DOKTORI (PhD) ÉRTEKEZÉS
LEPOSSA ANITA
KESZTHELY
VESZPRÉMI EGYETEM
*(25*,.210(= *$='$6È*78'20È1<,.$5
NÖVÉNYTERMESZTÉSI ÉS KERTÉSZETI TUDOMÁNYOK DOKTORI ISKOLA
3URJUDPYH]HW
DR. DEBRECZENI BÉLÁNÉ MTA doktora
7pPDYH]HW N
DR. MÁTÉ FERENC MTA doktora
DR. ÖRDÖG VINCE a biológiai tudományok kandidátusa
TALAJALGÁK MENNYISÉGI VIZSGÁLATA A BALATON-FELVIDÉKI NEMZETI PARKBAN,
VALAMINT TALAJBÓL IZOLÁLT
ALGATENYÉSZETEK NÖVÉNYI NÖVEKEDÉST BEFOLYÁSOLÓ HATÁSAINAK KIMUTATÁSA
Készítette:
LEPOSSA ANITA
KESZTHELY 2003
TALAJALGÁK MENNYISÉGI VIZSGÁLATA A BALATON-FELVIDÉKI NEMZETI PARKBAN,
VALAMINT TALAJBÓL IZOLÁLT ALGATENYÉSZETEK NÖVÉNYI NÖVEKEDÉST BEFOLYÁSOLÓ HATÁSAINAK KIMUTATÁSA
Értekezés doktori (PhD) fokozat elnyerése érdekében Írta:
Lepossa Anita
Készült a Veszprémi Egyetem „Növénytermesztési és Kertészeti Tudományok” Doktori Iskolája keretében
7pPDYH]HW N Dr. Máté Ferenc Dr. Ördög Vince Elfogadásra javaslom (igen / nem) (igen / nem)
………. ………
aláírás aláírás
A jelölt a doktori szigorlaton …... % -ot ért el,
Veszprém, Keszthely …...………
a Szigorlati Bizottság elnöke
Az értekezést bírálóként elfogadásra javaslom:
Bíráló neve: …... …... igen /nem
………
(aláírás) Bíráló neve: …... …... igen /nem
………
(aláírás) A jelölt az értekezés nyilvános vitáján…...% - ot ért el
Veszprém/Keszthely, ………
a Bíráló Bizottság elnöke
$GRNWRUL3K'RNOHYpOPLQ VtWpVH«
………
az EDT elnöke
Tartalomjegyzék
Oldal
Kivonatok 7
1. Szakirodalmi összefoglaló 9
1.1. Mit nevezünk talajalgának? 9
1.2. A talajalgológia, és helyzete Magyarországon 10
1.3. $WDODMDOJiNUDKDWy|NROyJLDLWpQ\H] N 12
1.3.1. Életmód 12
1.3.2. Fény 13
1.3.3. Víz 16
1.3.4. + PpUVpNOHW 19
1.3.5. pH 20
1.3.6. Tápanyagok 22
1.4. AWDODMDOJiNPHQQ\LVpJLHO IRUGXOiVD 27
1.4.1. Sejtszámlálási módszerek 27
1.4.2. A talajalgák kvantitatív változásai 36
1.5. $ILWRHGDIRQV]HUHSHpVMHOHQW VpJH 40
1.5.1. 7DODMpOHWEHLQGtWiVDWDODMYpG pVV]HUNH]HWMDYtWyKDWiV 41
1.5.2. N2-fixáció 43
1.5.3. Interspecifikus kapcsolatok 45
1.6. $GROJR]DWFpONLW ]pVHL 48
2. Kísérleti rész 49
2.1. Anyag és módszer 49
2.1.1. Mintavételi területek jellemzése 49
2.1.2. Talajminta-J\ MWpVDOJDPHQQ\LVpJLYL]VJiODWRNKR] 51
2.1.3. Sejtszámlálás, biomassza-számítás 51
2.1.4. Környezeti paraméterek mérése 53 2.1.5. 7DODMDOJiNUDKDWyWpQ\H] NVWDWLV]WLNDLYL]VJiODWD 54 2.1.6. Talajalgák izolálása, egyfaj-WHQ\pV]HWHN HO iOOtWiVD
faji határozás
55
2.1.7. Laboratóriumi algatenyésztés 55
2.1.8. Biotesztek algakivonatok auxin- és citokinin-V]HU
hatásának vizsgálatára
56 2.1.9. Csíranövény-WHV]WpO DOJDWHQ\pV]HWKDWiViQDNYL]sgá-
latára
57
2.2. Kísérleti eredmények 58
2.2.1. Mintavételi területeken tett megfigyelések 58
2.2.2. A sejtszámlálás eredményei 59
2.2.3. Vizsgált környezeti paraméterek 60
2.2.4. Algatársulások jellemzése, számított biomassza 62 2.2.5. Környezeti hatások vizsgálata variancia-analízissel 65 2.2.6. Környezeti hatások vizsgálata regresszió-analízissel 74 2.2.7. Izolált és laboratóriumban tenyésztett algatörzsek
jellemzése
83 2.2.8. Auxin- és citokinin-V]HU KDWiVWPXWDWyDOJDNLYRQDWRN 87 2.2.9. eO DOJDWHQ\pV]HWKDWiVDPXVWiUPDJRNFVtUi]iViUD 91
2.3. Eredmények értékelése, következtetések 93
Összefoglalás 106
Új tudományos eredmények 109
Köszönetnyilvánítás 112
Felhasznált irodalom 113
Mellékletek
KIVONAT
Talajalgák mennyiségi vizsgálata a Balaton-felvidéki Nemzeti Parkban, valamint talajból izolált algatenyészetek növényi növekedést befolyásoló
hatásainak kimutatása
Egyéves vizsgálatsorozatban a Balaton-felvidéki Nemzeti Park tala-
MDLEDQ pO DOJiN DEXQGDQFLiMiQDN KDYRQNpQWL YiOWR]iViW PpUWHP N|]YHt-
OHQVHMWV]iPROiVLPyGV]HUUHO$YL]VJiOWN|UQ\H]HWLWpQ\H] NQHND]DOJiN
mennyiségi változásaiban megmutatkozó hatását variancia- és regresszió- analízissel értékeltem. Az eredmények azt mutatták, hogy a talajalgák
PHQQ\LVpJpWDODSYHW HQEHIRO\iVROyWpQ\H] ND WDODMPpO\VpJpVDP Ye- lés, míg a cianobaktériumok és kovaalgák számára a talajtípus is megha- tározó. Évszak-hatás az eukarióta algák esetében mutatkozott nyári maximummal. Biotesztekkel bizonyítható volt a talajokból izolált egyes cianobaktérium-pV]|OGDOJDWHQ\pV]HWHNpO EHYRQDWiQDNpVYL]HVVHMWNi- vonatának növényi növekedést szabályozó hatása.
ABSTRACT
Quantitative analysis of soil algae in the Balaton Upland National Park, and demonstration of plant growth regulating effects of algal cultures
isolated from soil
Monthly variations of the algal abundance in the soils of the Balaton National Park were measured by a direct cell counting method in a one-year- long investigation. Variance- and regression analyses were used to evaluate the effect of the environmental factors on the quantitative changes of algae. Results showed basic role of soil depth and cultivation, while significant effect of soil
type could be determined on cyanobacteria and diatoms. Seasonal effect developed in the abundance of eukaryotic algae by maximum values during summer. Plant growth regulating effect of both living cells and water extract of some cyanobacterial and green algal strains isolated from soil was proved by biotests.
AUSZUG
Quantitative Untersuchung von Bodenalgen auf dem Gebiet vom Nationalpark Balaton-Oberland, und Nachweis beeinflussender Auswirkungen von aus dem Boden isolierten Algenkulturen auf das
Wachstum verschiedener Pflanzenarten
Im Rahmen einer einjährigen Untersuchungsreihe habe ich die monatlichen Veränderungen der Abundanz der in den Böden vom Nationalpark Balaton- Oberland lebenden Algen mit einer direkten Zellenzählungsmethode gemessen.
Die Auswirkungen der untersuchten Umweltfaktoren auf die quantitativen Veränderungen der Algen habe ich mit Varianz- und Regressionsanalyse bewertet. Die Ergebnisse haben gezeigt, dass die Bodentiefe und die Bewirtschaftung die Häufigkeit der Bodenalgen grundsätzlich beeinflussen, während für die Zyanobakterien und für die Kieselalgen auch der Bodentyp bestimmend ist. Die Wirkung der Jahreszeiten hatte sich bei den Eukariotenalgen mit einem Maximum im Sommer gezeigt. Mit Hilfe von Biotests konnte die Wachstum regulierende Wirkung des lebendigen Belages einzelner, aus dem Boden isolierten Zyanobakterien- und Grünalgenkulturen bewiesen werden.
1. SZAKIRODALMI ÖSSZEFOGLALÓ 1.1. Mit nevezünk talajalgának?
$] HOQHYH]pV YROWDNpSSHQ J\ MW IRJDORP PHO\QHN V] NHEE YDJ\ WiJDEE pUWHOPH]pVpWWHNLQWYHUpV]LQWFVDNREOLJiWWDODMEDQ pO DOJiNWHNLQWKHW NYDOyMá-
EDQWDODMDOJiNQDNPiVUpV]U OYLV]RQWDIDNXOWDWtYWDODMODNyOHYHJ -, kéreglakó, fal- és sziklabevonatot aONRWy ]X]PyDONRWy pV QpKD PpJ D KDYRQ pV MpJHQ pO NULRILWDDOJiNDWLVHEEHDFVRSRUWEDVRUROMiN(WWOpV*lUWQHU(EE ODIHl-
IRJiVEHOL NO|QEVpJE O LV DGyGKDW D NDWHJyULD IDMVSHNWUXPiUD YRQDWNR]y QDJ\
eltérés: míg Round (1981 cit. Ettl és Gärtner 1995) csupán 100 közönséges ta- lajlakó fajról írt, addig orosz talajokból 1500 fajt jegyeztek le (Sirenko 1999).
A talajalgákat - HOV VRUEDQWHUP KHO\-igényük alapján – az irodalom alap-
YHW HQNpWIpOHEHVRUROiVEDQLVPHUWHWL-1.2. táblázat).
1.1.táblázat: 6]iUD]I|OGLpO KHO\HNDOJiL+RIIPDQQV]HULQW
-WDODMDOJDWDODMRQLOOWDODMEDQpO
- epilita (sziklafalak bevonatát alkotó)
-KLSROLWDN|YHNNDYLFVRNDODWWDWDODMRNPpO\HEEUpWHJpEHQpO
- kazmoendolita (sziklaUHSHGpVHNEHQpO
-NULSWRHQGROLWDSRUy]XVN ]HWHNUHJHLEHQpO
- litofita alga
- endolita
-HXHQGROLWDN ]HWHNEHMiUDWRWYiMy
-EDUODQJLDOJDEDUODQJRNIDOiQEHYRQDWRWNpSH]
- hó-pVMpJDOJDKyERUtWRWWDWHUOHWHNHQIHOV]tQLMpJNULVWiO\RNEDQpO
- epifita alga (s]HUYH]HWWHEEQ|YpQ\HNHQSOIiNNpUJpQYDJ\OHYHOHLQpO
-HSL]RRQDOJDiOODWRNRQSOODMKiURNRQFVLJiNRQt]HOWOiE~DNRQpO
1.2.táblázat: Aero-terresztris algák csoportosítása Ettl és Gärtner (1995) szerint -HXWHUUHV]WULVDWDODMIHOV]tQLpVPpO\HEEUpWHJHLEHQpO
-KLGURWHUUHV]WULVQHGYHVDOM]DWRQpO
-DHURWHUUHV]WULVWDODMIHOV]tQHQpVOHYHJ EHQpO
terresztris (fakultatív aerofita)
- hipolita (kvarc-pVPpV]N|YHNDODWWpO
-HSLILWDNpUJHQOHYpOHQPRKiQVWEpO
- xilofita (fán lakó)
- litofita (sziklabevonatot alkotó)
aero-terresztris algák
aerofita (fakultatív terresztris)
- fikobionta (zuzmóalkotó)
$IHQWLHNE OLVNLW QLNKRJ\DWiUJ\DOWDOJDFVRSRUWUDQHPOHKHWHJ\pUWHOP
definíciót adni. Míg Hoffmann csoportosításában külön kategóriát képviselnek, Ettl és Gärtner felosztása jóval rugalmasabb, a kategóriák között átfedést enged.
