Effects of relatedness on social behaviour in house sparrows
Zoltán Tóth
Supervisor:
Dr. András Liker
Associate professor
Department of Limnology, University of Pannonia
Doctoral School of Chemistry and Environmental Sciences University of Pannonia
2010 Veszprém
ROKONSÁG HATÁSA A SZOCIÁLIS VISELKEDÉSRE HÁZI VEREBEKNÉL
Írta:
Tóth Zoltán
Készült a Pannon Egyetem Kémiai és Környezettudományi Doktori Iskolája keretében Témavezető: Dr. Liker András
Elfogadásra javaslom (igen / nem)
……….
A jelölt a doktori szigorlaton …... % -ot ért el,
Az értekezést bírálóként elfogadásra javaslom:
Bíráló neve: …... …... igen /nem
……….
Bíráló neve: …... …... igen /nem
……….
A jelölt az értekezés nyilvános vitáján …...% - ot ért el.
Veszprém, ……….
a Bíráló Bizottság elnöke A doktori (PhD) oklevél minősítése…...
………
Az EDT elnöke
“There is no end to our researches. Our end is in the other world.
It is a sign of contraction of the mind when it is content; or of lassitude.
No noble spirit stays within itself; it ever aspires and rises above its strength.
It scars beyond its deeds; if it does not advance and does not press forward, and does not back and does not clash within itself, it is only half alive.
Its pursuits are boundless and formless; its food is wonder, the chase, ambiguity.”
/Michel de Montaigne/
C
ONTENTSABSTRACTS...6
I.INTRODUCTION...9
1. Altruism and other forms of social behaviour...9
2. Hamilton’s idea of inclusive fitness ...10
3. Interpretation of inclusive fitness theory ...11
4. Revised versions of the Hamilton’s rule ...13
II.THESIS OBJECTIVES...15
III.GENERAL METHODS...18
1. Model species ...18
2. Study subjects...19
a) The studied house sparrow population...19
b) Free-living wintering flocks...19
c) Captive house sparrow flocks...20
3. Genetic analyses and kinship estimation ...23
IV.STUDY 1:GENETIC RELATEDNESS IN WINTERING GROUPS OF HOUSE SPARROWS (PASSER DOMESTICUS)...26
V.STUDY 2:EFFECTS OF RELATEDNESS ON SOCIAL-FORAGING TACTIC USE IN HOUSE SPARROWS ...39
VI.STUDY 3:KINSHIP AND AGGRESSION: DO HOUSE SPARROWS SPARE THEIR RELATIVES?...49
VII.STUDY 4:WHOM DO THE SPARROWS FOLLOW?THE EFFECT OF KINSHIP ON SOCIAL PREFERENCE IN HOUSE SPARROW FLOCKS...58
VIII.STUDY 5:INVESTIGATION OF SOCIAL NETWORK POSITIONS IN CAPTIVE HOUSE SPARROW FLOCKS...66
IX.CONCLUSIONS...78
X.SUMMARY...81
XI.REFERENCES...83
XII.ACKNOWLEDGMENT...97
XIII.THESIS POINTS...98
XIV.LIST OF PUBLICATIONS...102
XV.APPENDIX...104
A
BSTRACTThe author used the general framework of Hamilton’s (1964) inclusive fitness theory to address several questions about how genetic relatedness among individuals may affect different aspects of social behaviour in a relatively simple social system, the wintering flocks of house sparrows (Passer domesticus). For this purpose, intense ringing and observation of wild and captive birds, in combination with pedigree and molecular genetic data on relatedness were used to investigate (i) the kinship structure of free-ranging flocks and (ii) various social interactions among differently related group-mates in captive flocks. Although aggregation of related individuals was not found in the wild, most house sparrows had at least a few close relatives in their feeding groups, which may provide opportunity for the evolution of kin-biased behaviours in natural circumstances. In accordance with that, in the captive flocks sparrows exploited their close kin and non-kin flock-mates differently by scrounging (i.e. feeding from food found by others) during social foraging and preferred to follow their same-brood siblings during various social activities. On the other hand, sparrows did not mitigate their aggression towards relatives in any aspect of agonistic behaviours, and kinship did not influence individuals’ centrality (i.e. pattern of connections to flock-mates) in a non- random network constructed from within-group followings. Although indirect fitness effects were not measured in our studies to prove undoubtedly the operation of kin selection, this thesis provides new evidence for the presence of kin-biased social behaviours in a species that neither breed cooperatively nor live in permanent family groups.
A
BSTRACTDer Autor untersuchte die Beziehung zwischen genetischer Verwandtschaft und sozialen Verhaltensweisen in sozial relativ einfach organisierten Wintertrupps von Haussperlingen (Passer domesticus). Die Ergebnisse beruhen auf Volieren- und Freilandbeobachtungen. Um festzustellen welche Rolle Verwandschaft bei der sozialen Organisation innerhalb einer Gruppe spielt, benutzte er Pedigree-Daten und genetische Methoden um den Verwandtschaftsgrad zwischen den Individuen zu bestimmen. Außerdem wurde (i) die Verteilung verwandter Tiere innerhalb freilebender Wintergruppen festgestellt, und (ii) in Volieren, welche Verhaltensweisen zwischen Tieren mit verschiedenem Verwandschaftsgrad auftreten. Die Ergebnisse zeigten dass, in allen freilebenden Gruppen die meisten Individuen kontakt zu mindestens ein paar nahe verwandten Individuuen hatten. In den Volieren benutzten die Tiere die „Schmarotzer”-Taktik seltener gegenüber verwandten als nicht verwandten Individuen und bildteten bevorzugt Geschwistergruppen. Die in der Dissertation beschriebenen Untersuchungen liefern neue Ergebnisse über die Bedeutung von Verwandschaft für soziale Interaktionen in einer Art, für die weder kooperatives Brüten, noch stabile Familiengruppen typisch sind.
K
IVONATA szerző Hamilton (1964) inkluzív fitnesz elméletéből kiindulva vizsgálta, hogy az egyedek közötti genetikai rokonság miképpen befolyásolja a szociális viselkedés különböző aspektusait egy viszonylag egyszerű szociális rendszerben, a házi verebek (Passer domesticus) téli csapataiban. Ennek érdekében szabadon élő és fogságban tartott madarak megfigyelésével, illetve pedigré információk alapján és molekuláris genetikai módszerekkel végzett rokonságbecslés segítségével elemezte (i) a szabadon élő verébcsapatokban a rokon egyedek előfordulását, illetve (ii) fogságban tartott csapatokban azt, hogy a különböző fokú rokonságban álló csapattársak miként viselkednek egymással szociális interakcióik során. A vizsgálatok alapján a vadon élő csapatokban a rokon egyedek gyakori együttes előfordulása nem jellemző, azonban a legtöbb egyednek volt legalább kisszámú közeli rokona a táplálkozó csapatokban, ami lehetőséget teremt rokon-megkülönböztető viselkedésformák kialakulására természetes körülmények között. Ezzel összhangban, a fogságban tartott madarak bizonyos szociális viselkedéseik során megkülönböztették rokonaikat többi csapattársuktól: különböző mértékben használták ki az un. „potyázó” taktika révén a rokon, illetve nem-rokon társak által talált táplálékfoltokat csapatos táplálkozáskor, valamint szívesebben követték azonos fészekaljból származó testvéreiket a különböző szociális aktivitások során. Ugyanakkor az egyedek nem különböztették meg rokonaikat az agresszív interakciók alkalmával, és a közöttük lévő rokonság azt sem befolyásolta, hogy az egyes madarak mennyire központi szerepet töltenek be (milyen a társakhoz fűződő kapcsolataik mintázata) egy, a csapaton belüli követésekből modellezett hálózatban. Habár nem bizonyítható egyértelműen, hogy a megfigyelt rokon megkülönböztető viselkedések kizárólag indirekt fitnesz következményekkel járnak (azaz kizárólag a rokonszelekció következményei), a disszertációban bemutatott vizsgálatok új eredményeket szolgáltatnak az egyedek közötti rokonság szociális viselkedést befolyásoló hatására egy olyan faj esetében, amelyre sem a kooperatív költés, sem a tartós családi csoportok nem jellemzőek.
