R. Meredith
C H A P T E R 7
I. I n t r o d u c t i o n 262 I I . Structure 262
1. M o l e c u l a r Structure 263 2. Crystalline and A m o r p h o u s R e g i o n s 266
3. Fine Structure 268 4. M o i s t u r e A b s o r p t i o n 269 I I I . C r e e p and R e l a x a t i o n 270
1. I n t r o d u c t i o n 270 2. C r e e p and C r e e p R e c o v e r y 272
a. Small Stresses 272 b. Larger Stresses 273 3. R e l a x a t i o n 276
a. C o n s t a n t Extension 276 b. C o n s t a n t T w i s t 279 4. Influence of H u m i d i t y and T e m p e r a t u r e 279
a. C r e e p 279 b. R e l a x a t i o n 282 I V . Stress-Strain R e l a t i o n s 283
1. L o a d - E x t e n s i o n Curves under Standard C o n d i t i o n s 283
a. Effect of R a t e of Extension 286 b. Influence of H u m i d i t y and T e m p e r a t u r e 286
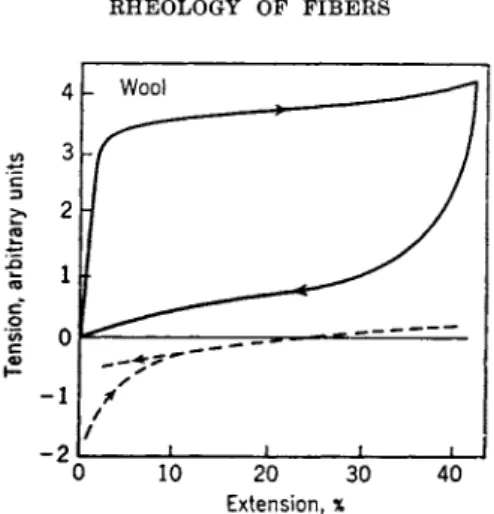
2. R e c o v e r y from Strain 289
a. Resilience 290 b. Crease Resistance 291 3. Flexural R i g i d i t y 292 V . D y n a m i c Properties 293
1. Elastic M o d u l u s and Internal F r i c t i o n 293 2. Influence of Static Strain and D y n a m i c Strain A m p l i t u d e 293
3. Effect of F r e q u e n c y and H u m i d i t y 295
4. Flexural R i g i d i t y 295 5. T o r s i o n a l R i g i d i t y 296 V I . Q u a n t i t a t i v e T h e o r e t i c a l Interpretation 298
1. T h e r m o d y n a m i c a l A p p r o a c h 298 2. R e a c t i o n - R a t e T h e o r y 300
a. T h r e e - E l e m e n t M o d e l 300 6. T w o - S t a t e T h e o r y 303 3. Distributions of R e l a x a t i o n T i m e s 305
V I I . Strength 307 1. Tensile Strength 307
261
2. Shear Strength
3. Resistance t o R e p e a t e d B e n d i n g N o m e n c l a t u r e
311 311 312 I. Introduction
Since some knowledge of fiber structure is essential t o an understanding of the rheological behavior of fibers, the first section describes the general features of fiber structure, with some detailed description of particular fibers where this is of special interest. T h e section on creep and relaxation emphasizes the important part played b y time. T h i s is followed b y a section on stress-strain relations with a controlled time factor. I n the section on d y n a m i c properties the response t o sinusoidally varying stress or strain has been recorded. A short section on quantitative theoretical interpreta- tion gives references for further reading and, finally, rupture properties are discussed.
T h i s survey is necessarily brief and touches only lightly on certain as- pects. A m u c h more complete a c c o u n t is t o b e found in a m o n o g r a p h b y the present author.1
Fibers m a y be divided into three main classes: (i) cellulose fibers, (ii) protein fibers, and (Hi) synthetic fibers. T h e natural cellulose fibers c o m - prise flax, c o t t o n , jute, h e m p , ramie, and other minor vegetable fibers such as k a p o k and coir fiber. Viscose rayon and c u p r a m m o n i u m rayon are man- made regenerated cellulose fibers, while cellulose acetate rayon is a cellulose ester fiber. W o o l , silk, and the m a n y varieties of hair fiber are natural protein fibers ; regenerated protein fibers that have been used commercially m a y b e made from the casein in milk ( A r a l a c ) , from the vegetable proteins in peanuts (Ardil), or zein ( V i c a r a ) . Synthetic fibers are m a d e b y the polymerization of m o n o m e r s t o form such materials as the polyamides, polyesters, p o l y v i n y l s , or polyethylene. Glass and asbestos can be p r o d u c e d in the form of filaments or fibers, b u t they will b e mentioned in this chapter only incidentally since their rheological properties d o n o t have the interest- ing characteristics possessed b y the organic fibers which consist of linear macromolecules.
I t is proposed t o discuss briefly those features of fiber structure which have a bearing on the interpretation of the rheological behavior of fibers.
Further information on the structure of fibers will b e found in b o o k s b y A s t b u r y ,l a M e y e r ,2 O t t ,3 Mauersberger,4 H e r m a n s ,5 R . D . Preston,6 J. M . Preston,7 H i l l ,8 Urquhart and H o w i t t ,9 and W o o d s .9 a
^ . M e r e d i t h , e d . , ' ' M e c h a n i c a l Properties of T e x t i l e F i b e r s . ' ' N o r t h - H o l l a n d Publishing, A m s t e r d a m , 1956.
1 o W . T . A s t b u r y , " F u n d a m e n t a l s of Fibre S t r u c t u r e . " Oxford U n i v . Press, N e w Y o r k , 1933.
II. Structure
1. MO L E C U L A R ST R U C T U R E
Fibers consist essentially of long chain molecules in which comparatively simple groups of a t o m s are joined together b y a condensation or addition polymerization reaction t o form a long chain of atoms j o i n e d b y primary valency linkages. R o t a t i o n a b o u t single b o n d s allows the chain molecule t o b e a v e r y strong b u t flexible unit, and this fundamental property is re- flected in the m a c r o s c o p i c rheological behavior of a fiber. Of course, the degree t o which the individual molecules can b e n d and coil is restricted b y the mutual interaction of active groups along the molecule, and from this point of v i e w the chemical constitution of the different fibers is important.
Close packing is hindered b y b u l k y side groups which m a y even form elec- trovalent or covalent primary linkages (as in w o o l ) , but in most cases the intermolecular forces are in the nature of secondary b o n d s such as h y d r o - gen b o n d s or v a n der W a a l s ' forces.
T h e fundamental building unit in cellulose fibers is the glucose residue linked as in cellobiose and expressed b y the formula
C H2O H H O H
I I I
c — ο c — c
/ \ H H
/ I | \
- O — ,
/ \ |
I / O H ΗΧ ι -
Ο C C
c
| \ O H H / \ — O — ' \ / I
H \ |
I X
\ / HC C C O
H O H C H2O H
A d j a c e n t glucose residues are identical except that each one is turned through 180 deg. a b o u t the chain axis with respect t o the next one. T h e chain molecules in cellulose are extremely long compared with their diame-
2 Κ . H . M e y e r , " N a t u r a l and S y n t h e t i c H i g h P o l y m e r s . " Interscience, N e w Y o r k , 1942.
3 E . O t t , e d . , " C e l l u l o s e and Cellulose D e r i v a t i v e s . " Interscience, N e w Y o r k , 1943.
4 H . R . Mauersberger, " M a t h e w s ' T e x t i l e F i b r e s . " W i l e y , N e w Y o r k , 1947.
5 P . H . Hermans, " P h y s i c s and C h e m i s t r y of Cellulose F i b r e s . " Elsevier, Amster- d a m , 1949.
6 R . D . Preston, " M o l e c u l a r Architecture of P l a n t Cell W a l l s . " C h a p m a n & H a l l , L o n d o n , 1952.