Közismerten vannak algafajok, melyek az evolúció során sikeresebben adaptá-
OyGWDNYDODPHO\V]iUD]I|OGLpO KHO\KH]YDJ\N|UQ\H]HWKH] Többségük azonban az igényelt ökológiai feltételek biztosításával vizes közegben is életképes, és jól szaporítható. Ezért úgy gondolom, hogy nem szabad éles határvonalat húzni a talajalga-NDWHJyULD N|Up pV D] iOWDOiQRV IHOIRJiVQDN PHJIHOHO HQ GROJR]DWRm- bDQWDODMDOJiQDNWHNLQWHNPLQGHQWDODMRQpVWDODMEDQHO IRUGXOyDOJafajt.
1.2. A talajalgológia, és helyzete Magyarországon
A talajalgák biológiájának, ökológiájának és rendszertanának gazdag nem- zetközi irodalma van az esettanulmányoktól (Fehér 1936, 1948; Fehér és Frank 1936, Lund 1947, Parker 1961, Parker és Bold 1961, Kiss 1968, 1987-1988;
Shtina és Nekrasova 1971, Komáromy 1975, 1976, 1983, 1985; Davey és
5RWKHU\ D QDJ\REE OpOHJ]HWYpWHO P YHNLJ )HKpU 6KLHOGV pV
Durrell 1964, Lund 1967, Starks és mtsai 1981, Metting 1981, 1992, 1994;
Shubert és Starks 1985, Wild 1988, Archibald 1990, Ettl és Gärtner 1995). A környezethatás-vizsgálatok eredményei alapján általánosságban megállapítható, hogy a talajok algaközösségét befolyásoló legfontosabb WpQ\H] N D] pOHWPyG IpQ\ QHGYHVVpJ K PpUVpNOHW S+ pV D IHOYHKHW WiSDQ\DJWDUWDORP $] |VV]e-
J\ OWLVPHUHWHNHOOHQpUHVHPDGKDWyiWIRJyNpSHERQ\ROXOW|NRV]LV]WpPD|VV]e-
WpWHOpUH pV P N|GpVpUH YRQDWNR]yDQ H]pUW D] VHP MyVROKDWy SRQWRVDQ KRJ\ D
talajok DOJDN|]|VVpJH PLNpQW UHDJiO D N|UQ\H]HWL WpQ\H] N YiOWR]iVDLUD 2NDLW D]DOiEEIHOVRUROWWpQ\H] NEHQOiWRP
1) az algákhoz - mint mesterséges rendszertani kategóriába - sorolt szervezetek óriási diverzitása;
2) korábbi vizsgálatok mérési nehézségei, az algák mennyiségi meghatározá-
ViQiO DONDOPD]RWW PyGV]HUHN IRO\DPDWRV IHMO GpVH UpYpQ D] HUHGPpQ\HN
összehasonlítása nehézségekbe ütközik;
3) a faji határozás nehézségei, a taxonómia állandó változása;
4) többségében kis mintaszámú vizsgálatok alapján vontak le általános érvé-
Q\ N|YHWNH]WHWpVHNHW
5) DPpUpVHNQDJ\UpV]HNHYpVN|UQ\H]HWLWpQ\H] HJ\LGHM YL]VJiODWiUDWHUMHGW
ki;
6) DWDODMDOJiN|NROyJLiMiUyO|VV]HJ\ OWLVPHUHWHNW~OQ\RPyUpV]WODERUDWyULXPL NtVpUOHWHNE OpVPHJILJ\HOpVHNE OV]iUPD]QDN
7) valamint a korábban már említett, csoportosítás körüli viták.
Hazánkban a talajalgákkal foglalkozó kutatások a múlt században indultak
PHJ $ KDUPLQFDV pYHNEHQ tUyGRWW D] HOV pV PLQGHGGLJ HJ\HWOHQ EHV]iPROy PDJ\DURUV]iJL WDODMRNEDQ D] DOJiN PHQQ\LVpJpU O DPLNRU D 0DJ\DU .LUiO\L
BányDPpUQ|NL pV (UG PpUQ|NL ) LVNROD 1|YpQ\WDQL ,QWp]HWpQHN SURIHVV]RUD
Fehér Dániel (1936) soproni homokos vályogtalajra települt lucfenyves, vala-
PLQW0LVNROFRQHJ\ PHV]HV YiO\RJRQWHQ\pV] ENNHUG WDODMiQDNNYDQWLWDWtY
algavizsgálatait végezte el. Megfigyelései szerint a leggazdagabb algaflóra az
V]LFVDSDGpNG~VKyQDSRNEDQILJ\HOKHW PHJ
$]LG MiUiVLWpQ\H] NQHNDWDODMDOJiNpOHWpUHJ\DNRUROWKDWiViUDKtYWDIHOD
figyelmet Kiss István (1955, 1959, 1968, 1987-88). Vizsgálataihoz abból a régi megfig\HOpVE OLQGXOWNLKRJ\DWHUP WDODMYDJ\DSDUODJIHOOHWHLG QNpQWKLr-
WHOHQPHJV]tQHVHGLNV~J\YpOWHKRJ\D]H]WRNR]yHJ\VHMW Q|YpQ\HNW|PHg-
SURGXNFLyMD D] ~Q WDODMYLUiJ]iV pV D VHNpO\ YL]HNHW KDVRQOy PyGRQ V]tQH]
"vízvirágzás" biológiai és meteorológiai szempontból rokon jelenségek. Igyeke-
]HWW UiPXWDWQL DUUD D SiUKX]DPUD DPHO\ D WDODMEDQ pO V]tQHV Q|YpQ\L PLNUo-
V]HUYH]HWHN KLUWHOHQ IHOV]DSRURGiVD pV D] LG MiUiV EL]RQ\RV KHO\]HWHL N|]|WW LVPHUKHW IHO 0HJILJ\HOpVHL V]HULQW D WDODMEDQ pV D WDODj felületén a fényviszo-
Q\RNWyOIJJ HQPiV-PiVDOJiNV]DSRURGKDWQDNIHOVHQQHNPHJIHOHO HQHOWpU V]tQH] GpVHNM|KHWQHNOpWUH
A magyarországi talajok algaflórájáról rendelkezésünkre álló gazdag iroda- lom nemzetközileg elismert talajalgológus kutatónk, P. Komáromy Zsuzsanna
QHYpKH] I ] GLN .RPiURP\ -77, 1979, 1982, 1983, 1984,
$]RUV]iJV]iPRVWHUOHWpWpULQW %XGDLKJ0iWUDKJ%NNKJ+Rr- tobágyi NP, Kiskunsági NP) florisztikai kutatásaiban erdei talajok, majd ho- moktalajok összehasonlító algológiai vizsgálatát végezte el. Ennek
HUHGPpQ\HNpSSHQ D] HUG WiUVXOiVRN WDODMDLEDQ DChlorophyta és Xanthophyta divíziók dominanciáját írta le, míg az alföldi meszes és sós talajokat a többség-
EHQOpY FLDQREDNWpULXPRNNDOMHOOHPH]WH$WDODMalgák dominanciájának, gyako- riságának és diverzitásának a legújabb módszertani vizsgálatán túl arra a
NpUGpVUH NHUHVWH D YiODV]W KRJ\ D NO|QE|] WDODMWtSXVRNQDN PLpUW YDQ HOWpU
algaflórája, és ezt fizikai, kémiai és biológiai okokra vezette vissza (Buczkó 1986). 1985-ben, fiatalon bekövetkezett halála után Magyarországon a talajal-
JiNNDOIRJODONR]yNXWDWiVRNVDMQiODWRVPyGRQPHJV] QWHN
7DODMDOJiNUDKDWy|NROyJLDLWpQ\H] N
1.3.1. Életmód
A talajalgák – a szervezettebb növényekhez hasonlóan – alapvet HQ IRWRDXWRWUyI pO OpQ\HN VRN IDMQiO PHJILJ\HOKHW D]RQEDQ D IRWRDX[RWUyI pOHt- mód is (vagyis energiaszükségletük fedezésére a napfényt használják, bizonyos
QpONO|]KHWHWOHQ V]HUYHV DQ\DJRN SUHNXU]RURNEyO W|UWpQ V]LQWHWL]iOiViUD Yi- szont nem képesek). Els NpQW 7UHERX[ FLW 0HWWLQJ tUWD OH WDODMDl- gáknál a fakultatív fotoheterotróf, illetve kemoheterotróf életmódra való áttérés
NpSHVVpJpW H]W N|YHW HQ SHGLJ W|EE NtVpUOHW LV LJD]ROWD PLQG HXNDULyWD DOJiN
mind cianobaktériumok esetében (Parker 1961, Sarma és Khattar 1993). Jelen- legi ismereteink szerint a zöld- és sárgászöld algáknak mintegy 40-50%-a fakul- tatív kemoheterotróf, a cianobaktériumok pedig többségükben fakultatív
fotoheterotrófok (Metting 1981), néhány törzsük esetében kimutatták sötétben és szerves anyag jelenlétében a kemoheterotróf szaporodást is, noha ilyen kö-
UOPpQ\HNN|]|WWD IHMO GpVMyYDO ODVVDEEPLQWIpQ\HQ 7DQGHDXGH 0DUVDFpV
Houmard 1993 cit. Adams 2000). Heterotróf szaporodás esetén a szerves szén-
IRUUiVW HJ\V]HU EE FXNURN V]HUYHV VDYDN pV DPLQRVDYDN DGMiN DPHO\HN IHOYe-
KHW VpJH HJ\EHQ IDML KDWiUR]yEélyeg is. Willson és Forest (1957) talajalgák
L]ROiOiVD VRUiQPHJILJ\HOWpNKRJ\SHSWRQRVpOHV]W NLYRQDWRWWDUWDOPD]yWiSRl-
GDWEDQD]DOJiNJ\RUVDEEDQIHMO GWHNPLQWVWHULOYt]EHQW|UWpQ NLWHQ\pV]WpVNRU
hosszabb inkubációnál azonban az algaflóra elszegényedése mutatkozott.
9DOyV]tQ KRJ\D]DOJiNHVHWpEHQDKHWHURWUyIpOHWPyGOHKHW VpJHHJ\IDMWD W~OpOpVLVWUDWpJLDPHUWQHKH]HQHONpS]HOKHW KRJ\NHGYH] WOHQIpQ\YLV]RQ\Rk- nál és korlátozott szervesanyag-tartalom mellett a talajalgák sikerrel verseng- hetnek obligát heterotróf szervezetekkel.
