Global and Planetary Change 205 (2021) 103609
Available online 5 August 2021
0921-8181/© 2021 The Author. Published by Elsevier B.V. This is an open access article under the CC BY license (http://creativecommons.org/licenses/by/4.0/).
Research Article
Chronostratigraphy of clinothem-filled non-marine basins: Dating the Pannonian Stage
Imre Magyar
a,b,*aMTA-MTM-ELTE Research Group for Paleontology, Budapest, Hungary
bMOL Hungarian Oil and Gas Plc., Budapest, Hungary
A R T I C L E I N F O Editor: Zhengtang Guo Keywords:
Clinoform Clinothem Chronostratigraphy Biostratigraphy Magnetostratigraphy Neogene
Pannonian basin Pannonian Stage
A B S T R A C T
Clinoforms and clinothems are important and common architectural elements in the sedimentary record of many basins worldwide. The influence that they exert on chronostratigraphic interpretations, however, is often underestimated. This is especially true in non-marine basins, such as the Central European to Central Asian Neogene Paratethyan basins, where chronostratigraphic division and correlation is usually based on fossils of environmentally sensitive molluscs recovered from outcrops of shallow-water deposits. In the subsurface, the diachronous boundary between the foreset and topset parts of the shelf-edge scale clinoforms, where significant lithological and paleontological changes can be observed with the transition from deep to shallow water, was routinely identified with one of the regional stage or substage boundaries, inflicting serious confusion in chro- nostratigraphy. In the late Neogene brackish lacustrine basin fill of the Pannonian basin (Pannonian Stage, 11.6 to 2.6 Ma), where extended endemic radiation and long-term gradual morphological change in various groups of the biota offer good marker species for high-resolution biostratigraphy, molluscs and dinoflagellate algae are used to establish a clinothem chronostratigraphy. The biozone and clinothem boundaries are correlated with the geological time scale through mammal stratigraphic considerations, magnetostratigraphy, and radiometric dating. For the time being, the beginning of lacustrine deposition at 11.6 Ma and the intervals between 8.8 and 7.1 Ma and 4.6–2.6 Ma are accurately dated in drill cores. These time intervals can be correlated through the dense seismic network across the basin. For other time intervals of the late Miocene and Pliocene, the uncertainty of dating remains high on the order of several 100 kys.
1. Introduction
The study of clinoform depositional surfaces and clinothem forma- tion has recently become an individual discipline (e.g. Henriksen et al., 2009; Patruno and Helland-Hansen, 2018; Pellegrini et al., 2020). In- terest in clinothem formation is mostly focused on the sedimentological processes, their forcing agents and variability through time, and quan- titative assessment and modelling of the connection between sediment delivery/deposition and clinoform geometry. Much attention was also paid to the identification and dating of chronostratigraphic surfaces in clinothems at various scales (e.g. Gani and Bhattacharya, 2005; Lease and Houseknecht, 2017). With the advent of computational seismic chronostratigraphy, the determination of chronostratigraphic bound- aries in seismic datasets is no longer considered a challenge. In spite of all these achievements, however, the use of traditional local and regional chronostratigraphic units, which had been established prior to
the recognition of clinothems in the given succession, remains a source of much confusion and miscorrelation in many basins worldwide. Shelf- edge scale clinoforms are usually difficult to recognize in the field (e.g.
Olariu et al., 2020), and the definition of chronostratigraphic units was often based on lithological or paleontological changes which later turned out to be facies change within a clinothem. In particular, the transition between the usually fine-grained foreset and generally coarser-grained topset parts of the shelf-edge scale clinothems is often interpreted mistakenly as a sign of basinwide base-level drop, a phe- nomenon generally used to mark chronostratigraphic (or sequence stratigraphic) boundaries. Shelf-edge scale clinothem deposition im- plies, however, that similar or identical changes in depositional envi- ronment, and thus in lithology and fossils, occur at considerably different times in different points of the same basin. An oversimplified lithological or biostratigraphical approach can easily fail to grasp the chronostratigraphic markers that commonly cross lithostratigraphic
* Corresponding author at: MTA-MTM-ELTE Research Group for Paleontology, Budapest, Hungary.
E-mail address: immagyar@mol.hu.
Contents lists available at ScienceDirect
Global and Planetary Change
journal homepage: www.elsevier.com/locate/gloplacha
https://doi.org/10.1016/j.gloplacha.2021.103609
Received 15 May 2020; Received in revised form 31 July 2021; Accepted 3 August 2021
(facies) boundaries. Lateral sediment accretion, where time elapses more laterally than vertically, partly violates Steno’s rule of super- position (Gani, 2017), and it is perceived as counterintuitive by many geologists. The influence that lateral sediment accretion exerts on chronostratigraphic interpretations is thus clearly underestimated in many cases. This is especially true for the restricted marine to lacustrine to fluvial Neogene Paratethys basins, where the boundary between the foreset and topset parts of the shelf-edge clinothems were routinely identified with various regional stage or substage boundaries originally defined by shallow-water dwelling molluscs (e.g. T˘ar˘apoanc˘a, 2004;
Tilita et al., 2007; Jipa and Olariu, 2009; Leever et al., 2009; Novak Zelenika and Malvic, 2011; Mat¸enco and Radivojevi´c, 2012; Casas- Gallego et al., 2020).
The recognition of clino-stratified successions is essential for the realistic reconstruction of the paleogeographic and sedimentary evolu- tion of depositional basins in both space and time. This case study demonstrates how the traditional biostratigraphy, based on shallow- water dwelling endemic molluscs, is reconciled with seismic stratig- raphy in the lacustrine, clinothem-bearing late Neogene Pannonian Stage, Pannonian basin, Central Europe. (In this paper the term “Pan- nonian Stage” is applied for the entire upper Neogene (upper Miocene and Pliocene) basin fill of the Pannonian basin, as indicated by Hilgen et al., 2012). The objective of this study is threefold: 1) to display the spatial position of Pannonian chronostratigraphic units (in this case, biochronozones) across the entire clinothem-filled Pannonian basin in schematic figures, using data from various countries of the region; 2) to unfold how the clinothem-bearing non-marine basin fill is calibrated to
the geological time scale; 3) finally, to demonstrate the need for similar integration of surface and subsurface data in order to establish a consistent chronostratigraphy in other clinothem-filled non-marine Paratethyan basins.
2. Stratal geometry of the upper Neogene in the Pannonian basin
The Pannonian basin (Fig. 1) is a Neogene back-arc basin formed in the collision zone of Europe and Africa (Horv´ath et al., 2006). The early to late Miocene rifting in various sub-basins was followed by a general thermal subsidence in the late Miocene. Coeval geographical separation from the Paratethys Sea, which once stretched from Central Europe to Central Asia (Popov et al., 2004), led to the formation of a giant, deep, brackish-water lake in the basin, similar to the present-day Caspian Sea.
