CHAPTER SIX
Microbial Activities of Soil As They Affect Plant Nutrition
J. H . Q U A S T E L
I. Microbial Activities of Soil and Soil Structure 672
A. Organic Matter in Soil 672 B. Soil as a Biological System 687 C. Organic Matter and Soil Aeration and Structure 689
D. Soil Improvers (Conditioners) 692 II. Nitrogen Metabolism in Soil 697
A. Main Steps in the Nitrogen Cycle in Soil 697
B. Formation of Ammonia in Soil 699
C. Amino Acids in Soil 699 D. Nitrification in Soil 700 E. Effects of Organic Substances on Nitrification of Ammonium I o n s . . . . 706
F. Oxidation of Nitrite in Soil 710 G. Conversion of Other Nitrogen Compounds to Nitrate 712
H. Denitrification in Soil 713 III. Sulfur Metabolism in Soil 717
A. Forms of Sulfur in Soil 717 B. Oxidation of Sulfur in Soil 718 C. Metabolism of Sulfur Amino Acids in Soil 722
D. Sulfate Reduction in Soil 723 E. The Cycle of Sulfur Transformations in Soil 724
F. Influence of Manganese Dioxide 724 G. Herbicidal Effects of Thiosulfates 725 IV. The Rhizosphere and Plant Nutrition 726
A. Microorganisms in the Rhizosphere 726 B. Nutrients, Vitamins and Cofactors, and Other Organic Substances in the
Rhizosphere 727 C. The Microflora of the Rhizosphere and Plant Growth 730
D. Relations of Amino Acids, Auxins, and Herbicides in the Rhizosphere
to Plant Growth 731 V. Studies of Soil Metabolism 733
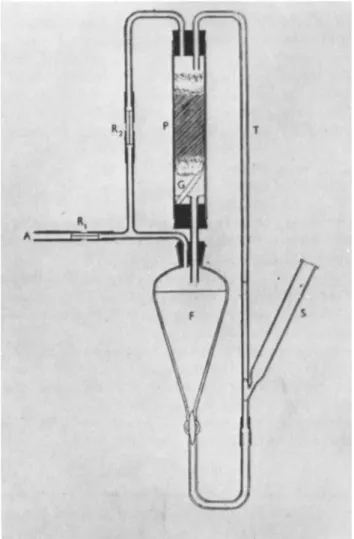
A. The Perfusion Technique 733 B. Metabolic Studies and Manometric Studies of Soil 738
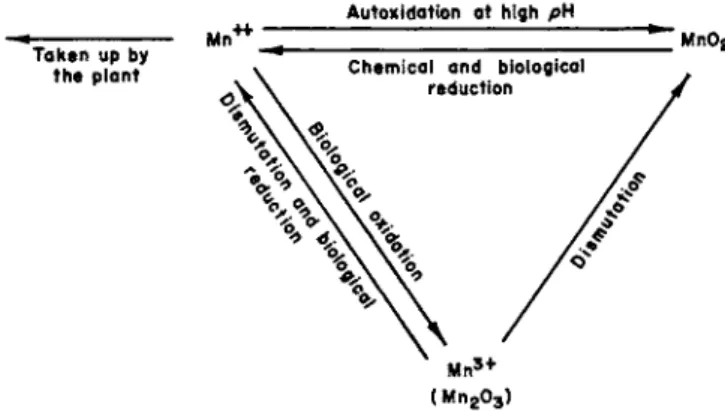
C. Role of Manganese and of Iron 739
VI. Concluding Remarks 742
References 743 671
672 J. H . QUASTEL
I. Microbial Activities of Soil and Soil Structure
A. ORGANIC M A T T E R I N SOIL
T h e organic m a t t e r of soil is p r i m a r i l y derived from t h e decomposi- tion products of plants a n d from the products of autolysis of t h e micro- biological population w h i c h develop at the expense of substances of p l a n t origin. A d y n a m i c equilibrium exists in t h e soil between t h e large n u m b e r s of bacteria, protozoa, yeasts, fungi, etc., t h a t inhabit the soil a n d t h e organic m a t t e r continually being supplied b y the decaying a n i m a l a n d vegetable tissues. T h i s equilibrium is such t h a t t h e ratio of carbon to nitrogen in soil organic m a t t e r is r e m a r k a b l y constant, a p p r o x i m a t e l y 10. Autotrophic bacteria, w h i c h obtain their carbon from atmospheric carbon dioxide, a n d t h e nitrogen fixers, such as species of Azotobacter, p l a y a f u n d a m e n t a l p a r t i n w i t h d r a w i n g carbon dioxide a n d nitrogen from the air into the organic m a t t e r of soil.
Soil n o r m a l l y contains a variety of substances c o m m o n l y classed as metabolites of plants a n d animals. T h e speed of change of these sub- stances m a y be v e r y great, t h e r a t e depending on t h e t e m p e r a t u r e , h y d r o g e n ion concentration, aeration, w a t e r availability, a n d m i n e r a l content of soil. A n u m b e r of substances, derived from animals, plants, or microorganisms, are, however, relatively resistant to breakdown, a n d such substances m a k e u p for t h e most p a r t w h a t is c o m m o n l y k n o w n as soil organic m a t t e r . Such substances h a v e physical a n d chemical properties w h i c h together help to d e t e r m i n e t h e structure of a soil.
T h e s e substances a r e also i n a continuous state of change, b u t a n equi- l i b r i u m is achieved so t h a t their total q u a n t i t y r e m a i n s fairly constant i n a n y one locale, t h e a m o u n t being largely dependent on climate a n d geographical conditions a n d t h e types of p l a n t being supplied.
Humus
A m o n g t h e most characteristic of the products w h i c h compose soil organic m a t t e r is t h e black colloidal material, h a v i n g h i g h hydrophilic power, k n o w n as h u m u s , whose presence helps to m a k e possible t h e c r u m b structure of soil so necessary for its fertility. H u m u s is a com- plex m i x t u r e of w h i c h h u m i c acid is a n i m p o r t a n t constituent. It is a p p a r e n t l y formed i n at least t h e following w a y s : (a) b y bacterial attack on cellulose, (b) b y chemical transformations of lignin, (c) from fungal mycelia. H u m i c acid from soil is stated to contain about 5 % nitrogen; it cannot be wholly, therefore, a lignin or cellulose condensa- tion product. Oden (179) believed it to be a tetrabasic acid w i t h a n equivalent weight of about 300.
6. M I C R O B I A L A C T I V I T I E S O F S O I L A N D P L A N T N U T R I T I O N 673
M a n y of the characteristics of soil organic m a t t e r are consistent w i t h the view t h a t lignin, or lignin derivatives, represents a major con- stituent. T h i s is m a d e m o r e likely b y t h e fact t h a t lignin is v e r y resist- a n t to microbiological attack. Lignins from various sources v a r y in composition; thus, t h e lignin from grass roots is not identical w i t h the lignin of oak leaves or of pine needles.
Various views h a v e been advanced to account for the formation of h u m u s . It was originally thought t h a t h u m u s is derived from cellulose in the soil. T h u s Beckley (18) found t h a t sugars on t r e a t m e n t w i t h acids give rise to h y d r o x y m e t h y l f u r f u r a l d e h y d e , w h i c h readily con- denses to form a substance closely resembling h u m u s . H e found indi- cations of t h e presence of this aldehyde in a dunged soil a n d in rotting straw in w h i c h h u m u s was being formed. A n alternative view (53) was advanced t h a t h u m u s w a s derived from t h e oxidation of quinones.
Fischer (57) suggested t h a t lignin is t h e p a r e n t substance of h u m i c acid. It w a s a l r e a d y k n o w n that, d u r i n g t h e bacterial decay of wood, cellulose disappears w i t h i n t h r e e years w h e r e a s t h e lignin suffers b u t little diminution in t h a t period. L i g n i n a n d h u m i c acid have, chemi- cally, m u c h in common ( 5 8 ) , a n d studies of t h e fractionation of organic m a t t e r i n m i n e r a l soils (4) indicated t h a t lignin w a s a source of h u m i c m a t t e r of soil. Moreover, there was a correlation between the formation of h u m i c acid a n d loss of lignin, b u t not t h a t of cellulose or pentosan.
H u m i c acid of soil is a t r u e acid a n d m a y be obtained from diverse soils such as peat a n d agricultural soil ( 1 7 9 ) . It readily gives rise to colloidal solutions. Its salts w i t h alkali metals are soluble a n d behave as colloidal electrolytes. T h e salts w i t h other metals are insoluble.