1.3.2. Fény
Ismert, hogy a talajalgák szaporodására és a sejtek differenciálódási folya-
PDWDLUDKDWiVVDO YDQ D PHJYLOiJtWiVLG WDUWDPD pV DIpQ\ VSHNWUiOLV|VV]HWpWHOH
Cianobaktériumoknál a heterociszta képzése, a nitrogenáz aktivitása és az
DNLQpWD IHMO GpVH LV IpQ\W O IJJ IRO\DPDW +XEHU 3URVSHUL pV PWVDL
1992, Adams és Duggan 1999). Anabaena cylindrica fonalakban a nagy fényin-
WHQ]LWiVQ|YHOWHDNHWW VKHWHURFLV]WiNHO IRUGXOiViQDNJ\DNRULViJiWDNLQpWiLQDN
csírázásához pedig nélkülözhetetlen a második fotorendszer (PS-,, P N|GpVH
(Adams és Carr 1981a,b; Chauvat és mtsai 1982). Fruktózzal kiegészített tápol- datban azonban az akinéták sötétben is kicsíráztak (Neely-Fisher és mtsai 1989). Lazaroff (1966 cit. Tandeau de Marsac 1994) sötétben tartott Nostoc
IDMRNQiOILJ\HOWHPHJKRJ\DY|U|VIpQ\LQGXNiOWDDKRUPRJyQLXPRNNpS] Gé- sét. Eukarióta talajalgák esetében a zoosporogenezis és az ivaros szaporodás folyamatában nélkülözhetetlen a fény.
Feltételezések szerint egyes talajalgák földrajzi elterjedését korlátozhatják a
IDMRNLJpQ\HLQHNQHPPHJIHOHO VXJiU]iVYLV]RQ\RN$]~QNURPDWLNXVDGDSWá- cióra képes cianobaktériumok sejtjeik színanyag-tartalmának megváltozásával alkalmazkodnak a környezet fényviszonyaihoz, így vörös fényben kékes-zöld
V]tQ HN ]|OG IpQ\EHQ SHGLJ Y|U|VHV iUQ\DODW~DN $ WDODMDOJiN D W|EEL PLNUo- szervezethez képest jól tolerálják az UV-sugárzást, ami felhasználható az alga- tenyészetek baktérium-PHQWHVtWpVpUH.DONypV)HOI|OG\+RVV]~LGHM - sugárzásnak kitett talajokban a cianobaktériumok jobban elviselték a rövid hul- lámhosszú fényt, mint az eukarióta algák (Franz és Woodwell 1973 cit. Metting 1981).
$IpQ\V]HUHSHDODSYHW D]DOJiNWDODMRNEDQPHJILJ\HOKHW YHUWLNiOLVHOWHr- jedésében is (Kiss 195/HJQDJ\REEV U VpJEHQ D IHOV]tQLQpKiQ\ FHQWLPpWe-
UHV WDODMUpWHJEHQ IRUGXOQDN HO PpO\HEEUH KDODGYD D] DOJD VHMWV]iP XJUiVV]HU HQ PtJ D IDMV]iP ODVVDEE WHPEHQ FV|NNHQ $OH[DNKLQD
Lukešová 1993). Többen gondolják azt, hogy a talajalgák fototróf CO2-
DVV]LPLOiFLyMD D WDODMV]HOYpQ\ IHOV QpKiQ\ PLOOLPpWHUpUH YDJ\ FHQWLPpWHUpUH
korlátozódik, és a mélyebb rétegek algaflórája csupán inaktív, kitartó alakokból
iOO PHO\HN D FVDSDGpNNDO OHPRVyGWDN YDJ\ DPHO\HNHW D PH] JD]GDViJL WHYé- kenység, a gyökérfejl GpVLOOHWYHWDODMIDXQDMXWWDWRWWRGD/XQG6KLHOGVpV
Durrell 1964, King és Ward 1977). Megfigyelések szerint azonban a maximális
IpQ\EHKDWROiVL]yQDDODWWLWDODMUpWHJHNE OL]ROiOWDOJDIDMRNQDJ\UpV]HQHPNpSH]
akinétát, hipnospórát, vagy egyéb kitartó alakot, ami a heterotróf életmód lehe-
W VpJpW IHOWpWHOH]L )HKpU pV )UDQN NtVpUOHWHLNEHQ KRVV]DEE LGHLJ -2 méter mély talajszintben tartott algatenyészeteket vizsgálva a cianobaktériumok sötéthez való nagyobb alkalmazkodóképességét állapították meg. Szerves anya- got nem tartalmazó, kiizzított talajban 6-12 hónap elteltével is klorofilljukat megtartó, életképes algasejteket találtak. Bristol-Roach (1927) egy méternél is
PpO\HEE WDODMUpWHJE O L]ROiOW DNWtY ]|OGDOJiNDW $Q\DJFVHUpW IRO\WDWy
endedafikus alga-együttes létezésére Tchan és Whitehouse (1953) adtak bizo-
nyítékot. Klorofill-fluoreszcencia méréssel homoktalajokban 4 mm-es mélység-
W ONH]G G HQD] DOJD VHMWV]iP HU WHOMHV FV|NNHQpVpW ILJ\HOWpNPHJ$IpQybe- hatolás mértéke a talajok nedvességtaUWDOPiWyO IJJ HQ FP-es mélységig volt kimutatható, és ez szoros korrelációt mutatott az alga sejtszámlálási ered- ményekkel.
$PHJYLOiJtWRWW]yQiEyODWDODMPpO\HEEUpWHJHLEHNHUO DOJiNEL]RQ\tWRt- tan folytatnak anyagcserét egy ideig, és az is valószín KRJ\HJ\HVDNWtYPRz- gásra képes cianobaktériumok vagy hormogóniumaik, a kovamoszatok és az eukarióta algák zoospórái képesek oda visszavándorolni (Burges és Raw 1967 cit. Wild 1988). A Chlamydomonas zöldalga például ostorai segítségével vizes közegben úszó, a talajban pedig kúszó mozgásra képes. Amikor ostorai szilárd
N|]HJJHO pULQWNH]QHN SpOGiXO WDODMV]HPFVpYHO WiUJ\OHPH]]HO YDJ\ IHG OHPHz-
]HOD]RVWRUYpJHNNHORGDWDSDGpVH]]HODYpOHWOHQV]HU PR]JiVVDODVHMWDV]i- lárd közeg mentén 1,5- PV VHEHVségre képes (Bloodgood 1990).
(ONpS]HOKHW QHN W QLN D] LV KRJ\ D PR]JiV V]HPSRQWMiEyO SDVV]tY VHMWHN– magas talajvíz esetén - kapilláris vízemeléssel jutnak ismét a talajfelszín köze- lébe.
$YiQGRUOiVOHKHW VpJHpVD]RNDPHJILJ\HOpVHNDKRODPpO\HEEWDlajréte-
JHNEHQHO IRUGXOyDOJiNDIHOV]tQLYDJ\IHOV]tQ-közeli rétegekben is megtalálha-
WyNPHJNpUG MHOH]LN|QiOOyV]XEWHUUiQDOJDIOyUDOpWH]pVpW%ULVWRO-Roach (1927)
pV%HOOYLV]RQWDWDODMV]HOYpQ\ IHOV QpKiQ\ FHQWLPpWHUpEHQ LOOHtve arid területeN YLOiJRVDEE KRPRNN|YHLQ pO DOJDSRSXOiFLyN PLNURUpWHJ]HWWVpJpW Ii-
J\HOWpN PHJ $ ViUJiV]|OG DOJiNDW ÄiUQ\pNNHGYHO N´-nek is mondják, mert
HO IRUGXOiVXN D IHOV]tQ DODWWL UpWHJHNEHQ J\DNRULEE )OLQW FLW 0HWWLQJ
1981). Megfigyelések szerint a talaj kiszáradásával a cianobaktériumok a felszín
DOi K~]yGQDN DKRO PpJ D IRWRV]LQWp]LVKH] HOHJHQG IpQ\ iOO UHQGHONH]pVUH D]
UV-sugárzás viszont lényegesen kisebb. A Microcoleus vaginatus – a talajok
pO EHYRQDWiW DONRWy HJ\LN OHJJ\DNRULEE FLDQREDNWpULXPIDM– nem rendelkezik UV-V] U SLJPHQWHNNHOH]pUWDPLNRUDWDODMWYt]pULDIHOV]tQUHYiQGRUROPDMG
a kiszáradással visszatér a felszín alatti zónába. Rendszeres napi mozgását ten- gerparti homokban már Flint (1958 cit. Metting 1981) is leírta, és a szintén te- lepkpS] Oscillatoria fajok illetve Spirulina subsalsa esetében is megfigyeltek negatív fototaxist 24 órás periódusban (Garcia-Pichel és mtsai 1994 cit. Stal 2000). A Scytonema és Nostoc fajok viszont UV-V] U SLJPHQWMHLN- mint pl. a
NRFVRQ\DKYHO\EHQOpY Virgásbarna színt adó scytonemin - védelmében gyak- ran alkotnak telepet kitett felszíneken.
Davey (1989) a fényintenzitás és stressz-rezisztencia összefüggését figyelte meg Prasiola - antarktiszi zöldalga - esetében, ahol az intenzív megvilágítás növelte a fagyás és kiszáradás károsító hatását. A nagy fényintenzitás más kör- nyezeti stresszorok - például a tápanyaghiány -KDWiViWLVIHOHU VtWKHWL(]DNDp-
FVRODWIRUGtWRWWLUiQ\EDQLVP N|GLNV|WpWEHQWDUWYDDQHKH]HQV]DSRURGyDOJiN
túlélési aránya javul (Reynolds és mtsai 1981 cit. Davey 1989), a fotoinhibíció hatása pedig kifejezettebb másodlagos stresszorok, mint például nitrogénhiány
1HDOHpV5LFKHUVRQYDJ\DODFVRQ\K PpUVpNOHW*HLGHUPHOOHWW
$WDODMDOJiNV]iPiUDDIpQ\KDWiVDPiVWpQ\H] NNHOV]RURVDQ|VV]HNDpcso- lódva jelentkezik. Ezek a talajtípusból adódó szerkezeti tulajdonságok (a pórus-
WHUHN QDJ\ViJD pV HORV]OiVD D] H]W EHIRO\iVROy WDODMP YHOpV D WDODMERUítottság mértéke és a vegetáció, melyek mind meghatározzák a talajba jutó fény mennyi-
VpJpWpVPLQ VpJpWDODSYHW HQEHIRO\iVROYDH]]HOD]DOJiNpOHWpt.