I. I
NTRODUCTION1. Altruism and other forms of social behaviour
All animals have to face challenges during their life and make decisions about how to cope with their varying social and physical environment. Similarly to other phenotypic traits, behaviour (i.e. collection of adopted decisions) is also under the optimizing pressure of natural selection and considered adaptive only if they maximize the performing individual’s fitness through the net benefits associated with their performance. In that sense, only that behaviour will be favoured by natural selection and will be continuously inherited in the genome of individual lineages, which is able to contribute efficiently to the performing individual’ success in terms of survival and/or reproduction (Danchin et al. 2008).
From an evolutionary point of view, a behaviour can be viewed as social if it has fitness consequences for both the actor, i.e. performer of given behaviour and the recipient, toward which the behaviour is directed. Different forms of social behaviour can be categorized according to whether they are beneficial (increase direct fitness) or costly (decrease direct fitness) for these individuals (West et al. 2007); direct fitness is defined as the probability of successful personal reproduction (Foster et al. 2006a). According to Hamilton’s (1964) classification, benefits and costs are calculated on the basis of lifetime consequences of the behaviour and net fitness effect. Following Hamilton’s terminology, behaviours that are beneficial to the actors and costly to the recipients (+/–) are selfish, whereas behaviours that are costly to the actors but beneficial to the recipients (–/+) are altruistic. Those behaviours which entail direct fitness benefits for both actors and recipients (+/+) are called mutually beneficial (West et al. 2006), while behaviours that has negative fitness consequences for both actor and recipient (–/–) are spiteful (Hamilton 1970).
For evolutionary biology, it has been a challenging problem to explain how altruistic behaviours being costly to perform but beneficial to other individuals could evolve and remain evolutionary stable in natural populations. Interestingly, different interpretations of altruism in the relevant literature often generated semantic confusion about this form of social behaviour (e.g. Wilson 2005, Wilson & Hölldobler 2005, Traulsen & Nowak 2006). For example, ‘reciprocal altruism’ (Trivers 1971), the alternate helping through repeated interactions between unrelated individuals cannot be viewed as altruistic in the above sense because it provides delayed direct fitness benefit for the performing individuals. Similarly, the
term ‘weak altruism’ introduced by Wilson (1975, 1977) does not describe solely altruistic acts as it can be applied by definition to behaviours that increase actors’ fitness directly.
Furthermore, terms like ‘cooperation’ or ‘helping’ can also be misleading sometimes as they are used to describe behaviours that are either beneficial (+/ ) or costly (–/ ) to the actors.
However, the precise distinction between mutually beneficial and altruistic behaviours is of great importance for an obvious reason. In contradistinction to mutually beneficial behaviours, altruistic behaviours do not entail direct fitness benefit for the actor (not even in the long term) that could outweigh the cost of performing the behaviour, thus altruism cannot be explained by the usual interpretation of the Darwinian natural selection (that restricts analyses to direct fitness costs and benefits; Sachs et al. 2004).
2. Hamilton’s theory of inclusive fitness
The concept of inclusive fitness introduced by Hamilton (1964) provided a revolutionary explanation for the evolution of altruistic behaviour by taking the indirect fitness benefits of behaviour into account. Hamilton suggested that the inclusive fitness consequence of an altruistic trait should include the trait’s contribution to the bearer’s fitness (direct fitness effect, see above) and also its contribution to the fitness of all individuals that possess the same genes and have been affected by the behaviour of the bearer (indirect fitness effects; Grafen 1984, Frank 1998, Danchin et al. 2008). For example, an individual that helps to raise its sibling’s offspring would suffer costs associated with the helping behaviour (as helping is likely to hinder its own reproduction), but also gains benefits from the reproductive success of its sibling as they are closely related due to common ancestry, and the helping individual also shares a certain amount of genes with the offspring to be raised. Applying the above-mentioned idea, Hamilton (1964, 1970) developed a mathematical theory to expand the Darwinian concept of natural selection from a theory that focused solely on individuals’ direct fitness to different one that takes account of social interactions, suggesting that natural selection leads to organisms acting as if they are maximizing their inclusive fitness (Grafen 2006). Hamilton summarized his idea known as Hamilton’s rule (Frank 1998), which can be expressed by the formula:
rxy×b-c>0 (1)
This equation involves three terms: c, the cost of direct fitness for the actor; b, the direct benefit for the recipient of the altruistic behaviour; and rxy, the degree of relatedness between the actor (x) and the recipient (y). According to this equation, we can expect altruistic behaviour to evolve when the benefit of the recipient multiplied by its relatedness with the actor exceeds the cost of the altruistic individual. Inclusive fitness theory, also known as kin selection after Maynard Smith (1964), was a major breakthrough and deeply influenced our understanding of social behaviour by providing a simple evolutionary explanation of altruistic traits: genes causing a fitness cost to their altruistic bearers can be rewarded if they contribute to enhance the replication of the same genes in related recipients (Dawkins 1976, Danchin et al. 2008).
The most common way of obtaining indirect fitness benefits is when altruistic behaviour is directed toward individuals who are genetically closely related to the actor due to common ancestry, e.g. an actor may help its close relatives’ reproduction since this way it can indirectly contribute to the passing of its own genes to the next generation. There are two mechanisms that can favour the evolution of this type of altruism (Hamilton 1964): kin recognition, when altruistic behaviour is preferentially directed toward relatives, and limited dispersal, which, by creating a neighbourhood of relatives, allows for acting altruistically toward all neighbours (West et al. 2007). Another way to obtain indirect fitness benefits is to behave altruistically only toward individuals who share the same altruistic genes as the actor.
This latter mechanism, coined as ‘green beard effect’ by Dawkins (1976), generates special assortment of altruistic individuals and requires a single gene (or a number of linked genes) that genetically codes both altruistic behaviour and some recognizable phenotypic trait, such as a “green beard” (Hamilton 1964, 1975, Dawkins 1976, Jansen & van Baalen 2006).
3. Interpretation of inclusive fitness theory
Kin selection theory has been the subject of much debate and misunderstanding, originating either from confused redefinition of terms (i.e. what kind of social interactions we consider altruistic), the misuse of related jargon (i.e. various interpretation of kin selection) or from the inappropriate application of Hamilton’s rule (for reviews see Frank 1998, West et al.
2002, 2007, Foster et al. 2006ab). Group selection, introduced by Wynne-Edwards (1962), was an alternative idea for the explanation of the evolution of altruism, emphasizing the importance of selection at group level, but was strongly criticized and refuted by many works
from theoretical point of view (Maynard Smith 1964, 1976, Grafen 1984, Frank 1986b). Ever since, new forms of group selection models have been developed (Wilson 1975, Wilson &
Colwell 1981, Wilson 2005, Wilson & Hölldobler 2005, Traulsen & Nowak 2006), arguing that kin selection theory provides an inappropriate explanation for the evolution of altruism and emphasizing the importance of multiple levels of selection acting on within-population groups (West et al. 2007). These works also state quite often that colony-level selection in insects (Arthropoda, Insecta) is unquestionably necessary to explain the evolution of eusociality (Wilson 2005, Fletcher et al. 2006), a phenomenon that could not be explained prior to Hamilton’s theory. However, the concept of different levels of selection is entirely consistent with the principle of inclusive fitness theory, i.e. organisms are expected to behave as if they were maximizing their inclusive fitness (Grafen 2006, Gardner et al. 2007). These two ideas (i.e. kin selection & colony-level selection) were shown to be mathematically identical and proved to be just different ways of conceptualizing the same evolutionary process (Grafen 1984, Frank 1986a, 1998, Taylor 1990, Queller 1992, Bourke & Franks 1995, Gardner et al. 2007). It was also shown that there is no theoretical or empirical example of group selection that cannot be explained by individual-level selection or kin selection (Lehmann et al. 2007, West et al. 2007). Inclusive fitness and kin selection theory proved to be useful for understanding specific biological cases, making testable predictions and also for constructing a general theoretical overview (Grafen 1984, Frank 1998, Queller 2004, West et al. 2008). In fact, inclusive fitness provided a unifying framework for all possible explanations of cooperation between interacting individuals, and became the central paradigm in social evolution theory (e.g. Frank 1998, Sachs et al. 2004, Foster et al. 2006b, Lehmann &
Keller 2006, West et al. 2007, 2008).