W a k s m a n ( 2 5 5 - 2 5 7 , 261) has emphasized t h e lack of satisfactory methods of characterizing h u m i c m a t t e r a n d has pointed out t h a t m a n y of t h e products referred to as h u m u s a n d h u m i c acid are not homo- geneous. H e uses the t e r m h u m u s to designate "organic residues which h a v e u n d e r g o n e extensive decomposition b y microorganisms w h e t h e r i n peat bog or in compost, in forest soil or in m i n e r a l soils . . . . T h e chemical composition of h u m u s varies considerably, depending on t h e n a t u r e of the p l a n t residues, on t h e conditions of decomposition, on the extent of decomposition a n d t h e microorganisms active in t h e decom- position process. H u m u s is not in a static condition b u t undergoes con- tinuous change, t h e r a n g e of change depending u p o n t h e above condi- tions. H u m u s is, therefore, not constant i n chemical composition."
W a k s m a n has published (255) t h e figures given in Table I, showing analyses of soil h u m u s derived from a v a r i e t y of sources. T h e results given in T a b l e I show that, w i t h certain exceptions, n a t u r a l h u m u s formations tend to be low in carbohydrate a n d to have a high lignin
674 J. H . QUASTEL
a n d protein content. T h e s p h a g n u m peats a r e exceptional because (a) the carbohydrates of t h e s p h a g n u m p l a n t are m o r e resistant to decom- position t h a n those of t h e grasses; this in t u r n produces a low protein accumulation because insufficient carbon is released to build u p the protein of t h e microorganisms, a n d (b) t h e h i g h l y acid conditions of s p h a g n u m bogs reduce the r a t e of decomposition b y microorganisms.
Constituents of the carbohydrate group, e.g., hemicelluloses of t h e uronic acid type, a r e also contained i n t h e resistant fraction of t h e h u m u s . Soil h u m u s , after a variety of transformations proceeding at different
T A B L E I
CHEMICAL NATURE OF SOIL HUMUS®
Per cent of total humus in ash-free material Lignins and lignin-
Nature of humus Carbohydrates like derivatives Proteins Hardwood-spruce forest, F layer 27 .16 43.18 9.11 Hardwood-spruce forest, H layer 17 .00 57.00 8.49 Lowmoor peat, Newton 11 .85 44.08 25.84 Lowmoor peat, Florida 7 .43 51.24 25.62 Highmoor peat, young sphagnum 37 .04 34.72 5.33 Highmoor peat, old sphagnum 21 .76 53.25 5.85
Alpine humus 17 .95 35.18 35.77
Chernozem soil, Kansas 13. .88 40.81 34.74 Prairie soil (Carrington loam) 11. 85 49.29 30.38
eF r o m Waksman (255).
rates on t h e various p l a n t products, tends to h a v e t h e following com- position: (a) lignin-protein complexes, 6 0 - 8 0 % ; (b) certain resistant hemicellulose a n d w a x y substances, 5 - 1 2 % ; (c) cellulose a n d other products i n a n active state of transformation, 5 - 2 5 % . W h e n t h e h u m u s is treated w i t h alkali, a large p a r t of groups (a) a n d (b) are brought into solution leaving group (c) undissolved, the latter being " h u m i n . "
W h e n t h e alkali extract is acidified, t h e major p a r t of (a) a n d some of (b) a r e precipitated giving t h e " h u m i c acids," while a p a r t of group (b) a n d possibly a p a r t of (c) a r e left in solution to be t e r m e d the
"fulvic acid" fraction. T e n n e y a n d W a k s m a n (244, 245) h a v e shown t h a t the decomposition of p l a n t products i n t h e soil proceeds m u c h m o r e slowly u n d e r anaerobic conditions t h a n u n d e r aerobic. This is t r u e especially of t h e lignins a n d organic nitrogenous complexes w h e n compared w i t h t h e decomposition of celluloses a n d hemicelluloses.
Although in fresh p l a n t residues t h e ratio of carbon to nitrogen varies from 2 0 0 : 1 to 5 0 : 1 , the h u m u s in t h e soil shows a n a r r o w ratio of C : N
6 . M I C R O B I A L A C T I V I T I E S O F S O I L A N D P L A N T N U T R I T I O N 6 7 5
approximating 1 0 : 1 , w i t h variations depending on factors including the n a t u r e of organic residues a n d extent of decomposition. It is unlikely t h a t t h e organic nitrogenous complexes in the h u m u s are free proteins;
t h e y a r e often regarded as lignoproteinates. T h e y a r e accompanied b y pectins, uronic acids, fats, waxes, etc. Organic m a t t e r of the soil m a y be divided ( 2 6 0 ) into two groups:
1. T h e h u m u s nucleus, consisting of lignoproteins often combined w i t h bases, e.g., calcium, iron, a l u m i n u m , a n d probably also w i t h sili- cates a n d phosphates. T h e s e lignoproteinates, possibly forming com- plexes w i t h t h e hemicelluloses, form the fraction designated as h u m i c acid a n d are responsible for the characteristic color a n d organic col- loidal properties of soil.
2 . T h e r e m a i n i n g constituents of h u m u s , consisting of cellulose, hemicelluloses, waxes, fats, starches, etc., i n various a m o u n t s a n d pro- portions according to the location, e.g., forest soils, h i g h l a n d peats.
T h e s e together form t h e fraction k n o w n as " h u m i n . "
T h e lignoprotein complexes a r e not completely resistant to decom- position a n d m a y be broken down b y a v a r i e t y of fungi including t h e mycorhiza associated w i t h the roots of trees.
2. Lignin
According to H i b b e r t ( 9 2 , 9 3 ), t h e older ideas of Klason t h a t lignin is a coniferyl t y p e (e.g., coniferyl alcohol, coniferaldehyde, or oxyconiferyl alcohol) of condensation p o l y m e r h a v e been largely confirmed. Investi- gations h a v e shown (a) t h a t a large yield of vanillin is formed from spruce {Picea) wood ( a r o u n d 2 5 % ) a n d a still larger combined yield of vanillin (I) a n d syringaldehyde ( I I ) ( a r o u n d 4 5 % ) is formed from
CHO CHO
Vanillin Syringaldehyde (I) (Π) m a p l e {Acer) wood; (b) t h a t spruce lignin is a m i x t u r e of condensa- tion polymers formed from simple u n i t s containing a n aromatic r i n g h a v i n g a three-carbon side chain (in support of this view, such products as a-hydroxypropiovanillone a n d vanilloyl m e t h y l ketone h a v e been isolated) ; a n d (c) t h a t hydrogénation of wood a n d of extracted lignins produces a h i g h yield of cyclohexane derivatives h a v i n g a three-carbon side chain.
676 J. H . QUASTEL
T h e lignins of all softwoods seem to be identical, consisting of guaiacylpropane units connected to each other through ether a n d car- bon-carbon linkages. Those of the hardwoods differ b y t h e presence of derivatives containing the syringyl radical in addition to the guaiacy]
radical.
F r e u d e n b e r g a n d his colleagues (63c, e, f, g, i ) , using a lignin ob- tained b y alternate extraction of spruce wood m e a l w i t h cupric am- m o n i u m hydroxide a n d hot dilute sulfuric acid with subsequent methyla- tions a n d oxidations of the m e t h y l a t e d products, w e r e able to isolate veratric acid ( I I I ) , isohemipinic acid ( I V ) , a n d dehydrodiveratric acid ( V ) . F r e u d e n b e r g concluded, from these a n d other studies, that lignin COOH COOH COOH COOH
HOOCkv^,OCH3 CH30 OCH
(IV) (V) is formed as a result of a p r e l i m i n a r y condensation between t h e side
chains of one molecule a n d the aromatic nucleus of another, thus:
CHOH.COCH3
All or most of t h e reactive groups a r e located at the alpha carbon atoms of the side chains. Hibbert (92, 93) impressed b y t h e fact t h a t t h e principal water-soluble ethanolysis products isolated from spruce a n d m a p l e wood represent ene-diol a n d 1,2-diketone oxidation systems (VI,
Ç—OH
'OCH3 lX ^ O C H3
6. M I C R O B I A L A C T I V I T I E S O F S O I L A N D P L A N T N U T R I T I O N 677
V I I ) a n d t h a t these substituted p h e n y l p r o p a n e compounds a r e ap- p a r e n t l y t h e building units from w h i c h lignin is formed, suggests t h a t t h e latter "is derived from a series of monomolecular h y d r o g e n transporting p l a n t catalysts, which, d u r i n g t h e life of the p l a n t a n d i n t h e subsequent post-mortal stage, u n d e r g o conversion into m o r e complex condensation p o l y m e r products." T h e p l a n t catalysts in question are R C O C H O H C H3, R C O C O C H3, R C H O H C O C H3, R - C H2 #C O * C H2* O H , a n d molecules of similar n a t u r e w h e r e R = guaiacyl or syringyl.