1.3.3. Víz
A talajok mikrobiális életközössége számára a talaj vízpotenciálja az egyik
OHJIRQWRVDEE WpQ\H] 0LQWKRJ\ D WDODMRNDW W|EEQ\LUH YiOWR]y QHGYHVVpJYLV]o-
Q\RN MHOOHP]LN D WDODMDOJiN NO|QE|] NpSSHQ DONDOPD]NRGWDN D] LG V]DNRV
kiszáradáshoz. A FLDQREDNWpULXPRN HJ\ UpV]H I NpQW D Nostocales és Stigonematales rendekbe sorolt fajok) akinéta-képzéssel, a zöldalgák zigospóra ill. hipnospóra formájában, míg mások látható morfológiai változások nélkül képesek elviselni a kiszáradást különféle fiziológiai és biokémiai adaptációs
mechanizmusok révén. Ilyenek például az extracelluláris poliszacharid termelé- se (kocsonyaburok, kocsonyahüvely), a sejtek és fonalak aggregációja, az er VHEE VHMWIDOV]HUNH]HW pV YDVWDJViJ D YDNXyOXP KLiQ\D IpQ\W|U V]HPFVpN ps olajcseppek akkumulációja, vagy a viszkózus protoplazma. Laboratóriumi kí- sérletekben megfigyelték, hogy a baktériumok és protozoák mellett a mikroalgák is inaktiválódnak olyan alacsonyabb vízpotenciál értéken, amelyen
D]pOHV]W - és fonalas gombák még képesek fenntartani anyagcseréjüket (Harris 1981 cit. Metting 1991). Alacsony vízpotenciálú talajban a mozgó alakok hely- változtatása is gátolt.
$ PLNURDOJiN NO|Q|VHQ D FLDQREDNWpULXPRN NLWDUWy DODNMDL KRVV]~ LGHM
kiszáradást képesek elviselni. Bizonyítékként szolgálnak erre, hogy 69-87 éves
KHUEiULXPL J\ MWHPpQ\HNE O pOHWNpSHV DOJDVHMWHNHW WXGWDN L]ROiOQL /LSPDQ
1941 cit. Whitton 2000, Parker és mtsai 1969 cit. Bell 1993, Kondratyeva és Kislova 1999). A cianobaktériumok akinétái hideggel és kiszáradással szemben
UH]LV]WHQVHN K YHO V]HPEHQ YLV]RQW pU]pNHQ\HN 1LFKROV pV $GDPV $]
Anabaena cylindrica akinétái sötét és száraz körülmények között 5 évig életké- pesek maradtak, vegetatív sejtjei ezeket a feltételeket csupán 2 hétig tudták elvi- selni (Yamamoto 1975). A kovamoszatok és sárgászöld algák kevésbé
WROHUiQVDN D Yt]KLiQ\UD (]W EL]RQ\tWMD D] DULG WHUOHWHN WDODMDLEDQ ULWND HO IRr- dulásuk (Lund 1947, Cameron 1964), laboratóriumi kísérletekben pedig a kiszá- radással szemben nagyobb érzékenységük.
A talaj nedvességtartalma és a talajalgák száma közötti összefüggésre töb- ben is rámutattak (Shtina és Bolyshev 1963, Busygina 1976, Shimmel és Darley
$KPHG $] DOJDYHJHWiFLyEDQ EHN|YHWNH] pYV]DNRV YiOWR]iVRN PHQQ\LVpJLMHOOHJ HNDIHOYHKHW Yt]WDUWDORPpVK PpUVpNOHWIJJYpQ\pEHQPtJ
a fajösszetétel éves szinten állandónak mondható (Lund 1947). Tchan és Whitehouse (1953) a változó talajnedvességgel magyarázták vizsgálataikban az alga sejtszám napi ciklusát.
Az arid területek talajalgái számára az egyetlen, viszonylag rendszeresen
KR]]iIpUKHW Yt]IRUUiV D KDUPDW PHO\HW Yt]J ] IRUPiMiEDQ LV NpSHVHN D VHMWHN
anyagcseréjük fenntartásához hasznosítani (Friedmann és Galun 1974 cit. Bell 1993). A szárazság növekedésével a talajok mikrobiális tevékenysége a felszíni rétegekre koncentrálódik (Skuji ã $ 1HJHY VLYDWDJ PLNURILWD WDODMEHYo- natainak vastagsága 1-15 mm között változhat a területre hullott éves csapa-
GpNPHQQ\LVpJW OIJJ HQpVH]]HOHJ\WWYiOWR]LNDEHYRQDWRWDONRWyWiUVXOiVRN
felépítése is: a legszárazabb területeken a bevonatot csupán cianobaktériumok képezik (többségében Microcoleus vaginatus), míg a csapadékosabb területek bevonata cianobaktériumok, zöldalgák, gombák, mohák és zuzmók társulásából áll (Shem-Tov és mtsai 1999).
A talajalJiN V]iPiUD IRQWRV KRJ\ D NH]G G V]iUD] LG V]DNUD LG EHQ UHa- gálni tudjanak kitartó vagy ellenálló alakok képzésével. Evans (1959 cit.
Metting 1981) megfigyelései szerint minél hosszabb a száraz periódus, annál
W|EE LG W LJpQ\HO D] ~MUDQHGYHVHGpV pV D] DNWtY DQ\DJFVHUpM VHMWHN NpS]pVH D
hirtelen közegváltás (a spóra vízbe kerülése) pedig egyenesen gátolhatja a kitar- tó alakok csírázását. A fokozatos kiszáradás tehát nagyobb túlélési esélyt jelent, ahol a sejtek egyéni fiziológiai változásain túl - a kolóniákban és
F|QyELXPRNEDQ pO DOJiN HVHWpEHQ- az elpusztuló perifériális sejtek védelmet
Q\~MWDQDN D EHOMHEE OpY NQHN EL]WRVtWYD H]]HO D SRSXOiFLy IHQQPDUDGiViW ÒMDEENXWDWiVLHUHGPpQ\HNV]HULQWD]DOJiNDWDODMRNNLV]iUDGiVDVRUiQ D] NHW pU R]PRWLNXV Vtresszre a szervezettebb növényekhez hasonlóan reagálnak (Kobayashi és mtsai 1997). Talajlakó zöldalgákkal végzett kísérletben a külön-
E|] VWUHVV]RURN Vy-, sav-, és szárazságstressz) hatására a Stichococcus bacillaris és Chlorella vulgaris tenyészetek abszcizinsav (ABA)-termelése a kezeletlen kontrollhoz képest 5-10-V]HUHVUH Q WW PLQGHPHOOHWW D QDSRV We-
Q\pV]WpVVRUiQPHJILJ\HOKHW YROWD]LVKRJ\ D]$%$-szint a tenyészetek öre- gedésével párhuzamosan emelkedett (Maršálek és mtsai 1992b).
Lényegesen nagyobb az alga sejtszám és a fajgazdagság azokon a területe-
NHQ DKRO D Yt] YLV]RQ\ODJ N|QQ\HQ KR]]iIpUKHW $] DOJDIHMO GpV -60%-os vízkapacitás mellett a legkifejezettebb (Stokes 1940). A cianobaktériumok fej-
O GpVpQHN pV QLWURJpQN|WpVpQHN D -100%-os abszolút talajnedvesség kedvez (Shtina 1972 cit. Zimmerman és mtsai 1980). A vízzel telített talajokban mérhe-
W LQWHQ]tYHEEV]DSRURGiVWpVQLWURJpQN|WpVWXJ\DQDNNRUD]DODFVRQ\R[LGiFiós- redukciós potenciál is indokolhatja (Stewart és Pearson 1970, Fogg és mtsai 1973). Tchan (1953) megfigyelése szerint az eukarióta algák aktivitása viszont
FV|NNHQWD]HU WHOMHVHV ]pVWN|YHW NLVSRWHQFLiOpUWpNQHNN|V]|nKHW HQ + PpUVpNOHW
$ K PpUVpNOHW N|]YHWOHQO D VHMWDQ\DJFVHUpW N|]YHWHWW PyGRQ D WDODM Io- lyadék- pV Ji]QHP DQ\DJDLQDN GLII~]LyMiW pV UHGR[SRWHQFLiOMiW EHIRO\iVROYD
hat a mikroszervezetekre (Stotzky 1972 cit. Metting 1992). A 60 °C -os napi
K LQJDGR]iV~ VLYDWDJL N|UQ\H]HWEHQ D IDJ\RWW WXQGUD-talajokban, a sarkvidék
MHJHVWHUOHWHLQpVDK IRUUiVRNYL]pEHQpO DOJiNMyOV]HPOpOWHWLNKRJ\HPLk-
URV]HUYH]HWHN PLO\HQ WiJ W U NpSHVVpJJHO UHQGHONH]QHN D V]pOV VpJHV K Ppr- sékleti értékek elviseléséhez. Laboratóriumi kísérletekben a Stichococcus, Microcoleus, Protococcus és Schizothrix fajok (Kärcher 1931, Cameron és Blank 1966 cit. Metting 1981) sikeresen túlélték a -195 °C-os, napokon át tartó hideget légszáraz talajban, más algák pedig elviselték a 160 °C-os forróságot
7UDLQRUFLW%HOO$]RNDYL]VJiODWRNPHO\HNQpOMHOHQW VQHWWyIo- toszintézisW PpUWHN D] DOJDWHOHS IDJ\iVD DODWWL K PpUVpNOHWHQ -DPHV FLW
Metting 1981, Davey 1989), a sejtekben krioprotektív anyagok jelenlétére en-
JHGQHNN|YHWNH]WHWQL7HDUOH+DVRQOyHOYHQP N|GQHNHJ\HVW|U]VJ\ j- temények, melyeknél a krioprezervációs és fagyasztva-szárítási eljárásokkal az
DOJiNKRVV]~LGHM WiUROiViWROGRWWiNPHJDEHIHUW ]pVYDJ\iWIHUW ]pVYHV]pO\pW
ezáltal a minimálisra csökkentve (Blackwell és Chapman 1993).
$WDODMDOJiNpOHWpEHQD]H[WUpPK PpUVpNOHWLpUWpNHNOHJW|EEV]|UIL]LROygi- ai szárazsággal társulnak, ami segítséget jelenthet a túlélésben. Vogel (1955 cit.
%HOO NtVpUOHWHL D]W PXWDWWiN KRJ\ D V]pOV VpJHVHQ PDJDV K PpUVpNOHWHW
PHJHO ] NLV]iUDGiVVRUiQD]DOJiNK W U NpSHVVpJHMREEYROWSpongiochloris typica zöldalga esetében egy órán át 100 °C-on szárított talajban egy év eltelté- vel az inokulumhoz képest a sejtszám mindössze két nagyságrenddel csökkent (Trainor és McLean 1964).
$ ]|OGDOJiN pV D FLDQREDNWpULXPRN HOWpU K PpUVpNOHWL RSWLPXPPDO UHn- delkeznek (Venkataraman 1964, Ruble és Davis 1988), és a magasabb értékek többnyire az utóbbi algacsoportnak kedveznek. Davey (1989) antarktiszi tala- jokból izolált Phormidium faj és Prasiola faj összehasonlító laboratóriumi vizs- gálata azt eredményezte, hogy a többszöri fagyasztást és felengedést köveW HQD
zöldalga jobb életképességet mutatott, ami alacsonyabb megvilágítás mellett
Q|YHOKHW YROW $ FLDQREDNWpULXP Yt]W|EEOHW PHOOHWW PXWDWRWW MREE W~OpOpVL
arányt, a dehidratációra toleráns és a fagyással szemben érzékeny volt, ellenke-
] OHJD]|OGDOJiYDO. Burke és mtsai (1976 cit. Davey 1989) feltételezik, hogy a fagyás folyamata extracellulárisan zajlik a zöldalga esetében, míg intracellulárisan a cianobaktériumnál, amely a sejtre nagyobb veszélyt jelent.