The area of Lake Pannon at its largest extent was ca. 250,000 km2, and its depth was several hundred metre to more than one thousand metre in the various sub-basins (Bal´azs et al., 2018). The Pannonian basin, as a mature hydrocarbon exploration area, has an extensive coverage of 2D seismic lines and 3D seismic cubes (Kov´acs, 2018). These seismic data show that the basin was filled with lateral sediment accretion (shelf- edge progradation) during the Pannonian (Fig. 2). The original deposi- tional surfaces appear as consecutive clinoforms in the seismic images, reflecting shelf-edge progradation from all around the basin margin towards its center (e.g. Trkulja and Kirin, 1984; Marton, 1985; B´erczi and Phillips, 1985; Pog´acs´as, 1987; Horv´ath and Pog´acs´as, 1988; Mat- tick et al., 1988; Pog´acs´as et al., 1988, 1992, 1993, 1994; Juh´asz, 1992;
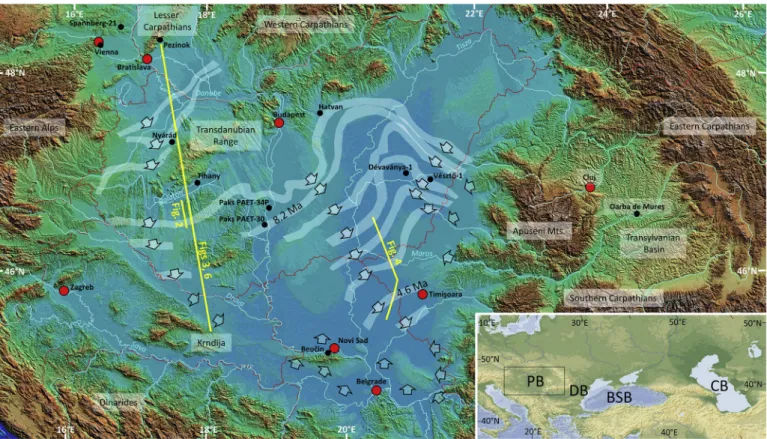
Fig. 1. Present-day topography of the Pannonian basin with dip directions (indicated by arrows) of late Neogene shelf-edge scale clinoforms as observed in seismic data. White bands indicate the geographical position of the shelf-edge slopes (where they could be mapped) at consecutive moments of time (according to Magyar et al., 2013). Lack of data around the intra-basinal highs and inselbergs is due to either shallow-water sedimentation or Quaternary erosion. The 8.2 Ma and 4.6 Ma old slopes were dated by seismic correlation from the magnetostratigraphic reference drill cores of PAET-34P and PAET-30, and of D´evav´anya-1 and V´eszt˝o-1, respectively. Black dots indicate locations of boreholes and outcrops referred to in the text. International boundaries and largest cities (for reference) are in red. Inset:
The Paratethyan basins referred to in the text: PB: Pannonian basin, DB: Dacian basin, BSB: Black Sea basin, CB: Caspian basin. (For interpretation of the references to colour in this figure legend, the reader is referred to the web version of this article.)
Ujsz´aszi and Vakarcs, 1993; Csat´o, 1993; Molenaar et al., 1994; Sacchi et al., 1998, 1999; Safti´c et al., 2003; Magyar et al., 2007; T´oth-Makk, 2007; Csat´o et al., 2007, 2013, 2015; Mat¸enco and Radivojevi´c, 2012;
Sztan´o et al., 2013a, 2016; Bal´azs et al., 2018). Progradation from the northwest, supplied by the paleo-Danube River system, was faster and thus covered a longer traverse than progradation from the other di- rections (SW, S, E, and NE; Fig. 3). As a consequence, the last deepwater setting where sediment transport systems eventually met and completed the filling of the basin was located in the SE part of the basin (Fig. 1;
Pog´acs´as and R´ev´esz, 1987; Vakarcs et al., 1994; Magyar et al., 2013; ter Borgh et al., 2014).
Lake Pannon clinoforms are typically 200–600 m high, indicating an original water depth of several hundred metre to more than one thou- sand metre in the various sub-basins (Bal´azs et al., 2018). The width of the slope is 5–15 km, with a dip of 3–5◦in its upper part. Individual clinothems are usually smaller than 25 km along strike (Mattick et al., 1988) and reflect frequent avulsion of the sediment entry points (Uhrin et al. 2009). The clinothems are organized into alternating prograda- tional and aggradational clinothem sets, resulting in flat and ascending
shelf-edge trajectories (Uhrin and Sztan´o, 2012; Sztan´o et al., 2013a).
These fourth-order sedimentary cycles, probably controlled by the climate (Juh´asz et al., 2007; Sztan´o et al., 2013a; Gong et al., 2019), are expressed in the topset shelf deposits as parasequence sets (Sztan´o et al., 2013b). The aggradational clinothem sets contain thick sandy bottom- sets, whereas the progradational ones often onlap on older surfaces at, or not far beyond, the toe of slope (Sztan´o et al., 2013a; Gong et al., 2019).
3. Chronostratigraphy of the upper Neogene clinothem-bearing basin fill in the Pannonian basin – A brief history
The non-marine nature of the upper Neogene sedimentary forma- tions in the Pannonian basin, as well as those in the entire Paratethyan area that was compartmentalized into large brackish lacustrine basins in the late Neogene (Palcu et al., 2021), lead to significant uncertainties in their correlation to the standard stages and to the geologic time scale.
The upper Neogene of the Central Paratethys was divided into four chronostratigraphic stages in 1975: Pannonian, Pontian, Dacian, and Romanian, to establish a uniform chronostratigraphic nomenclature and Fig. 3. Schematic cross section displaying the large-scale clinoform architecture of the Pannonian (upper Miocene and Pliocene) basin infill, with highlighted foresets along a 300 km long profile from the northern basin margin (Lesser Carpathians in Slovakia) to the southern margin (Krndija hills in Croatia). In addition to the seismic line in Fig. 2, the drawing is based on seismic sections published by Ujsz´aszi and Vakarcs (1993), Sacchi et al. (1998, 1999), Magyar et al. (2007), Uhrin and Sztan´o (2012), T¨or˝o et al. (2012), Sztan´o et al. (2016), Sebe et al. (2020), and subsurface data archive of MOL Plc. Orange colored basement represents all pre- Pannonian (older than upper Miocene) formations. For simplicity, the thin Quaternary cover is not indicated. The vertical scale is approximate. Location of pro- file in Fig. 1.
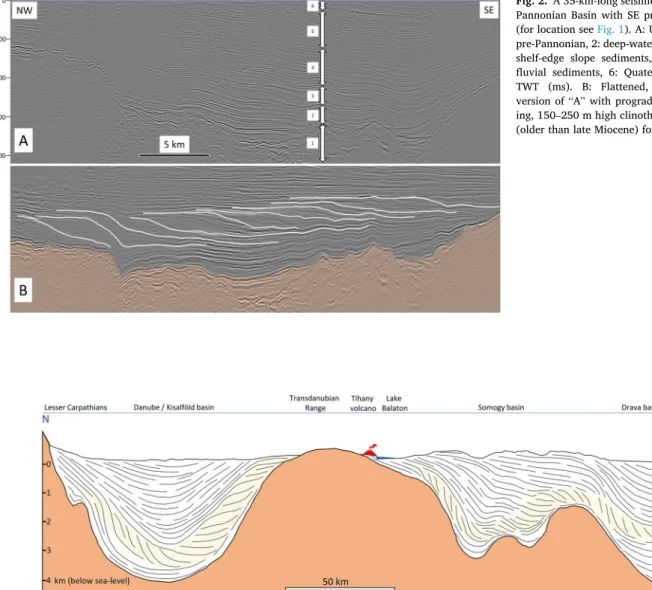
Fig. 2.A 35-km-long seismic profile from the central Pannonian Basin with SE prograding clinoform sets (for location see Fig. 1). A: Uninterpreted version. 1:
pre-Pannonian, 2: deep-water marls and turbidites, 3:
shelf-edge slope sediments, 4: deltaic deposits, 5:
fluvial sediments, 6: Quaternary. Vertical scale in TWT (ms). B: Flattened, vertically exaggerated version of “A” with prograding and slightly aggrad- ing, 150–250 m high clinothems. The pre-Pannonian (older than late Miocene) formations in orange.
shallow-water mollusc biostratigraphy was the most important tool.