7 J. M . Preston, e d . , " F i b r e S c i e n c e . " T e x t i l e Institute, M a n c h e s t e r , 1953.
8 R . Hill, e d . , " F i b r e s from S y n t h e t i c P o l y m e r s . " Elsevier, A m s t e r d a m , 1953.
9 A . R . Urquhart, and F. O . H o w i t t , ed., " T h e Structure of T e x t i l e F i b r e s . " T e x - tile Institute, Manchester, 1953.
9 a H . J. W o o d s , " P h y s i c s of F i b r e s . " Institute of P h y s i c s , L o n d o n , 1955.
ter; a convenient w a y of expressing this length is b y using the term degree of polymerization ( D . P . ) which represents in this case the average number of glucose residues in the chain molecule. T h e D . P . of ordinary viscose r a y o n is a b o u t 350 and that of native cellulose fibers such as c o t t o n and ramie is at least 10,000. T h e ratio of the length t o the diameter of the chain molecules is thus v e r y high and the D . P . of undegraded native cellu- lose is m u c h higher than that of regenerated cellulose. T h i s has an influence o n the relative mechanical properties of native and regenerated cellulose, as will b e seen later.
W o o l and all mammalian hair fibers are c o m p o s e d mainly of the protein called keratin; silk filaments consist of the protein called fibroin. T h e s e proteins are polypeptides which have the basic structure
where the amino-acid residue, R, is different for successive units of struc- ture in the p o l y p e p t i d e chain. A s m a n y as 18 different amino acids have been identified from w o o l keratin and silk fibroin contains a b o u t 15 differ- ent amino acid residues. T h u s the protein chain molecules are far more complex than those of cellulose. A striking feature of the p o l y p e p t i d e chain is its richness in peptide b o n d s and the C O — and N H — groups m a y give rise, under suitable conditions, t o h y d r o g e n b o n d s between these groups in neighboring chain molecules. T h e proteins m a y therefore b e expected t o show intensive interchain secondary b o n d i n g b u t the possibilities are con- siderably restricted b y the side chains, consisting of amino-acid residues, which occur so frequently along the main chain and which must separate the main chains sufficiently t o allow for their a c c o m m o d a t i o n . T h e p o l y - peptide chains can interact b y means of their side chains t o form "salt linkages" (ionic in nature) or covalent linkages via the cystine groups.
These more stable covalent linkages confer on keratin a network structure and they are responsible for m a n y of its peculiar elastic properties. T h e regenerated protein fibers are made from " g l o b u l a r " proteins in which the p o l y p e p t i d e chains are folded in specific w a y s t o give relatively c o m p a c t molecules. Destruction of this specific configuration b y " d e n a t u r a t i o n "
allows the molecules t o b e drawn into alignment when spun into fibers.
T h e synthetic fibers h a v e m u c h simpler structures than a n y of the natural fibers and the ideas i n v o l v e d in explaining their rheological proper- ties are simpler. Polythene, i.e., polymerized ethylene, has the chemical formula ( C H2)n. I t is the simplest t y p e of linear macromolecule and b e - haves t o some extent like frozen rubber. Fibers of polythene m a y b e quite highly crystalline, caused b y the lack of side groups on the chain mole- cules, which allows close approach of the chain molecules over extended
Γ C O
\
C H I N H R
lengths: the molecules are held together side b y side b y v a n der W a a l s ' forces.
A number of synthetic fibers are based on vinylchloride ( C H2: C H C 1 ) polymerized with vinyl acetate ( C H3C O O C H : C H2) t o form V i n y o n ( 9 0 % vinyl chloride and 1 0 % v i n y l acetate) or with acrylonitrile ( C H2: C H C N ) , t o form the fibers k n o w n as V i n y o n Ν and D y n e l (60 % v i n y l chloride and 4 0 % acrylonitrile). T h i s c o p o l y m e r has a higher softening point than V i n y o n which it has superseded. If these fibers are heated, a considerable thermal contraction occurs and t h e y show long-range rubberlike elasticity from the shortened length. T h e X - r a y p h o t o g r a p h indicates p o o r crystal- linity. Acrylonitrile itself can b e polymerized and formed into fibers (e.g., Orion, Acrilan, and Creslan) which are p o o r l y crystalline. T h e long chain molecules interact b y virtue of the polar C N groups t o form h y d r o g e n b o n d s .
A n o t h e r fiber-forming material is vinylidene chloride (CH2:CCl2) which can b e c o p o l y m e r i z e d with v i n y l chloride t o form filaments, manufactured under the trade names Saran and V e l o n ; t h e y contain a b o u t 85 % vinylidene chloride and 1 5 % vinyl chloride. V i n y l alcohol ( C H2: C H ( O H ) ) forms the basis of y e t another synthetic fiber in the v i n y l family. W h e n polymerized it is highly crystalline and can be p r o d u c e d with a high degree of orientation.
Fibers such as V i n y l o n and K u r a l o n are based on polyvinylaic ohol which has been treated with formaldehyde t o form cross linkages between the chain molecules in the noncrystalline regions and so render the fiber in- soluble in water.
T h e best k n o w n synthetic fiber is nylon, a p o l y a m i d e formed b y the condensation polymerization of dicarboxylic acids and diamines. T h e c o m - monest t y p e is n y l o n 66, m a d e from adipic acid and hexamethylene diamine:
it has the formula [ — N H ( C H2)6N H — C O ( C H2)4C O — ] » . T h e outstanding characteristic of the n y l o n chain molecule is the occurrence at intervals along it of the peptide linkage — C O — N H - — , with the possibility of hy- drogen b o n d i n g between the C O and N H groups of neighboring molecules.
N y l o n is spun from the molten p o l y m e r and crystallizes on cooling even before it is cold-drawn b y some 300 or 400 % t o orientate the chain mole- cules and so strengthen the fiber. I n the noncrystalline regions the chain molecules have considerable freedom t o kink and fold owing t o the relatively large distance between the C O — N H groups in the main chains, which are the o n l y points where there is an appreciable degree of interaction between adjacent chains. T h e l o w elastic m o d u l u s and considerable elasticity are n o d o u b t dependent on this lack of interaction o v e r an appreciable fraction of the length of the chain molecule.
B y introducing b u l k y side chains at n o t t o o frequent intervals along the main chain, thus preventing their close approach and introducing a v e r y
p r o n o u n c e d t e n d e n c y for the chains t o fold, a highly extensible "elastic n y l o n " can be p r o d u c e d . F o r example, b y replacing the h y d r o g e n a t o m o n every other nitrogen a t o m in n y l o n 6 6 b y the group — ( C H2) 3— C H3, fibers with striking long-range reversible elasticity at ordinary tempera- tures are obtained. T h i s material has a crystalline c o m p o n e n t with m o d - erate orientation.
D a c r o n polyester fiber (Terylene in E n g l a n d ) is a relatively n e w syn- thetic fiber m a d e b y polymerizing ethylene glycol and terephthalic acid.
T h e chemical constitution of the chain molecule of p o l y e t h y l e n e tereph- thalate m a y b e written ( — 0 · C H2 · C H2 · O — C O — < ^ ^ > — C O — )w . T h e chain repeat distance of 1 0 . 7 5 A . is only slightly less than that required for a fully extended chain containing one chemical unit t o the geometrical repeating unit with successive ester groups in the trans configuration t o each other. T h e degree of crystallinity is a b o u t 5 5 t o 7 5 % and the length of the crystallites has been estimated t o be 6 0 t o 8 0 A .
2 . CR Y S T A L L I N E A N D AM O R P H O U S RE G I O N S
A t this stage it is appropriate t o consider the structure resulting from the interaction of the long chain molecules.