1.3.5. pH
$]LRQL]iOyWpQ\H] NpVDS+WpQ\OHJHVszerepének megítélése nehéz, mert
W|EEQ\LUH PiV KDWiVRNNDO HJ\WW SO WiSHOHPHN YiOWR]y IHOYHKHW VpJH UpYpQ
közvetetten befolyásolják a talajok algaflóráját. Az arid talajokat általában lúgos
NpPKDWiV pV QDJ\REE LRQHU VVpJ MHOOHP]L PtJ D FVDSDGpNRV pJKDjlaton kiala- kult talajok többnyire savanyúak, a viszonylagosan nagy agyagtartalom követ-
NH]WpEHQ SHGLJ J\HQJH NDWLRQ NLFVHUpO NpSHVVpJ pV– különösen foszforra és egyes mikroelemekre - kisebb tápanyag-V]ROJiOWDWyNpSHVVpJMHOOHP]L NHW
Az algák kémhatás szempontjából szinte valamennyi területet meghódítot-
WiND)|OG|Q(J\HVIDMDLND]HU VHQVDYDVN|UQ\H]HWKH]DONDOPD]NRGWDNS+
Cyanidium caldarium, Doemel és Brock 1971 cit. Lukešová és Hoffmann
PtJ PiVRN HU VHQ O~JRV N|]HJEHQ WDOiOWiN PHJ pOHWIHOWételeiket (pH=13,5 Spirulina platensis, Soeder és Stengel 1974 cit. Lukešová és Hoffmann 1995). Általánosan elfogadott nézet, hogy a cianobaktériumok legin- kább a semleges és gyengén lúgos talajokat kedvelik (Shields és Durrell 1964, King és Ward 1977, Komáromy 1982), a zöld- és sárgászöld algapopulációk
GRPLQDQFLiMD SHGLJ VDYDQ\~ WDODMRNEDQ ILJ\HOKHW PHJ 6KLHOGV pV 'XUUHOO
1964, King és Ward 1977, Lukešová 1993), ami a pH-val szembeni nagyobb
W U NpSHVVpJNNHO LV PDJ\DUi]KDWy +XQW pV PWVDL /XNHãRYá és Hoffmann (1995) vizsgálataiban hét – savanyú talajból izolált – zöldalga faj szaporodása mégis semleges (pH=7) pH-pUWpNQpO PXWDWNR]RWW HJ\|QWHW HQ
jobbnak a savasabb kezelésekhez képest. Ez azt bizonyítja, hogy a zöldalgák a savas kémhatást sokkal inkább tolerálják, mint kedvelik.
A pH szoros korrelációban van a talaj kalcium-karbonát tartalmával. Meg- figyelések szerint a talaj meszezése növeli az alga sejtszámot, a diverzitást, ezen belül is a cianobaktériumok arányát (Stokes 1940, Sukala és Davis 1994, Lukešová és Hoffmann 1996). Ezzel szemben Komáromy (1975) savanyú tala- jokban mért nagyobb egyed- és fajszámot.
Egyes talajlakó cianobaktérium, zöldalga és kovamoszat fajok jól tolerálják a nag\ LRQHU VVpJHW 7|EE PHJILJ\HOpV HPOtW JD]GDJ FLDQREDNWpULXP-flórát sós és félsivatagos területeken (Durrell 1962, Ali és Sandhu 1972). Érdekes megfi- gyelést tett Jaag (1945 cit. Büdel és mtsai 1994), miszerint a talajlakó és epilita cianobaktériumok vöröses színét okozó gloeocapsin savas közegben vörössé,
O~JRVN|]HJEHQV]tQWHOHQQpYDJ\NpNHVV]tQ YpYiOLN
1.3.6. Tápanyagok
$]HO ] HNE OPiULVPHUWD]D]iOWDOiQRVQp]HWKRJ\VDYDQ\~EEWDODMRNEDQ
a zöldalgák, alkáli talajokban a cianobaktériumok képeznek többséget. Lund (1947) ezt azzal magyarázta, hogy a kovamoszatok és cianobaktériumok számá-
UDO~JRVN|]HJEHQDV]HUYHVpVV]HUYHWOHQDQ\DJRNN|QQ\HEEHQIHOYHKHW N
A szervesanyag-tartalom nagymértékben meghatározza a talajok algaflórá- jának összetételét és dominanciaviszonyait. Erre a következtetésre jutott Bristol- Roach (1927) és Stokes (1940) a szervestrágyázás hatását vizsgálva. Friedmann és Galun (1974 cit. Metting 1981) természetes körülmények között tapasztaltak nagyobb algadiverzitást a nagyobb szervesanyag-tartalmú mikrokörnyezetben.
Érdemes azonban megjegyezni, hogy ezekben az esetekben a hatás közvetve a szervesanyag jobb vízvisszatartó képességével is magyarázható, ráadásul a na- gyobb humusztartalom következtében sötétebb talajok gyorsabb felmelegedése is közismerten serkenti a mikrobiológiai aktivitást. Magyarország homoktalaja- in végzett vizsgálataiban Komáromy (1979) a cianobaktériumok dominanciáját
tUWDOHPtJDV]HUYHVDQ\DJEDQJD]GDJWDODMYDJ\V]HUYHV|VV]HWHY NHWWDUWDOPa- zó dúsító tenyészet az ostoros formák (Euglenophyta) elszaporodását segítette
HO 3ULQJVKHLPpVHGLYt]LyWDJMDLQDNW|PHJSURGXNFLyMDYROWPHJILJ\Hl-
KHW WUiJ\DOpYHO V]HQQ\H] G|WW V]LNHV YL]HNEHQ LV .LVV 9UDQi pV
Votruba (1995) laboratóriumi kísérletei hasonló eredményt hoztak: az oldható humuszsav és humát a zöldalgák szaporodását serkentette, a cianobaktériumokra pedig hatástalan volt. Acea és Carballas (1999) kísérletei- ben viszont a talajjal kevert búzaszalma és baromfitrágya gátolta a talajalgák
IHMO GpVpW
Makroelemekkel (C, N, P, Ca, Mg, K, S) végzett táplálási kísérletek azt
PXWDWWiN KRJ\ D WDODMDOJiN pOHWIRO\DPDWDLEDQ D WiSHOHPHN DODSYHW HQ KDVRQOy
szerepet töltenek be, mint az edényes növényeknél (O'Kelley 1974 cit. Pipe és
Shubert 1984), és a tápanyagfelvétel is lényegében azonos folyamatok útján játszódhat le.
A nitrogén az egyik legfontosabb esszenciális elem, cianobaktériumokban mennyisége a szárazsúly 10%-iW LV HOpUKHWL 7HUPpV]HWHV N|UQ\H]HWNE O D Wa- lajalgák többnyire szervetlen formában (légköri dinitrogén, nitrát és ammóniumsók, kisebb mennyiségben nitrit), illetve szerves nitrogéntartalmú
YHJ\OHWHN IRUPiMiEDQ XUHD pV DPLQRVDYDN YHV]LN IHO PHO\HN HO V]|U D VHMt- ben ammóniává alakulnak, ami aztán az anyagcsere-utakra kerül. Ennek megfe-
OHO HQD]1+4
+-1 D]DOJDVHMWHNV]iPiUDDOHJN|QQ\HEEHQIHOYHKHW NO|Q|VHQ
pH>7 kémhatású közegben és gyengébb fényviszonyok mellett. A cianobaktériumok sejtjeiben fénymikroszkóp alatt - festés nélkül is - jól látható cianoficin szemcsék nitrogénraktárként szolgálnak D NHGYH] WOHQ WiSDQ\DJ- ellátottság esetére, és egyes fajok (Spirulina sp., Anabaena sp.) lebomló fikocianin-tartalmukat is képesek hasznosítani. Antarktiszi talajok algapopulá-
FLyLQDN NpV Q\iUL LG V]DNEDQ PHJILJ\HOW FV|NNHQpVH QHP D Yt]KLiQ\QDN Ka- nem sokkal inkább a talajok kis NO3-
-N és NH4+
-N koncentrációjának, illetve az alacsony N/P aránynak tudható be. Davey és Rothery (1992) az algák N-limitált
IHMO GpVpYHOPDJ\DUi]WiN D]WLVKRJ\DV]DSRURGiVLLG V]DNU|YLGHEEPLQWD]W DK PpUVpNOHWLpUWpNHNOHKHW YpWHQQpN$WiSROGDWRVDOJDWHQ\pV]WpVLVW|EEQ\LUH
N-OLPLWiOW XJ\DQDNNRU D W~O VRN IHOYHKHW QLWURJpQ pV IRV]IRU JiWROKDWMD D] Dl- gák szaporodását a talajban (Lund 1947). A cianobaktériumok légköri N-kötése – a heterociszta-NpS] GpV pV D QLWURJHQi]-szintézis gátlása révén – MHOHQW VHQ FV|NNHQDPPyQLXPpVQLWUiWMHOHQOpWpEHQ$QDJ\PHQQ\LVpJEHQIHOYHKHW Nö-
W|WWQLWURJpQQHNDFLDQREDNWpULXPRNIHMO GpVpUHJ\DNRUROWQHJDWtYKDWiViUDUL]s-
I|OGHNP WUiJ\i]iVDVRUiQILJ\HOWHNIHO:KLWWRQ
$] pGHVYL]HNW O pV WHQJHUHNW O HOWpU HQ D WDODMRN yULiVL IRV]IRUNpV]OHWWHO
rendelkeznek, nagyrészt gyengén oldható ásványi foszfátok (pl. kalcium- foszfát), illetve nagy molekulasúlyú szerves anyagok formájában. Az algák számára azonban csak az oldható ortofoszfát, illetve a kis molekulasúlyú szer-
YHV IRV]IiWRN KDV]QRVtWKDWyN 0LQWKRJ\ D IRV]IRU IHOYHKHW VpJpW LOO ROGKDWyVá-
JiWV]iPRVN|UQ\H]HWLWpQ\H] tJ\DWDODMS+-értéke, nedvesség- és nitrogéntar- talma, Ca2+, Mg2+, Fe2+, Fe3+ és Al3+ koncentrációja is befolyásolja, ismert a foszfor puffer-szerepe, továbbá faji eltérések is vannak, ezért megleheW VHQ
nehéz az optimális foszfor-szükségletet megállapítani. Ehhez még az is hozzájá- rul, hogy a cianobaktériumok polifoszfát testecskéiben, és a többi divíziók tag- jainak (Euglenophyta, Chlorophyta) sejtjeiben a környezet foszfor- koncentrációjának akár több ezerszeresét is képesek tartalékolni. Egy átlagos cianobaktérium sejt száraz tömegének – normális ellátottság mellett - csupán 0,6%-át teszi ki a foszfor. Az azonban nem vitatott, hogy a közeg foszfor- ellátottsága meghatározó lehet a talajalgák IHMO GpVpUH .LQJ pV :DUG
vizsgálataikban a foszfort találták a legfontosabb limitáló tápelemnek. Hiánya
VWLPXOiOMD OHJW|EE FLDQREDNWpULXPQiO D] DNLQpWiN NpS] GpVpW :RON 6XWKHUODQG pV PWVDL IRV]IiWDGDJROiV PHOOHWW SHGLJ HU WHOMHVHEE V]aporo-
GiV pV IRNR]RWW QLWURJpQN|WpV YROW PpUKHW UL]VNXOW~UiN WDODMDLEDQ :LOVRQ pV
Alexander 1979, Roger és mtsai 1987).