Four books subsequently were published under the auspices of the In- ternational Union of Geological Sciences - Regional Committee of Mediterranean Neogene Stratigraphy to define, describe, and charac- terize each chronostratigraphic stage (Papp et al., 1985; Stevanovi´c et al., 1990; Marinescu and Papaianopol, 1995; Papaianopol et al., 2003). The definition of the Pontian, Dacian and Romanian stages, however, came from the Eastern Paratethys, and their identification and
correlation in the Pannonian Basin remained highly subjective.
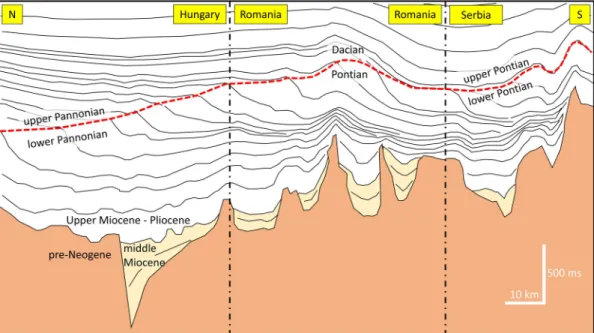
The trouble was further enhanced when the distinct lithological and paleontological boundary observed in exploration boreholes at the transition from offshore to deltaic deposits was interpreted by oil geol- ogists as a result of a basin-wide base-level drop and thus a chro- nostratigraphic boundary. This view was maintained even after the early seismic surveys imaged the clinoform geometry of the basin fill, because the foresets often showed a toplap indicating erosion. This is a common Fig. 4. A simplified line drawing of a 145 km long composite seismic time profile from the eastern part of the Pannonian basin, based on seismic sections published by Magyar et al. (2006), Sztan´o et al. (2013a), Tilita et al. (2007), Mat¸enco and Radivojevi´c (2012) and subsurface data archive of MOL Plc. The line crosses two international borders (for location see Fig. 1). Orange: pre-Neogene basement, pale orange: middle Miocene, white: upper Miocene–Pliocene. The vertical scale is in two-way-time. The top of the profile is at about 900 m below sea-level, and the deepest point of the upper Miocene–Pliocene basin fill in the middle of the Hungarian section is at about 5500 m below sea-level. The boundary between the foreset and topset parts of the clinoforms, indicated by a dotted red line, has been traditionally interpreted as a major chronostratigraphic boundary. In Hungary, Romania, and Serbia, different regional (sub)stage names (chronostratigraphic terms) have been applied for the same diachronous lithostratigraphic surface. (For interpretation of the references to colour in this figure legend, the reader is referred to the web version of this article.)
Fig. 5.A depth profile from Lake Pannon displaying the relationship between seismic-scale morphology, depositional environments, and habitat and diversity of molluscs and dinoflagellates.
phenomenon caused by the dynamics of sedimentologial processes in highly progradational settings, but a more careful analysis reveals its local nature that excludes the interpretation of regional erosional sur- faces obstinately adhered to the foreset-topset boundary (ter Borgh et al., 2014). This boundary was correlated with different chro- nostratigraphic surfaces in different countries: it was thought to corre- spond to the base of the “upper Pannonian” in Hungary, to that of the
“upper Pontian” in the former Yugoslavia (Slovenia, Croatia, Serbia), and that of the “Dacian” in Romania (Fig. 4). Apart from the different terminologies used in different countries, there was consensus about the
chronostratigraphic marker status of the slope/shelf boundary, which was in fact a highly diachronous surface across the basin. The resulting conceptual and terminological confusion required a solution with a novel approach.
4. Pannonian biochronology and correlations to the geologic time scale
4.1. Relative biochronology
The highly endemic biota of Lake Pannon displayed significant ver- tical gradient along the paleo-depositional surfaces (clinoforms; Fig. 5).
The littoral to shallow sublittoral zones on the shelf (clinoform topsets) were inhabited by an extremely diverse mollusc fauna, whereas the several-hundred-meter-deep profundal environment (clinoform bot- tomsets) was scarcely populated by a few highly adapted deepwater dwellers (Korp´as-H´odi et al., 1992; Juh´asz and Magyar, 1992; Geary et al., 2000). In contrast, fossil dinocyst algal assemblages are diverse in the profundal sediments and impoverished in shallow sublittoral de- posits. Consequently, molluscs can be better used to biostratigraphically subdivide the shallow water sediments, whereas dinoflagellates are more useful in the offshore deposits (Magyar et al., 1999; Magyar and Geary, 2012).
In the littoral deposits of Lake Pannon, a series of taxon-range zones (or rather lineage zones) were established on the basis of subsequent appearances of new morphologies within apparently anagenetic evolu- tionary lineages (Müller and Magyar, 1992a; Geary et al., 2002, 2010).
The highest temporal resolution is offered by the littoral cockle lineage starting with Lymnocardium edlaueri and ending with Prosodacnomya vodopici (Müller and Magyar, 1992b; Magyar et al., 1999, 2000; Magyar and Geary, 2012; Fig. 6). By extending each of these shallow water mollusc biozones from the topset (where the given species actually oc- curs) to the entire seismically delineated clinothem, it became possible to determine the physical position of the mollusc biostratigraphic units within the basin (Fig. 7). Correlation of these units with the geologic time scale was attempted by a combination of mammal biostratigraphic correlation, radioisotopic dating, and magnetostratigraphy.
4.2. Correlations to land mammal biozones
The shallow lacustrine deposits and/or the immediately overlying deltaic and fluvial sediments often yield mammal remains. The Euro- pean Neogene mammal “zones” (MN zones) were originally introduced as consecutively numbered typical mammal associations (“reference faunas”; Mein, 1975) separated by distinct evolutionary and migration events, but there is a tendency to use them as “normal” biozones in stratigraphy (Agustí et al., 2001). The upper Miocene to Pliocene in- cludes zones MN8 to MN16, assigned into the mammal “stages” of Astaracian (MN7-8), Vallesian (MN9-10), Turolian (MN11-13), Rusci- nian (MN14-15), and Villanyian (MN16-17, the latter being partly Pleistocene). The mammal stages and zones were dated magnetostrati- graphically in Spain, and in spite of the obvious asynchronicity of some individual first occurrences across geographical-paleoecological gradi- ents (Van der Meulen et al., 2011), these dates are commonly accepted and used in the entire area of Europe (Hilgen et al., 2012). Thus, the Lake Pannon mollusc biozones can be associated with various “mammal zones”, and their age can be constrained by mammal biochronological data (Table 1, Fig. 8).
4.3. Radioisotopic dating
Only a few radiometric ages can be directly linked to the fossiliferous sedimentary successions and biozones of the Pannonian Stage. Some of them were obtained in the 1980s and 1990s with K-Ar method, and have large errors (e.g. Bal´azs and Nusszer, 1987; Kov´acs, 1992). Two Ar-Ar datasets, however, gave substantial support for dating the Lake Fig. 6.Members of the shallow-water-dwelling Prosodacnomya lineage from
Lake Pannon. These cockles apparently display anagenetic evolution and extended gradual morphological change, thus they are excellent markers for lineage zones. From oldest to youngest: A: Lymnocardium edlaueri; B: Lymno- cardium ponticum; C: Lymnocardium decorum; D: Prosodacnomya carbonifera; E:
Prosodacnomya dainellii; F: Prosodacnomya vutskitsi; G: Prosodacnomya vodopici.