In the last decade, powerful methods for understanding the evolution of altruism have been developed by applying a new approach (Fletcher & Doebeli 2006, 2009, Taylor et al.
2007) that divides the direct fitness effects of altruism on individuals into two parts: one that is due to the self and the other that is due to the ‘interaction environment’. This methodology emphasizes the fundamental importance of the latter to explain altruistic behaviour and provides a more general concept compared to the inclusive fitness approach, enabling, for instance, also the analysis of interspecific mutualism (Fletcher et al. 2006). However, being mathematically equivalent, the results of these direct fitness analyses can also be interpreted in terms of inclusive fitness, which can be considered as a more natural way of understanding fitness consequences originating from the interactions of related individuals (Taylor et al.
4. Revised versions of the Hamilton’s rule
Inclusive fitness theory and Hamilton’s rule can be applied in any situation involving conflict or cooperation (Hamilton 1964, Treisman 1977). However, it is often neglected that limited dispersal, the same mechanism that favours the evolution of altruistic behaviour toward relatives, may also lead to increased competition between related individuals, reducing or even completely removing the net advantage of altruism (West et al. 2002, Griffin & West 2002, Griffin et al. 2004). The importance and extent of how this kind of competition influences selective processes in nature have been demonstrated by a recent work on fig wasps (West et al. 2001). The level of fighting between fig wasp males has always been a classic example of how kin selection operates as less fighting was predicted to occur in species where competing males were highly related (Hamilton 1979, Trivers 1985). However, West et al. (2001) found that fighting level did not correlate with relatedness across the fig wasp species; in fact, competition between relatives was so intense that it possibly removed any advantage of being altruistic. Instead, fighting levels could be well explained by the direct benefit of winning: males fought more aggressively when there were fewer females to compete for within a fruit.
The effect of competition between relatives can be incorporated into the Hamilton’s rule in several ways (Grafen 1984, Queller 1994, Frank 1998, West et al. 2002). For instance, as West et al. (2002) pointed out, the influence of competition between relatives on kin selection can be investigated by extending Hamilton’s rule to include all individuals whose fitness is affected by an altruistic behaviour. This idea can be summed up in the following equation:
rxy×b-c-rxe×d>0, (2)
where rxy is the genetic relatedness between the altruist and the beneficiary of its altruism (i.e.
the standard r, see above), rxe is the altruist’s relatedness to the competitors of the beneficiary, and d is the general decrease in fitness of these competitors associated with the altruistic act.
If the altruist is unrelated to the competitors of the beneficiary (rxe=0) or the altruistic act leads to no increase in the general level of competition (d=0), then the classic equation for Hamilton’s rule holds (West et al. 2002). As the altruist becomes more related to the
competitors of the beneficiary (increasing rxe, for example, when competition becomes more local) and/or the altruistic act increases the general level of competition (increasing d), the advantage of being altruistic is reduced. In the extreme, if an individual equally related to both the beneficiary and its competitors, then altruism cannot be favoured, irrespective of the values of b and c (West et al. 2002). Following another concept, Queller’s (1994) model incorporates competition between relatives into the r term of Hamilton’s rule. He showed that standard estimations of relatedness usually overestimate the importance of kin selection and relatedness should be measured at the correct scale, i.e. relatedness to the beneficiary of the altruism has to be measured with respect to the individuals with which the beneficiary will compete (defined here as rc), rather than with respect to the global population (rxy). If competition between relatives does occur (rxe>0), then the effective relatedness will be lower (rc<rxy).
All in all, recent refinements of Hamilton’s original rule provided sufficient conceptual solutions to allow for the fact that limited dispersal may induce both high relatedness and increased competition between interacting individuals, and showed that kin selection theory and Hamilton’s rule are correct, but care must be taken when applying them to a specific social system (Griffin & West 2002, West et al. 2002, Griffin et al. 2004).
II. T
HESIS OBJECTIVESI used the above general framework of inclusive fitness theory to address several questions about how relatedness (more precisely, genetic relatedness due to common ancestry) among individuals may affect different aspects of social behaviour in a relatively simple social system. I aimed to investigate whether individuals of a gregarious bird species discriminate their kin during social interactions in the non-reproductive period, and if they do, such behaviours can be linked to indirect fitness benefits or not. In this thesis, therefore, I looked into five main issues to explore the possible relationship between genetic relatedness and social behaviour in wintering flocks of house sparrows (Passer domesticus). All of the studies presented here were carried out by the Ornithological Group at the University of Pannonia. For all studies I participated in most stages of the work, from bird ringing, behavioural observations through data analyses until the preparations of related manuscripts.
Study 1. What is the kin composition of wintering house sparrow flocks like?
To explore the relationship between genetic relatedness and social behaviour in the species, it is essential to gain adequate information about the kin composition of wintering sparrow flocks, i.e. whether house sparrows have the possibility to interact with related group- mates in natural circumstances. For this purpose, we used intense ringing and observation of wild birds, in combination with molecular genetic data on relatedness and investigated whether feeding aggregations of house sparrows include also aggregations of related individuals (Chapter IV).
Study 2. Do house sparrows discriminate their kin during social foraging?
An important, well studied type of social interactions of sparrows occurs during social foraging, when individuals use alternative behavioural tactics (“producer” and “scrounger”
strategies) to maximize their food intake while adjusting their behaviour to various ecological conditions. Hamilton’s rule predicts kin-discriminative behaviour during social foraging if either the cost of being scrounged or the benefit of the scrounger is high; in the former case reduced scrounging from kin, whereas in the latter case increased scrounging from kin can be expected. To test whether relatedness affects foraging tactic use in house sparrows, we
differently related individuals. Specifically, we tested whether sparrows use the aggressive and non-aggressive form of scrounging tactic at a different rate and with different success against closely related and unrelated flock-mates (Chapter V).
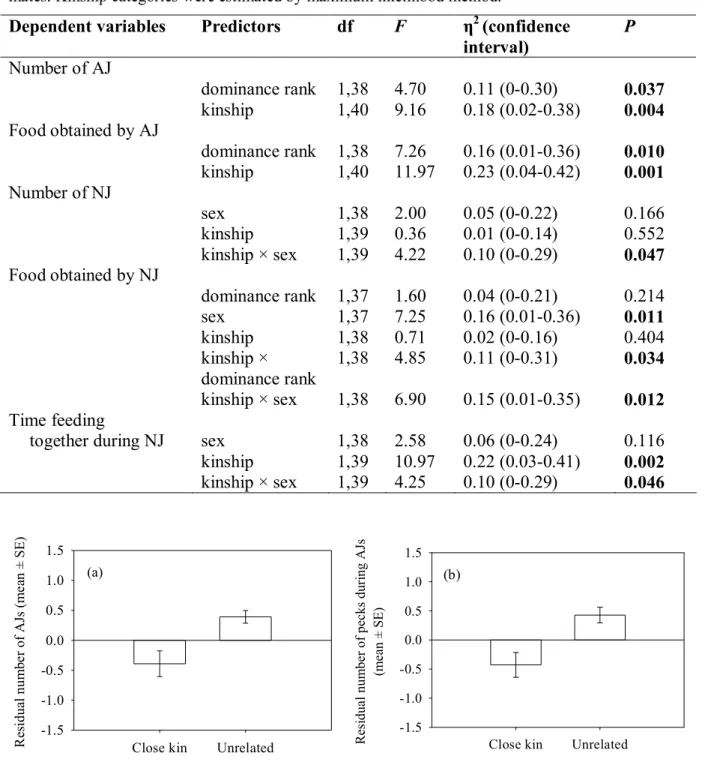
Study 3. How does relatedness influence aggressive interactions in the sparrow flocks?