A ként a talajalgák többnyire szulfátok és tioszulfátok formájában veszik fel, de néhány faj (Chlorella sp., Euglena sp., Synechococcus sp.) esetében ki-
PXWDWWiN KRJ\ PHJIHOHO N|UOPpQ\HN N|]|WW NpSHVHN V]HUYHV 6-tartalmú ve- gyületek, mint cisztein, cisztin, esetleg glutation és tiocianát felvételére is. Itt érdemes újból megemlíteni a dolgozat elején leírtakat, miszerint egyes algafajok képesek kemoheterotróf életmódra áttérni sötétben, szulfidot használva elekt- rondonorként, elemi kén felszabadulása mellett. A szulfát felvétele aktív folya-
PDWPHO\HWDK PpUVpNOHWpVDS+EHIRO\iVROpVJiWOyGKDWV]XOILWLOOWLRV]XOIiW
jelenlétében. Laboratóriumi kísérletekben az eukarióta talajalgák nagyobb érzé- kenységet mutattak szulfit-stressz (100mM) hatására, mint a cianobaktériumok (Miszalski és mtsai 1995).
Hunt és mtsai (1979) szoros korrelációt mutattak ki a talajalgák mennyisé-
JHpVDIHOYHKHW Wápelem-WDUWDORPN|]|WWV]iQWyI|OGLNtVpUOHWHNEHQDP WUiJ\á-
]iV HJ\pUWHOP HQ Q|YHOWH D WDODMDOJiN PHQQ\LVpJpW pV D IDMJD]GDJViJRW 6KWLQD
1969, 1991; Sukala és Davis 1994). Eukarióta talajalgák tenyésztésénél a táp- anyagoknak lényeges szerepe van a motilis sejtek differenciálódási folyamatai- ban is. Ezt figyelték meg O’Kelley és Deason (1962 cit. Metting 1981) a Protosiphon botryoides zoosporogenezise során, ahol a tápanyagok és a fény indukálták, a nitrogén, kén és kalcium hiánya pedig gátolta a zoospórák képzé- sét. Két Anabaena törzset vizsgálva Rodríguez és mtsai (1990) kimutatták, hogy aerob környezetben, és kötött nitrogén jelenlétében kalciumra csupán nyom- elem-mennyiségben van szüksége a cianobaktériumnak; ugyanakkor nem mér-
KHW V]DSRURGiV KD D QLWURJpQIRUUiVW HJ\HGO D OpJN|UL GLQLWURJpQ NpSH]L
Feltételezik, hogy a kalciumnak nem a nitrogén-fixációnál van katalizáló hatása,
KDQHP VRNNDO LQNiEE QLWURJHQi]W YpG V]HUHSH OHKHW D NLDODNXOy
heterocisztákban.
Az esszenciális mikroelemek, és szükséges mennyiségük ismerete az alga- tenyésztésben jut kiemelt szerephez. Egy általános tápoldat összetételében sze- repel a mangán, nikkel, cink, bór, vanádium, kobalt, réz, és molibdén. A nehézfémek esetében gyakran igen kis különbség van kedYH] pVPpUJH] Gy]LV N|]|WWH]pUWDWDODMDOJiNUDNO|Q|VHQYHV]pO\HVKRJ\DWDODMRNEDQLG V]DNRVDQ NLDODNXOyDQR[LNXVIHOWpWHOHNSO0QHVHWpEHQLOOHWYHDS+NHGYH] WOHQYiOWo-
]iVDL D WDODMVDYDQ\RGiVVDO Q D NDWLRQRN IHOYHKHW VpJH pV PR]JpNRQ\ViJD D WDODMEDQ D V]NVpJHVQpO QDJ\REE IHOYpWHONHW VHJtWLN HO -HOOHP] KRJ\ D Wo-
[LNXVKDWiVU|YLGLGHLJLVIHQQiOOKDWSOKLUWHOHQHV ]pVNRUYt]]HOWHOtW G|WWSó-
UXVWHUHN YDJ\ SRQWV]HU KDWiVD OHKHW SO FVXSiQ QpKiQ\ KDWiURV WDODMV]HPFVH
mikrokörnyezetében érvényesül). A túlzott mennyiségben rendelkezésre álló
IpPHND]DOJiNIHMO GpVpEHQPRUIROyJLDLIL]LROyJLDLpVELRNpPLDLYiOWR]iVRNDW
okozhatnak. A kadmium és cink nagyobb koncentrációi gátolhatják az utódsej- tek leválását, ezáltal fonalas alakot kölcsön|]YH D QRUPiOLVDQ HJ\VHMW
cianobaktériumoknak, vagy hatásukra megváltozhat a pigment-összetétel úgy,
KRJ\ PHJQ D NDURWLQRLGNORURILOO DUiQ\ pV FV|NNHQ D ILNRFLDQLQ PHQQ\LVpJH
Egyéb ionok nagy koncentrációja, mint például a Ca2+ vagy PO43-MHOHQW VPpr- tékben csökkentheti a cink toxicitását cianobaktériumokra. A kadmium vagy réz
W~O]RWWNRQFHQWUiFLyMDKDWiViUDSHGLJMHOHQW VHQFV|NNHQYDJ\PHJLVV] QKHWD
fotoszintézis (Bednarz és Starzecka 1992, Robinson és mtsai 2000). Az arzén, kadmium, nikkel, ólom, palládium és cink kis koncentrációban (0,005-0,025 ppm) általánosan serkentették Nostoc fajok esetében a nitrogén-kötést; az As, Ni és Pd nagyobb dózisainál (0,025-0,125 ppm) is ez volt tapasztalható, míg a Cd, Pb és Zn ilyen mennyiségben már gátló hatást mutattak. A vizsgálatból az is
NLGHUOW KRJ\ D] pU]pNHQ\VpJ KDWiUD IDMRQNpQW HOWpU +HQULNVVRQ pV 'D6LOYD
1978). Phormidium fragile tenyészetekben a biomassza, a fehérjetartalom, klo- rofill-D pV NDURWLQRLGRN PHQQ\LVpJH D Q|YHNY KLJDQ\-koncentrációval fokoza- tosan csökkent, míg a nehézfém felvéteOHXJUiVV]HU HQPHJQ WW.KDOLO
0LQWDUUyODK PpUVpNOHWLKDWiVRNDWWiUJ\DOyIHMH]HWEHQPiUV]yHVHWWHJ\HV DOJiN V]pOV VpJHV N|UQ\H]HWL IHOWpWHOHNKH] LV NpSHVHN DONDOPD]NRGQL 7|EE
megfigyelés szól arról, hogy nehézfémekkel természetes úton (pl. ércbányák kitermelési vonalán) vagy szándékosan szennyezett talajon kiterjedt cianobaktérium bevonatokat találtak, ahol a kis fonalvastagságú Oscillatoriales rend tagjai (Plectonema és Schizothrix fajok) domináltak. Nostoc commune
WHOHSHNHW Q\HUVRODMMDO pV N RODMV]iUPD]pNRNNDO WHUKHOW WDODMRNEDQ PXWDWWDN NL
(Whitton 2000).
$WDODMDOJiNPDJXNLVHO VHJtWKHWLNDWiSDQ\DJRNMREEIHOYHKHW VpJpWSpl-
GiXO V]HUYHV VDYDN N|UQ\H]HWEH ERFViWiViYDO NHOiWNpS] N pV HQ]LPHN WHUPHOé- sével. Zöld- és sárgászöld algák extracelluláris enzim-produkciójáról szemle- cikkében Metting (1981) közöl összefoglaló táblázatot.
$WDODMDOJiNPHQQ\LVpJLHO IRUGXOiVD
1.4.1. Sejtszámlálási módszerek
Azért, hogy valós képet alkothassunk a talajalgáknak az edafonban betöl-
W|WW V]HUHSpU O D WiUVXOiVRN |VV]HWpWHOpQHN pV P N|GpVpQHN YL]VJiODWiQ W~O PHQQ\LVpJLPHJKDWiUR]iVRNUDLVV]NVpJYDQ7DODMDOJiNPHQQ\LVpJLHO IRUGu- lásával csupán a múlt század elején kezdtek el foglalkozni (Robbins 1912, Esmarch 1914 cit. Fehér 1954), és a vizsgálati módszerek folyamatos fejleszté-
VpQHN N|V]|QKHW HQ D WDODMRN DOJDSRSXOiFLyMiQDN V]iPV]HU VtWpVpUH MHOHQOHJ W|EE PpUpVL OHKHW VpJ LV DGRWW &ODUN pV 'XUUHOO 6KLHOGV 0HWWLQJ
1994, Gärtner 1996, Whitton 2000):
A) Közvetett módszerek a sejtszám becslésére
• Hígított tenyészetek módszere: a talajmikrobiológiában közismert sejt- számlálási módszer, amelynél ismert hígítású talajoldatok agarlemezen
IHMO G DOJDWHOHSHLQHNV]iPiEyON|YHWNH]WHWQHNDNLLQGXOiVLVHMWV]iPUD$
módszer HO Qyei KRJ\ D NXOW~UiN pUWpNHOpVpUH NpV EE LV OHKHW VpJ YDQ H]HQW~OPLQ VpJLYL]VJiODWRNUDpVL]ROiOiVUDLVIHOKDV]QiOKDWyNDV]pOHVz- tett tenyészetek. Hátrányai: eszköz-, munka- pV LG LJpQ\HV D IHOKDV]QiOW
tápoldat összetétele, valamint a tenyésztési körülmények nagyban befo-
O\iVROMiN D IHMO G WHOHSHN IDM|VV]HWpWHOpW QDJ\REE KtJtWiVQiO HJ\HV EDk- térium- és gombafajok uralkodóvá válnak, ezzel lassítják az algák
IHMO GpVpW PtJ D] pU]pNHQ\HEE DOJDQHP]HWVpJHNHW HO LV Q\RPKDWMiN D]
antibiotikummal dúsított tápközeg használata viszont korlátozza a
FLDQREDNWpULXPRN IHMO GpVpW $ PyGV]HU DODSHOYHNpQW PHJIRJDOPD]RWW
"egy sejt = egy telep" feltételezés a fonalas és a cönóbiumot vagy telepet
NpSH] IDMRNHVHWpEHQQHPiOOMDPHJDKHO\pWDPLMHOHQW VHQFV|NNHQWLD
módszer pontosságát.
• MPN (Most Probable Number) - módszer (Allen, 1957): hasonlóan az
HO EE LVPHUWHWHWW PyGV]HUKH] LWW LV LVPHUW KtJtWiV~ WDODMROGDW D NLLQGXOiVL DODS D]]DO D NO|QEVpJJHO KRJ\ D WHQ\pV]HWHN WiSROGDWEDQ IHMO GQHN $ IHQWHEE IHOVRUROW HO nyök és hátrányok e módszerre is érvényesek. Meg-
MHJ\]HQG KRJ\ D &)8-módszernél használt agarral szilárdított tápoldat
WHUPpV]HWHVHEEN|]HJHWEL]WRVtWDWDODMDOJiNIHMO Géséhez.