T h e s e substances probably p l a y a n i m p o r t a n t role in p l a n t m e t a b - olism, b u t their m o d e of action is at present obscure.
T h e r e seems to be a g r e e m e n t t h a t in t h e p l a n t lignin is attached to carbohydrates, possibly polyuronic acids. P r e s u m a b l y such complexes exist i n the soil, for t h e y a r e only v e r y slowly broken down b y micro- organisms.
a. Biosynthesis of lignin. F r e u d e n b e r g (63a) a n d his colleagues have studied biosynthesis of lignin b y introducing radioactive substances into spruce saplings ( 6 3 h ) . T h e y found t h a t radioactive lignin is formed from D-coniferin, b u t not from L-coniferin, a n d from radioactive p h e n y l - alanine, t h e latter being converted first into radioactive coniferin.
T h e r e is r a p i d transformation of p h e n y l a l a n i n e into coniferyl alcohol a n d of t h e latter into its glucoside.
L i g n i n m a y be formed in vitro b y bubbling air t h r o u g h a dilute ( < 0 . 5 % ) solution of coniferyl alcohol i n presence of laccase (mush- room) at 2 0 ° C at p H 5.5-6.5. C a m b i u m laccase m a y be used instead of m u s h r o o m ( 6 3 d ) . Moreover lignin can also be formed from coniferyl alcohol b y using horseradish (Armoracia lapathifolia) peroxidase a n d dilute h y d r o g e n peroxide. Biosynthetic a n d n a t u r a l lignins seem to be identical, w i t h molecular weights about 10,000 ( 6 3 b ) . Laccase a n d peroxidase cannot attack coniferin; only coniferyl alcohol is attacked to form lignin. D u r i n g t h e vegetative period t h e tissues of t h e c a m b i u m a n d adjacent cells contain m u c h coniferin. This is split b y a ß-glu- cosidase present i n these cells to form coniferyl alcohol w h i c h is t h e n transformed into lignin. T h e cell dries as soon as the hollow spaces of t h e cell w a l l a r e filled w i t h lignin.
Intermediates between coniferyl alcohol a n d lignin a r e n u m e r o u s a n d complex; t h e y include: dehydrodiconiferyl alcohol, dZ-pinoresinol, guaiacylglycerol-ß-coniferyl ether, a n d coniferyl aldehyde. All these intermediates as well as lignin itself are optically inactive. T h i s is due to t h e fact t h a t e n z y m a t i c attack on coniferyl alcohol causes t h e removal of a phenolic h y d r o g e n atom, leaving a radical w h i c h exists i n a variety of mesomeric forms a n d w h i c h a r e optically inactive. These on combin-
678 J. H . Q U A S T E L
ÇH2OH CH2OH I
CH
π II
Λ
xι
•k^ O C H , CH Οο
II
Dehydrodiconiferyl alcohol
(vm)
2OH
OH 3
where R = CH3 or C1 2C2 1O10 (K)
Lignin can undergo further dehydrogenations in presence of laccase or peroxidase. I n t h e p l a n t lignin synthesis takes place, b u t no further reactions occur w h e n t h e cell walls are fully lignified.
I n soil, however, lignin is attacked b y laccases or peroxidases from wood-rotting fungi a n d breakdown takes place. T h e presence of these e n z y m e s in the fungi enables t h e organisms to obtain breakdown products, p r e s u m a b l y for their nourishment. T h e b r e a k d o w n of lignins in this m a n n e r is analogous to the b r e a k d o w n of phenols a n d is p a r t of the humification process of soils.
A review of the methods of h u m u s analysis, including t h e lignin fractions, is given b y W a k s m a n ( 2 5 8 ) .
3. Carbohydrate Decomposition in Soil
Carbohydrates represent a major constituent of t h a t p a r t of soil organic m a t t e r w h i c h undergoes relatively r a p i d transformation. T h e y ing w i t h each other form various a r r a n g e m e n t s , e.g. ( V I I I ) , w h i c h m a y combine ( I X ) with m e t h a n o l or cane sugar.
6 . M I C R O B I A L A C T I V I T I E S O F S O I L A N D P L A N T N U T R I T I O N 6 7 9
are quickly broken down, b u t t h e y a r e also continually being resyn- thesized b y the great v a r i e t y of organisms t h a t are nourished b y soil organic matter. A t the same time, decaying p l a n t m a t e r i a l enriches the soil w i t h a constant supply of a variety of carbohydrates.
T h e continued presence of carbohydrates in the soil stimulates t h e proliferation of those organisms t h a t can attack these substances a n d derive from t h e m t h e e n e r g y a n d t h e carbon for their growth. Soil, therefore, n o r m a l l y contains m a n y types of carbohydrate-splitting organisms.
Various factors influence t h e n u m b e r s a n d types of organisms t h a t respond to carbohydrates. Such factors are t h e availability of nitrogen, the presence or absence of oxygen, t h e h y d r o g e n ion concentration, a n d the moisture content of t h e soil.
Carbohydrate metabolism in a soil, therefore, u n d e r field conditions depends v e r y m u c h on t h e t y p e of soil, on its structure a n d its location.
Forest soils, w h i c h a r e u s u a l l y acidic i n n a t u r e , show a great pre- ponderance of fungi w h e r e a s limed, m a n u r e d soils of n e u t r a l or alkaline p H show a preponderance of bacteria. T h e forms of metabolism of carbohydrate will v a r y according to t h e site of t h e soil a n d its previous treatment. Anaerobic soils, such as those presented b y bogs a n d fens, show a different metabolic behavior, t h e t y p e of carbohydrate break- down associated w i t h anaerobic life being dominant. T h e m a i n product of carbohydrate b r e a k d o w n in t h e soil is bacterial or fungal m a t t e r . It has been estimated t h a t between 4 0 a n d 7 0 % of t h e carbon of cellu- lose t h a t is decomposed in soil is transformed into microbial cells. T h e m a i n carbohydrates in soil are represented b y cellulose, hemicellulose, starches including lichenin, dextrans, a n d levans, a n d t h e simple sugars.
T h e rates of attack of these substances in soil v a r y v e r y considerably;
t h e simpler sugars are broken down relatively quickly w h e r e a s certain hemicelluloses, including t h e polyuronides, are m o r e resistant to attack.
a. Cellulose breakdown. A large v a r i e t y of organisms attack cellu- lose, including anaerobic a n d aerobic bacteria a n d fungi. Actinomycetes a n d t h e filamentous fungi, including Fusarium, Aspergillus, Pénicil- lium, a n d Trichoderma, a r e all involved. According to W a k s m a n , in the breakdown of wood b y fungi, t h r e e stages m a y be distinguished:
(a) attack b y U r e d i n e a e a n d Ustilagineae t h a t decompose sugars a n d starches, (b) attack b y fungi belonging to t h e genera Mucor, Pénicil- lium, a n d Aspergillus t h a t b r i n g about t h e destruction of sugars a n d starches a n d decompose hemicellulose a n d polyuronides, a n d (c) de- velopment of t h e t r u e wood-destroying fungi belonging to t h e Poly- poraceae a n d Agaricaceae t h a t attack both cellulose a n d lignin. Protozoa a n d a variety of wood-boring insects (termites, cockroaches, etc.) are also involved in cellulose decomposition in soil. T h e products of cellu-
680 J. H . QUASTEL
lose decomposition m a y be, besides carbon dioxide, organic acids (chiefly acetic, butyric, a n d valeric acids), alcohols, hydrocarbons (e.g., m e t h a n e ) a n d hydrogen. These are usually produced b y anaerobic organisms. Aerobic bacteria a n d fungi seem to b r i n g about the complete destruction of cellulose.
Cellulose is insoluble in water, a n d p r e s u m a b l y organisms t h a t can break down a n d flourish on cellulose m u s t h a v e the m e a n s of producing from it some soluble product t h a t can pass into t h e cell. T h e conven- tional view is t h a t a n exoenzyme brings about hydrolysis of t h e cellu- lose molecule with subsequent liberation of glucose, w h i c h is destroyed w i t h i n the cell—the products formed v a r y i n g w i t h the n a t u r e of the organism. It is not k n o w n w h e t h e r cellulose dextrins a r e produced in the e n z y m a t i c attack on cellulose, b u t the evidence indicates that two types of hydrolytic attack take place, one accomplishing t h e initial attack, possibly r u p t u r i n g cross linkages or securing considerable chain shortening, a n d t h e other bringing about hydrolysis of t h e fragments.