D u r i n g the formation of a fiber there is a t e n d e n c y for the chain mole- cules t o orientate themselves preferentially in the direction of the fiber axis, either as the result of the growth process in natural fibers or of the drawing which takes place during the spinning of regenerated or synthetic fibers. Consider, for example, a material spun into a filamentary f o r m : the molecules will b e arranged in a rather haphazard w a y , being entangled and crimped or folded in some of the m a n y possible configurations m a d e pos- sible b y their flexibility. T h e later stages of the spinning process in which the filaments are stretched tend t o align the molecules in the direction of the fiber axis. A s they are drawn into parallelism s o m e of the chain mole- cules will find themselves in positions favorable t o strong interaction with neighbors and will p r o d u c e crystalline regions or smaller bundles of chains if the regularly arranged material is not large enough t o be described as crystalline. I n other regions the chain molecules will n o t be so f a v o r a b l y placed and m a y remain in a partially orientated state. T h u s in the fiber there will b e a continuous gradation of degrees of orderliness of arrange- ment, ranging from crystalline areas t o patches with only traces of align- ment. I t is convenient t o describe the regions of perfect order as crystalline and those of c o m p l e t e disorder as a m o r p h o u s : all those regions which d o n o t show perfect order m a y be called noncrystalline.
F r o m the breadth of the interferences in X - r a y photographs of fibers, the dimensions of the crystalline regions or "crystallites" h a v e been esti-
mated. F o r example, in ramie the cellulose crystallite has been estimated t o have a length of at least 600 A . and a breadth of 50 t o 60 A . Filaments of viscose rayon give smaller values, a b o u t 300 A . in the direction of the fiber axis and a b o u t 40 A . across. I t is generally assumed that in fibers the aver- age length of the individual chain molecules is m u c h greater than that of the crystallites so that the c o n c e p t of a network structure arises. T h e junctions of the network are crystallites or chain bundles in the noncrystal- line regions and the links are chain molecules which m a y pass through several j u n c t i o n points.
W i t h these ideas of long chain molecules, crystalline regions, and network structure in mind w e m a y next consider the structure of particular fibers in a little more detail.
X - r a y examination shows that the alignment of the chain molecules with the fiber axis varies considerably in different fibers, at least in the crystal- line regions. Optical methods using polarized light, which give the average orientation of b o t h crystalline and noncrystalline parts of the fiber, show similar ranges of orientation. Flax, ramie, h e m p , and jute are highly orientated, c o t t o n shows an intermediate orientation while viscose rayon m a y have m e d i u m or high orientation depending on the a m o u n t of stretch applied during its formation.
Silk is perhaps the simplest protein fiber, consisting of fibroin in w h i c h nearly half the amino-acid residues are from glycine and one quarter from alanine. I t is believed that the crystalline parts consist mainly of these c o m p a c t amino acids. Other more complex amino acids such as serine and tyrosine occur with moderate frequency and fall in the noncrystalline regions. T h e r e is n o cystine t o form disulfide bridges between the p o l y - peptide chains so that, on the whole, the interaction between the chains is predominately of the weaker t y p e , with hydrogen bonding likely t o b e important.
W o o l and hair fibers consist of keratin, which contains a wide range of amino-acid residues of which none forms more than 16 % b y weight. A p p r o x - imately half of the total number of side chains are either basic, acidic, or cystine: side-chain interactions m a y thus be expected t o be far more im- portant in keratin than in fibroin. Keratin is n o t highly crystalline b u t such X - r a y reflections that occur indicate g o o d crystalline orientation in the direction of the fiber axis. In the unstretched form the X - r a y p h o t o - graph shows a repeat distance along the fiber axis of 5.1 A . c o m p a r e d with a b o u t 7 A . for the fully extended p o l y p e p t i d e chains in fibroin: the un- stretched keratin is said t o b e in the α-form and it is inferred that the chain molecules are folded in some w a y . T h i s is significant Theologically, for keratin fibers immersed in water show perfect long-range elasticity of form, e.g., at ordinary temperatures the range of elastic extension is as m u c h as
7 0 % . A s extension proceeds, the X - r a y pattern corresponding to the α- f o r m b e c o m e s progressively weaker and is replaced b y a different one, correspond- ing t o the jö-form, and similar t o that of fibroin. In other words, stretched keratin contains fully extended chain molecules which are produced from the regularly folded α-keratin. T h i s transformation is reversible and the perfect elasticity of keratin appears t o b e related t o this reversible trans- formation between folded and straightened chains. T h e nature of the α- f o l d is still under discussion.10
T h e r e are t w o points of view with regard t o the explanation of the long- range elasticity of keratin. O n the one hand, the fact that the crystallites h a v e specifically folded chains suggests that folding of this t y p e m a y b e expected t o occur in the noncrystalline regions also. T h e long-range elas- ticity of keratin might then b e explained in terms of the α-β transformation.
On the other hand, it m a y b e argued that the heterogeneity of the degree of order in the packing of the p o l y p e p t i d e chains in the fiber as a w h o l e sug- gests that the extensibility of the fiber has its origin in the noncrystalline parts and that the elasticity of keratin is of a rubberlike t y p e , g o v e r n e d b y e n t r o p y considerations.11 T h e rheological properties of keratin fibers shed some light on this subject and, on balance, appear t o favor the first p o i n t of v i e w .
3. FI N E ST R U C T U R E
T h e rheological properties of fibers will d e p e n d t o some extent on the w a y in which the fundamental building units of the fiber are assembled:
this is especially the case with natural fibers of biological origin such as c o t t o n , flax, and w o o l . F o r details reference m a y b e m a d e t o the b o o k s already m e n t i o n e d ; in w h a t follows, a brief sketch of s o m e of the outstand- ing structural features of the various fibers will b e given.
A n essential feature of all native cellulose fibers is the existence of fibrils which range in size from 50 A . diameter (and possibly less) up t o the order of a few microns. In highly swollen fibers some of these fibrils b e c o m e visible under the optical microscope and t h e y can be separated and examined un- der the electron microscope. A spiral structure is frequently observed which m a y v a r y from a left-hand t o a right-hand screw texture in successive layers. T h e angle between the direction of the fibrils in the cell wall and the fiber axis is small in flax, ramie, and h e m p fibers ( < 6 d e g ) b u t in c o t t o n the spiral angle near the surface is a b o u t 30 deg. and coir fiber has a spiral angle of 45 deg. C o t t o n fibers have a thin outer primary cell wall with a criss-cross fibrillar structure of a b o u t 70 deg. spiral angle, inside of w h i c h there is a secondary cell wall, which forms the major part of the fiber, with
1 0 A . E l l i o t t , Textile Research J. 22, 783 (1952).
1 1 Κ . H . M e y e r , A . J. A . v a n der W y k , W . G o n o n , and C . H a s e l b a c h , Trans. Fara- day Soc. 48, 6 6 9 (1952).
a 30-deg. spiral angle that alternates m a n y times along the length of the fiber from a left-hand t o a right-hand spiral. T h e s e reversals in the spiral structure are peculiar t o c o t t o n and occur a b o u t 20 times in a centimeter length. W h e n the fiber first dries out, the tubular wall collapses and the spiral structure causes the fiber t o c o n v o l u t e or twist. T h e average number of convolutions or 180 deg. twists ranges from 40 t o 80 per centimeter, de- pending on the variety and maturity of the c o t t o n , and these c o n v o l u t i o n s reverse their direction of rotation a b o u t the fiber axis at those places where the underlying spiral structure reverses.
T h e regenerated cellulose fibers are simpler t o deal with, for the "cellu- l o s e " from which t h e y are m a d e is purified during the processes of manu- facture and t h e y h a v e a simpler fine structure. H o w e v e r , m a n y c o m m e r - cially p r o d u c e d viscose rayons d o h a v e a skin and a core differing in molecular orientation and density w i t h a fairly sharply defined b o u n d a r y between t h e m : the orientation usually decreases as the core is approached.