Pannon sediments.
Vasiliev et al. (2010) dated two volcanic ash layers from the top of the marine middle Miocene Sarmatian Stage that directly underlies the Pannonian succession. The samples were taken at Oarba de Mures¸, Transylvanian basin (Fig. 1), where a more than 1 km thick Sarmatian is overlain by Pannonian sediments in a continuous, deep-water succession (Sztan´o et al., 2005). The calculated 40Ar/39Ar ages were 11.62 ±0.12 Ma and 11.64 ±0.14 Ma (Vasiliev et al., 2010), indicating that the Sarmatian/Pannonian boundary roughly corresponds to the 11.63 Ma old Serravallian/Tortonian boundary (Fig. 8; Hilgen et al., 2012).
The other dataset refers to the Tihany maar volcano at Lake Balaton (N´emeth et al., 1999, 2001). Radiometric dating of this volcanic body gave first 7.92 ±0.22 Ma by K-Ar method (Balogh and N´emeth, 2005), then 7.96 ±0.03 Ma by Ar-Ar method (Fig. 6; Wijbrans et al., 2007). The volcanic body overlies the L. decorum Zone. An allochtonous sedimen- tary inclusion, i.e. a piece of country rock within the volcanic body, however, contained specimens of Prosodacnomya carbonifera, indicating that the First Appearance Datum (FAD) of Prosodacnomya is older than the volcano (Sztan´o et al., 2013b). Therefore, the FAD of Prosodacnomya can be dated as slightly older than 7.96 Ma (Figs. 7, 8).
Most recently, the authigenic 10Be/9Be dating method is being applied to lake Pannon sediments and overlying fluvial deposits (ˇSujan et al., 2016, 2017). The first results are encouraging; they do reflect the heterochronous evolution of sedimentary environments, but for the time being, they are not calibrated against other geochronometric data (such
as Ar-Ar dating or magnetostratigraphic interpretations).
4.4. Magnetostratigraphy
Magnetostratigraphic investigations in the Pannonian basin started in the late 1970s, when the thick Pleistocene to Pliocene sedimentary fill of eastern Hungary was cored continuously in wells D´evav´anya-1 and V´eszt˝o-1, in 1116 and 1200 m length, respectively (Fig. 1; Cooke et al.
1979). These drillings did not reach the sediments of Lake Pannon; they ended in fluvial Pliocene deposits overlying the lacustrine layers.
Seismic correlation showed, however, that the lowermost part of the cored fluvial interval stratigraphically corresponds to the shelf-edge clinoform of Lake Pannon some 120–130 km to the S. According to the magnetostratigraphic interpretation of the cores, sedimentation in this part of the basin has been continuous since at least the early Plio- cene, and the polarity zones could be correlated with the global polarity timescale without any difficulties down to the Gauss/Gilbert boundary (base of C2An.3n, 3.6 Ma; Cooke et al., 1979; Elston et al., 1994; N´ador et al., 2003). Correlation of the older polarity zones was less straight- forward, but according to the most preferred alternative, the lowermost identified polarity change is correlated with the base of the Nunivak subzone (C3n2n, 4.63 Ma; Elston et al., 1994; Fig. 8). The mineral car- rying the magnetic signal in the Quaternary fluvial sediments was proved to be magnetite (Püsp¨oki et al., 2016). As the Pliocene and Quaternary fluvial sedimentary facies are similar to each other, it is Fig. 7. Schematic cross section displaying the spatial distribution of chronostratigraphic units (clinothem sets) with different colors along the profile of Fig. 3. The units are defined by littoral biozones in their topsets: A: Lymnocardium edlaueri Zone; B: Lymnocardium ponticum Zone; C: Lymnocardium decorum Zone; D: Proso- dacnomya carbonifera Zone; E: Prosodacnomya dainellii Zone; F: Prosodacnomya vutskitsi Zone; G: Prosodacnomya vodopici Zone (the littoral marker fossils do not occur in deep-water deposits). Basinal sediments with no traceable connection with clinoforms and fluvial deposits in the top of the sequence are not assigned to any of the biochronozones and thus remain white. Orange colored basement represents all pre-Pannonian (older than upper Miocene) formations. For simplicity, the thin Quaternary cover is not indicated. Age intervals of the identified mammal zones, the radioisotopic age of the Tihany volcano, and magnetostratigraphic data from the Paks drill cores PAET-30 and PAET-34P (projected along strike from 100 km) are also indicated. Dating of the base of the lacustrine Pannonian stage came from magnetostratigraphy and Ar-Ar geochronology (see the text for details and references). Location of profile in Fig. 1.
Table 1
Examples of correlation between land mammal biozones and Pannonian littoral mollusc marker species (biozones).
Locality example MN Zone Age (Ma)
(Hilgen et al., 2012)
Reference (latest) Mollusc species/ zone Reference (latest)
Vienna (V¨osendorf,
Hennersdorf) MN9 11.2–9.9 Harzhauser et al.
(2004) Lymnocardium edlaueri Papp, 1985, Harzhauser and Mandic (2004)
Pezinok MN10 (lower
part) 9.9–9.6 Joniak (2016) Lymnocardium edlaueri (upper
part) Fordinal (1997)
Ny´ar´ad MN11 8.9–7.6 Kordos (1987) Lymnocardium ponticum Strausz (1942)
Tihany MN11 Kordos (1987) Lymnocardium decorum Sztan´o et al. (2013b)
Hatvan MN12 7.6–6.8 Bernor et al. (2003) Prosodacnomya dainellii Müller and Magyar (1992b)
reasonable to suppose that detrital magnetite is the major magnetic mineral in the Pliocene succession as well.
Later magnetostratigraphic research of Lake Pannon deposits yielded more ambiguous results. The investigated cores were drilled in marginal locations, where obvious unconformities hindered the straightforward correlation with the Global Polarity Timescale (GPTS). These correla- tions were usually based on the identification of the exceptionally long normal polarity zone C5n (if penetrated; e.g. Lantos et al., 1992; Elston et al., 1994). Although the correlations usually gave meaningful results, precise seismic and biostratigraphic interpretations often revealed sig- nificant controversies between the correlations (Magyar et al., 1999, 2007). It turned out that the main magnetic carrier in the lacustrine to deltaic deposits of Lake Pannon is greigite, a diagenetic mineral (Babinszki et al., 2007). When formed shortly after deposition, greigite may reflect the original polarity that prevailed during sedimentation.
When it is late diagenetic, however, its polarity can be irrelevant to the polarity prevailing during sedimentation. Therefore, an uncontrolled factor of uncertainty was introduced into all these measurements and interpretations.
In spite of these uncertainties, several attempts were made to mag- netostratigraphically date the Sarmatian/Pannonian boundary (Vasiliev et al., 2007, 2010; Paulissen et al., 2011; ter Borgh et al., 2013), and two of these were successful. Using a paleomagnetic logging tool for the downhole measurement of the magnetic field in borehole Spannberg-21 in the Vienna basin, an 1800-m-long paleomagnetic record was attained from the middle and upper Miocene. Correlation with the GPTS showed that the base of the Pannonian Stage is 11.6 Ma old (Paulissen et al., 2011; Fig. 8). Similarly, the magnetostratigraphic investigation of the Beoˇcin section in northern Serbia lead to the interpretation that the base of the Pannonian Stage is 11.63 ±0.04 Ma old (ter Borgh et al., 2013;
Fig. 8).