T h e evidence for this is the observation t h a t insoluble dextrans prepared b y cold acid hydrolysis of cellulose a r e a p p a r e n t l y utilized m o r e readily a n d b y m o r e varieties of organisms t h a n u n t r e a t e d cellulose itself.
P r i n g s h e i m (189) postulated t h e existence of two hydrolytic e n z y m e systems, cellulase a n d cellobiase, the former producing cellubiose from cellulose a n d t h e latter breaking down t h e disaccharide to glucose.
Simola ( 2 2 0 ) , in a study of two aerobic spore-forming organisms n a m e d Cellulobacillus myxogenes a n d Cellulobacillus mucosus, obtained results supporting Pringsheim's view. Both glucose a n d cellobiose w e r e identi- fied a m o n g t h e products of breakdown.
Cellulases are n o w k n o w n to be w i d e l y distributed in p l a n t products, particularly in seeds, in t h e digestive secretions of invertebrates, a n d in extracts of various molds a n d bacteria. T h e y a p p e a r to be produced b y the n o r m a l intestinal flora of insects. T e r m i t e s can exist for long periods on a diet of p u r e cellulose w h e n t h e n o r m a l flora representatives of two orders of flagellates are present. Cellulases develop in fungi a n d bacteria w h e n these are g r o w n w i t h cellulose as substrate, a n d it is possible t h a t t h e y a r e to be regarded as adaptive enzymes. Cellulases of invertebrates are obtained from t h e gut; snails represent a rich source.
E n z y m a t i c hydrolysis of cellulose leads almost wholly to rf-glucose, b u t cellobiose is probably a n intermediate i n t h e process.
T h e variable resistance of cellulose to e n z y m a t i c attack has been ascribed to chain length (degree of p o l y m e r i z a t i o n ) , but there is strong evidence t h a t the principal factor is the degree of crystallinity. A com- parison of the equilibrium moisture contents of cellulose of widely dif-
6. M I C R O B I A L A C T I V I T I E S O F S O I L A N D P L A N T N U T R I T I O N 681
ferent reactivities showed a direct correlation w i t h t h e extent of e n z y m a t i c hydrolysis. H i g h l y variable results h a v e been recorded con- cerning t h e degradation of cellulose b y microorganisms, this being doubtless due to variations i n the physical state of t h e cellulose. It is well k n o w n t h a t native cotton (Gossypium) cellulose a n d wood are m u c h m o r e resistant to e n z y m a t i c hydrolysis t h a n regenerated cellulose.
T h e r e is, unfortunately, little evidence as to t h e n a t u r e of t h e initial attacks b y e n z y m e s on cellulose, beyond t h e fact t h a t t h e y are pri- m a r i l y hydrolytic. W i n o g r a d s k y (276) p u t forward t h e view t h a t a n oxidation took place, the t e r m i n a l alcohol groups being oxidized to aldehydes w h i c h w e r e t h e n oxidized to carboxyl groups. Cellulose at- tacked b y bacteria was observed to have acid properties. I n support of this view, W a l k e r a n d W a r r e n (263) isolated a g u m from cellulose suspensions decomposed b y Cytophaga a n d showed it to be a n acid, yielding furfuraldehyde a n d carbon dioxide on t r e a t m e n t w i t h 1 2 % hydrochloric acid, a p r o p e r t y common to polyuronides. Because of its low reducing power, t h e y considered t h e g u m to be a n acidic t y p e of oxycellulose a n d stated t h a t it was a n intermediate stage in cellulose breakdown. T h i s interpretation, however, according to N o r m a n a n d his colleagues (176, 177), is not justified, as oxidation would not, itself, cause i m m e d i a t e chain splitting to such a n extent t h a t all t h e oxidized fragments would be w a t e r soluble. T h e r e was no evidence t h a t the residual cellulose w a s m o r e oxidized t h a n t h e original sample, a n d uronic acid groupings, previously introduced b y chemical m e a n s , w e r e actually preferentially removed b y t h e organism.
T h e latter stages of cellulose decomposition are not m u c h m o r e sat- isfactorily understood t h a n t h e earlier stages. I n general, t h e aerobic organisms effect extensive oxidations, t h e major product being carbon dioxide. Two-thirds of t h e carbon removed from cellulose decomposed b y Cytophaga is removed as carbon dioxide, a n d almost all the r e m a i n - der can be accounted for in t h e g u m synthesized b y t h e organism
( 2 6 3 ) . N o n e of the species of Vibrio a p p a r e n t l y produces acid from cellulose. T h e two aerobic sporeformers, studied b y Simola ( 2 2 0 ) , yield about 1 0 % volatile acids acetic a n d formic from cellulose, t h e acetic acid being in considerable excess. Anaerobic organisms give larger yields of acid products a n d even alcohol from cellulose—so m u c h so t h a t attempts h a v e been m a d e to develop large-scale fermentation processes involving cellulose, b u t so far w i t h little success. Over fifty years ago, Omelianski showed t h a t certain anaerobes will decompose cellulose to give high yields of fatty acids, chiefly acetic a n d butyric, together w i t h carbon dioxide a n d either h y d r o g e n or m e t h a n e , but it is n o w consid- ered unlikely t h a t m e t h a n e is a product of cellulose decomposition. It
682 J. H . QUASTEL
is m o r e likely t h a t it is produced b y t h e action of t h e m e t h a n e organ- isms t h a t b r i n g about a reduction of carbon dioxide b y h y d r o g e n or b y hydrogénation. T h e products of anaerobic b r e a k d o w n of cellulose are those t h a t m i g h t be expected from the anaerobic decomposition of glu- cose, n a m e l y , acetic, butyric, a n d lactic acids, ethyl alcohol, carbon dioxide, a n d hydrogen.
Although celluloses are vigorously attacked b y fungi, little is k n o w n of the intermediate products. As is well k n o w n , fungi produce from sugars a variety of acids, such as citric, oxalic, fumaric, succinic, a n d gluconic—but n o n e of these acids is reported d u r i n g the fungal attack of cellulose, although of course t h e y m a y be formed a n d r a p i d l y broken down. M u c h work, w i t h selective inhibitors, is required to t h r o w light on t h e action of cellulose degradation b y fungi. T h e most obvious prod- uct of cellulose metabolism is synthesized microbial substance. As
m u c h as 5 0 % of t h e carbon of cellulose finds its w a y into t h e m y c e l i u m of fungi. Available nitrogen is required for this process, a n d it is cal- culated t h a t t h e ratio between cellulose decomposed a n d nitrogen im- mobilized is about 35 to 1.
T h e r m o p h i l i c bacteria h a v e been studied chiefly w i t h a view to the development of a n industrial fermentation process to obtain acids or alcohol from cellulose. Vigorous a n d rapid decomposition of cellulose at 65° will take place in t h e presence of these organisms w h i c h are easily obtained from h i g h l y m a n u r e d soils. T h u s , i n the presence of a thermophilic organism, 4 2 g m of cellulose gave rise to 21.6 g m of ace- tic acid, 10.3 g m of ethyl alcohol, 11.9 g m of carbon dioxide, a n d a n u n - determined q u a n t i t y of hydrogen.
A fact of importance in the breakdown of carbohydrates in soils is t h a t association of organisms of different metabolic character, such as bacteria a n d fungi, m a y be m o r e effective t h a n the organisms taken separately. T h u s , a p u r e culture of a v a r i e t y of Aspergillus brought about a smaller r a t e of breakdown ( 2 8 % i n 48 days) of oat (Avena) straw cellulose t h a n w h e n it w a s present in a mixed culture ( 3 9 % decomposition in t h e same t i m e ) . A p p a r e n t l y , the presence of carbo- hydrates m o r e easily available t h a n cellulose, a n d p e r m i t t i n g the devel- opment of a vigorous flora, seems to pave t h e w a y for a n attack on cellulose ( 1 7 5 ) .