• Klorofill-DH[WUDNFLyVPyGV]HUGXUYDEHFVOpVLPyGV]HUD]RQEDQHJ\V]HU
és gyors. Azt veszi alapul, hogy a talaj obligát heterotróf mikroszervezete-
LW OHOWpU HQPLQGD]HXNDULyWDDOJiNPLQGSHGLJDFLDQREDNWpULXPRNVHMt- jei tartalmaznak a-NORURILOOW DPL VSHNWURIRWRPpWHUUHO PpUKHW $]
extrakcióhoz több kivonószer is használható, nemzetközileg elfogadott az etanolos módszer (egyes vélemények szerint azonban a metanolos és ace- tonos kivonás cianobaktériumoknál nagyobb pontosságot ad). A módszer
KiWUiQ\DKRJ\QHPWHV]LOHKHW YpD]HJ\HVWD[RQyPLDLFVRSRUWRNHlkülö- nítését, a talajok növényi maradványai pedig torzíthatják az eredményt (Tsujimura és mtsai 2000). Mint biomassza index meglehet sen pontatlan annak következtében, hogy a sejtek klorofill tartalma jelenW VPpUWpN Ia-
MRN N|]|WWL HOWpUpVW PXWDW IDMRQ EHOO SHGLJ D IHMO GpVL Vtádium és a kör- nyezeti ténye] NQDJ\EDQEHIRO\iVROMiNDPHQQ\LVpJpW
B) Közvetlen sejtszámolási módszerek
• Fénymikroszkópos sejtszámlálás talajminta vizes oldatából, számláló (Bürker)–kamra vagy kalibrált tárgylemez segítségével. Az optikai mód- szerek HO Qyei:DV]HUYH]HWHNWHQ\pV]WpVLN|UOPpQ\HNWiSN|]HJK Ppr- séklet, fény stb.) okozta szelekciója, valamint a kultúrakészítés eszköz-,
LG -pVPXQNDLJpQ\HVP YHOHWHHOPDUDGHátrányai: a vízmintákkal ellen- tétben a talajoldat átlátszatlan mivolta megnehezíti az algasejtek felismer-
KHW VpJpW %UNHU-kamrában a fonalas alakok nehezen, vagy egyáltalán
nem számolhatók. A minta tárolhatósági ideje korlátozott, a vizsgálatot mihamarabb el kell végezni.
• Klorofill-autofluoreszcencián alapuló sejtszámlálás (Tchan 1952 cit.
Oesterreicher 1988): a talajok aktuális algatartalmának gyors és nagy pon- tosságú sejtszám-meghatározási módszere. Elve: fluoreszcens mikroszkóp alatt a talajszuszpenziót megvilágítva a klorofillt tartalmazó sejtek (eukarióta algák és cianobaktériumok egyaránt) vörös autofluoreszcenciát
PXWDWQDN PtJ D WDODM HJ\pE |VV]HWHY L iVYiQ\L DQ\agok és szerves tör-
PHOpNHNHJ\iOWDOiQQHPJHUMHV]W GQHNtJ\DV|WpWKittérben nem látsza-
QDN YDJ\ D Y|U|VW O HOWpU V]tQW HPLWWiOQDN SO OiVG iEUD 5RVW
1992). A vizsgálathoz legalább 200-szoros nagyítás javasolt a kisebb sej-
WHNpV]RRVSyUiNpV]UHYHKHW VpJHPLDWW(O Q\HL már egészen kis sejtek is
MyOOiWV]DQDNDVHMWDJJUHJiWXPRNN|QQ\HEEHQIHOLVPHUKHW NpVtJ\DVHMt-
V]iP MREEDQ EHFVOKHW ]|OG -580 nm) és kék (450-490 nm)
JHUMHV]W IpQ\ Yáltott alkalmazásával a cianobaktériumok biztonsággal
PHJNO|QE|]WHWKHW ND]HXNDULyWDDOJDVHMWHNW ODIRQDODV-HJ\VHMW - és a
WHOHSHW YDJ\ F|QyELXPRW NpSH] IDMRN NO|Q-külön is leszámolhatók, az
HUHGPpQ\HN SHGLJ |VV]HYHWKHW N D N|]|V V]iPROiVL DODSQDN(sejtszám)
N|V]|QKHW HQ)pQ\PLNURV]NySHJ\LGHM KDV]QiODWiYDODGRPLQiQVIDMRN
nemzetségek azonnal regisztrálhatók. Mindezek ismeretében a talaj alga-
ELRPDVV]iMDLVVRNNDOSRQWRVDEEDQEHFVOKHW $PyGV]HUDVHMWHNpOHWNé-
SHVVpJpU OLVNpSHWDGPHUWD]HOpusztult sejtek lebomló klorofill-tartalma gyengébb intenzitással vagy egyáltalán nem fluoreszkál. Hátrányai: a flu-
RUHV]FHQVPLNURV]NySL]iOiVDV]HPHWHU VHQLJpQ\EHYHV]LH]EHKDWiUROMD D] HJ\ DONDORPPDO PpUKHW PLQWiN V]iPiW D PpUpV SRQWRVViJiW D
talajszuszpenzió homogenitása nagymértékben befolyásolja; a klorofill- extrakciós módszerhez hasonlóan csupán a mintavételkor aktív anyagcse-
UpM VHMWHNpUWéNHOKHW N7VXMLPXUDpVPWVDL
$ODSYHW HQ YDODPHQQ\L VHMWV]iPOiOiVL PyGV]HU HVHWpEHQ D] HUHGPpQ\HN
meJEt]KDWyViJiQDNEL]WRVtWpNDDPHJIHOHO PLQWDYpWHODN|UOWHNLQW PLQWDNe-
]HOpVpVDOHKHW OHJSRQWRVDEEDQNLYLWHOH]HWWVHMWV]iPOiOiV$WDODMDOJiNJ\ MWé- sének módszerét minden esetben a vizsgálat célja határozza meg (Metting 1994, Lukešová 1993, Lepossa 1999). Általános szabályai természetesen vannak, ezek a statisztikai elemzésekkel alátámasztott számban, random elvégzett mintavétel,
VWHULOHV]N|]|NKDV]QiODWDSRQWRVMHO|OpVpVPHJIHOHO PLQWDWiUolás.
$ ILWRHGDIRQ ELRPDVV]iMD PHJEHFVOKHW D] HJ\VpJQ\L térfogatú vagy tö-
PHJ WDODMUD YRQDWNR]WDWRWW VHMWV]iPEyO pV D] DOJDVHMW iWODJRV WpUIRJDWiEyO $]
LURGDORPEDQIHOOHOKHW DOJDVHMWV]iP-adatok összehasonlítása azonban meglehe-
W VHQ QHKp] D V]iPROiVL HJ\VpJ pV DONDOPD]RWW PyGV]HUHN VRNIpOHVpJH N|YHt- keztében (1.3. táblázat).
Két közvetett mérési módszer összehasonlítása során Tsujimura és mtsai (2000) szignifikáns korrelációt mértek a kolóniák számának és a klorofill-a
NRQFHQWUiFLyORJDULWPXVDLN|]|WWP YHOHWOHQWHUOHWHQ0 YHOWWDODMRQH]DNDp- csolat nem volt bizonyítható, amit részben a klorofill-extrakciós módszernek a növényi maradványok okozta túlbecslésével magyaráztak. A hígított tenyésze- tek módszerét pedig a talajalgák mennyiségi változásainak inkább térbeli, mint-
VHP LG EHOL YL]VJiODWiUD DMiQOMiN $ N|zvetett és közvetlen sejtszámolások eredménye akár 44%-os eltérést is mutathat (Tchan és Whitehouse 1953). Mód- szer-összehasonlító munkájában Oesterreicher (1988) arra a megállapításra
MXWRWW KRJ\ PHJIHOHO N|UOPpQ\HN N|]|WW HOYpJH]YH D KtJtWRWW WHQ\pV]HWek módszere és a fluoreszcens mikroszkópos számlálás eredménye nem különbö-
]|WW MHOHQW VHQ QRKD N|YHWNH]HWHVHQ QDJ\REE VHMWV]iPRW PpUW D] XWyEEL Pyd-
V]HUUHO9L]VJiODWDLEDQUiYLOiJtWDUUDKRJ\DSRUy]XVDEEV]HUNH]HW WHUMHGHOPHV
nyershumusz talajok nem hasonlíthatók össze tömör talajokkal, amennyiben –
DPLQW D OHJW|EE V]HU] QpO V]HUHSHO– az összehasonlítás alapját a talaj száraz súlya adja, és javasolja a sejtszám-pUWpNHNWDODMWpUIRJDWKR]W|UWpQ YLV]RQ\tWiViW
1.3. táblázat:7DODMDOJiNPHQQ\LVpJLHO IRUGulása
Sejtszám (103 sejt/g talaj)
Biomassza
NJpO W|PHJ
/ha)
Mélység
(cm) *\ MW KHO\
Sejtszámlá- lási módszer
Irodalom
2,8-36,5 (Ch+D) 1,1-15 búza monokultúra trágyázási tartamkísér- letben, kezeletlen talaj, Anglia
0,09-105,5 (Ch+D) 0,03-41
0-2,5
búza monokultúra trágyázási tartamkísér- letben, szerves trágyázott talaj, Anglia
MPN Bristol Roach, 1927
0,17-59 0,5-139 NXNRULFDP YHOpVDOyOLVWiOOy- és PK- trágyázott agyagtalaj, USA
0,03-62,5 0,07-147 NXNRULFDP YHOpVDOyOLVWiOOy- és PK- trágyázott, meszezett agyagtalaj, USA 4,8-85,4 11-202 NXNRULFDP YHOpVDOyONH]HOHWOHQDJ\DJWa-
laj, USA
2,3-212 5,4-499 NXNRULFDP YHOpVDOyOPHV]H]HWWDJ\DJWa- laj, USA
6->230 14->542 NXNRULFDP YHOpVDOyO13.-trágyázott agyagtalaj, USA
28,3->230 67->542
0-15
NXNRULFDP YHOpVDOyO13.-trágyázott, meszezett agyagtalaj, USA
MPN Stokes, 1940
<2-4,8 <1,6-3,7 NH]HOHWOHQSRG]RORVEDUQDHUG WDODM2Uosz- ország
69-117 54-92 N-WUiJ\i]RWWSRG]RORVEDUQDHUG WDODM
Oroszország
45-264 35-207 PHV]H]HWWSRG]RORVEDUQDHUG WDODM2Uosz-
ország
24-287 19-225
0-5
N-trágyázott és meszezett, podzolos barna
HUG WDODM2URV]Rrszág
? Balezina, 1975
1.3. táblázat folytatása
Sejtszám (103 sejt/g talaj)
Biomassza
NJpO W|PHJ
/ha)
Mélység
(cm) *\ MW KHO\
Sejtszámlá- lási módszer
Irodalom
0-0,18 (C) <0,001 P YHOHWOHQKXPXV]RV|QWpVWDODMRQ86$
0,05-1600 (C) 0,0003-12 0-0,5
humuszos öntéstalaj, tavaszi búza alól, USA MPN Zimmerman és mtsai, 1980 220,8 35 szántott talaj, trágyakezelés nélkül, Oroszor-
szág
501,2 79 szántott talaj, N60P60K60 trágyakezeléssel, Oro- szország
202,4 32 szántott talaj, N120P120K120 trágyakezeléssel, Oroszország
634,8 100 VHNpO\P YHOpV WDODMWUiJ\DNH]HOpVQpONO
Oroszország
1426 224 VHNpO\P YHOpV WDODM160P60K60 trágyakezelés- sel, Oroszország
961,6 151
0-1
VHNpO\P YHOpV WDODM1120P120K120 trágyakeze- léssel, Oroszország
?