W h e n r a y o n fibers are ground in a ball mill, splitting takes place in the direction of the fiber axis owing t o the anisotropic structure b u t the fi- brillae are observed t o b e not so well formed as with native cellulose fibers.
W o o l fibers are m a d e u p of several distinct parts. T h e main b o d y of the fiber is the cortex c o m p o s e d of long spindle-shaped cells; within this is a medulla, or central channel, c o m p o s e d of a network of cells enclosing air spaces. C o v e r i n g the cortex is the cuticle c o m p o s e d of a layer of scales rather like those of a fish and outside this is a v e r y thin continuous m e m b r a n e called the epicuticle, which is o n l y visible under the electron microscope.
T h e elastic properties are controlled b y the behavior of the cortex : recently this part of the fiber has been found t o consist of t w o different halves called the paracortex and orthocortex, which differ in their cystine content and on that a c c o u n t m a y b e expected t o h a v e different mechanical properties, for the behavior of w o o l keratin has been shown t o depend closely on the sulfur linkages between the chain molecules and the number of these linkages is proportional t o the cystine content.
R a w silk is a twin filament consisting of t w o filaments of fibroin e m b e d d e d in a matrix of silk g u m . T h i s silk g u m is mainly sericin and is r e m o v e d b y the chemical process of d e g u m m i n g t o leave isolated filaments of de- g u m m e d silk. Since these filaments are spun rapidly from solution b y the silk w o r m and are n o t the p r o d u c t of a slow growth process, they have a simpler fine structure than w o o l fibers in m u c h the same w a y that regen- erated cellulose fibers are simpler than native cellulose fibers.
4. MO I S T U R E AB S O R P T I O N
T h e a m o u n t of water held within a fiber can h a v e a considerable influence on the rheological behavior. T h e weight of absorbed water expressed as a fraction of the b o n e - d r y weight of the fibers is called the moisture regain: a
T A B L E I
M O I S T U R E R E G A I N OF V A R I O U S F I B E R S AT 2 5 ° C .
Moisture regain (%) at relative humidity of
Fiber 65% 95%
C o t t o n 7 . 0 1 5 . 0
Viscose 1 3 . 3 2 7 . 5
A c e t a t e 6 . 0 1 3 . 8
W o o l 1 4 . 7 2 7 . 7
Silk 1 0 . 0 2 3 . 8
N y l o n 4 . 2 7 . 7
D a c r o n 0 . 5 0 . 9
Orion 2 . 0 2 . 5
few examples of the moisture regains of fibers at 65 and 9 5 % relative h u m i d i t y are given in T a b l e I. I t will b e seen that w o o l , silk, and viscose r a y o n h a v e high regains, c o t t o n , acetate rayon, and n y l o n show interme- diate values whilst D a c r o n and Orion h a v e v e r y l o w regains.
A t any given relative humidity, a fiber has a greater capacity for holding water if brought into equilibrium with this h u m i d i t y from a higher relative h u m i d i t y than is the case when brought from a lower relative h u m i d i t y ; this fact must b e considered when conditioning fibers for rheological tests. T h e data in T a b l e I refer t o fibers which h a v e been conditioned from dryness t o the stated relative humidity, i.e., they have been brought along the absorp- tion isotherm as distinct from the desorption isotherm, which would be fol- lowed in conditioning them from the w e t state.
Since the mechanical properties depend on the moisture regain of the fiber, it is usual t o measure these properties in a standard atmosphere which has been defined as 65 ± 2 % r.h. at 20° C ; of course, this is n o t necessary for some of the synthetic fibers like Polythene, V i n y o n , or Saran which have negligible moisture regain at relative humidities normally encountered.
III. C r e e p a n d R e l a x a t i o n 1. IN T R O D U C T I O N
T h i s section is concerned with the simpler time-dependent p h e n o m e n a of creep, or slow deformation under constant stress, and relaxation, or d e c a y of stress at constant deformation.
W h e n a fiber is stretched under constant load, the extension is found t o increase rapidly at first and more slowly later: when the load is r e m o v e d , the fiber contracts, initially at a fast rate, w h i c h decreases almost t o zero after a sufficiently long time. In s o m e fibers, under the proper conditions, the r e c o v e r y after stretching proceeds until the initial length is attained
while in others, or the same fibers under different conditions, there remains a residual extension or 'set' even after prolonged times of retraction. A l - ternatively, if a fiber is rapidly stretched and held stretched, the tension developed decreases rapidly and m a y reach an equilibrium v a l u e : when the fiber is released and allowed t o recover freely, the tension on restretching t o the same extension as before will exceed the equilibrium value and m a y equal the tension originally developed on first stretching, showing that a
"tension r e c o v e r y " takes place rather similar t o the creep r e c o v e r y w h i c h occurs after a creep test.
Studies of creep in filaments of silk were m a d e as early as 1835 b y W e b e r and a detailed review of early w o r k on the subject of creep in filamentous materials is t o b e found in a b o o k b y L e a d e r m a n .12 Creep tests on fibers h a v e generally been m a d e under constant load rather than under constant stress, because of the greater experimental simplicity. A l t h o u g h the distinction between experiments m a d e under constant load and constant stress is hardly significant from the p o i n t of v i e w of theoretical interpretation when the strain is small, this is n o t generally true for the strains which are c o m m o n l y encountered with fibers and m o r e satisfactory data for theoretical inter- pretation are obtained from relaxation tests in which the geometrical form of the fiber remains constant during the experiment.
Stretching a fiber m a y i n v o l v e t w o main processes w h i c h m a y b e called b o n d stretching and chain straightening. T h e b o n d s with which w e are concerned are n o t the primary covalent linkages in the chain molecules themselves b u t rather those linking the molecules together t o form a net- w o r k structure. Before a b o n d can contribute effectively t o the extension of a fiber it must b e orientated in the direction of the fiber axis and the shorter
"chains of b o n d s " will orientate first. T h e breaking of one b o n d m a y allow the stress t o pass t o another one in parallel with it : thus a broken b o n d m a y remain in a condition where reformation is possible when the fiber is re- leased. Set will arise if the degree of b r e a k d o w n of the b o n d network is sufficiently drastic or if new b o n d s are formed which are sufficiently stable.
If they are not, the residual unbroken b o n d s in the network will tend t o pull the system b a c k toward the unstrained state b u t this will b e resisted b y the action of the new b o n d s . Creep r e c o v e r y arises from the gradual b r e a k d o w n of these b o n d s as t h e y b e c o m e stressed in resisting contraction. T h e rate of creep r e c o v e r y will therefore d e p e n d o n the degree of b o n d b r e a k d o w n , the number of new b o n d s , and the stability of these with respect t o the strength of the unbroken b o n d s . T h e breaking and building of b o n d s involves internal energy changes b u t these will b e almost inextricably mixed u p with c o n -
12 H . Leaderman, " E l a s t i c and Creep Properties of F i l a m e n t o u s Materials and other H i g h P o l y m e r s . " T e x t i l e F o u n d a t i o n , W a s h i n g t o n , 1943.
figurational changes caused b y chain straightening and these configuration changes will add an entropy term t o the elastic force within the fiber.
2. CR E E P A N D CR E E P RE C O V E R Y
a. Small Stresses
A detailed investigation of the creep and creep recovery of single fila- ments of viscose rayon, acetate rayon, silk, and n y l o n under small stresses has been made b y Leaderman.12 T h e specimens he used were "mechanically c o n d i t i o n e d " b y allowing the specimen t o creep for 24 hr. under load, then removing the load and allowing 24 hr. r e c o v e r y : in a n y subsequent 24-hr.
test the constant applied load did not exceed the conditioning load. B y this technique, the specimens were brought into an elastic state in which re- c o v e r y of strain was perfect so that the same filament could be used for m a n y experiments.
