléndiammin-klorid komplex oligo(etilén-glikol) (OEG) származékait (16. ábra), melyek hasonló, vagy akár két-, háromszor nagyobb SOD-utánzó aktivitást mutattak a standard salen komplexekhez képest [53].
1.9.1. Szuperoxid gyök-anion reakciója SOD utánzó vegyületekkel NBT (nitroblue tetrazolium) reagens jelenlétében [37,45]
A
szuperoxid dizmutáz enzimek és modelljeik aktivitásának mérése közvetlen és közvetett módszerekkel valósítható meg. A közvetlen mérések azonban csak költséges és nagy időfelbontású technikákkal valósítható meg, mint a stopped-flow és az impulzus radiolízis. Helyettük szélesebb körben közvetett módszereket alkalmaznak amelyet mi is választottunk méréseink kivitelezéséhez ahol a relatív sebességi állandókat határozzák meg egy vagy több referencia reagenssel szemben. A szuperoxid gyök-anion forrásaként egy enzimatikus reakciót választottunk, melynek során xantin oxidáz jelenlétében a xantin uronsavvá alakul (17. ábra). E reakció eredményeképpen szabadul fel a szuperoxid gyök-anion.A
NBT egy sárga színű tetrazolium-só, melyet a szuperoxid gyök-anion kékeslila színű mono-, illetve diformazánná redukál. A reakció megfelelő mennyiségű xantin oxidáz hozzáadására indul. A keletkező diformazán 560 nm-en követhető, ahol ΔA560 = 0,024-0,028 min-1 ~1 μmol/perc szuperoxid keletkezésének feleltethető meg. AAz alkalmazott módszer lényege, hogy mérjük a NBT redukciója során keletkezett diformazán abszorbanciaváltozását 560 nm-en különböző SOD utánzó komplex koncentrációk mellett. Az abszorbanciaváltozás (ΔA560), arányos a szuperoxid gyök- anion koncentráció-változásával, amennyiben SOD-utánzó vegyület nincs jelen (16)
ahol ε560 = a diformazán moláris elnyelési együtthatója, l = pedig az alkalmazott küvetta úthossza. Ha a rendszerben nincs SOD utánzó vegyület, akkor a (14) sebességi egyenlet érvényes, ha azonban SOD-utánzó vegyület is jelen van, akkor megindul a kompetitív reakció, ami szintén a szuperoxid gyök-anion koncentrációjának csökkenését okozza, így a sebességi egyenlet módosul, ahol a harmadik tag elhanyagolhatóan kicsi (17):
Ha a kompetitív reakciók azonos mennyiségű szuperoxidotfogyasztanak, akkor a (18) egyenlet érvényes:
ahol az IC50 érték az a SOD utánzó komplex koncentráció, amely a NBT redukciójának 50%-os inhibícióját okozza. Az inhibíció értékét a következő két módszerrel számolhatjuk ki:
2 2
2 SOD O [NBT]O
dt O d
NBT
SOD k
k
dt ε l d A dt
NBT d dt O
d 560
560 2
IC50 NBT NBT
SOD
k k
x x 0
ΔA ΔA I ΔA
ΔA .100 ΔA I(%) ΔA
o x o
(15)
(16)
(17)
(18)
(19) (20)
2 2
2 NBT O SOD O
dt O d
SOD
NBT k
k
A (19) és (20) egyenletekben ΔA0 a komplexet nem tartalmazó rendszer abszorbancia növekménye, a ΔAx érték pedig az x koncentrációban hozzáadott komplex jelenlétében mért abszorbancia különbség.
N N N N
NO2
H3CO
2
N H N N N
NO2
H3CO
2 O2
NH NH HN
N O
O N
H NH HN
NH O
O O
O2 O2 SOD-utánzó komplex
O2+ H2O2 kataláz O2+ H2O
xantin uronsav
xantin oxidáz
17. ábra A SOD-utánzó aktivitás mérése NBT reagens jelenlétében.
A gyakorlatban az abszorbancia változását mérjük növekvő [MnII(Ln)2] komplex koncentrációt alkalmazva, az IC50 értékeket leolvassuk az I(%) vs. komplex koncentráció görbéről. Az 1 komplexre vonatkozó mérési eredményeket tüntettük fel az alább látható ábrákon és táblázatokban. Az összes többi komplex esetében is hasonlóképpen történt az IC50 értékek meghatározása. Egy alternatív megoldás lehet, ha nem a 18/a. ábrán bemutatott exponenciális telítési görbét ábrázoljuk, hanem a
x x o
A A
A vs. [Mn] függvény szerinti egyenest. Ekkor y = 1 értéknél olvasható le az [Mn] = IC50 (18/b. ábra). A 29.
táblázat tartalmazza a MnII(Ln)2 komplexek mért IC50 értékeit és az ezekhez tartozó, ezekből a (27) egyenlet alapján számított kSOD sebességi állandókat.