Inclusive fitness theory can be applied in any situation involving also conflict and competition, in which case it predicts restricted level of aggression between related individuals (at least by its original form, i.e. without local kin competition). To test whether genetic relatedness affects agonistic interactions between house sparrows, we observed captive flocks in which birds could interact with differently related individuals. Specifically, we tested whether sparrows show reduced aggression against kin compared to unrelated flock-mates, and also whether the presence of relatives influences their success, as measured by the achieved rank of birds in dominance hierarchy (Chapter VI).
Study 4. Are kin group-mates more associated with each other in the flocks than unrelated birds?
Socio-positive interactions (such as joining others in non-aggressive activities) are often entailed by important fitness consequences to the participants and genetic relatedness among group-members has been found to affect such affiliative behaviour in various vertebrate species. To test whether kinship affects social preference in house sparrows, we observed captive flocks in which birds could join differently related individuals in different social activities. Particularly, we investigated whether preference between sparrow flock- mates increases with genetic relatedness or familiarity from early developmental period influencing affiliation between flock-mates (Chapter VII).
Study 5. Does the presence of kin affect network position of individuals in the flocks?
In groups of interacting individuals there are often differences between group- members in the number of their social interactions and how central or peripheral they are within the social network (in terms of connectivity to others). To investigate the effect of relatedness on the network positions of house sparrows in their flocks, we used within-group
significantly different from equivalent random networks, and (ii) if they are, how kinship among group-mates affects individuals’ centrality in the network (Chapter VIII).
III. G
ENERALM
ETHODS1. Model species
The house sparrow (Passer domesticus) is the member of the Old World sparrow family Passeridae, and despite the recent population declines in Western Europe, is one of the most widely distributed bird species all over the world. Sparrows live in rural and urban habitats close to humans and are well known for their gregariousness and adaptive capabilities (Anderson 2006). For several reasons, this species seemed to be an ideal subject for the study of kin-biased social behaviours in a non-reproductive context.
Firstly, house sparrows are characterized by sedentary behaviour: after settling at a breeding site, adult birds may live within a small area throughout the year (Summers-Smith 1963, Perrins 1998, Anderson 2006). Natal dispersal is also limited in the species, although significant variation is found among populations and the reported frequency of dispersal may depend on the spatial scale on which it is studied. For example in a meta-population living in an archipelago, only 9.6% of female and 5.7% of male offspring dispersed from their natal island before their first breeding (Altwegg et al. 2000). On the other hand, Fleischer et al.
(1984) reported much higher frequencies of dispersal (52% of females, 27% of males) among neighbouring (usually within 1 km) colonies breeding in a farmland area. However, ringing data suggest that the majority of young sparrows settle close (usually < 2 km) to their natal area (Anderson 2006) and a significant proportion may remain in their natal colony. Such limited movements could result in most sparrows staying with their relatives (e.g. siblings or parents) in the non-breeding flocks at feeding and roosting sites around the breeding colonies.
Secondly, sparrows are highly social, found in flocks at and around their feeding and roosting sites throughout the non-breeding season. These flocks usually consist of ten to 30 or more individuals and flock-members perform various activities together such as foraging, roosting, and dust bathing (Anderson 2006). Flock-members also form social hierarchy and gain social status through aggressive interactions, competing frequently for resources with each other, particularly for food and roosting sites (e.g. Møller 1987, Liker & Barta 2001).
Because sparrows perform such wide variety of interactions, their wintering flocks are frequently used as model systems in studies of social behaviour, especially aggression and status signalling (Møller 1987, Liker & Barta 2001, Bókony et al. 2006; see Nakagawa et al.
2007 for a review) and problem solving in groups (Liker & Bókony 2009). The species is also
one of the best known producer–scrounger systems (Liker & Barta 2002, Lendvai et al. 2004, 2006), as sparrows usually feed in flocks and use both producer (actively searching for food) and scrounger (exploiting other’s food findings) tactics during foraging (Barnard & Sibly 1981, Johnson et al. 2001, Liker & Barta 2002). However, despite the intense research on the social behaviour of the species, we essentially lack adequate information about the kinship structure of sparrow flocks and how relatedness impacts their social behaviour.
2. Study subjects
a) The studied house sparrow population
We have been studying the house sparrow population since 2004 in and around the territory of the Zoo of Veszprém, in Veszprém (47º05′ N, 17º54′ E), north-western Hungary, where sparrows are resident all year-round. They breed in high numbers (>50 pairs) in the zoo, using both nest boxes and natural nesting sites, and for the whole zoo area the population size is estimated several hundreds in each autumn when young birds are present in large numbers (Bókony et al. 2008). For our study purposes, non-breeding adults were captured with mist-nets and were ringed at regular intervals (1 or 2 days weekly) during autumn and early winter (September – December) in each year, with some additional ringing of breeding adults during spring and summer. Breeding birds were also occasionally caught using a nest- trap fitted on nest boxes during the nestling feeding period. In addition to adults, we ringed 9- 11 days old nestlings both in nest boxes and at natural nest sites in the zoo throughout the years. All birds (including nestlings) were ringed with a numbered metal ring and three colour rings for individual identification. We also took a small amount of blood from each bird from the brachial vein and stored it in Queen’s lysis buffer (Dawson et al. 1998) until genetic analysis (see below).
b) Free-living wintering flocks
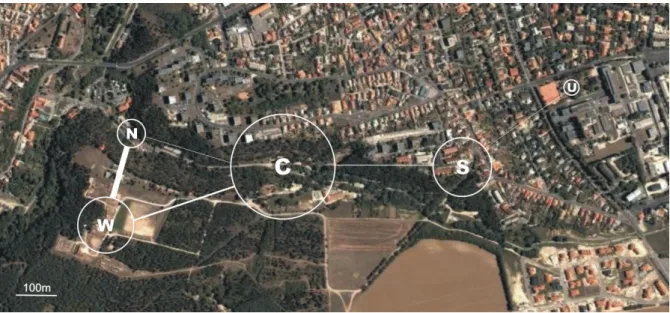
For the analyses of kinship structure (Study 1), fieldwork was conducted in 2004 – 2006, which included ringing and blood-sampling of free-ranging sparrows and their subsequent observations at five study sites to collect data on group composition and associations between individuals. Four of our five study sites were situated in and around the Zoo of Veszprém (northern site, N; western site, W; central site, C; and southern site, S),
whereas one additional site was on the nearby campus of the University of Pannonia (U), south-east from the zoo (Fig. III.1).
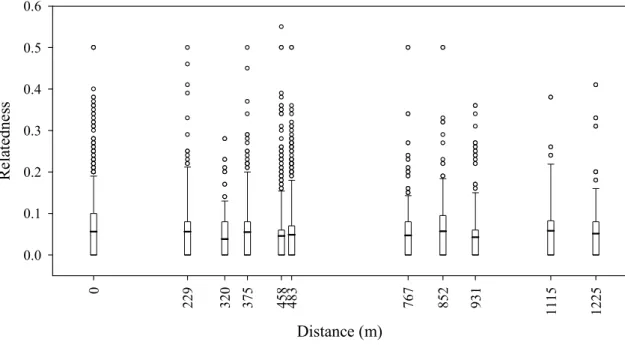
Figure III.1. Map of the study area. Study sites are indicated by the centre of the circles, and the diameter of the circles is proportional to core-flock size (number of ringed house sparrows observed exclusively at that site) in 2006. Lines connecting the circles indicate the movements of commuter birds (i.e. those moving between sites), line width is proportional to the number of commuters between the respective sites in 2006.
Each site had a group of shrubs where the sparrows regularly roosted during the day, and some adjacent open area where they could feed (see below). Distances between sites ranged between 229 – 1225 m (mean SE = 669 111 m), and they were separated from each other by forest tracks (typically mature Austrian pine Pinus nigra) or built-up areas (Fig.