Most recently, Kelder et al. (2018) investigated four several- hundred-meter long drill cores from the vicinity of Paks, central Hungary (Fig. 1). Thermal demagnetization of the samples revealed composite behavior of the natural remanent magnetization, where the low-temperature and high-temperature components had the same po- larity, recording the early diagenetic greigite formation, whereas the medium-temperature component had an antiparallel polarity, repre- senting a late diagenetic overprint. Based on the low-temperature and high-temperature components, magnetostratigraphic interpretation of two cores (PAET-30 and PAET-34P) was possible. The two drillings are 18 kms apart (Fig. 1), but seismic correlation between them supported the synchronicity of the polarity zones in the cores, which represent the time interval of ca. 9 Ma to ca. 6.8 Ma. The abundant fossil material from the cores ensured the age calibration of several biozones: for instance, the base of the Prosodacnomya carbonifera Zone at 8.1 Ma, that of the P. dainellii at 7.6 Ma, and of the P. vutskitsi at 7.1 Ma (Magyar et al., 2019) (Figs. 7, 8). The shelf-edge slope at the location of PAET-30 was dated as 8.2 Ma (Fig. 1).
4.5. State of the art geochronology of the Pannonian Stage
For the time being, the geochronology of the Pannonian Stage thus depends on 4 pivotal points: 1) the Sarmatian/Pannonian boundary at 11.6 Ma, based on Ar-Ar dating and magnetostratigraphy; 2) the Tihany volcano radiometric age at 7.96 Ma, restricting the age of the underlying littoral mollusc biozones; 3) the Paks magnetostratigraphic record spanning 8.77 Ma to 7.14 Ma, age-calibrating several mollusc and dinoflagellate biozones; and 4) the D´evav´anya and V´eszt˝o magneto- stratigraphic record, spanning 4.63 Ma to 2.59 Ma (Fig. 8). The cali- brated biozones and seismic horizons allow long-distance chronostratigraphic correlations and dating in far-away locations of the Pannonian basin. Where the seismic projection of these pivotal data is not available, mollusc and mammal geochronology facilitates the age estimation of the sediments. The accuracy of these estimations is vari- able, but the error is usually less than 500 kys. Further enhancement of dating resolution can be expected from discovery and dating of volcanic ash layers in the sedimentological record and, possibly, from the future calibration of the authigenic 10Be/9Be data.
5. The other Paratethyan basins – A discussion
Chronostratigraphic practices where diachronous lithological or lithostratigraphical boundaries within clinothems are considered syn- chronous surfaces may occur in many sedimentary basins worldwide, and they do occur in other Paratethyan basins. In the late Neogene succession of the Dacian basin (Fig. 1), for instance, the base of the Pliocene Dacian stage above the underlying Pontian is defined by the first appearance of the endemic shallow-water cockle Pachydacna (Marinescu and Papaianopol, 1995). In the deep subsurface, the offshore to deltaic transition is identified with this Pontian-Dacian boundary (e.g.
Jipa and Olariu, 2009; Casas-Gallego et al., 2020). As the basin is filled by clinothems, however (e.g. Leever et al., 2009; Fongngern et al., 2015;
Magyar et al., 2020), the deep to shallow water transition, and thus the possible first occurrence of Pachydacna in boreholes, have a different age from one location to the other.
Similar patterns may characterize the upper Neogene and Pleisto- cene stratigraphy in the Black Sea and Caspian basins as well (Fig. 1).
The best candidates are superpositions where predominantly muddy offshore formations (“stages”) are overlain by sandy shallow-water formations (“stages”). Synchronous basinwide regression producing such successions can be a real phenomenon, but it is often confused with normal regression by prograding clinothem formation, a process that can be slow and lengthy, resulting in significantly diachronous deep/
shallow transitions both in lithological characteristics and in the envi- ronmentally sensitive, benthic paleontological record across the basin.
Fig. 8.Time scale for the deposits of Lake Pannon with the key chronostrati- graphic and geochronological data discussed in the text. Correlation between Lake Pannon mollusc biochronology and magnetostratigraphic data was carried out only in the Paks cores (indicated by gray). The geomagnetic polarity time scale is according to Raffi et al. (2020). C.P.: Central Paratethys; mM: middle Miocene; Sar: Sarmatian.
In the southern Caspian, for instance, seismic data show that the Akchagylian – Apsheronian transgressive-regressive sedimentary cycle produced several hundred m high clinothem sets (e.g. Abdullayev et al., 2012). The Akchagylian is usually described as a transgressive, mud- prone brackish marine sediment, whereas the Apsheronian is consid- ered a regressive, shallower, brackish lacustrine depositional sequence (e.g. Richards et al., 2018; Krijgsman et al., 2019; Lazarev et al., 2020).
Although diachrony was recognized at the base of the Akchagylian Stage defined by a flooding event (Forte et al., 2015), clinothem building as a possible source of diachrony within the Akchagylian–Apsheronian suc- cession was not addressed by any of the above field-based studies. Thus, the position of the Akchagylian-Apsheronian and lower Apsheronian- upper Apsheronian boundaries within the clinothems remains elusive:
are they chronostratigraphic (synchronous) or lithostratigraphic (prob- ably diachronous) in nature?
In both the Dacian and Caspian examples above, the identification of stage and substage boundaries in the subsurface would be possible by tracing biochronologically calibrated seismic surfaces from the topsets to the bottomsets of clinoforms, similarly to the biochronostratigraphic subdivision of the Pannonian Basin system. There is tremendous litera- ture on the late Miocene, Pliocene and Pleistocene fossils of the Dacian, Black Sea and Caspian basins, upon which the chronostratigraphic units (regional stages) were defined (e.g. Nevesskaja et al., 2001; Popov et al., 2016), and these species are often closely related to the late Miocene biota of Lake Pannon (e.g. Magyar, 2015; Richards et al., 2018; Neu- bauer et al., 2020). The Eastern Paratethyan species, however, are still not known well enough in terms of their paleoecology and phylogenetic relationships. Chronostratigraphy of the shelf-edge scale clinothems, however, cannot be assessed paleontologically without considering the paleoecological changes along a clinoformal surface from profundal to shallow-water environments, and the evolutionary changes within each environment.
6. Conclusions
Regression, i.e. basinward migration of the shoreline, may take a significant lapse of time before the entire basin is filled with shelf-edge scale clinothems. In such basins, the chronostratigraphic boundaries are clinoform surfaces which can be easily tracked in the subsurface on seismic sections but are difficult to observe in the field or in well logs, and they should not be confused with the major lithological and bio- facies changes that occur in each clinothem at (or close to) the shelf break. In the basin fills of long-lived lakes, which were abundant in the Neogene of the Paratethyan area (in the Pannonian, Dacian, Black Sea and Caspian basins), this diachronous shelf break transition was iden- tified with traditional regional chronostratigrahic (stage and substage) boundaries that had been defined in outcrops of the topset deposits. At the same time, however, the evolution of the rich endemic biota in these basins offers good stratigraphic markers if the paleontological changes are interpreted laterally, following the accretion of the shelf edge. The clinothem and biozone boundaries can be dated by mammal strati- graphic correlation, magnetostratigraphy, and radiometric dating methods and then tracked along clinoformal seismic surfaces into the deep subsurface. In the late Miocene to Pliocene (Pannonian) brackish lacustrine deposits of the Pannonian basin, dating with such methods usually bears a several 100 kyr error, except for the base of the lacustrine sequence (11.6 Ma) and the 9–6.8 Ma and 4.6–2.6 Ma intervals, where high-quality magnetostratigraphy in drill cores decreases the uncer- tainty to less than 100 ky and age-calibrates the dense reflection seismic network of the basin.