H e attempted t o see h o w far the superposition principle of B o l t z m a n n13 could b e used t o describe the observed creep behavior. T h i s principle states, in effect, that, at any instant, the deformation in a b o d y showing delayed elasticity does not result only from the load acting at that instant but also from the entire previous loading history. F o r the superposition principle t o apply, the magnitude of the delayed c o m p o n e n t of extension at a n y time must be proportional t o the load and the total extension caused b y a c o m - plex loading history must be equal t o the sum of the extensions produced b y the separate loading effects, removal of load being equivalent t o the ap- plication of a negative load.
Creep tests of 24-hr. duration and 24-hr. creep-recovery tests made b y Leaderman show b y their coincidence when suitably plotted that this prin- ciple is o b e y e d b y viscose rayon, acetate rayon, and silk u p t o " l o a d s " of 3 , 3 . 5 , and 6 X 108 d y n e s / c m .2, respectively. B e y o n d these loads the first assumption with respect t o proportionality of the delayed elastic extension and load breaks d o w n b u t the second assumption concerning the summation of separate loading effects holds, at least u p t o twice the loads quoted a b o v e . N y l o n o b e y s a modified form of the superposition principle for small strains u p t o about 2 % extension. All the a b o v e results are for tests made in the standard atmosphere of 6 5 dz 2 % r.h. and 20° C .
T h e validity of the principle m a y b e tested b y applying more c o m p l e x loading procedures. Suppose that a long-duration creep and recovery test is first carried out on a mechanically conditioned specimen. N e x t a sequence of short-duration tests is m a d e in which the load is applied for 10 min. fol- lowed b y 10 min. free recovery and this loading cycle is repeated. F r o m the superposition principle, if the extensions at times t, (10 + t), (20 + t),
1 3 L . B o l t z m a n n , Pogg. Ann. Phys. 7, 624 (1876).
T A B L E I I
C O N S T A N T L O A D OF 3 X 1 08 D Y N E S / C M2 FOR 20 HR. AT 6 5 % R . H .
Instantaneous Delayed extension extension Fiber Total extension, %
%
of total extensionViscose I 0.57 51 49
Viscose I I 0.30 71 29
A c e t a t e 0.76 70 30
Silk 0.34 69 31
N y l o n 2.16 40 60
N y l o n I I 1.26 50 50
(30 + t), etc., are a, 6, c, d, etc., then the extensions at these times in a continuous creep test without r e c o v e r y will be a, (a + 6 ) , (6 + c ) , (c + d), etc. G o o d agreement between calculated and observed extensions was found in experiments m a d e on viscose rayon.
S o m e typical data showing the relative amounts of instantaneous and delayed extension for mechanically conditioned specimens under a constant
" l o a d " of 3 X 1 08 d y n e s / c m .6 for 20 hr. at 6 5 % r.h. (from Leaderman's curves) are recorded in T a b l e I I . Under these conditions, ordinary viscose rayon I has a delayed extension equal t o a b o u t half the total extension, the same as for n y l o n I I . Stretching viscose rayon I in the swollen state t o p r o d u c e viscose I I has the effect of increasing the a m o u n t of instantaneous extension t o 7 0 % of the total extension. Its creep behavior is then similar t o that of silk and acetate rayon. T h e n y l o n I I sample is less extensible than n y l o n I and its percentage of instantaneous extension is higher.
b. Larger Stresses
A t higher stresses than those which h a v e just been considered, the in- stantaneous and delayed extensions are n o longer proportional t o the stress.
T y p i c a l behavior of an ordinary viscose rayon is shown in Fig. 1, which is based on data b y O'Shaughnessy14 for specimens which had received n o mechanical conditioning. T o emphasize the nature of the creep curves, the ratio of strain t o stress has been plotted against the logarithm of the time for a series of constant loads corresponding t o stresses from 2.6 t o 13.3 X 108 d y n e s / c m .2.
I t will b e seen that at l o w stresses the creep curve is convex t o the log time axis: at high stresses it is c o n c a v e t o the log time axis. A t intermediate stresses, the creep curve is sigmoid in shape. T h i s is a general description of the creep curves observed with fibers over a time range from 10 t o 105 sec.
1 4 M . T . O ' S h a u g h n e s s y , Textile Research J. 18, 263 (1948).
12
T h e curves suggest that each is part of a c o m m o n sigmoidal curve which m a y b e called the creep function. A t l o w stresses the relaxation mechanisms responsible for the high rate of creep at intermediate stresses h a v e not been invoked in the experimental time interval, at high stresses the relaxation p r o d u c e d b y these mechanisms has already taken place before observations have been started. It appears that increase of stress is equivalent t o a shift in the time scale.
R e c e n t l y the creep curves observed for nylon over a range of stresses have been generalized b y Catsiff, Alfrey, and O'Shaughnessy15 t o give a "master creep c u r v e , " which respresents the data as a single sigmoidal curve b y the application t o the individual curves of scale factors and shifts along the log time axis. A typical result of this process is shown in Fig. 2. T o reconstruct the creep curve at any particular stress one proceeds as follows: First the master curve is translated along the log time axis until its inflexion point falls at the characteristic retardation time for the given stress, next the ordinates of the curve are multiplied b y a scale factor, and finally the in- stantaneous extension for the given stress is added. T h u s , t o describe the creep behavior of a particular fiber one requires a master creep curve, and plots or tables of characteristic retardation time, scale factor, and instan- taneous extension for a range of stresses. D a t a for the creep and r e c o v e r y of n y l o n obtained b y A b b o t t1 6 show the general features indicated b y the master creep curve.
1 5 E . Catsiff, T . Alfrey, and M . T . O'Shaughnessy, Textile Research J. 23, 808 (1953).
1 6 N . J. A b b o t t , Textile Research J. 21, 227 (1951).
3
Time, seconds
F I G. 2. Master creep curve for nylon at 3 0 % r.h. and 3 6 ° C , showing its synthesis b y fitting together individual creep curves shifted vertically and laterally and mul- tiplied in ordinate. T h e numbers refer t o constant loads in units of 108 d y n e s / c m .2. (Catsiff et al.lb)
A n a t t e m p t t o analyze the creep curves of v i s c o s e17 and acetate rayon has been m a d e b y M a r k and Press.18 F o r the particular viscose-rayon creep curve chosen b y these authors, the following equation was deduced from the experimental d a t a :
e ( % ) = 0.6 + 1.9 + 1.0 (1 - e~tm) + 0.9 logio (t + 1)
D y n a m i c modulus measurements of v e r y short duration g a v e a value for the instantaneous extension of 0.6 % . B e t w e e n 0.01 and 1 sec. a v e r y slightly retarded elastic extension of 1.9% with retardation time less than 1 sec.
was postulated. B e y o n d t = 120 sec. the creep was mainly logarithmic and is represented b y the last term of the a b o v e equation. T h e remaining term on the right-hand side represents delayed creep with a retardation time of λ = 50 sec. T h i s term assumes that the delayed extension follows an ex- ponential law based on the rheological equation ρ = Ee + ηβ and that the retardation time λ = η/Ε.
T h e molecular interpretation of the terms in the a b o v e equation is that the first term represents the stretching of strong intermolecular b o n d s . I m - mediately afterward, the material responds with a fast elastic mechanism,
1 7 J. J. Press, / . Appl. Phys. 14, 224 (1943).
18 H . M a r k and J. J. Press, Rayon Textile Monthly 24, 297, 339, 405 (1943).
p r o b a b l y associated with the straightening of kinked chain molecules.
Following this comes a considerably retarded elasticity with a retardation time of 50 sec. which represents the displacement of larger units such as clusters of entangled chain molecules. Superimposed on these processes is a logarithmic creep which represents b o t h the molecular mechanisms with longer retardation times and a certain a m o u n t of crystallization.