III.1). In 2004 regular ringing was restricted to two zoo sites (W and C), whereas in 2005 all sites were involved. In 2006, we estimated the maximum number of birds present during the observations at each site, and median of these flock sizes were 21, 32, 100, 65, and 12 individuals for site N, W, C, S and U, respectively. In total, 410 house sparrows were ringed and blood-sampled before the start of the 2005 observation period, whereas the cumulative number of ringed individuals was 1244 by the start of the 2006 observation period (not all observed subsequently, see below).
c) Captive house sparrow flocks
The effect of relatedness on different aspects of social behaviour (Studies 2-5) was investigated on captive birds. We captured house sparrows with mist nets during autumn in 2005–2006 within the territory of the Zoo of Veszprém. Because we had monitored the breeding of ringed birds and had also ringed the nestlings (see above), we had pedigree information for many individuals by the time of capture. We allocated the captured birds into two flocks in 2005 and formed one flock in 2006, so that each contained same-brood siblings (Table III.1).
Table III.1. Characteristics of the captive house sparrow flocks.
Flock 1 Flock 2 Flock 3
Study period 2005 October -
December
2005 October- December
2006 October - November Number of individuals
Males Females
21 14 7
23 11
12
17 10 7 Number of same-brood siblings 9
(3 dyads, 1 triad)
10
(3 dyads, 1 tetrad)
10 (5 dyads)
Conducted studies Study 2-5 Study 2-5 Study 3-5
Number of scroungings (i.e.
exploitation of other’s food findings;
Study 2):
Flock total
Per individual (mean ± SE)
166
7.90 ± 1.72
438
19.04 ± 1.80 - - Number of agonistic interactions
(Study 3):
Flock total
Per individual (mean ± SE)
542
25.81 ± 3.95
788
34.26 ± 5.56
766
45.06 ± 6.89 Number of followings (Studies 4-5):
Flock total
Per individual (mean ± SE)
80
4.21 ± 0.49
138 6.0 ± 0.55
250
14.71 ± 1.19
All siblings and most other birds were juveniles of the year (49 out of the 61 individuals). Sex ratio of the flocks (Table III.1) was similar to those of free-living foraging flocks at our study site (41–63%, calculated from ringing data; see also Bókony et al. 2008) and those reported by Breitwisch & Hudak (1989). Adults were caught either in September after cessation of reproduction or, in a few cases, in June–July together with their offspring (and were held together in outdoor aviaries), so dependent youngs’ lives were not risked by our work. Upon capture we measured body mass (± 0.1 g), tarsus length (± 0.1 mm) and took small blood samples (approximately 100 μl) for kinship analyses. Each individual was ringed with a numbered metal ring and three colour rings. We also marked the captive birds by
painting small, coloured signs with nontoxic paint (using Deco painter, Marabu, Germany) on their crown feathers to facilitate quick individual recognition during the observations. Birds were held in outdoor aviaries (approximately 5(W) × 4(L) × 3(H) m) in the zoo (Fig III.2).
Figure III.2. One of the aviaries (with attached hiding-place for observation) where we held the studied house sparrow flocks in captivity (photo by Á.Z. Lendvai)
In 2005, flock 1 and 2 were held separately in two aviaries ca. 5 m apart, with partial visual barriers (bushes) between them. Aviaries contained roosting trees and small boxes for sleeping and resting. Water, sand, and fine gravel (to facilitate digestion) were provided and multivitamin droplets were regularly added to the water. In order to be able to investigate social foraging tactic use in 2005 in Flock 1 and 2, feeding in these flocks took place on a grid (1.2 × 1.2 m) that contained 144 (12 × 12) equidistant wells (diameter 2.5 cm, depth 1.2 cm) for presenting food (Fig III.3; Lendvai et al. 2004).
Figure III.3. Sparrows became easily accustomed to the grid and used it successfully during social foraging (photos by Z. Tóth).
In 2006 in Flock 3, food was presented in three plastic bowls (diameter approximately 20 cm, depth 9 cm), each fixed on the top of a wooden pool (approximately 90 cm high). We provided millet, oat, wheat and sunflower seeds ad libitum during a 4-week acclimatization period and between different observations. The birds apparently became familiar with the aviaries during acclimatization, and they had also learned to use the grid by the time of the observations in Flock 1 and 2. Throughout the study, we did not observe severe aggression resulting in visible injuries, similarly to the previous studies on captive sparrow flocks (e.g.
Lendvai et al. 2004, 2006). Birds did not lose weight in captivity, their weight even increased slightly during the study (weight of all captive birds at the start of captivity [mean ± SE]:
27.85 ± 0.19 g; at the end of captivity: 28.23 ± 0.19 g; paired t test: t60 = −2.93, n = 61, P = 0.005). After the observations, we released all birds at the site of capture. To facilitate their survival after release, we provided bird food on feeders where we observed the released birds several times during winter. Some of them were also recorded as breeding adults in the following spring.
3. Genetic analyses and kinship estimation
DNA extraction from the stored blood samples was performed with standard phenol- chloroform procedure, or with Qiagen DNeasy Tissue Kit (QIAGEN, Hilden, Germany), following the producer’s instructions. Seven highly polymorphic microsatellite loci were used for genotyping (17.4 0.7 alleles per locus; Table III.2). Primers for four dinucleotide loci (Pdo1, Pdo2 [Neumann & Wetton 1996], Pdo5 [Griffith et al. 1999] and Pdo8 [Griffith et al.
2007]), one trinucleotide locus (Pdo9, Griffith et al. 2007) and one tetranucleotide locus (Pdo3, Neumann & Wetton 1996) were developed specifically for house sparrows. Another dinucleotide locus (McyU4) was originally isolated for the superb fairy-wren (Malurus cyaneus; Double et al. 1997), and was used successfully in genetic studies of sparrows (e.g.
Jensen et al. 2003). In each primer pair (Sigma-Aldrich, Budapest, Hungary) forward primers were fluorescently labelled on the 5’-end with HEX, JOE or FAM-6 dyes (Applied Biosystems, Foster City, CA, USA). PCR reactions consisted of approximately 100 ng of template DNA, 0.5 μM of each primer, 0.2 mM dNTPs, 2 mM MgCl2, 1 unit of Taq DNA polymerase (Fermentas AB, Vilnius, Lithuania) and the 10X Taq buffer in a final volume of 25 μl. To resolve alleles, all amplified PCR products were analyzed on an ABI Prism 3100
Genetic Analyzer (Applied Biosystems) at the Biomi (Gödöllő, Hungary) using ROX-labelled ILS-600 internal standard (Promega, Madison, WI, U.S.A.). The data were analyzed with the GeneScan software (Applied Biosystems). Genetic analyses were conducted in the Molecular Biodiversity Group, Institute of Genetics, HAS Biological Research Center, Szeged.
We used the ML-Relate computer software (Kalinowski et al. 2006) to calculate maximum likelihood estimates of relatedness coefficients and relationship categories between individuals from genotypic data. This method accommodates null alleles during the relatedness estimations which had high frequency at two loci (Pdo2: 0.27, Pdo8: 0.26; Table III.2), and is considered to be more accurate than other estimators (Milligan 2003). Allele frequencies, pair-wise genetic relatedness and kinship category estimations were performed by entering all individuals’ genotype in the program as if they were a single population, since no prior reference data was available about the studied sparrow population (for pair-wise relatedness coefficients in our captive flocks see Appendix 1). We used ML-Relate to calculate the likelihood of four common relationships: U – unrelated, HS – half-siblings, FS – full-siblings, PO – parent-offspring (no other relationships are allowed by the software) and to determine the relationship that had the highest likelihood for each pair of individuals (Kalinowski et al. 2006). In the statistical analyses of Studies 1, 2 and 3, birds with which a given individual had HS, FS and PO relationships were pooled and considered as “close kin”.
The “unrelated” flock-mates were those birds with which a given individual was estimated to be unrelated (U) according to ML-Relate (for average genetic relatedness between close kin and unrelated flock-mates in the above-mentioned Studies see Chapter IV/2b, Chapter V/2b and Chapter VI/2b, respectively). This categorization is likely to reflect real relationships with reasonable accuracy, because in our total sample of birds with known pedigree (including free-ranging genotyped sparrows and individuals studied in the captive flocks), 85.4% and 89.9% of the assigned dyads (n = 1225) matched the real (pedigree-based) relationships in the close kin and the unrelated group, respectively. Furthermore, for the same set of dyads with pedigree-based kinship information, pair-wise relatedness coefficients were estimated 0.423 0.032 for full siblings and parent-offspring dyads (n = 48) and 0.042 0.002 for unrelated birds (offspring from different broods of different parents; n = 1177 dyads).