Declaration of Competing Interest I declare no conflict of interest.
Acknowledgements
MOL Hungarian Oil and Gas Plc. is thanked for securing access to subsurface data. The pictures of fossils in Fig. 5, Fig. 6 are from the unique photo archive of Lajos Tam´as Katona, Bakony Museum of the Hungarian Natural History Museum, Zirc. Vikt´oria Baranyi kindly pro- vided information and pictograms of dinocysts for Fig. 5. David Houseknecht, Cornel Olariu, an anonymous reviewer, volume editor Liviu Matenco and guest editor Attila Bal´azs are thanked for their very careful and useful reviews of the earlier versions of the paper. This is MTA-MTM-ELTE Paleo contribution No. 339, supported by the Hun- garian National Research, Development and Innovation Office NKFI 116618 project.
References
Abdullayev, N.R., Riley, G.W., Bowman, A.P., 2012. Regional controls on lacustrine sandstone reservoirs: the Pliocene of the South Caspian Basin. In: Baganz, O.W., Bartov, Y., Bohacs, K., Nummedal, D. (Eds.), Lacustrine Sandstone Reservoirs and Hydrocarbon Systems, AAPG Memoir, vol. 95, pp. 71–98.
Agustí, J., Cabrera, L., Garc´es, M., Krijgsman, W., Oms, O., Par´es, J., 2001. A calibrated mammal scale for the Neogene of Western Europe. State of the art. Earth Sci. Rev.
52, 247–260.
Babinszki, E., M´arton, E., M´arton, P., Kiss, L.F., 2007. Widespread occurrence of greigite in the sediments of Lake Pannon: implications for environment and
magnetostratigraphy. Palaeogeogr. Palaeoclimatol. Palaeoecol. 252, 626–636.
Bal´azs, E., Nusszer, A., 1987. Unterpannonischer Vulkanismus der Beckengebiete Ungarns. Ann. Hungarian Geol. Inst. 69, 95–113.
Bal´azs, A., Magyar, I., Matenco, L., Sztan´o, O., T˝ok´es, L., Horv´ath, F., 2018. Morphology of a large paleo-lake: analysis of compaction in the Miocene-Quaternary Pannonian Basin. Glob. Planet. Chang. 171, 134–147. https://doi.org/10.1016/j.
gloplacha.2017.10.012.
Balogh, K., N´emeth, K., 2005. Evidence for the Neogene small-volume intracontinental volcanism in Western Hungary: K/Ar geochronology of the Tihany Maar Volcanic Complex. Geol. Carpath. 56, 91–99.
B´erczi, I., Phillips, R.L., 1985. Process and depositional environments within Neogene deltaic-lacustrine sediments, Pannonian Basin, Southeast Hungary. Geophys. Trans.
31, 55–74.
Bernor, R.L., Feibel, C., Viranta, S., 2003. The vertebrate locality of Hatvan, Late Miocene (Middle Turolian, MN12), Hungary. In: Petculescu, A., Stiuc˜a, E. (Eds.), Advances in Vertebrate Paleontology “Hen to Panta”. Emil Racovitza Institute of Speleology, Bucharest, pp. 105–112.
Casas-Gallego, M., P´erez-Rodríguez, I., Fenton, J.P.G., Gold, D.P., Marza, A., Tudor, E., 2020. Integrated biostratigraphic, palaeoenvironmental and wireline log characterisation of upper Miocene to lower Pliocene successions of the eastern Dacian Basin (SE Romania). Newsl. Stratigr. 53, 71–92. https://doi.org/10.1127/
nos/2019/0531.
Cooke, H.B.S., Hall, J.M., R´onai, A., 1979. Paleomagnetic, sedimentary and climatic records from boreholes at D´evav´anya and V´eszt˝o, Hungary. Acta Geol. Hung. 22, 89–109.
Csat´o, I., 1993. Neogene sequences in the Pannonian basin, Hungary. Tectonophysics 226, 377–400.
Csat´o, I., Kendall, C.G., Moore, P.D., 2007. The Messinian problem in the Pannonian Basin, Eastern Hungary – Insights from stratigraphic simulations. Sediment. Geol.
201, 111–140.
Csat´o, I., Granjeon, D., Catuneanu, O., Baum, G.R., 2013. A three-dimensional stratigraphic model for the Messinain crisis in the Pannonian Basin, eastern Hungary. Basin Res. 25, 121–148. https://doi.org/10.1111/j.1365- 2117.2012.00553.x.
Csat´o, I., T´oth, S., Catuneanu, O., Granjeon, D., 2015. A sequence stratigraphic model for the Upper Miocene–Pliocene basin fill of the Pannonian Basin, eastern Hungary.
Mar. Pet. Geol. 66, 117–134. https://doi.org/10.1016/j.marpetgeo.2015.02.010.
Elston, D.P., Lantos, M., H´amor, T., 1994. High resolution polarity records and the stratigraphic and magnetostratigraphic correlation of Late Miocene and Pliocene (Pannonian s.l.) deposits of Hungary. In: Teleki, P.G., Mattick, R.E., K´okay, J. (Eds.), Basin Analysis in Petroleum Exploration. A case study from the B´ek´es basin, Hungary. Kluwer Academic Publishers, Dordrecht, pp. 111–142.
Fongngern, R., Olariu, C., Steel, R.J., Kr´ezsek, Cs, 2015. Clinoform growth in a Miocene, Paratethyan deep lake basin: thin topsets, irregular foresets and thick bottomsets.
Basin Res. 28, 770–795.
Fordinal, K., 1997. Molluscs (Gastropoda, Bivalvia) from the Pannonian deposits of the western part of Danube Basin (Pezinok clay-pit). Slovak Geol. Mag. 3, 263–283.
Forte, A.M., Sumner, D.Y., Cowgill, E., Stoica, M., Murtuzayev, I., Kangarli, T., Elashvili, M., Godoladze, T., Javakhishvili, Z., 2015. Late Miocene to Pliocene stratigraphy of the Kura Basin, a subbasin of the South Caspian basin: implications for the diachroneity of stage boundaries. Basin Res. 27, 247–271. https://doi.org/
10.1111/bre.12069.
Gani, M.R., 2017. Mismatch between time surface and stratal surface in stratigraphy.
J. Sediment. Res. 87, 1226–1234.
Gani, M.R., Bhattacharya, J.P., 2005. Lithostratigraphy versus chronostratigraphy in facies correlations of Quaternary deltas: Application of bedding correlation. In:
Bhattacharya, J.P., Giosan, L. (Eds.), River deltas – Concepts, Models and Examples, vol. 83. SEPM Special Publication, pp. 31–48.
Geary, D.H., Magyar, I., Müller, P., 2000. Ancient Lake Pannon and its Endemic Molluscan Fauna (Central Europe; Mio-Pliocene). In: Rossiter, A., Kawanabe, H.
(Eds.), Ancient Lakes: Biodiversity, Ecology, and Evolution. Advances in Ecological Research, vol. 3, pp. 463–482.
Geary, D.H., Staley, A.W., Müller, P., Magyar, I., 2002. Iterative changes in Lake Pannon Melanopsis reflect a recurrent theme in gastropod morphological evolution.
Paleobiology 28, 208–221.
Geary, D.H., Hunt, G., Magyar, I., Schreiber, H., 2010. The paradox of gradualism:
phyletic evolution in two lineages of lymnocardiid bivalves (Lake Pannon, central Europe). Paleobiology 36, 592–614.