W e g e n e r19 has represented his data on creep and creep r e c o v e r y of viscose, c u p r a m m o n i u m , and cellulose acetate rayon over a wide range of stresses at 55 % r.h. and 18° C . b y a power law represented b y log β = A ± Β log t over the time range from 30 t o 3000 sec. A formula of this t y p e is o b v i o u s l y applicable t o only a limited time range.
T h e creep of w o o l fibers under constant stress has been investigated b y S p e a k m a n ,20 w h o found that u p t o 150 min. the creep curves could b e de- scribed b y the formula I = l0 (1 + ßtV3), where the parameter β indicates the rate of creep. A m i n i m u m value of β was observed for a stress of 9 Χ 108 d y n e s / c m .2: for C o t s w o l d w o o l , β was 0.022, 0.008, and 0.013 at constant stresses of 6, 9 and 15 Χ 108 d y n e s / c m .2, respectively. T h e a b o v e creep formula has been found t o hold for soft metal wires when multiplied b y a term of the form ekt t o allow for the plastic flow which occurs in these metals.
R i p a and S p e a k m a n21 have studied the creep of w o o l fibers under a load of 7 Χ 108 d y n e s / c m .2 and found an exponential approach t o a limiting exten- sion (eiim.) of a b o u t 75%. T h e rate constant found from their linear p l o t of log (eiim. — e) against time varied widely from fiber t o fiber of Lincoln w o o l or of human hair. B y separating the fibers into groups according t o their creep rates they showed that the rate constants were inversely proportional t o the sulfur content of the fibers, which emphasizes the i m p o r t a n c e of the disulfide cross links in the rheology of keratin fibers. F r o m creep at different temperatures, they deduced a value of 26 k c a l . / m o l e for the activation energy, which is of the same order of magnitude as the 20 k c a l . / m o l e o b - tained b y the hydrolysis of cystine with dilute hydrochloric acid.
3. RE L A X A T I O N
a. Constant Extension
T h e p h e n o m e n o n of relaxation is closely related t o that of creep and methods of expressing creep behavior might b e expected t o be satisfactory for describing relaxation of stress. T h i s is perhaps best demonstrated b y taking the data obtained b y M e r e d i t h22 for viscose and acetate r a y o n at 65 % r.h. and 25° C . for a number of constant extensions and replotting them
1 9 W . Wegener, Zellwolle, Kunstseide, Seide 46, 298, 342 (1941).
2 0 J. Β . Speakman, J. Textile Inst. 17, T472 (1926).
2 1 O . R i p a and J. B . Speakman, Textile Research J. 21, 215 (1951).
2 2 R . Meredith, J. Textile Inst. 45, T438 (1954).
1 1 02 1 04 1 06 1 08 1 01 0 1 01 2 1 014 1 016
Time, seconds
F I G . 3 . C o m p o s i t e curves for relaxation of tension at 6 5 % r.h. and 2 5 ° C . under constant extensions shown in the k e y . C o n t i n u o u s curve and left-hand scale refer t o acetate r a y o n : dashed c u r v e and right-hand scale refer t o viscose r a y o n .
as stress divided b y strain against the logarithm of the time. W h e n this was d o n e it was observed that the slopes of the curves were the same for different constant extensions at any given value of stress over strain, i.e., the family of curves obtained at different constant extensions appeared t o b e part of a c o m m o n curve which shifted t o lower values of l o g t as the extension was increased. A c o m p o s i t e or master relaxation curve was therefore m a d e b y translating each individual curve along the log time axis until a fit of the overlapping ends was obtained. T h e result is shown in Fig. 3 for viscose r a y o n and cellulose acetate r a y o n . S i g m o i d curves are obtained w h i c h are rather similar t o the master creep curve of Fig. 2 . T h e c o m p o s i t e creep curve gives the correct relation between stress o v e r strain and log time for a constant extension of 1%: t o obtain the curves corresponding t o higher constant extensions, a relation between extension and the a m o u n t of shift ( Δ log t) is required. F o r acetate r a y o n the relation was found t o b e approxi- mately Δ logio t — 12 logio e and for viscose rayon
Δ logio t ^ 26 >J/logio (1 + e),
where e is expressed as a percentage and t is in seconds. I t is clear from F i g 3 that at l o w and high extensions the relative rate of d e c a y of stress is small, whereas at intermediate extensions of 5 t o 10 % the rate of stress d e c a y is appreciably higher.
W e g e n e r23 has m a d e a detailed study of the d e c a y of tension with time
2 3 W . Wegener, Melliand Textilber. 30, 9 0 , 1 3 8 , 1 8 4 , 2 2 9 , 2 8 2 , 3 8 8 , 4 4 3 , 5 0 1 , 5 5 8 ( 1 9 4 9 ) .
for viscose rayon, c u p r a m m o n i u m rayon, and cellulose acetate rayon when extended b y various amounts at 6 5 % r.h. and 20° C . T h e tension d e c a y was observed over a rather restricted time range from 1 sec. t o 5 min. b u t his results agree with those already given in Fig. 3 as far as the shape of the relaxation curve and the rate of d e c a y of stress are concerned. H e repre- sented his data b y the formula p/p0 = [(t + a)/a]~n where p0 is the stress at time t = 0 and a and η are constants. T h i s formula reduces t o the f o r m log e = A + Β log ρ when t ^> a, as is the case for t > 10 sec. A p o w e r law of this latter t y p e can also b e used t o describe the tension d e c a y in c o t t o n and flax y a r n s22 over a time range of 3 4ο t o 105 sec.
T h e relaxation behavior of w o o l and hair fibers is rather m o r e compli- cated than that of the cellulosic fibers in that there is a possibility, under suitable conditions, of the b r e a k d o w n of the disulfide cross linkages be- tween adjacent chain molecules. T h e relaxation of tension in w o o l fibers23a'24 extended b y different amounts in water at 25° C . is shown in Fig. 4. T h e r e is a rapid initial d e c a y of stress within the first second, followed b y a slower d e c a y , and later b y a further slow d e c a y which accounts for m o s t of the tension d e c a y at the higher extensions. T h e influence of p H throws some light on this slow relaxation p r o c e s s .25 I n acid the slow relaxation is in- hibited or deferred whereas in alkali it is hastened. T h e b o n d s whose break- d o w n leads t o the slow relaxation are thus stable at l o w p H and unstable at high p H , which is consistent with the idea that these b o n d s m a y b e
2 30 B . J. R i g b y , Australian J. Sei. Research A8, 176 (1955).
2 4 G . C . W o o d , Textile Inst. 45, T462 (1954).
2 5 J. B . Speakman, and S. Y . Shah, J. Soc. Dyers Colourists 57, 108 (1941).
disulfide linkages between the p o l y p e p t i d e chains, their hydrolysis being catalyzed b y h y d r o x y l ions. S o m e further e v i d e n c e favoring this hypothesis is p r o v i d e d b y the experiments of Reese and E y r i n g26 w h o studied the in- fluence of reducing agents on this slow relaxation.
Lasater, N i m e r , and E y r i n g27 are using the change in the rate of relaxa- tion of w e t c o t t o n fibers t o study the reaction of c o t t o n t o acids and alkalis.
b. Constant Twist
S o m e measurements of the d e c a y of torque in twisted bundles of fibers and yarns m a d e from c o t t o n were m a d e b y P e i r c e28 w h o found that the re- sults could be described empirically over a time range from several seconds t o a d a y b y the equation
Τ = c + a e x p . ( - #1 / 3)
where Τ is the torque, c is the elastic couple which persists for the duration of the experiment and a is a couple which d e c a y s with time at a rate c o n - trolled b y β. T h e same law was found t o hold for m a n y materials, e.g., silk, w o o l , tin foil, aluminium foil, and glass. P e i r c e29 also expressed this torsional relaxation in terms of a spectrum of relaxation times which h e translated into activation energies and with which he attempted t o explain the values so found in physical terms (see Section V I - 3 ) .