Table III.2. Number of alleles, allele sizes and their estimated frequencies in seven highly polymorphic microsatellite loci (combined results for all the genotyped house sparrows, i.e. both free- ranging and captive birds, n=258).
Pdo1 Pdo2 Pdo3 Pdo5 Pdo8 Pdo9 Mcyu4
Number of alleles 18 18 15 20 18 18 15
Allele sizes (allele frequencies)
154 (0.124)
169 (0.021)
109 (0.003)
203 (0.145)
189 (0.005)
369 (0.003)
177 (0.008) 158
(0.003)
171 (0.015)
113 (0.023)
205 (0.008)
193 (0.008)
372 (0.003)
179 (0.176) 166
(0.005)
173 (0.015)
117 (0.013)
218 (0.003)
195 (0.145)
375 (0.092)
181 (0.008) 168
(0.008)
177 (0.023)
121 (0.056)
224 (0.005)
197 (0.018)
378 (0.239)
183 (0.066) 172
(0.041)
179 (0.003)
125 (0.084)
226 (0.031)
199 (0.233)
381 (0.015)
185 (0.003) 174
(0.305)
181 (0.010)
129 (0.142)
228 (0.008)
201 (0.054)
384 (0.023)
187 (0.059) 178
(0.003)
183 (0.097)
133 (0.157)
230 (0.020)
204 (0.005)
387 (0.074)
189 (0.107) 180
(0.013)
185 (0.045)
137 (0.122)
233 (0.013)
208 (0.026)
390 (0.156)
191 (0.074) 182
(0.013)
187 (0.204)
141 (0.137)
235 (0.013)
210 (0.132)
393 (0.036)
193 (0.084) 184
(0.140)
189 (0.075)
146 (0.074)
237 (0.013)
212 (0.045)
396 (0.036)
195 (0.117) 186
(0.036)
191 (0.118)
150 (0.058)
239 (0.084)
216 (0.003)
402 (0.015)
197 (0.100) 188
(0.091)
193 (0.036)
154 (0.048)
241 (0.003)
218 (0.003)
405 (0.026)
199 (0.153) 190
(0.008)
195 (0.015)
159 (0.043)
243 (0.074)
220 (0.013)
409 (0.239)
201 (0.038) 192
(0.099)
197 (0.003)
163 (0.031)
245 (0.117)
222 (0.008)
414 (0.008)
205 (0.005) 194
(0.051)
199 (0.003)
167 (0.010)
247 (0.240)
224 (0.023)
417 (0.008)
207 (0.003) 196
(0.038)
201 (0.037)
249 (0.107)
226 (0.020)
420 (0.010) 198
(0.020)
203 (0.008)
251 (0.069)
228 (0.003)
423 (0.008) 208
(0.005)
205 (0.005)
253 (0.038)
230 (0.003)
426 (0.010) NULL
(0.269)
255 (0.008)
NULL (0.257) 259
(0.003)
IV. S
TUDY1: G
ENETIC RELATEDNESS IN WINTERING GROUPS OF HOUSE SPARROWS(P
ASSER DOMESTICUS)
Abstract
Social behaviour of group-living animals is often influenced by the relatedness of individuals, thus understanding the genetic structure of groups is important for the interpretation of costs and benefits of social interactions. In this study we investigated genetic relatedness in feeding aggregations of free-living house sparrows (Passer domesticus) during the non-breeding season. This species is a frequent model system for studies of social behaviour (e.g. aggression, social foraging), but we lack adequate information on the kin structure of sparrow flocks. During two winters, we ringed and observed sparrows at feeding stations, and used resightings to identify stable flock-members and to calculate association indices between birds. We genotyped the birds using seven highly polymorphic microsatellite loci, and estimated pair-wise relatedness coefficients and relatedness categories (close kin vs.
unrelated) by maximum likelihood method. We found that most birds were unrelated to each other in the flocks (mean SE relatedness coefficient: 0.06 0.002), although most individuals had at least a few close relatives in their home flock (14.3 0.6 % of flock- mates). Pair-wise association between individuals was not significantly related to their genetic relatedness. Furthermore, there was no difference between within-flock versus between-flock relatedness, and birds had similar proportions of close kin within and outside their home flock. Finally, relatedness among members of different flocks was unrelated to the distance between their flocks. Thus, sparrow flocks were not characterized by association of relatives;
nevertheless, the presence of some close kin may provide opportunity for kin-biased behaviours to evolve.
Liker A., Bókony V., Kulcsár A., Tóth Z., Szabó K., Kaholek B. & Pénzes Zs. 2009. Genetic relatedness in wintering groups of house sparrows (Passer domesticus). Molecular Ecology, 18, 4696–4706.
1. Introduction
In group-living animals the genetic relatedness among group-members often influences the costs and benefits of their social interactions, for example via kin selection (Hamilton 1964, Krause & Ruxton 2002). Benefits gained through increased inclusive fitness of related group-members may explain various kin-biased behaviours such as helping breeding relatives rearing offspring (Russell & Hatchwell 2001, Baglione et al. 2003), association between related males at display sites (Petrie et al. 1999, Shorey et al. 2000, Krakauer 2005), increased anti-predator behaviour in the presence of close kin (Dunford 1977) and differential exploitation of flock-mates during social foraging (Ha et al. 2003).
Thus, information about the fine-scale genetic structure of social groups is important when we are to get a realistic understanding of the pay-offs of social interactions between group-mates.
In birds, relatedness has usually been studied during breeding, typically in relation to helping behaviour, inbreeding and incest avoidance. In these contexts, a relatively high number of studies focused on cooperatively breeding groups (see examples above) and the genetic relationships of breeders within a population (e.g. Foerster et al. 2006, Lebigre et al.
2008, Szulkin & Sheldon 2008). Few studies investigated relatedness in non-breeding social units, although knowledge on relatedness in such groups is as important as in breeding groups. Firstly, group formation is widespread during migration and wintering, and individuals may aggregate during these periods even in solitarily breeding species (e.g.
Ekman 1989). Secondly, social interactions are frequent in non-reproductive groups, including aggression for food and shelter, exploitation of others’ food resources by scrounging, and alarming flock-mates upon predation risk (Elgar 1989, Giraldeau & Caraco 2000, Krause & Ruxton 2002). All of these interactions involve some conflicts between group-mates’ interests, so the presence of related individuals may provide opportunity for the evolution of kin-biased behaviour.
In this study we investigated genetic relatedness in feeding aggregations of non- breeding house sparrows (Passer domesticus). Sparrows may aggregate at the same feeding and roosting sites throughout the whole non-breeding period, or even year after year (e.g. we are aware of traditional daytime gathering and feeding sites that have been used for 5-10 years; A. Liker & V. Bókony, personal observations). Although the stability of flock composition has not been studied, it is likely to change dynamically to some extent because birds may use several feeding sites (Beer 1961). On the other hand, the highly sedentary
nature of the house sparrows (Summers-Smith 1963, Anderson 2006) suggests that a substantial proportion of the birds in feeding aggregations may be resident around the sites and are present regularly at the same feeding sites, yielding some stability in the social structure. Since dispersal is often limited in the species (Summers-Smith 1963, Altwegg et al.
2000, Anderson 2006; but see Fleischer et al. 1984), related individuals may regularly forage and roost as members of the same flock.
We used intense ringing and observation of wild birds, in combination with molecular genetic data on relatedness, to investigate whether feeding aggregations of house sparrows include aggregations of related individuals. Specifically, we tested (1) whether the degree of association among birds at feeding sites reflects their genetic relatedness; (2) whether birds are more closely related to their flock-mates, i.e. to birds with whom they regularly feed together, than to members of other flocks that use different feeding sites; and (3) whether the distance between flocks correlates with genetic relatedness between birds from these different flocks. Since female house sparrows move to farther distances than males during natal dispersal (Altwegg et al. 2000), we also investigated these questions separately for the sexes.