Gong, C., Sztan´o, O., Steel, R.J., Xian, B., Galloway, W.E., Bada, G., 2019. Critical differences in sediment delivery and partitioning between marine and lacustrine basins: a comparison of marine and lacustrine aggradational to progradational clinothem pairs. GSA Bull. 131, 766–781.
Harzhauser, M., Mandic, O., 2004. The muddy bottom of Lake Pannon – a challenge for dreissenid settlement (Late Miocene; Bivalvia). Palaeogeogr. Palaeoclimatol.
Palaeoecol. 204, 331–352.
Harzhauser, M., Daxner-H¨ock, G., Piller, W.E., 2004. An integrated stratigraphy of the Pannonian (Late Miocene) in the Vienna Basin. Aust. J. Earth Sci. 95-96, 6–19.
Henriksen, S., Hampson, G.J., Helland-Hansen, W., Johannessen, E.P., Steel, R.J., 2009.
Shelf edge and shoreline trajectories, a dynamic approach to stratigraphic analysis.
Basin Res. 21, 445–453. https://doi.org/10.1111/j.1365-2117.2009.00432.x.
Hilgen, F.J., Lourens, L.J., Van Dam, J.A., 2012. The neogene period. In: Gradstein, F.M., Ogg, J.G., Schmitz, M., Ogg, G. (Eds.), The Geologic Time Scale 2012. Elsevier B. V, pp. 923–978. https://doi.org/10.1016/B978-0-444-59425-9.00029-9.
Horv´ath, F., Pog´acs´as, Gy, 1988. Contribution of seismic reflection data to
chronostratigraphy of the Pannonian basin. In: Royden, L.H., Horv´ath, F. (Eds.), The Pannonian Basin. A Study in Basin Evolution: AAPG, Memoir, 45, pp. 97–105.
Horv´ath, F., Bada, G., Szafi´an, P., Tari, G., Ad´´ am, A., Cloetingh, S., 2006. Formation and deformation of the Pannonian Basin: constraints from observational data. In: Gee, D.
G., Stephenson, R.A. (Eds.), European Lithosphere Dynamics: Geological Society, London, Memoirs, 32, pp. 191–206.
Jipa, D.C., Olariu, C., 2009. Dacian Basin: Geo-Eco-Marina, 3. Special Publication, pp. 1–264.
Joniak, P., 2016. Upper Miocene rodents from Pezinok in the Danube Basin, Slovakia.
Acta Geol. Slovaca 8, 1–14.
Juh´asz, Gy, 1992. Pannonian s.l. formations in the Hungarian Plain: distribution, facies and sedimentary environment. F¨oldtani K¨ozl¨ony 122, 133–165.
Juh´asz, Gy, Magyar, I., 1992. Review and correlation of the Late Neogene (Pannonian s.
l.) lithofacies and mollusc biofacies in the Great Plain, eastern Hungary. F¨oldtani K¨ozl¨ony 122, 167–194.
Juh´asz, Gy, Pog´acs´as, Gy, Magyar, I., Vakarcs, G., 2007. Tectonic versus climatic control on the evolution of fluvio-deltaic systems in a lake basin, Eastern Pannonian Basin.
Sediment. Geol. 202, 72–95.
Kelder, N.A., Sant, K., Dekkers, M.J., Magyar, I., van Dijk, G.A., Lathouwers, Y.Z., Sztano, O., Krijgsman, W., 2018. Paleomagnetism in Lake Pannon: problems, pitfalls, ´ and progress in using iron sulfides for magnetostratigraphy. Geochem. Geophys.
Geosyst. 19, 3405–3429. https://doi.org/10.1029/2018GC007673.
Kordos, L., 1987. Neogene vertebrate biostratigraphy in Hungary. Ann. Hungarian Geol.
Inst. 70, 393–396.
Korp´as-H´odi, M., Pog´acs´as, Gy, Simon, E., 1992. Paleogeographic outline of the Pannonian s.l. of the southern Danube-Tisza Interfluve. Acta Geol. Hung. 35, 145–163.
Kov´acs, L., 1992. Pannonian (s.l.) and Quaternary formations of borehole B´acsalm´as 1.
In: Annual Report of the Hungarian Geological Institute of 1990, pp. 193–210.
Hydrocarbons in Hungary. In: Kov´acs, Zs (Ed.), 2018. Results and Opportunities.
Hungarian Energy and Public Utility Regulation Authority, Budapest, p. 330. ISBN 978-615-00-2316-8.
Krijgsman, W., Tesakov, A., Yanina, T., Lazarev, S., Danukalova, G., van Baak, C.G.C., Agustí, J., Alçiçek, M.C., Aliyeva, E., Bista, D., Bruch, A., Büyükmeriç, Y., Bukhsianidze, M., Flecker, R., Frolov, P., Hoyle, T.M., Jorissen, E.L., Kirscher, U., Koriche, S.A., Kroonenberg, S.B., Lordkipanidze, D., Oms, O., Rausch, L., Singarayer, J., Stoica, M., van de Velde, S., Titov, V.V., Wesselingh, F.P., 2019.
Quaternary time scales for the Pontocaspian domain: Interbasinal connectivity and faunal evolution. Earth Sci. Rev. 188, 1–40. https://doi.org/10.1016/j.
earscirev.2018.10.013.
Lantos, M., H´amor, T., Pog´acs´as, Gy, 1992. Magneto- and seismostratigraphic correlations of Pannonian s.l. (Late Miocene and Pliocene) deposits in Hungary.
Paleontologia i Evoluci´o 24-25, 35–46.
Lazarev, S., Jorissen, E.L., van de Velde, S., Rausch, L., Stoica, M., Wesselingh, F.P., Van Baak, C.G.C., Yanina, T.A., Aliyeva, E., Krijgsman, W., 2020. Magneto-
biostratigraphic age constraints on the palaeoenvironmental evolution of the South Caspian basin during the Early-Middle Pleistocene (Kura basin, Azerbaijan). Quat.
Sci. Rev. 222 https://doi.org/10.1016/j.quascirev.2019.105895.
Lease, R.O., Houseknecht, D., 2017. Timing of Cretaceous shelf margins in the Colville basin. In: Arctic Alaska (abs.): AAPG Pacific Section Annual Meeting, Anchorage, Alaska, May 21–24, 2017, pp. 51–52. http://www.searchanddiscovery.com/abstract s/pdf/2018/90302ps/abstracts/ndx_lease.pdf.
Leever, K.A., Matenco, L., Rabagia, T., Cloetingh, S., Krijgsman, W., Stoica, M., 2009.
Messinian sea level fall in the Dacic Basin (Eastern Paratethys): palaeogeographical implications from seismic sequence stratigraphy. Terra Nova 22, 12–17. https://doi.
org/10.1111/j.1365-3121.2009.00910.x.
Magyar, I., 2015. Palaeobiogeography of lacustrine cockles as a tool to reconstruct palaeogeography in the Late Neogene Paratethys. Hantkeniana 10, 143–146.
Magyar, I., Geary, D.H., 2012. Biostratigraphy in a Late Neogene Caspian-Type Lacustrine Basin: Lake Pannon, Hungary. In: Baganz, O.W., Bartov, Y., Bohacs, K., Nummedal, D. (Eds.), Lacustrine Sandstone Reservoirs and Hydrocarbon Systems:
AAPG, Memoir, 95, pp. 255–264. https://doi.org/10.1306/13291392M953142.