T h e relaxation of torque in a twisted n y l o n monofilament was found b y H a m m e r l e and M o n t g o m m e r y30 t o b e approximately linear with log time from 2 0 t o 2 0 , 0 0 0 sec. and this was interpreted in terms of a distribution of relaxation times such that the contribution t o the torque from a mechanism with a given relaxation time is inversely proportional t o the magnitude of the relaxation time (see Section V I - 3 ) .
4. TH E IN F L U E N C E O F HU M I D I T Y A N D TE M P E R A T U R E
One w o u l d expect the rates of creep and of relaxation t o increase with increasing h u m i d i t y or temperature, and this is found t o b e the case.
a. Creep
Steinberger31 m a d e a series of measurements on the creep of single fila- ments of cellulose acetate rayon and c u p r a m m o n i u m r a y o n covering a range of relative humidities from 0 t o 1 0 0 % at a constant load equivalent t o a nominal stress of a b o u t 6 . 5 Χ 1 08 d y n e s / c m .2 T h e effect of increasing the relative h u m i d i t y in steps of a b o u t 1 0 % can be seen for cellulose acetate
2 6 C . E . Reese and H . Eyring, Textile Research J. 20, 743 (1950).
2 7 J. A . Lasater, E . L . Nimer, and H . Eyring, Textile Research J. 23, 237, 481 (1953).
2 8 F. T . Peirce, J. Textile Inst. 14, T390 (1923).
29 ρ f . Peirce, in "Essays in R h e o l o g y , " p . 47. Pitman, L o n d o n , 1946.
3 0 W . G . Hammerle and D . J. M o n t g o m e r y , Textile Research J. 23, 595 (1953).
3 1 R . L . Steinberger, Textile Research 6, 191, 267 (1936).
% r.h.
01 ι ι ι 1 1 1 Ι Ο2 Ι Ο3 Ι Ο4 1 0 5 1 0 6
T i m e , s e c o n d s
F I G . 5 . Influence of h u m i d i t y on the creep of acetate rayon at 6 . 5 Χ 1 08 d } ^ n e s / c m .2 and 2 8 ° C . (Steinberger.*1)
rayon from Fig. 5. F r o m 0 t o 4 0 % r.h. the a m o u n t of creep is small, but a b o v e this relative humidity it rises rapidly. It will b e noticed that each curve appears t o b e part of a c o m m o n sigmoidal curve that is shifted along the log time axis t o shorter times as the relative humidity is raised, thus resembling the family of creep curves obtained for viscose rayon at constant relative humidity and various loads. N y l o n1 2 under a constant " l o a d " of 1.8 Χ 108 d y n e s / c m .2 gives a similar family of creep curves for relative humidities of 0, 62, and 1 0 0 % r.h. T h e creep of viscose and acetate rayon under constant loads from 1.2 t o 9.4 Χ 108 d y n e s / c m .2 at 30 t o 6 5 % r.h.
has also been studied b y Pinte and H e n n o .32 B y wetting and reconditioning fibers t o r e m o v e internal strain set u p b y creep, it is possible t o show h o w m u c h of the strain p r o d u c e d b y creep is elastic and h o w m u c h represents permanent change in the fiber. Press17 found that u p t o 12 % extension, the recovery of ordinary viscose rayon could b e divided roughly into three ranges. Extensions up t o % % are retarded and c o m p l e t e l y elastic, repre- senting the m a x i m u m m o v e m e n t of molecular segments before weaker sec- o n d a r y b o n d s in the noncrystalline regions begin t o break. F r o m % t o 4 % , extension is not c o m p l e t e l y recoverable until the sample is swollen in water : swelling reduces the steric hindrance and allows the weakened restoring forces t o bring chain segments b a c k t o their original state of order. A b o v e 4 % of the extensions are not completely recoverable because of molecular segments having been m o v e d t o new equilibrium positions.
3 2 C . Pinte and M . H e n n o , Bull. inst. textile France 6, 3 5 ( 1 9 4 8 ) .
T h e effect of temperature on the a m o u n t of creep of dry viscose rayon m a y b e quite large. T h u s , although the general form of the creep curve re- mained unchanged, Smith and Eisenschitz33 found that the a m o u n t of creep at a n y time was seven times as great at 114° C . c o m p a r e d with that at 20° C : lowering the temperature t o —180° C . reduced the a m o u n t of creep to one-sixth of that at 20° C . If all the relaxation times of the mechanisms responsible for creep are similarly affected b y a change of temperature, the shape of the creep curve should b e unaltered for moderate temperature changes, i.e., a change of temperature should merely displace the creep curves on the logarithmic time plot along the time axis. T h i s kind of effect was found for cellulose acetate r a y o n and n y l o n b y L e a d e r m a n .12 Under a stress of 3.7 X 108 d y n e s / c m .2 in the temperature range from 51 t o 75° C , a decrease in temperature of 10° C . corresponded t o a multiplication of the time scale b y nine for dry cellulose acetate r a y o n : for nylon, a decrease in temperature of 10° C . corresponded t o a multiplication of the time scale b y six. T h e rate of creep of w o o l has been shown b y S p e a k m a n20 t o increase
A v i t h increasing temperature.
Keratin fibers show perfect elasticity of form when stretched quickly b y 3 0 % or less in water at r o o m temperature. If the time under strain, or the temperature or the strain is increased, the recovery is incomplete and the material acquires a " s e t . " T o b e able t o retain a set in fibers has important technological advantages and the conditions required t o p r o d u c e a set h a v e been m u c h studied.34 T w o possible effects in the b o n d network can lead t o set, namely, irreversible b o n d b r e a k d o w n and b o n d rebuilding. T h e set m a y be recovered if the conditions under which it was p r o d u c e d are altered and it is necessary t o distinguish between " c o h e s i v e " set (produced b y dehydra- tion with the formation of h y d r o g e n b o n d s which are easily broken when the fiber is placed in w a t e r ) , " t e m p o r a r y " set (which requires h o t water or steam for its removal and is associated with the formation of m o r e stable b o n d s than the h y d r o g e n b o n d ) , and " p e r m a n e n t " set (which resists the action of prolonged steaming). A l t h o u g h keratin fibers can be permanently set b y relaxation at high temperatures, this can only occur in the presence of water since strained fibers heated at 100° C . in dry air d o n o t acquire even temporary set. Steam is quite effective b u t aqueous solutions which are alkaline or which contain a setting agent such as sodium bisulfite are more effective. W h e n treated fibers are placed in boiling water, for example, there is a slow creep recovery and the a m o u n t of truly permanent set is only a fraction of the setting extension. I t is believed that the disulfide b o n d s play a primary part in the setting of keratin. F o r example, the basis
3 3 H . de W i t t Smith and R . Eisenschitz, , / . Textile Inst. 22, T170 (1931).
3 4 L . Peters and H . J. W o o d s , in " M e c h a n i c a l Properties of T e x t i l e F i b r e s "
( R . M e r e d i t h , e d . ) , C h a p t e r 11, N o r t h - H o l l a n d Publishing, A m s t e r d a m , 1956.
of cold permanent-waving systems is the reduction of the disulfide b o n d followed b y its reoxidation in the strained state.