2. Materials and methods
a) Observations of feeding aggregations
We operated feeding stations at two sites (W and C) during the winter of 2004 – 2005, and at all five study sites during the winter of 2005–2006 (for detailed description of the study area and the feeding stations see Chapter III/2b). Seeds (mixture of wheat, sunflower, corn grit and millet) were provided on bird tables (one per site) erected on poles near the roosting bushes. In addition, the same food was provided on a feeding platform placed on the ground at the two sites used in 2004–2005 (Bókony et al. 2008). At site C we also observed birds feeding in a nearby (70 m) outlet of the zoo’s raccoons (Procyon lotor) where the raccoons’
food was provided in three containers erected on separate poles (see Bókony et al. 2008), and this food was regularly consumed by the same groups of sparrows that visited the bird table (so we pooled observations at the bird table and at the raccoon feeders for site C). The provision of seeds on the bird tables started several weeks before the beginning of observations and continued until the end of the study, while the raccoon feeders were operated continuously over the whole year.
We recorded the occurrence of ringed birds at the feeding stations during regular observations that were conducted between 5 January and 11 February in 2005, and between 11 January and 3 February in 2006. The weather during these periods was typically cold (below 0C) with frequent snowfalls, so the sparrows readily used the feeders at all sites. In 2005, 20 and 40 observations were conducted, respectively, at site W and C (20 at the bird table and 20 at the raccoon feeders). In 2006, 13 observations were conducted at site N, 10 at W, 22 at C (12 at the bird table and 10 at the raccoon feeders), 14 at S, and 10 at U.
Observations were conducted from remote hidden locations (ca. 50 m) using 20–60× spotting scopes, and they usually lasted 60 minutes, although some of them had to be interrupted earlier because of zoo maintenance work or bad weather conditions. Most observations of a given site were carried out on different days, and usually all sites were observed on each day of the fieldwork. In some cases two observations per day were conducted at the same site, and in these cases there was at least an hour gap between subsequent observations. During each observation we recorded the identity and sex of each colour-ringed sparrow that fed on the feeders or stayed nearby (e.g. near the platform or below the table on the ground) and was recognizable unambiguously by its colour rings. In 2005 additional recordings were collected from high resolution videotapes at the container feeders of site C (see Bókony et al. 2008), that were added to the records collected by the observers. Note that in some observations we could not identify all colour-ringed birds present at the station, e.g. when a large number of birds used simultaneously the feeder but stayed there only for brief periods. In total we collected 5220 records of resightings (including multiple records of the same individuals during a single observation) for 436 ringed birds during the two years of the study. Two (V.
Bókony and A. Liker) and three (V. Bókony, A. Kulcsár and A. Liker) observers, working simultaneously at different sites, conducted the data collection in 2005 and 2006, respectively.
b) Selection of birds for the analyses
For the subsequent analyses we selected those individuals that had been observed in at least 3 independent observations. Multiple recordings of the same individual within one observation were counted as one, and the above criterion was applied separately for 2005 and 2006. The mean SE number of observations for the selected birds was 11.5 4.4 (n = 79 birds) and 4.9 1.7 (n = 128 birds) in 2005 and 2006, respectively. All of these individuals (n
= 188, because 19 birds were included in both years’ analyses) were genotyped and used in the statistical analyses.
For some analyses, we categorized the selected birds into two groups, according to the number of sites they were observed at. Firstly, birds were assigned to the “core flock” of a specific site if they had been recorded exclusively at that site in all of their observations (n = 66 and 103 individuals in 2005 and 2006, respectively). Secondly, birds observed at more than one site within a year were categorized as “commuters”, i.e. these birds moved between sites at least once between the observations (n = 13 and 25 individuals). This categorization was conducted for the two study years separately, and individuals present in both years were categorized for each year. The size of the core flocks correlated strongly with the estimated median flock sizes of the sites (see above; Pearson correlation: r = 0.94, n = 5 sites, P = 0.0195, data from 2006).
c) Statistical analyses
In the first set of analyses we used the records of all birds from the observations to calculate half-weighted association indices (Cairns & Schwager 1987) between each pair of individuals (i.e. including both core-flock and commuter birds). This calculation was performed using the SOCPROG 2.3 software (Whitehead 2007), with observations as sampling units (n = 60 and 69 units for 2005 and 2006, respectively). The half-weighted association index is a measure of the proportion of observations in which the two birds occurred together, taking the lowest value (0) for dyads that were never recorded in the same observation (e.g. they were observed exclusively at separate sites, or at the same sites but in different observations), whereas the highest value (1) is assigned to dyads that are exclusively recorded together (i.e. always occurred at the same site and in the same observations). This index was calculated separately for the two years. To test whether association in feeding groups was related to kinship between birds (as estimated from the genotypic data, see Chapter III/3), we calculated correlations between relatedness coefficients and association indices. Correlation of pair-wise matrices was tested by the Mantel randomization tests with 10000 permutations as implemented in SOCPROG 2.3. We analysed the two years separately, and ran separate analyses for the pooled dataset of all dyads and for sex-specific (male-male, female-female, and male-female) dyads. In a subset of analyses, we also investigated the core flock of each site separately (i.e. commuters were excluded).
In the second set of analyses we focused on core-flock individuals, and tested whether relatedness of birds within a core flock (i.e. among individuals using exclusively the same feeding site) differed from the relatedness of these individuals to birds assigned to other core flocks (pooling individuals from all other sites). To do this, first we compared relatedness coefficients between the two dyad types (within versus between core flocks) by randomization test (Mann-Whitney test with 10000 permutations). After that we calculated the proportion of close kin (rML [mean ± SE] = 0.22 ± 0.004; for detailed description about kinship categories see Chapter III/3) present within and outside the core flock for each individual (number of close kin in the flock per number of flock-mates; number of close kin outside the flock per number of non-flock-mates). Then we tested for differences between the two proportions by randomization test (paired Wilcoxon test with 10000 permutations), using individuals as data points. We analysed the two years separately, and within each year a separate analysis was run for each core flock. These analyses were performed in the R statistical computing environment (R Development Core Team 2008).
Finally, we tested whether the spatial distances between core-flock individuals were correlated with their genetic relatedness. We assigned all individuals of a core flock to a single spatial location (feeder site). Within-flock distance was set to 0 m, whereas between- flock distances were calculated from the geographic coordinates of the sites. In this analysis we only included data from 2006, yielding 10 pair-wise distances for the 5 sites. The same distance was assigned to all dyads of each pair of core flocks. We analysed the relationship between pair-wise spatial distances and relatedness coefficients by matrix correlations and Mantel randomization tests with 10000 permutations using SOCPROG 2.3. Separate analyses were run for the pooled dataset of all dyads and for sex-specific dyads. To augment these spatial analyses, we also tested for sex-biased dispersal based on the mean corrected assignment index (mAIc) described by Goudet et al. (2002), using Fstat 2.9.3.2 software with 10000 randomizations (Goudet 1995). A significantly lower mAIc value for one sex indicates that dispersal is biased towards that sex. This method was chosen because it is accepted as the most powerful tool for detecting sex-biased dispersal from genotypic data within a single population (Goudet et al. 2002).