T h e n a t u r e of b r e a k d o w n of a n organic substance i n soil is clearly determined b y competitive, as well as b y adaptive a n d symbiotic, fac- tors in the soil population.
b. Hemicelluloses and polyuronic acids. T h i s group contains hexosans, as m a n n a n s a n d galactans, pentosans of formula (CsHsO*)^ pectins, a n d polyuronides. T h e y a r e present in considerable a m o u n t s in higher
6 . M I C R O B I A L A C T I V I T I E S O F S O I L A N D P L A N T N U T R I T I O N 6 8 3
green plants w h e n c e t h e y find their w a y to t h e soil. T h e largest group of t h e hemicelluloses a r e t h e pentosans w h i c h m a y b e present to t h e extent of 7% in p i n e needles a n d 3 2 % i n corn cobs. Precise knowledge of the constitution of t h e ill-defined polyuronide hemicelluloses is prac- tically nonexistent o w i n g to t h e difficulty of isolating homogeneous products. I n m a n y cases t h e u r o n i c acid constituent has not been iden- tified w i t h certainty.
Polyuronides a n d u r o n i c groupings a r e widely distributed i n soils a n d their content has a direct relation to soil fertility, as t h e y contrib- u t e greatly to t h e w a t e r retention a n d c r u m b structure of soil. Such polyuronides a r e pectic acid a n d alginic acid as well as a v a r i e t y of bacterial polysaccharides. Pectic acid, w h i c h is t h e m a i n constituent of t h e pectins, is composed of D-galactopyruronic acid units m u t u a l l y united b y 1,4-a-linkages. Alginic acid, on t h e other h a n d , is represented as a chain of D- m a n n o p y r u r o n i c acid residues joined b y l,4-/?-linkages.
A polyuronide representing t h e h i g h l y i m p o r t a n t group of specific bacterial polysaccharides, w h i c h m u s t form a significant proportion of soil carbohydrates, is built of cellobiuronic acid residues joined b y
^-linkages i n t h e 1 , 3 position. These yield glucose a n d glucuronic acids on hydrolysis.
M a n y mucilaginous polysaccharides contain uronic acid units ( 9 4 a ) . Seed mucilages of t h e plantains m a y h a v e a high proportion of xylopyranose end groups a n d a smaller proportion of galactopyranose end groups. T h e work of H i r s t a n d of others (e.g., 94b, 221a, 44a) has given information on the complex polyuronides present i n g u m arabic a n d other p l a n t g u m s t h a t m a y be found i n soils t h a t contain decaying vegetable m a t t e r . T h e r e seems to be little doubt t h a t t h e metabolism of both microorganisms a n d plants contributes greatly to the store of polyuronic acids in soil.
T h e products of hydrolysis of t h e hemicellulose of Iceland moss (Cetraria islandica) gave rise to glucose, m a n n o s e , galactose, a n d glu- curonic acids.
Mucilaginous polysaccharides contain u r o n i c acid units, a n d such compounds h a v e v e r y complex structures. T h e mucilage of t h e seed of t h e p l a n t a i n h a s a high proportion of xylopyranose e n d groups, galactopyranose units, galactouronic acid, a n d a m e t h y l pentose, as yet unidentified. Glucuronic acid is incorporated w i t h galactopyranose units i n g u m arabic, a n d p l a n t g u m s contain structures of this kind associated w i t h rhamnofuranoses, arabofuranoses, etc.
Hemicelluloses, especially t h e pentosans or xylans, a r e readily de- composed b y soil fungi, actinomycetes, a n d bacteria. T h e decomposition m a y take place m o r e r a p i d l y t h a n t h a t of cellulose but, as has been
684 J. H . QUASTEL
pointed out b y W a k s m a n a n d Starkey ( 2 6 0 a ) , t h e organic m a t t e r in certain peats a n d m i n e r a l soils m a y be free from cellulose u n d e r condi- tions w h e r e t h e r e is still m u c h hemicellulose present. T h i s is due to t h e greater resistance of certain hemicelluloses to decomposition a n d t h r o u g h t h e synthetic activities of a n u m b e r of microorganisms t h a t give rise to gums a n d slimes.
Hydrolysis of this group of substances m a y take place b y h e m i - cellulases w h i c h a r e widely distributed as extracellular e n z y m e s of microorganisms, a n d in seeds a n d in the digestive systems of inverte- brates. F u n g i , such as Aspergillus oryzae, are rich in such hydrolytic e n z y m e s w h i c h exert their optimal activity at p H 4 - 5 , b u t fungal hydrolysis of t h e xylans is r a r e l y complete. M a n n o g a l a c t a n s , found in soil t h r o u g h the breakdown of p l a n t seeds, are hydrolyzed to m a n n o s e or galactose b y fungi or b y snail enzymes w h e r e a s m a n n a n is hydro- lyzed to m a n n o s e b y various anaerobic bacteria.
T h e pectic acids, w h i c h consist of colloidal polygalacturonic acids, undergo hydrolysis b y enzymes existing in plants a n d microorganisms.
Pectinase, n o w k n o w n as pectin polygalacturonase, causes a fission of glycosidic linkages between neighboring anhydrogalacturonic acid units. A n o t h e r e n z y m e , once k n o w n as pectase, b u t n o w k n o w n as pectin methylesterase or pectin esterase, accelerates t h e removal of ester groups in pectinic acids. T h e s e e n z y m e s are present in bacteria a n d in molds a n d are of obvious i m p o r t a n c e in controlling t h e initial breakdown of t h e pectins in soil. A n o t h e r e n z y m e , provisionally t e r m e d a depolymerase, exists in yeasts a n d plants a n d p r e s u m a b l y plays its p a r t in soil metabolism. This e n z y m e hydrolyzes polygalacturonic acids of larger molecular weight only, a n d t h e hydrolysis does not proceed beyond t h e point w h e r e the degree of polymerization is about 5.
c. Estimation of polyuronic acids. Methods of estimation depend on t h e fact, first shown b y Lefevre a n d Tollens ( 1 3 1 ) , t h a t prolonged boiling w i t h 1 2 % H C l liberates carbon dioxide from u r o n i c acids.
Bartholomew a n d N o r m a n (17) h a v e worked out a m e t h o d of esti- m a t i o n of uronic acids in soils based on this observation. Some of their results a r e given in T a b l e II. T h e r e is some error d u e to C 02 evolution from hexosan groups.
T h e polyuronides, whose content i n soil seems to h a v e a direct relation to t h e fertility of soil, contribute greatly to the w a t e r retention or c r u m b structure of soil. Quastel a n d W e b l e y ( 2 0 3 ) , using a m a n o - m e t r i c technique, h a v e shown t h a t the addition of sodium alginate to a soil greatly improves its structure as shown b y increased c r u m b stability at h i g h w a t e r contents. I t was suggested t h a t alginate confers its soil-
6. MICROBIAL ACTIVITIES OF SOIL A N D P L A N T NUTRITION 685
T A B L E II
URONIC CONTENT OF VARIOUS SOILS EXPRESSED AS CARBON DIOXIDE YIELD0
C 02 yield per 100 gm Depth air dry cell Soil type (inches) (gm) Edina silt loam 0-4 0.082; 0.084 Edina silt loam 16-19 0.016; 0.016 Weiler silt loam
o-m
0.114; 0.118 Weiler silt loam 10-13 0.028; 0.030 Ames very fine sandy loam0-2M
0.232; 0.238 Webster silty clay loam 0-6 0.162; 0.162 Leon fine sand o - iy
2 0.108; 0.110Leon fine sand 0.019; 0.021
aD a t a from Bartholomew and Norman (17).
T h e relationship between cellulase a n d lichenase is not clear; t h e y seem to be different b u t further w o r k on this question is required.
e. Polymeric conversion of sucrose. A m o n g the sugars t h a t enter the soil a n d a r e swiftly metabolized, sucrose has special importance not only because of t h e quantities concerned, b u t because of the special re- actions in w h i c h it is e n z y m a t i c a l l y involved.
Sucrose, like other disaccharides, is r a p i d l y hydrolyzed to t h e con- stituent hexoses b y m a n y soil microorganisms, a n d thenceforward the metabolism of t h e sugars follows t h e familiar lines that occur in aerobic a n d anaerobic bacteria or fungi. Sucrose, however, gives rise to two products of considerable i m p o r t a n c e i n soil metabolism. T h e y , together w i t h t h e polyuronides, greatly affect soil structure a n d stability. These products are t h e polymers, dextrans a n d levans.