NBT λ = 560 nm
diformazán
18/a. ábra Az 1 komplex IC50 értékének meghatározása NBT reagens jelenlétében az
I(%) vs. [1] görbe alapján.
18/b. ábra Az 1 komplex IC50 értékének meghatározása NBT reagens jelenlétében az
x x o
A A
A vs. [1] egyenes alapján.
1.9.2. Szuperoxid gyök-anion reakciója SOD utánzó vegyületekkel citokróm c(III) reagens jelenlétében [39,40]
A mérést a 1.9.1. pontban leírtak szerint végeztük el oly módon, hogy a NBT reagens helyett citokróm c(III) reagenst alkalmaztunk referenciaként. A xantin uronsavvá való átalakulása során felszabaduló szuperoxid gyök-anion redukálja a vas(III)-tartalmú citokróm c vegyületet vas(II)-tartalmú citokróm c vegyületté (19. ábra). A reakcióban
IC50
IC50
keletkező redukált forma fényelnyelését követjük UV-látható spektrofotométerrel 550 nm-en. A citokróm c(III) mérhető abszorbancia-változásának a ΔA550 = 0,024-0,028 min-1 tartományban kell lennie. A referencia reakció sebességi állandója ebben az esetben: kcit = 2,6×105 M-1 s-1, ha pH = 7,4-7,8 [40]. A SOD-utánzó vegyületek sebességi állandóit a (21) egyenlet szerint határoztuk meg a (14) egyenlethez hasonlóan az NBT helyett a citokróm c(III) reagens adatait figyelembe véve:
Az abszorbancia változását mértük növekvő [MnII(Ln)2] komplex koncentrációt alkalmazva, az IC50 értékeket leolvastuk a 20/a. ábrán és 20/b. ábrán feltüntetett függ- vények alapján, (az ábrán az 1 komplexre vonatkozó adatokat ábrázoltuk). A 30. táblázat tartalmazza a MnII(Ln)2 komplexek mért IC50 értékeit és az ezekhez tartozó, ezekből a (30) egyenlet alapján számított kSOD sebességi állandókat.
O2
NH NH HN
N O
O N
H NH HN
NH O
O O
O2 O2 SOD-utánzó komplex
O2+ H2O2 kataláz O2+ H2O citokróm c( III)ox citokróm c( II)red
xantin uronsav
xantin oxidáz
19. ábra A SOD-utánzó aktivitás mérése citokróm c(III) reagens jelenlétében.
IC50
c(III)
cit cit
SOD
k k
λ = 550 nm kcit[cit c(III)]
(21)
20/a. ábra Az 1 komplex IC50 értékének meghatározása az I(%) vs. [1] görbe alapján citokróm c(III) reagens jelenlétében.
20/b. ábra Az 1 komplex IC50 értékének meghatározása citokróm c(III) reagens jelenlétében az
x x o
A A
A vs. [1] egyenes alapján.
Elméletileg lehetőség van mellékreakciók kialakulására is a rendszerben, mint például az indikátor reagens NBT és a SOD-utánzó komplex, továbbá a SOD-utánzó komplex és a xantin oxidáz között. Ezért végeztük el citokróm c(III) jelenlétében is a vizsgálatokat, ahol hasonló eredményeket kaptunk (IC50), mint az NBT-s kísérleteknél. Bizonyítottuk, hogy a komplexek a xantin oxidázt nem gátolják, ugyanis az uronsav-képződés a λ =296
IC50
IC50
nm-es hullámhosszon követhető, a komplexeink tehát minden esetben csakis a szuperoxid gyök-anionnal lépnek reakcióba.
2.0. Kataláz enzimek
A kataláz enzimek az egyik legfontosabb védelmi funkciót ellátó enzimek (21. ábra). A különböző biológiai folyamatok melléktermékeként keletkező hidrogén-peroxidot dioxigénné és vízzé alakítják át a katalizátorfehérjék közül a legnagyobb hatásfokkal (22).
Ezek a vegyületek a vas- vagy mangántartalmú metalloenzimek csoportjába sorolhatók.
21. ábra Az emberi kataláz enzim [55].