Magyar, I., Geary, D.H., Süt˝o-Szentai, M., Lantos, M., Müller, P., 1999. Integrated biostratigraphic, magnetostratigraphic and chronostratigraphic correlations of the Late Miocene Lake Pannon deposits. Acta Geol. Hung. 42, 5–31.
Magyar, I., Muller, P., Geary, D.H., Sanders, H.C., Tari, G.C., 2000. Diachronous deposits of Lake Pannon in the Kisalfold basin reflect basin and mollusc evolution.
Abhandlungen Der Geologischen Bundesanstalt 56, 669–678.
Magyar, I., Fogarasi, A., Vakarcs, G., Buk´o, L., Tari, G.C., 2006. The largest hydrocarbon field discovered to date in Hungary: Algyo. In: Golonka, J. Picha, F.J. (Eds.), The Carpathians and their foreland: geology and hydrocarbon resources. AAPG, Memoir 84, 619–632.
Magyar, I., Lantos, M., Ujsz´aszi, K., Kordos, L., 2007. Magnetostratigraphic, seismic and biostratigraphic correlations of the Upper Miocene sediments in the northwestern Pannonian Basin System. Geol. Carpath. 58, 277–290.
Magyar, I., Radivojevic, D., Sztan´o, O., Synak, R., Ujszaszi, K., P´ocsik, M., 2013.
Progradation of the paleo-Danube shelf margin across the Pannonian Basin during the Late Miocene and Early Pliocene. Glob. Planet. Chang. 103, 168–173. https://
doi.org/10.1016/j.gloplacha.2012.06.007.
Magyar, I., Sztano, O., Sebe, K., Katona, T.L., Csoma, V., G´ ¨or¨og, A., T´ ´oth, E., Szuromi- Korecz, A., ˇSujan, M., Braucher, R., Ruszkiczay-Rüdiger, Zs, Koroknai, B., W´orum, G., Sant, K., Kelder, N., Krijgsman, W., 2019. Towards a high-resolution chronostratigraphy and geochronology for the Pannonian Stage: Significance of the Paks cores (Central Pannonian Basin). F¨oldtani K¨ozl¨ony 149, 351–370.
Magyar, I., Krezsek, C., Tari, G., 2020. Clinoforms as paleogeographic tools:
Development of the Danube catchment above the deep Paratethyan basins in Central and Southeast Europe. Basin Res. 32, 320–331. https://doi.org/10.1111/bre.12401.
Marinescu, F., Papaianopol, I. (Eds.), 1995. Chronostratigraphie und Neostratotypen.
Neogen der Zentralen Paratethys, 9, Dacien. Editura Academiei Romane, Bucharest, p. 530.
Marton, Gy, 1985. A derecskei m´elyz´ona szeizmosztratigr´afiai vizsg´alata (Seismic stratigraphic study of the Derecske depression). Magyar Geofizika 26, 161–181 (in Hungarian, with English abstract).
Mat¸enco, L., Radivojevi´c, D., 2012. On the formation and evolution of the Pannonian Basin: constraints derived from the structure of the junction area between the Carpathians and Dinarides. Tectonics 31, TC6007. https://doi.org/10.1029/
2012TC003206.
Mattick, R.E., Phillips, R.L., Rumpler, J., 1988. Seismic stratigraphy and depositional framework of sedimentary rocks in the Pannonian basin in southeastern Hungary. In:
Royden, L.H., Horv´ath, F. (Eds.), The Pannonian Basin. A Study in Basin Evolution:
AAPG, Memoir, 45, pp. 117–145.
Mein, P., 1975. Proposition de Biozonation du N´eog`ene M´editerran´een `a partir des mammif`eres. In: Trabajos sobre Neogeno/Cuaternario, 4, p. 112. ´
Molenaar, C.M., R´ev´esz, I., B´erczi, I., Kov´acs, A., Juh´asz, Gy, Gajdos, I., Szanyi, B., 1994.
Stratigraphic framework and sandstone facies distribution of the Pannonian sequence in the B´ek´es basin. In: Teleki, P.G., Mattick, R.E., K´okay, J. (Eds.), Basin Analysis in Petroleum Exploration. A case study from the B´ek´es basin, Hungary.
Kluwer Academic Publishers, Dordrecht, pp. 99–110.
Müller, P., Magyar, I., 1992a. Continuous record of the evolution of lacustrine cardiid bivalves in the late Miocene Pannonian Lake. Acta Palaeontol. Pol. 36, 353–372.
Müller, P., Magyar, I., 1992b. Stratigraphic significance of the Upper Miocene lacustrine Cardiid Prosodacnomya (K¨otcse section, Pannonian basin, Hungary). F¨oldtani K¨ozl¨ony 122, 1–38.
N´ador, A., Lantos, M., T´oth-Makk, A., Tham´´ o-Bozs´o, E., 2003. Milankovich-scale multy- proxy records from fluvial sediments of the last 2.6 Ma, Pannonian basin, Hungary.
Quat. Sci. Rev. 22, 2157–2175.
N´emeth, K., Martin, U., Harangi, Sz, 1999. Miocene maar/diatreme volcanism at the Tihany Peninsula (Pannonian Basin): the Tihany Volcano. Acta Geol. Hung. 42, 349–377.
N´emeth, K., Martin, U., Harangi, Sz, 2001. Miocene phreatomagmatic volcanism at Tihany (Pannonian Basin, Hungary). J. Volcanol. Geotherm. Res. 111, 111–135.
https://doi.org/10.1016/S0377-0273(01)00223-2.
Neubauer, T.A., Anistratenko, O.Yu., Anistratenko, V.V., Kijashko, P., Stoica, M., van de Velde, S., Vinarski, M., Wesselingh, F.P., 2020. A Revision of the poorly known Pontocaspian Gastropod Genus Abeskunus, and its Central Paratethyan Origin. Hist.
Biol. https://doi.org/10.1080/08912963.2020.1720015.
Nevesskaja, L.A., Paramonova, N.P., Popov, S.V., 2001. History of Lymnocardiinae (Bivalvia, Cardiidae). Paleontol. J. 35 (Suppl. 3), 147–217.
Novak Zelenika, K., Malvic, T., 2011. Stochastic simulations of dependent geological variables in sandstone reservoirs of Neogene age: A case study of the Kloˇstar Field, Sava Depression. Geol. Croatica 64, 173–183.
Olariu, C., Steel, R.J., Vann, N.K., Tudor, E.P., Shin, M., Winter, R., Gan, Y., Jung, E., Almeida, F., Giacomone, G., Minisini, D., Brinkworth, W., Loss, M.L., Inigo, J., Gutierrez, R., 2020. Criteria for recognizing shelf-slope clinoforms in outcrop;
Jurassic Lajas and Los Molles formations, S. Neuqu´en Basin, Argentina. Basin Res.
32, 279–292.
Palcu, D.V., Patina, I.S., S¸andric, I., Lazarev, S., Vasiliev, I., Stoica, M., Krijgsman, W., 2021. Late Miocene megalake regressions in Eurasia. Sci. Rep. 11, 11471. https://
doi.org/10.1038/s41598-021-91001-z.
Papaianopol, I., Marinescu, F., Krsti´c, N., Macalet, R. (Eds.), 2003. Chronostratigraphie und Neostratotypen, Neogen der Zentralen Paratethys, 10, Romanien. Editura Academiei Romane, Bucharest, p. 527.
Papp, A., 1985. Die Mollusken-Fauna des Pannonien der Zentralen Paratethys. In:
Papp, A., J´ambor, ´A., Steininger, F.F. (Eds.), Chronostratigraphie und