3. Results
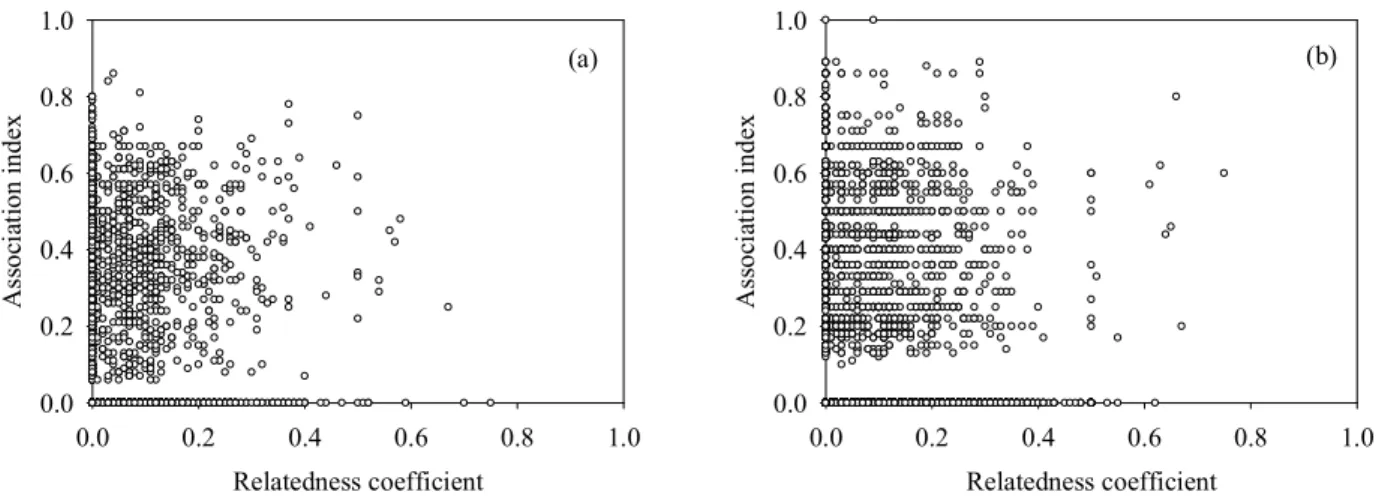
a) Association and relatedness
In 2005, there was no significant correlation between the relatedness of individuals and their association in the observations (Table IV.1, Fig. IV.1a). In 2006, relatedness coefficients and association indices correlated positively for the pooled (not sex-specific) dataset (Table IV.1). However, this correlation was weak (r = 0.02; Fig. IV.1b) and became non-significant when significance levels were adjusted for multiple testing. We did not find any significant correlation between relatedness and association indices when either sex- specific dyad types or core flocks were analysed separately (Table IV.1).
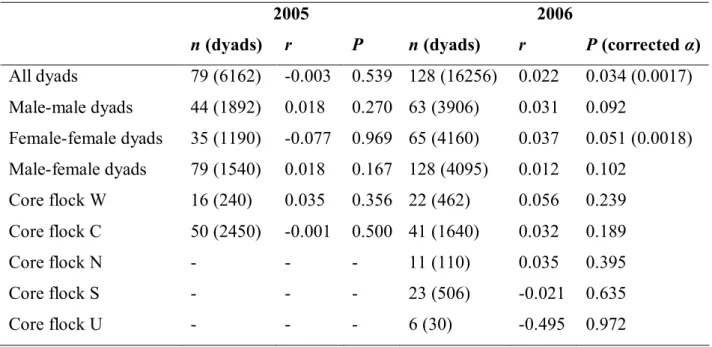
Table IV.1. Correlation between pair-wise genetic relatedness and association at feeding sites among house sparrows. For the analyses of all, male-male, female-female and male-female dyads, we used all available individuals (core flocks plus commuters) from the studied year. In the separate analyses of core flocks, only within-flock dyads were included (core flocks N, S and U were studied only in 2006). Correlation coefficients (r) were calculated by matrix correlation, and the associated P-values were calculated by Mantel randomization tests with 10000 permutations; n = number of individuals and the number of dyads used in the tests are given in parentheses. Note that P values are one-tailed, as provided by the software (SOCPROG 2.3). Corrected significance levels (α) are given for P < 0.05 results (sequential Bonferroni correction for 15 comparisons, one-tailed).
2005 2006
n (dyads) r P n (dyads) r P (corrected α) All dyads 79 (6162) -0.003 0.539 128 (16256) 0.022 0.034 (0.0017) Male-male dyads 44 (1892) 0.018 0.270 63 (3906) 0.031 0.092
Female-female dyads 35 (1190) -0.077 0.969 65 (4160) 0.037 0.051 (0.0018) Male-female dyads 79 (1540) 0.018 0.167 128 (4095) 0.012 0.102
Core flock W 16 (240) 0.035 0.356 22 (462) 0.056 0.239 Core flock C 50 (2450) -0.001 0.500 41 (1640) 0.032 0.189
Core flock N - - - 11 (110) 0.035 0.395
Core flock S - - - 23 (506) -0.021 0.635
Core flock U - - - 6 (30) -0.495 0.972
Relatedness coefficient
0.0 0.2 0.4 0.6 0.8 1.0
Association index
0.0 0.2 0.4 0.6 0.8 1.0
(a)
Relatedness coefficient
0.0 0.2 0.4 0.6 0.8 1.0
Association index
0.0 0.2 0.4 0.6 0.8 1.0
(b)
Figure IV.1. Genetic relatedness versus association at feeding sites between house sparrows in 2005 (a) and 2006 (b). Data points represent dyads. See Table IV.1 for statistical results.
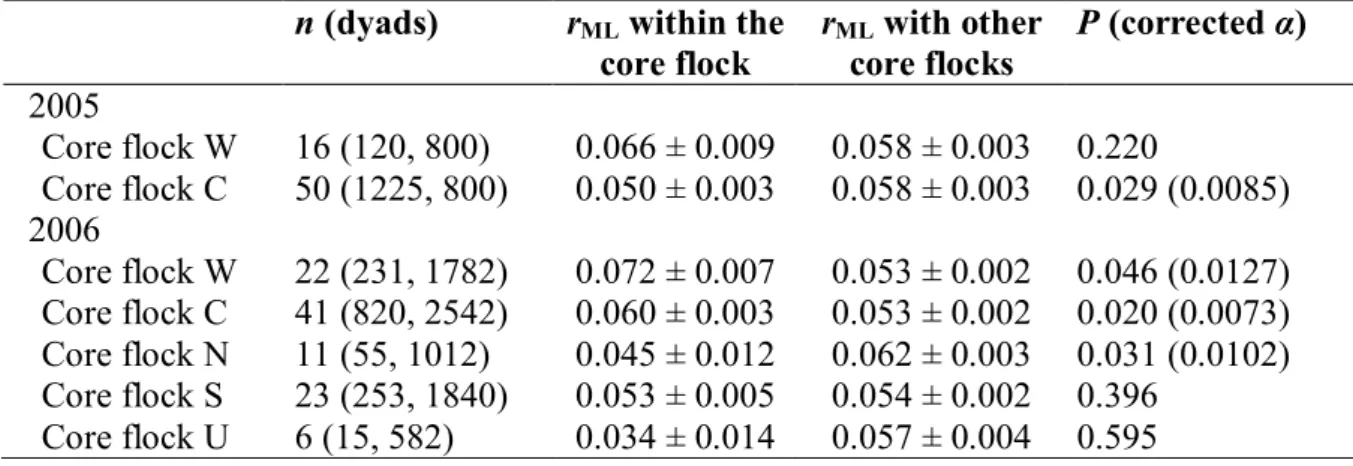
b) Relatedness within and between core flocks
The average relatedness within core flocks was low (Table IV.2), and in most cases it was not different from relatedness between birds belonging to different core flocks. Although we detected significant differences in some comparisons, these effects were small and inconsistent in both cases; among core flocks (e.g. N versus C in 2005) and between the two years (core flock C), and did not remain statistically significant after sequential Bonferroni correction (Table IV.2). We obtained qualitatively identical results when sex-specific dyads were analysed separately for each core flock (Appendix 2). The variance in relatedness across dyads was also small, both in the pooled sample of all dyads in each year (0.008) and within each core-flock (range: 0.003–0.013).
Almost all sparrows had at least one close kin (FS, HS or PO) both within (96% of birds) and outside their flocks (97%). On average, the proportion of close kin was 0.143 0.006 (range: 0–0.439) within core flocks and 0.149 0.006 (range: 0–0.500) between different flocks (Fig. IV.3). Individuals did not differ in the proportion of their close kin within and outside their core flocks in any but one core flock, and this latter difference became non-significant after Bonferroni correction (Table IV.2). We obtained qualitatively the same results when the sexes were analysed separately for each core flock (Appendix 3).