2H2O2 kataláz 2H2O + O2
2.1. Mangántartalmú kataláz enzimek
A Mn-tartalmú kataláz enzimeket a Thermus thermophilus [56], a Lactobacillus plantarum [57] és a Thermoleophilum album [58] baktériumokból izolálták először. Ezen enzimek röntgenszerkezete alapján megállapítást nyert, hogy az aktív centrum két egymástól 3,03 Å távolságra lévő mangániont tartalmaz a Lactobacillus plantarum esetében, melyek karboxiláthídon keresztül kapcsolódnak (22. ábra).
(22)
22. ábra A Lactobacillus plantarum-ból elkülönített kataláz enzim szerkezete és aktív centruma [59].
A Mn-katalázoknak négy oxidációs állapottal rendelkező formáját jellemezték eddig.
Ezek a következők: (II,II), (II,III), (III,III), (III,IV). Csak a (II,II) és (III,III) oxidációs állapotú formák mutattak nagymértékű katalitikus aktivitást a H2O2 bomlási folyamatában. A Mn-katalázokban a két mangán magot először ESR-spektroszkópiával mutatták ki. Ezzel a technikával jellemezték a hármas és négyes oxidációs állapotokat is [60-62]. A meghatározott antiferromágneses csatolási állandók alapján szerkezeti modellekkel összehasonlítva már a röntgenszerkezetek ismerete előtt sejtették, hogy az aktív helyen a mangánionok (μ-OH) (μ-karboxilát) híddal vannak összekapcsolva [63].
Az ESR-rel a Mn(II,II) katalázban található mangánionok antiferromágneses csatolását is sikerült igazolni. EXAFS spektroszkópiával vizsgálva az egyes oxidációs állapotokat a Mn-Mn távolság észrevehetően lerövidül 2,7 Å-re a katalitikusan inaktív Mn(III,IV) formában az eredeti 3,03 Å helyett. Ez az alak valószínűleg bisz(μ-oxo)-hidat tartalmaz [64]. A Mn-katalázok fiziológiai aktivitását kizárólag csak a (II,II) és (III,III) oxidációs állapotoknak lehet tulajdonítani. A ping-pong ciklusú mechanizmus a két egymásba könnyen átalakítható oxidációs állapotnak köszönhető (23. ábra) [65,66].
A B D C
MnIII MnIII E OH
O H
O O
O O NHis O
NHis OGlu
OH
MnII Mn
OH
O O
OH2
H2NHis OGlu H2O
O2 O O
HOOH
H2O MnII MnII
OH O H
O O
O O NHis H2O
NHis OGlu
H
O OH O OH
H2O
MnII MnII OH
O H
O O
O O NHis H2O
H2NHis OGlu
O O
OH H
H2O MnII MnII
OH O
O O
O O NHis OH2
H2NHis OGlu
O O HOOH
O O
O O
23. ábra A kétmagvú mangántartalmú kataláz enzim által katalizált reakciók mechanizmusa [67].
A mangán-tartalmú katalázokról spektroszkópiai módszerekkel megállapították, hogy az enzimatikus reakció során redukált Mn(II,II), illetve oxidált Mn(III,III) formában vannak jelen. A vegyes oxidatív állapotok az enzimatikus reakcióban nem játszanak szerepet.
[68]. A reakció (A-E) első lépésében a szubsztrátum közvetlenül az ún. terminális (szélső) oldali Mn-t támadja meg, leszorítva onnan a vízmolekulát (A→B). Az internális (belső) oldali fehérjerész karboxilát csoportja protonpuffer szerepet tölt be. Deprotonálja a H2O2- t, majd később átad egy protont, és ezáltal tud víz képződni. A koordinált peroxid terminális helyzetből (B) elfordul, majd a μ-η2-peroxo híd képződik (C). A szubsztrátum redukálódik, víz lép ki, és a mangán-ionok távolsága lecsökken 3,6 Å-ről 3,1 Å-re (C→D). A továbbiakban μ-oxo híd protonálásával a híd felhasad és újabb szubsztrátum molekula lép be a koordinációs övezetbe (D→E). Ezután intramolekulárisan két elektron
medierekre alapozza a katalitikus ciklust [69]. A fenti enzimeknek direkt felhasználása gyógyászati célokra korlátozott, a sejtmembránok alacsony permeabilitása és a nagy molekulatömeg következtében. Az irodalomban számos példa található különféle alacsony molekulatömegű átmenetifém-tartalmú, Mn-tartalmú komplexekre (1. táblázat), amelyeket a kataláz enzimek szerkezeti és/vagy működési modelljeiként vizsgáltak [70- 80].