Before leaving this subject of set, the p h e n o m e n o n of supercontraction in keratin fibers should b e mentioned. Steaming stretched fibers for 2 min. at high tensions is followed b y a contraction in steam or boiling water which, after a long time, reduces the length of the fiber t o as m u c h as 30 % less than its initial unstrained length. T h i s treatment hydrolyzes some of the disulfide b o n d s but does not allow sufficient time for appreciable b o n d re- building t o take place.
b. Relaxation
T h e d e c a y of tension in viscose rayon over a period of 5 min. has been studied b y W e g e n e r and L u y k e n35 at relative humidities of 45, 65, 85, and 1 0 0 % r.h. with constant extensions from 2 t o 1 8 % reached at a constant rate of extension of 0 . 6 6 % per sec. T h e curves are of the form given b y P/Po = [(t + a)/a]~n, where a and η are disposable constants. T h e rate of relaxation is higher at 65 t o 8 5 % r.h. than it is at 45 and 1 0 0 % r.h. I t is possible that the apparently l o w rate of relaxation at 100 % r.h. is a result of the relatively slow extension up t o the desired constant extension which allows the mechanisms with short relaxation times t o relax while the speci- men is being extended.
S p e a k m a n36 measured the d e c a y of tension in w o o l fibers stretched about 3 0 % at 25, 50, 75, and 1 0 0 % r.h. T h e d e c a y of stress remained linear with log time up t o 75 % r.h. b u t the slope of the relaxation curve increased at the longer times at 1 0 0 % r.h., giving a curve of the same shape as that for 3 0 % extension shown in Fig. 4. I t appears then that the relaxation of disulfide b o n d s is inhibited at relative humidities of 7 5 % r.h. and less.
A s the temperature is increased,24 the relaxation curve of w e t w o o l changes in shape and from 65 t o 95° C . it b e c o m e s c o n v e x t o the log time axis over the whole of its observed course (from 30 sec. t o 100 m i n . ) : the rate of relaxation is higher than at r o o m temperature and is characteristic of a single rate process : this is p r o b a b l y so because of the first rapid relaxation taking place before observations could be m a d e .
A t r o o m temperature the relaxation curve of n y l o n at 1 5 % extension shows an almost linear relation between stress and log time at all humidities.
H o w e v e r , the relaxation curve changes considerably with increase in tem- perature37 as shown in Fig. 6, which refers t o n y l o n in water. T h e shape of the curves for temperatures greater than 53° C . is rather similar t o that for w o o l2 4 in water or in alkali at r o o m temperature.
3 5 W . Wegener and E . L u y k e n , Melliand Textilber. 33, 37 (1952).
3 6 J. B . Speakman, Proc. Roy. Soc. B103, 377 (1928).
3 7 J. B . Speakman and A. K. Saville, / . Textile Inst. 37. Ρ271 (1946).
Ol ι ι ι I
1 0 1 02 1 03 1 04 1 05
T i m e , s e c o n d s
F I G . 6 . Influence of temperature on the relaxation of nylon in water at 1 5 % e x - tension. (Speakman and Saville.®)
I V . S t r e s s - s t r a i n Relations
1. LO A D -EX T E N S I O N CU R V E S UN D E R ST A N D A R D CO N D I T I O N S
T h e stress-strain curve of a fiber is a g o o d illustration of the resistance t o stretching or twisting; it shows graphically the influence of humidity and temperature on the elastic properties and enables us t o assess the effect of physical and chemical treatments. Since the stress corresponding t o a given strain depends quite appreciably on the time factor, it is necessary t o control either the rate of extension or the rate of loading. T o simplify quantitative theoretical interpretation, it is desirable t o use a constant rate of extension but for practical purposes a constant rate of loading m a y b e easier t o achieve and equally satisfactory. T h e load-extension curves obtained in a standard atmosphere of 6 5 % r.h. and 20° C . will b e considered first, and later the influence of the rate of extension, humidity, and temperature will b e con- sidered.
T y p i c a l stress-strain curves for various cellulose fibers are shown in Fig. 7.
T h e high Y o u n g ' s modulus and high strength of flax is characteristic of the bast fibers in general, all of which h a v e a high degree of crystallinity and a highly orientated molecular structure. T h e t w o stress-strain curves for c o t - t o n illustrate t w o p o i n t s : firstly, that different varieties of c o t t o n h a v e curves of typical shape b u t different elastic moduli and, secondly, that the
Ο 1 0 2 0 Extension, %
F I G . 7. Stress-strain curves of cellulose fibers at 6 5 % r.h. and 20° C . K e y : F = flax; CS = Sea Island c o t t o n ; CT = T e x a s c o t t o n ; A = acetate r a y o n ; VHT = high- tenacity viscose r a y o n ; V MT — m e d i u m - t e n a c i t y viscose r a y o n ; V0 = ordinary vis- cose r a y o n .
extensibility of c o t t o n is appreciably greater than that of the bast fibers, because of the spiralling of the fibrils about the fiber axis. Ordinary viscose rayon has an initial Y o u n g ' s modulus similar t o that of c o t t o n but the fiber yields at an extension of about 2 % and b e y o n d this point it shows a m u c h greater extension per unit increase in stress and reaches a higher extension at rupture than any of the natural cellulose fibers. T h e three curves for viscose rayon illustrate a general feature of man-made fibers, namely, that increasing the molecular orientation b y stretching the fibers during manu- facture increases the modulus of elasticity and the strength, and lowers the breaking extension. T h i s general principle is followed b y synthetic fibers such as nylon, D a c r o n , O r i o n ,38 etc. T h e stress-strain curves for these ma- terials shown in Fig. 8 correspond t o the 'regular' types in current p r o d u c - tion for textile purposes; higher resistance t o extension with higher strength and correspondingly lower breaking extension can easily b e obtained for special applications such as use in tire cords. One striking feature of n y l o n is its l o w initial modulus compared with D a c r o n and Orion (continuous filament). T h e w o o l fiber has a characteristic stress-strain curve consisting
3 8 G . H . Fremon, in "Fibres from Synthetic P o l y m e r s " ( R . Hill, e d . ) , p . 461.
Elsevier, A m s t e r d a m , 1953.
0 10 20 30 Extension, %
F I G. 8. Stress-strain curves of protein and synthetic fibers at 6 5 % r.h. and 20° C . K e y : Τ = T e r y l e n e ; Ν = nylon 66; S = silk; W = w o o l ; Oc = Orion (continuous filament); 04i, 04 2 = Orion (staple fiber); V = V i n y o n N .
of three regions: up t o 2 % extension the resistance t o extension is fairly high and is caused b y straining the b o n d n e t w o r k ; next the h y d r o g e n b o n d s yield and the p o l y p e p t i d e chains in the noncrystalline regions start t o un- fold. A t extensions of 20 t o 30 % the resistance t o extension increases and at this stage the transformation of the crystalline phase from the a- t o the ß-form begins, as shown b y X-rays. Silk has a much higher initial modulus than wool, as m a y be expected since the polypeptide chain molecules are already in the fully extended ß-form in the unstressed state of the fiber.
For further information, reference can be made to Meredith,39 Hermans,0
Speakman,40 Susich and Backer,41 F a r r o w4 1 a, Lucas et. a/.4 1b
T h e manner in which the torque in a filament develops as twist is inserted has been studied b y M o r t o n and Permanyer,42 using viscose rayon, acetate rayon, and nylon. T h e shapes of the torque-twist curves are similar t o the corresponding tensile load-extension curves. There is an initial linear part up to twists of about 3 turns per cm., followed b y a yielding of the fiber so
3 9 R . Meredith, J. Textile Inst. 36, T107 (1945).
4 0 J. B . Speakman, J. Textile Inst. 18, T431 (1927).
4 1 G . Susich and S. Backer, Textile Research J. 21, 482 (1951).
4 1α B . Farrow, J. Textile Inst. 47, T58 (1956).
4 16 F . Lucas, J. T . B . Shaw, and S. G . Smith, / . Textile Inst. 46, T440 (1955).
4 2 W . E . M o r t o n and F . Permanyer, J. Textile Inst. 40, T371 (1949).