Tret'yakova és mtsai, 1989 (cit. Shtina, 1991)
0-2200 <1728 0-5 OHFVDSROWW ]HJOiS2URV]RUV]iJ ? Busygina, 1976
27,4-214,8 (0-3 cm) 22-101 eUG 8NUDMQD
116,3-730 92-574 réti növénytársulás, Ukrajna
1105 868 W ]HJOiS8NUDMQD
64-213,3 50-168 sztyepp, Ukrajna
16,6-298,8 12-234
0-5
P YHOWFVHUQR]MRPWDODM8NUDMQD
? Sudakova, 1977
<60 <48 drénezetlen glejes podzol talaj, Oroszország
<220 <173 drénezett glejes podzol talaj lazítás nélkül
<400 <315
0-5
drénezett és lazított glejes podzol talaj
? Kondakova, 1984 (cit. Shtina, 1991)
1.3. táblázat folytatása
Sejtszám (103 sejt/g talaj)
Biomassza
NJpO W|PHJ
/ha)
Mélység
(cm) *\ MW KHO\
Sejtszámlá- lási módszer
Irodalom
5-208 <340 podzol talajok, Oroszország
5-80 <82 láptalajok, Oroszország
52-300 <445 réti öntéstalajok, Oroszország
48 16 Ny-szibériai csernozjom talajok, Oroszo.
2150 187 sötét gesztenyebarna talajok, Oroszország
988 515 réti szolonyec talajok, Oroszország
392-5146 74-429
0-10
V]W\HSSHVHG UpWLV]RORQ\HFWDODMRN2Uoszo.
? Shtina, 1959
0,08-4,6 (C) 0,07-3,6
0,17-28,12 (Ch) 0,1-23 kezeletlen savanyú homoktalaj 0,23-91,2 (C) 0,2-71
0,79-91,2 (Ch) 0,6-71
0-5
meszezett savanyú homoktalaj
MPN Sukala és Davis, 1994
54,159 8,5 golfpálya, USA
37,385 5,9 pázsit, USA
35,069 5,5 mocsaras mélyföld, USA
14,68 2,3 útmenti terület, USA
5,736 0,9
0-1
pagony, USA
Hígított tenyészetek
módszere
King és Ward, 1977
2,3 0,3 P YHOHWOHQWHUOHW.D]DKV]WiQ
150 24 0-1
OXFHUQDP YHOpV.D]DKV]WiQ
klorofill-a meghat.
Tsujimura és mtsai, 2000
1.3. táblázat folytatása
Sejtszám (103 sejt/g talaj)
Biomassza
NJpO W|PHJ
/ha)
Mélység
(cm) *\ MW KHO\
Sejtszámlá- lási módszer
Irodalom
524-46770 83-7346 0-1 lesiklópálya-területek a Tiroli-Alpokban (1870 m), Ausztria
Hígított teny. m. +
fluor
Oesterreicher, 1988 15-640 (C) 0,5-20
340-27000 (Eu) 106-8483
Y|U|VSDOiQNpS] G|WWWDODMpYHVWHUPpV]e- tes vegetáció, USA
2,5-210 (C) 0,8-66 1600-25000 (Eu) 503-7854
Y|U|VSDOiQNpS] G|WWWDODMpYHVWHUPé- szetes vegetáció, USA
0-16 (C) <5,0 4,6-120 (Eu) 1,5-37
0-2
Y|U|VSDOiQNpS] G|WWWDODMORPEKXOODWyHUG
USA
MPN Hunt és mtsai, 1979
295,5-6063 46-952 0-1
53,7-826 76-1167 1-10
P YHOWP Wrágyázott szántóföld, búza – burgonya - árpa vetésforgóban, Csehország 1126,5-6959 177-1094 0-1
95,9-356,6 85-504 1-10
P YHOpVDOyO-3 éve kivont parlagterület, Csehország
265,4-1414,9 42-222 0-1
119-556,6 169-787 1-10 8-pYHQHPP YHOWNDV]iOy&VHKRUV]iJ
2,4-94,8 0,1-5,5 avarszint
0,3-6,3 0,05-1,0 0-1
0,1-1,9 0,1-2,7 1-10
40-pYHVW|OJ\HUG &VHKRUV]iJ
Hígított tenyészetek
módszere
Lukešová, 1993
6,1-793,3 4,8-623 lombhullató-pVW OHYHO HUG k talajából, Magyarország
1,44-135,12 1,1-106
15-20
lombhullató-pVW OHYHO HUG NWDODMiEyOe- Európa
MPN Fehér, 1936
1.3. táblázat folytatása
Sejtszám (103 sejt/g talaj)
Biomassza
NJpO W|PHJ
/ha)
Mélység
(cm) *\ MW KHO\
Sejtszámlá- lási módszer
Irodalom
? 6,2-41,2
(száraz súly) 0-1 gyertyános-tölgyes, folyó-ártéri területen, Szlovákia
klorofill-a
meghat. Hindák, 1975 8,4-106 1,4-17 0-1 N|YHVYi]WDODM~MUDWHOHStWHWWHUG WHUOHW
Csehország
0,3-2,2 0,2-1,7 0-5 N|YHVYi]WDODMHUG WHUOHW&VHKRUV]iJ
Hígított tenyészetek
módszere
Lukešová és Hoffmann, 1996
Magyarázat:
Zöld színnel a sejtszámból kalkulált - a vizsgált talajrétegre vonatkozó - biomassza-értékeket tüntettem föl.
A számításhoz az alábbi képletet használtam:
ahol,
N = 1 g talajban mért sejtszám
msejt = sejttömeg, átlagosan 10 PiWPpU M J|PEDODN~VHMWHWpVJFP3
VHMWV U séget feltételezve (5,24 × 10-10 g)
talaj iWODJRVWDODMV U VpJJFP3
) (Metting 1994) 105 = szorzó kg/ha-ra
h = vizsgált talajréteg vastagsága (cm) Jelölések:
C = Cyanophyta Ch = Chlorophyta D = kovaalgák
Eu = valamennyi eukarióta alga
Biomassza (kg/ha) = N × msejt × talaj × 105 × h
1.4.2. A talajalgák kvantitatív változásai
$]LURGDORPEDQV]HUHSO DOJDVHMWV]iP- és biomassza-értékek közötti jelen-
W VHOWpUpVHNWiEOi]DW– az alkalmazott módszerekNO|QE|] VpJpQW~O- az algák mennyiségének tér-pVLG EHOLYiOWR]iVDLYDOLVpUWHOPH]KHW N$WDODjalgák
NYDQWLWDWtYMHOOHJ YL]VJiODWDLW|EEQ\LUHHJ\V]HULPpUpVHNE ODGyGySLOOanatnyi
iOODSRWRNDWWNU|] WDQXOPiQ\RN)HKpU6KWLQD6KWLQDpV%olyshev 1963, Balezina 1975, King és Ward 1977, Tret'yakova és mtsai 1989 cit. Shtina 1991, Lukešová és Hoffmann 1996), és viszonylag kevés információ áll rendel-
NH]pVUH D] DOJDVHMWV]iP LG V]DNRV GLQDPLNiMiUyO +XQW pV PWVDL
Zimmerman és mtsai 1980, Lukešová 1993).
Gyakoribbak a faj-|VV]HWpWHOEHOLYiOWR]iVRNDWD]LG -, talajtípus-, vagy más
N|UQ\H]HWL WpQ\H] N IJJYpQ\pEHQ OHtUy PHJILJ\HOpVHN .LVV :LOOVRQ pV
Forest 1957, Komáromy 1975, 1982, 1983, 1984, 1985; Shubert és Starks 1985, Sukala és Davis 1994). A vizsgálatok eredménye alapján a fitoedafonban bekö-
YHWNH] YiOWR]iVRNDW W|EEIpOHNpSSHQ PDJ\DUi]]iN DPL pUWKHW LV KLV]HQ- Fe- hér Dániel (1954) szavaival élve – QHP HJ\ YDJ\ NpW WpQ\H] KDQHP QpKD D WpQ\H] NQHNV]LQWHEHOiWKDWDWODQpVWDOiQrészben nem is ismert sorozata játszik
LWWG|QW V]HUHSHW$]RQEDQYDODPHQQ\LHVHWEHQYLVV]DYH]HWKHW ND]|VV]HIg-
JpVHNDWDODMDOJiNHO ] HNEHQOHtUWDODSYHW |NROyJLDLLJpQ\HLUHOiVGDlfe- jezet). A talajok algaszám-YiOWR]iVDLW HO LGp] OHJIRQWRVDEE WpQ\H] NQHN D]
DOiEELDNHPOtWKHW N
Az alga sejtszámnak a talajmélységgel párhuzamos csökkenése, és a talajba
MXWyIpQ\PHQQ\LVpJpQHNKDVRQOyYiOWR]iVDHJ\pUWHOP HQDNDSFVRODWV]RURVVá- gát mutatja. Megfigyelések szerint populáció-maximum a talajfelszínen és a felszínközeli rétegekben (0-5 cm) mutatkozik (Bristol-Roach 1927, Alexakhina 1971, Drew és Anderson 1977, Hindák 1985, Lukešová 1993), de vannak 10-20
FP PpO\VpJEHQ LV MHOHQW V ELRPDVV]iW IHOWpWHOH] YpOHPpQ\HN )HKpU
King és Ward 1977).

![1.3. táblázat folytatása Sejtszám (10 3 sejt/g talaj) Biomassza NJpO W|PHJ /ha) Mélység (cm) *\ MW KHO\ Sejtszámlá-lási módszer Irodalom 0-0,18 (C) <0,001 P YHOHWOHQKXPXV]RV|QWpVWDODMRQ86$ 0,05-1600 (C) 0,0003-12 0-0,5](https://thumb-eu.123doks.com/thumbv2/9dokorg/866904.46665/33.1188.213.986.234.696/táblázat-folytatása-sejtszám-biomassza-mélység-sejtszámlá-yhohwohqkxpxv-qwpvwdodmrq.webp)

![2.3. ábra: $PHWHRUROyJLDLWpQ\H] NDODNXOiVDDYL]VJiOWLG WDUWDPEDQ](https://thumb-eu.123doks.com/thumbv2/9dokorg/866904.46665/62.918.240.702.322.602/ábra-phwhruroyjldlwpq-h-ndodnxoivddyl-vjiowlg-wduwdpedq.webp)
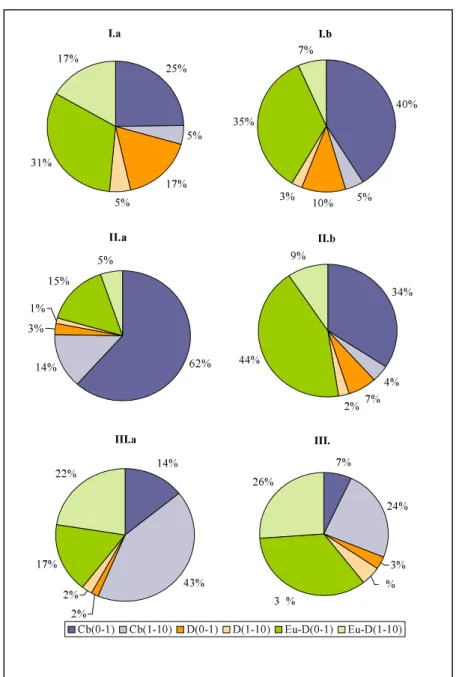
![2.3. táblázat $FVRSRUWRVtWyWpQ\H] NKDWiVDD]DO gacsoportok mennyiségére](https://thumb-eu.123doks.com/thumbv2/9dokorg/866904.46665/67.918.200.710.384.624/táblázat-fvrsruwrvtwywpq-h-nkdwivdd-do-gacsoportok-mennyiségére.webp)