W h e n sucrose is perfused t h r o u g h a n e u t r a l soil u n d e r aerobic con- ditions, a considerable portion of it is converted to a levan polysac- charide. T h i s reaction is not brought about b y glucose or fructose or b y
improving properties b y its combination as a n ion w i t h one or m o r e constituents of the soil particles.
d. Lichenin. This carbohydrate, of Iceland moss a n d other plants, is broken down b y lichenases found in m a n y fungi a n d certain bacteria
(Cytophaga) a n d i n m a n y invertebrates. T h e distribution of these enzymes is similar to t h a t of cellulose. T h e y are extracellular, dif- fusible enzymes t h a t m a y be extracted directly from the organisms used as sources; t h e y m a y be separated from cellulase a n d other e n z y m e s b y fractional precipitation of aqueous solutions b y ethanol a n d ethyl ether. Lichenase hydrolyzes lichenin quantitatively to d-glucose
686 J. H . QUASTEL
a m i x t u r e of these monosaccharides. T h e synthesis of t h e levan is se- cured b y specific microorganisms of soil, e.g., Bacillus subtilis. A sol- uble cell-free e n z y m e w h i c h brings about levan production from su- crose has been isolated from B. subtilis a n d Aerobacter levanicum b y H e s t r i n a n d his colleagues ( 9 1 ) . On t h e other h a n d , cells of Leuco- nostoc mesenteroides a n d related organisms h a v e yielded a soluble en- z y m e , forming d e x t r a n from sucrose. I n both cases t h e polymerization of one component of t h e sucrose molecule is accompanied b y a n accu- m u l a t i o n of a n equivalent a m o u n t of t h e other component i n t h e form of free hexose, e.g., fructose i n dextran production a n d glucose in levan production.
N o r d a n d Vitucci (174) investigated the carbohydrate metabolism of the wood-destroying fungi, all of w h i c h a r e basidiomycetes. Acetic acid h a d been previously shown to be t h e last identifiable metabolite of these organisms, b u t N o r d a n d Vitucci, b y starting w i t h sodium ace- tate instead of cellulose, found t h a t t h e oxalic acid salt was the last prod- uct. F r o m results based on t r a p p i n g procedures a n d t h e use of iden- tified intermediates as sole carbon sources, t h e y concluded t h a t t h e two p a t h w a y s of degradation shown in Scheme A m a y take place w i t h t h e fungi u n d e r investigation.
cellulose glucose
\
ethanol
ι
acetic acid ,
ι
glycolic acid glyoxylic acid
ι
oxalic acid-* I
SCHEME A
F u r t h e r w o r k b y N o r d a n d Vitucci demonstrated t h a t although t h e over-all chemical result of t h e action of m a n y wood-destroying fungi
succinic fumaric
ι
malic
ι
6 . M I C R O B I A L A C T I V I T I E S O F S O I L A N D P L A N T N U T R I T I O N 6 8 7
is t h e same, t h e modes of metabolism of the organisms m a y differ con- siderably. Lentinus lepideus, one of the k n o w n root fungi, growing on wood or i n a glucose or xylose m e d i u m produces a n oily substance w h i c h w a s found to be a m i x t u r e of esters of c i n n a m i c acid a n d anisic acid. M e t h y l ^ - m e t h o x y c i n n a m a t e w a s also produced b y a m e c h a n i s m depending on t h e carbohydrate portion of the wood (Scheme B ) .
^-pentosan xylose
\
wood ethanol acetaldehyde
ι
t \
glucose CH=CH- COOCH«
t "
cellulose OCH 3
p -methoxymethy 1 cinnamate SCHEME Β
Soil h a s enormous potentialities for t h e digestion or b r e a k d o w n of organic m a t t e r . I t is u n l i k e l y t h a t a n y form of carbohydrate w o u l d long w i t h s t a n d t h e action of t h e soil organisms t h a t develop u n d e r soil conditions. T h e p r e l i m i n a r y steps i n t h e decomposition of complex car- bohydrates consist of hydrolysis to t h e simple sugars; a v a r i e t y of e n z y m e s accomplishing such changes are k n o w n .
B . S O I L AS A B I O L O G I C A L S Y S T E M
Soil m a y b e considered as a complex biological system i n w h i c h hosts of organisms a r e competing w i t h each other, often for a limited supply of nutritional material. T h e y exercise profound effects on each other's development a n d chemical activities a n d establish between t h e m - selves a d y n a m i c equilibrium w h i c h is constantly c h a n g i n g w i t h changes i n t h e physical a n d chemical e n v i r o n m e n t of the soil.
Some conception of the enormous population of microorganisms i n soil is provided b y t h e fact t h a t t h e r e m a y be as m a n y as 5000 mil- lions of bacteria p e r g r a m of soil. T h i s corresponds to a weight of over 4 tons of bacterial substance p e r acre of soil. T h e n u m b e r s fluctuate
688 J. H . Q U A S T E L
greatly, depending on t h e availability of t h e food supply, moisture, aeration conditions, t e m p e r a t u r e , h y d r o g e n ion concentration of t h e soil, t h e types of plants growing i n t h e soil, etc. Results, given i n T a b l e III, a r e taken from the s t a n d a r d w o r k b y S. A. W a k s m a n , "Principles of Soil Microbiology" ( W i l l i a m s & W i l k i n s , Baltimore, M a r y l a n d ,
T A B L E III
NUMBER OF MICROORGANISMS I N SOIL I N RESPONSE το TREATMENTS0
Hydrogen ion Microorganisms found6
concentration of
Treatment of the soil the soil Bacteria Actinomycetes Fungi Unfertilized and unlimed 4.6 3000 1150 60
Lime only added 6 . 4 5410 2410 23
Salts and ammonium sulfate
added 4.1 2690 370 112
Salts, ammonium sulfate
and lime added 5 . 8 6990 2520 39
Stable manure and salts added 5 . 4 8800 2920 73
"From S. A . Waksman (see text).
6 In thousands per gram of soil, determined by plate method.
T A B L E IV
NUMBER OF MICROORGANISMS I N SOIL AS AFFECTED BY PROXIMITY TO ROOTS"
Microorganisms found6
Sample of soil
Plant taken Bacteria Actinomycetes Fungi
Rye Near roots 28,600 4,400 216
Away from roots 13,200 3,200 162
Sugar beet Near roots 57,800 15,000 222
Away from roots 32,100 12,200 176
Alfalfa Near roots 93,800 9,000 268
Away from roots 17,800 3,300 254
"From Starkey (229).
* In thousands per gram soil.
1932) ; t h e y show h o w t h e microorganisms of soil v a r y i n n u m b e r ac- cording to t h e t r e a t m e n t of t h e soil.
Results given i n T a b l e I V , t a k e n from Starkey ( 2 2 9 ) , show h o w t h e proportions of microorganisms i n soil v a r y according to their prox- i m i t y to plants.
T h e bacteria r a n g e i n n u m b e r , u s u a l l y from a few h u n d r e d thou-
6. M I C R O B I A L A C T I V I T I E S O F S O I L A N D P L A N T N U T R I T I O N 689
sand to several h u n d r e d million p e r g r a m of soil, though m a n y species do not develop u n d e r t h e o r d i n a r y plating technique. G r a y a n d T h o r n - ton (77) obtained direct counts of t h e order of 4000 millions bacteria i n a g r a m of m a n u r e d arable soil. Such figures w e r e over a h u n d r e d times greater t h a n those obtained b y a plating technique. Protozoa m a y reach figures of t h e order of o n e million p e r g r a m of soil (amoebae 280,000; flagellates 770,000; ciliates 1000—estimated on a n e u t r a l m a n u r e d arable soil at R o t h a m s t e d ) . Algae m a y exceed 100,000 p e r g r a m of soil. According to W a k s m a n a n d Starkey (260a) t h e n u m b e r s of actinomycetes r a n g e from a few t h o u s a n d to m a n y millions p e r g r a m of soil, a n d t h e n u m b e r s of fungi m a y exceed one h u n d r e d thousand p e r g r a m .
I n such a complex microbiological population t h e r e exist n u m e r o u s chemical interrelationships t h a t affect metabolic behavior a n d cell pro- liferation. Symbiotic associations take place a n d antibiotic developments occur. T h u s W a k s m a n a n d Woodruff (262) h a v e s h o w n that, b y t h e use of a proper extractant (e.g., e t h e r ) , it is possible to demon- strate t h a t soils contain substances of t h e actinomycin t y p e w h i c h in- hibit t h e g r o w t h of certain bacteria i n culture media. Such substances m a y be both bacteriostatic a n d bacteriocidal; their effects m a y be con- siderably modified b y soil constituents ( 2 5 9 ) .