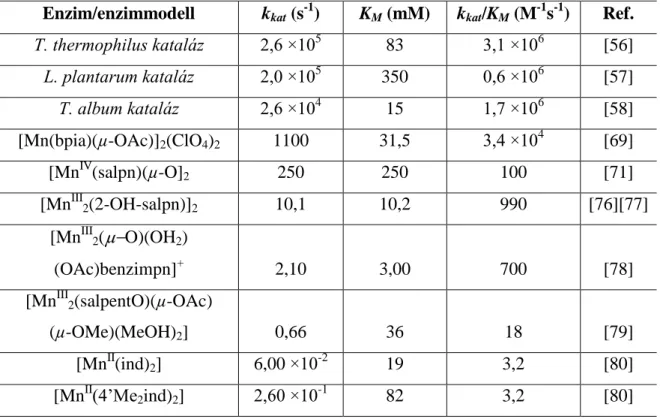
1. táblázat Néhány kataláz enzim és enzimmodell katalitikus aktivitása.
Enzim/enzimmodell kkat (s-1) KM (mM) kkat/KM (M-1s-1) Ref.
T. thermophilus kataláz 2,6 ×105 83 3,1 ×106 [56]
L. plantarum kataláz 2,0 ×105 350 0,6 ×106 [57]
T. album kataláz 2,6 ×104 15 1,7 ×106 [58]
[Mn(bpia)(µ-OAc)]2(ClO4)2 1100 31,5 3,4 ×104 [69]
[MnIV(salpn)(µ-O]2 250 250 100 [71]
[MnIII2(2-OH-salpn)]2 10,1 10,2 990 [76][77]
[MnIII2( O)(OH2)
(OAc)benzimpn]+ 2,10 3,00 700 [78]
[MnIII2(salpentO)(µ-OAc)
(µ-OMe)(MeOH)2] 0,66 36 18 [79]
[MnII(ind)2] 6,00 ×10-2 19 3,2 [80]
[MnII(4’Me2ind)2] 2,60 ×10-1 82 3,2 [80]
2.2. Ligandumszintézis
Az átmenetifém-tartalmú modellvegyületeink előállítására izoindolin alapvázú ligan- dumokat használtunk fel, amelyek szintetizálása alapvetően két lehetséges reakcióúton keresztül valósítható meg. A tervezettekhez hasonlóak szintézise során 1,2-diciano-
benzolból kiinduló olvadékfázisú-, és az 1,3-diimino-izoindolinból kiinduló folyadékfázisú reakciókat alkalmaztak [81]. Az olvadékfázisú folyamatok (24. ábra) magas hőmérsékletet igényelnek, ezáltal a reakcióidő rövidebb, mint a folyadékfázisú szintézis esetében [82]. Az izoindolin oldalkarokkal rendelkező kelátképzők kialakulását a kilépő ammónia képződésének vége jelzi.
CN CN
+ 2 NH
N
N - NH3
N N H H2N
N HN
HN N
24. ábra A HL1 ligandum előállítása 1,2-diciano-benzolból.
A másik lehetséges előállítási módszer az 1,3-diimino-izoindolinból kiinduló folyadékfázisú folyamat, amely mechanizmusát tekintve egy AN típusú lépéssel indul (25.
ábra) [83]. A fenti reakciók során oldószerként alkoholokat használnak, a legjobb hozamok elérése végett többnyire n-BuOH-t alkalmaznak.
+ 2
-2 NH3 n-BuOH
NH N
N NH
NH
NH
N NH H2N
N HN
HN N
25. ábra A HL1 ligandum előállítása 1,3-diimino-izoindolinból.
![4. ábra A SOD enzim működésének optimuma a lipid peroxiláció során [16].](https://thumb-eu.123doks.com/thumbv2/9dokorg/873393.46986/17.892.255.607.357.693/ábra-sod-enzim-működésének-optimuma-lipid-peroxiláció-során.webp)
![22. ábra A Lactobacillus plantarum-ból elkülönített kataláz enzim szerkezete és aktív centruma [59]](https://thumb-eu.123doks.com/thumbv2/9dokorg/873393.46986/36.892.128.744.106.334/ábra-lactobacillus-plantarum-elkülönített-kataláz-szerkezete-aktív-centruma.webp)
![23. ábra A kétmagvú mangántartalmú kataláz enzim által katalizált reakciók mechanizmusa [67]](https://thumb-eu.123doks.com/thumbv2/9dokorg/873393.46986/37.892.121.741.103.648/ábra-kétmagvú-mangántartalmú-kataláz-által-katalizált-reakciók-mechanizmusa.webp)