Cell adaptation occurs a n d t h e e n z y m e e q u i p m e n t of t h e soil micro- organisms undergoes changes i n response to changes in their environ- m e n t . K l u y v e r (114) commented, i n a recent lecture, t h a t t h e a m o u n t of microbial protoplasm i n e a r t h surpasses t h a t of a n i m a l protoplasm b y almost t w e n t y times. I t is obvious t h a t in dealing w i t h t h e subject of metabolic activities of t h e soil w e a r e dealing w i t h the metabolism of vast n u m b e r s of living cells, w h i c h greatly exceed t h e n u m b e r of cells found i n animals.
Considerations such as these, together w i t h increasing knowledge of t h e p h e n o m e n a of cell adaptations as a function of the environment, m a k e it necessary to investigate soil as a n integral biological whole ( 1 9 2 ) . Its organic m a t t e r m a y be, according to the circumstances, largely t h a t of living organisms, similar i n m a n y f u n d a m e n t a l respects to t h a t of plants or of animals, or it m a y be largely t h a t of the ligno- protein-polyuronide system of complexes w i t h relatively little living m a t e r i a l present. T h e study of all such m a t e r i a l — l i v i n g a n d non- living—is proper to the topic of soil organic m a t t e r .
C . O R G A N I C M A T T E R A N D S O I L A E R A T I O N A N D S T R U C T U R E
I t is n o w well k n o w n t h a t organic m a t t e r plays a n essential role in securing i n soil t h e structure, or tilth, t h a t is conducive to high fertility.
690 J. H . QUASTEL
I n fact, m a i n t e n a n c e cf the supply of organic m a t t e r to arable soils is of t h e greatest practical importance, for productive capacity is largely dependent u p o n it.
Aggregating substances a r e u n i f o r m l y distributed in soil crumbs.
T h e y consist, according to Kroth a n d P a g e ( 1 1 8 ) , of polar organic sub- stances, resulting from the decomposition of fresh organic m a t t e r , a n d of a group of substances, including iron a n d a l u m i n u m oxides, fats, waxes, a n d resins, t h a t are less efficient as soil aggregators. Bacterial slimes a n d fungal mycelia, it is well k n o w n , p l a y a n i m p o r t a n t role in soil aggregation a n d resistance of soils to erosion ( 7 1 , 166).
Following w o r k (154) on t h e binding action of mycelia a n d organic m a t t e r on soil particles, M a r t i n (153) showed t h a t addition of cultures of Bacillus subtilis to soil produces aggregations of soil particles owing to t h e organic substances formed b y t h e growth of t h e organism from a molecule such as sucrose. H e demonstrated t h a t addition of bacterial polysaccharides, such as t h e levans a n d dextrans, will affect soil struc- t u r e to a greater degree t h a n addition of casein or of lignin. Geoghegan a n d Brian (68a, b ) h a v e found t h a t bacterial a n d other polysaccharides aggregate soils. T h e y reported t h a t bacterial levans a n d dextrans h a v e m a r k e d aggregating effects on soil particles, t h e levans being formed b y a wide v a r i e t y of aerobic organisms, a n d the dextrans being formed b y various species of Leuconostoc. Soil crumbs containing small q u a n - tities of levans a r e v e r y stable b u t a r e e v e n t u a l l y broken down b y bac- terial action. T h e aggregating action of t h e products derived from micro- organisms is m u c h greater t h a n t h e direct binding effect of t h e micro- bial cells themselves. As soil aggregators, dextrans produced from sucrose b y t h e metabolic action of Leuconostoc a r e a p p a r e n t l y superior to t h e levans derived from sucrose b y t h e action of B. subtilis. A poly- saccharide formed b y Rhizobium a n d containing 6 7 % glucose a n d 2 0 % uronic acid residues (44) is a less efficent soil aggregator t h a n either t h e levans or t h e dextrans. U n d o u b t e d l y , t h e structure of the polysaccharide or polyuronide fraction h a s a strong influence on the aggregating properties of soil. H y d r o g e n bonding is considered a major m e c h a n i s m w h e r e b y polysaccharides a r e b o u n d to the soil particle.
H a worth, P i n k a r d , a n d Stacey (89) h a v e shown t h a t poor soils w i t h a low total organic content h a v e o n l y traces of polysaccharides; those w i t h higher total organic content h a v e 0.5-1.5 g m polysaccharide per kilogram moisture-free soil a n d possess a greater moisture-retaining capacity. Polysaccharide fractions containing levans w e r e isolated from such soils. S w a b y (237) finds t h a t a variety of substances, in- cluding proteins a n d polyuronides, affect soil aggregation.
T h e r e is n o w a n extensive literature on t h e formation of soil organic
6 . M I C R O B I A L A C T I V I T I E S O F S O I L A N D P L A N T N U T R I T I O N 6 9 1
m a t t e r b y various cropping systems a n d incorporation of crop residues, a n d its bearing on soil structure a n d soil fertility. A brief s u m m a r y has been given r e c e n t l y ( 1 9 1 ) .
1. Soil Aggregation
T h e structure of a soil is largely determined b y its c r u m b or aggregate formation. T h i s greatly influences w a t e r m o v e m e n t , aeration, a n d heat transfer. M a n y of the biochemical a n d chemical changes t h a t affect soil fertility t a k e place at the large surface areas presented b y soil aggre- gates. Substances t h a t stabilize crumbs, or aid i n soil aggregation, are u n i f o r m l y distributed in the c r u m b s of soil of good structure.
a. Polyuronides, polysaccharides, and related substances as soil aggregators. Polyuronides a n d u r o n i c groupings a r e present a m o n g h u m u s constituents a n d these a r e w i d e l y distributed in plants, com- posts, a n d soils. T h e u r o n i c acid content varies w i t h the soil t y p e ( 1 7 ) , a n d is a p p a r e n t l y related to its fertility. T h e polyuronides present in- clude pectic acid, alginic acid, a n d various bacterial polysaccharides.
M a n y mucilaginous saccharides contain u r o n i c acid units, a n d it is k n o w n t h a t complex polyuronides compose t h e p l a n t g u m s found in
soils containing decaying vegetable m a t t e r .
T h e aggregating effect of lignins, or lignin-like substances, in h u m u s has been investigated b y McCalla ( 1 4 8 ) a n d Alderfer et al. ( 1 ) . S t r a w composts in various stages of decomposition aggregate soils. M a t e r i a l containing m o r e t h a n 1 1 % hemicellulose a n d 1 8 % cellulose increases c r u m b formation. It seems t h a t conditions favorable for microbial ac- tivity facilitate t h e i m p r o v i n g action of t h e organic m a t e r i a l on c r u m b formation, but t h e y also hasten t h e destruction of the formed aggregates.
T h e m a r k e d effect of a polyuronide, alginic acid, in increasing soil aeration b y improving c r u m b stability a n d a p p a r e n t w a t e r retention was first shown b y Quastel a n d W e b l e y ( 2 0 3 ) [see also Quastel ( 1 9 0 ) ] . Using a m a n o m e t r i c technique ( 5 4 ) t h a t measures directly t h e avail- ability of oxygen to living cells in soil, t h e y w e r e able to demonstrate t h a t t h e presence of alginates greatly affects the air-water relationship of soil. T h e a m o u n t of w a t e r t h a t m a y be added to a soil before the availability of oxygen to t h e soil organisms falls t h r o u g h waterlogging a n d aggregate b r e a k d o w n is m u c h increased b y t h e addition of sodium alginate. T h e addition of 1 % sodium alginate increases the aeration capacity of a poor soil to t h a t found for a fertile g a r d e n soil. T h e effect of t h e alginate r a p i d l y increases w i t h increase of concentration to a m a x i m u m . E v e n a g a r d e n soil w i t h a fairly high content of organic matter, showing good aeration at a high w a t e r content, is affected favorably b y addition of alginate.
692 J. H . QUASTEL
2. Effects of Cellulose Products on Soil Aeration
Using their m a n o m e t r i c method, Quastel a n d W e b l e y (203) showed t h a t not o n l y a polyuronide, such as alginate, is effective in improving soil structure, b u t t h a t cellulose esters, such as cellulose acetate, e t h y l cellulose, a n d c a r b o x y m e t h y l cellulose, will improve air-water rela- tionships of a soil. T h e s e substances a t concentrations of 1 % (per d r y weight of soil) exercise effects similar to those d u e to about 0 . 5 % sodium alginate. T h e results a r e consistent w i t h those of Felber a n d G a r d n e r (55, 5 6 ) , w h o found t h a t addition of m e t h y l cellulose to soil secures considerable retention of moisture.
Addition of f a r m y a r d m a n u r e a n d horse d u n g greatly improves t h e air-water relationships a n d c r u m b stability of a soil. Quastel a n d W e b - ley (203) point out t h a t t h e effects of addition of these substances are twofold: (a) physical, d u e to the presence of fine s t r a w w h i c h has a binding effect on t h e soil c r u m b s — t h e effect h e r e is reversible, as wash- ing t h e soil results i n removal of t h e straw a n d the soil reverts largely to its original condition; (b) chemical, or physicochemical, due probably to the presence of polyuronides, or polysaccharides, affecting the soil particles—here t h e effect is irreversible as w a s h i n g t h e soil causes n o i m m e d i a t e diminution in c r u m b stability.
Addition of sewage sludges, composts, a n d refuses to soil improves t h e air-water relationships, u s u a l l y in proportion to the a m o u n t s of organic m a t t e r present. Alkaline extracts of soils a n d peats, afcer subsequent neutralization, m a y also be effective.
D . SOIL IMPROVERS (CONDITIONERS)
T h e conclusion reached b y t h e w r i t e r i n 1945 as a result of exper- iments w i t h alginate, etc., w a s t h a t it is u n l i k e l y t h a t polyuronides, or polysaccharides, will be successful as soil improvers o n a w i d e scale, though t h e y m a y be of importance in glasshouses w h e r e i m m e d i a t e im- provements i n soil structure m a y be v e r y beneficial. F o r field work, especially, substances m u s t be sought t h a t a r e less vulnerable to attack in t h e soil t h a n t h e polyuronides a n d polysaccharides. T h e s e substances are metabolites of the cell, subject to b r e a k d o w n b y a variety of soil organisms. T h e i r speed of decomposition i n the soil, w i t h consequent breakdown of t h e crumbs stabilized b y such substances, is determined, a m o n g other factors, b y availability of nitrogenous compounds whose mobilization b y t h e organisms attacking t h e polyuronides, etc., would also be deleterious to t h e plant. Moreover, t h e relatively large a m o u n t s of polyuronide salts required for structure i m p r o v e m e n t ( 5 - 1 0 tons per acre) would cause h a r m f u l effects to t h e soil d u e to t h e release of large quantities of cations present i n t h e salts.
6. M I C R O B I A L A C T I V I T I E S O F S O I L A N D P L A N T N U T R I T I O N 693
It b e c a m e clear t h a t t h e ideal soil conditioner should be a substance h a v i n g perhaps a m e c h a n i s m similar to t h a t of t h e polyuronides or polysaccharides i n aggregating soil particles b u t w h i c h would u n d e r g o a relatively slow r a t e of destruction in soil. It should, like t h e n a t u r a l l y occurring soil conditioners, be devoid of toxic effects on p l a n t or a n i m a l . It should n o t interfere w i t h t h e soil microbiological equilibria, involv- ing, for example, the growth of nitrogen-fixing organisms. N o r should it inhibit processes of soil nitrification; on t h e contrary, b y i m p r o v i n g soil aeration, it m i g h t accelerate t h e m . It should not remove, at a n y r a t e irreversibly, t h e trace elements t h a t a r e i m p o r t a n t for p l a n t nutrition.
T h i s ideal has been achieved to a great extent b y the introduction of the synthetic polyelectrolytes. *
1. Effects of Synthetic Polyelectrolytes on Aggregation, Aeration, and Water Relationships of the Soil (191)
H e d r i c k a n d M o w r y (90) found t h a t o n l y certain water-soluble polymeric electrolytes of h i g h molecular weight a r e effective as soil aggregators at t h e v e r y low concentrations required b y practical consid- erations. One of t h e most active substances w a s m a d e b y t h e hydrol- ysis of polyacrylonitrile. Other polymers of n e a r l y related structure also showed great improving effects on soil aeration a n d soil aggrega- tion. T h e two materials on w h i c h most w o r k w a s reported w e r e the sodium salt of hydrolyzed polyacrylonitriles a n d another carboxylated p o l y m e r used as a partial calcium salt'. Both polymers a r e polyanions.
Hedrick a n d M o w r y showed t h a t these substances, at concentrations of 0 . 1 % (per d r y w e i g h t soil) give better aeration values, i n a s a n d y loam, t h a n a n y other materials tested (e.g., sodium alginate, sodium c a r b o x y m e t h y l cellulose, pectin, lignin) at 1 % . T h e aggregate stabil- i t y is greatly increased, t h e effect being observed w i t h m a n y different soils. V e r y pronounced i m p r o v e m e n t i n t h e workability of all soils treated w i t h t h e polymers takes place. T h e y a r e c r u m b l y a n d friable at high w a t e r contents. W h i l e t h e p r i m a r y effect of the polyelectro- lyte on t h e soil is on c r u m b stability, t h e percolation r a t e t h r o u g h treated soil often shows a hundredfold increase over t h e r a t e i n u n - treated soil. T h e moisture equivalent is stated to be improved, a n d evi- dence from experiments on the wilting of plants indicates t h a t all the increase i n w a t e r held b y t h e soil is available for p l a n t growth. I n ad- dition to t h e m o r e r a p i d infiltration a n d percolation of water, a n d in- creased m o i s t u r e storage, a M i a m i silt l o a m treated w i t h t h e synthetic
* By the Monsanto Chemical Company, with whose scientific staff the writer was associated in the search for a synthetic soil improver.
694 J. H . Q U A S T E L
conditioners showed decreased surface evaporation, a n effect appar- ently due to t h e increased stability of t h e soil aggregates.
Allison (3) found from both laboratory a n d field studies t h a t alkali soils from t h e western U n i t e d States, w h i c h a r e low in permeability, on t r e a t m e n t w i t h t h e polymer,* give m a r k e d increases in permeability in proportion to t h e r a t e of t r e a t m e n t . H i g h water-stable aggregation in several saline a n d alkali soils is obtained after application of the conditioners at rates of 0.025 a n d 0 . 1 % . T h e aggregates prove to be w a t e r stable, t h e structural i m p r o v e m e n t persisting t h r o u g h a second growing season ( 1 5 7 ) .
Vallance (247) finds t h a t t r e a t m e n t of poorly aggregated soil w i t h Krilium, at rates of 0 . 0 1 5 - 0 . 0 7 5 % , increases water-stable aggregates u p to 5 3 - 7 0 % , b u t there is little effect in well-aggregated ( 5 5 % ) soils.
It cannot be overemphasized t h a t t h e soil conditioners, such as poly- uronides or synthetic polyelectrolytes, stabilize soil crumbs, a n d that, for their effective use, soils m u s t be worked into a good structure before, or at t h e t i m e of, application of t h e conditioners.
K r i l i u m produces a large increase in permeability of all soils, b u t the increases v a r y greatly w i t h the different soils. Swanson (239) has also shown t h a t K r i l i u m administration improves aggregation, porosity, a n d permeability a n d prevents slaking; it stabilizes existing structure and facilitates drying. I m p r o v e m e n t s of soil structure of a cultivated loess b y K r i l i u m application h a v e been noted, b u t t h e r e is a p p a r e n t l y no improving effect on the surface l a y e r of a n acid a n d degraded forest soil. Increases of porosity a n d permeabilities of Ohio clays a n d silts after application of K r i l i u m h a v e been reported b y M a r t i n a n d Volk ( 1 5 8 ) . It appears t h a t t h e a m o u n t of clay in t h e soil is a dominating factor in determining the aggregating power of t h e polyelectrolyte. As deter- m i n e d b y permeability m e a s u r e m e n t s , t h e polyelectrolyte t r e a t m e n t , at both 0.025 a n d 0.100% levels, seems to overcome the dispersive effect of a high content of exchangeable sodium ( 3 ) . I n a comparison of aggregating effects of the K r i l i u m polymers on n i n e western soils, v a r y - ing widely in a n u m b e r of chemical a n d physical characteristics, it was found t h a t regardless of p H , salinity, exchangeable-sodium percentage, or clay content of t h e various soils tested, t h e r e seems to be no signifi- cant difference between t h e abilities of t h e p o l y m e r conditioners to produce water-stable aggregates.
Administration of t h e polyelectrolyte to alkaline a n d saline soils facilitates removal of salt a n d exchangeable sodium after soil irrigation, a property of considerable importance in soil reclamation.
* Now known as Krilium, which is the trademark of the Monsanto Chemical Com- pany for substances such as hydrolyzed polyacrylonitrile sold as soil conditioners.