ERIKA PÉNZES - KÓNYA
*THE EXAMINATION OF SPATIAL HETEROGENITY OF THE TERRICOLOUS BRYOPHYTE VEGETATION IN
ACIDOFIL FOREST COMMUNITIES
Abstract: The bryophyte vegetation of three acidofil forest communities of four habitats on radiolarian bedrock were investigated in the Bükk mountains in Hungary. The frequency and the spatial pattern of 14 terricolous bryophyte spe- cies were recorded in permanent plots for three years using four-term local quadrat variance (4TLQV) spatial pattern analysis. The variograms of the occured terricolous bryophyte species were calculated with a program package called PASSAGE (Pattern Analysis, Spatial Statistics and Geographyc Exegesis). The examined bryophyte species were: Dicranum scoparium, Ceratodon purpureus, Leucobryum juniperoideum, Polytrichum formosum, Poilytrichum piliferum, Hypnum cupressiforme, Hungary / bryophytes/ Acidofil forests/ Spatial pattern /Radiolarian rocks /Life forms/ Quadrat variance method Introduction
Temperate forests have diverse structures considering the levels and the species compositions in them. If we examine the structure of the zonal temperate forest communities, taking a special attention to the importance of the bryophyte layer, it can be seen that in those communities where the density of the canopy level is high and the litter is thick, the bryophyte –lichen vegetation consist of epiphytic species and of the ones which occur on the bigger rocks or on tree trunks (Orbán 1989, Longton 1992, Kürschner 1999). The bryophyte species occuring on the soil of these communities are mainly Querco-Pinion species (Boros 1968, Kovács 1975, Simon 1975, Pénzesné & Orbán 2000) so the bryophyte flora of these communities have a connection with Pinus forests, too (Pócs 1967). We can find the list of the bryophyte species of these communities in many works but there are a few quantitative analysis of the terricolous bryophyte vegetation in acidofil forest communities in Hungary. The bryophyte vegetation of some forest community types have been examined with
*Department of Botany, Eszerházy Károly College, 3300 Eger, Pf.46., Hungary. ko- nya@ektf.hu
quantitative methods in Hungarian forest areas (Orbán 1989, Ódor & Standovár 2001, 2002), but not the acidofil oak forests.
Bryophytes can be occur as long-term dominant species mainly in peatlands and some tundra environments (Oechel & Van Cleve 1986). Several bryophyte species occur with great density in habitats wich are limited in size and duration.
(Longton 1982). The spatial mosaic structure of these habitats considering the light mosaic and the moving rocky surface favours for bryophytes to grow in big patches together with lichens. Even in these habitats bryophytes can also form important components of ecosystems, where some resources are limited for higher plants or there are some extreme abiotic factors decreasing the abundance of vascular plants. Such forest communities have been developped in the Bükk mountains on radiolarian bedrock type. Where radiolarian rocks reach the surface of the rubbling, homogeous rocky slopes bryophytes and lichens have great importance in moderating the extreme microclimate of the soil surface by covering it in extended areas (Pénzesné Kónya & Orbán 2000, Pénzes-Kónya 2003, Pénzes-Kónya 2004.).
Several studies have been published about the detection of the real spatial pattern of bryophyte and lichen species (Alpert 1985, During & Lloret 1996, Söderström & Herben 1997). The most bryophyte species are growing in two dimensions, so their patches can be examined with spatial pattern analysis which had been carried out for surfaces (Dale et al. 2002). The density data of the examined species must be recorded in a grid network of a contiguous quadrat.
These types of methods are called quadrat variance methods such as Paired Quadrat Variance (PQV), Four Term Local Quadrat Variance (4TLQV) and Nine Term Local Quadrat Variance (9TLQV) (Dale 1999, Rosenberg 2003).
The aim of the study was the following:
– To investigate the bryophyte vegetation of the different acidofil forest communities on the basis of the occurance, frequency and the cover estimation of the bryophyte species.
– To examine the heterogeneity of the spatial mosaic structure of the bryophyte vegetation on the soil layer in the examined communities, using spatial pattern analyses.
– To compare the life history traits, growth form, the spatial pattern of the bryophyte species occuring with great abundance on the soil surface of the examined communities.
Materials and Methods Sampling areas
During the selection of the sampling areas some circumstances must have been taken into account:
– The areas must be located on the same bedrock type in the Bükk mountains (in this case it is radiolarian rock).
– The sampling areas must be close to each other so there is no big dispersal limitation for the propagules of bryophytes and the macroclimatic fluctuations are the same considering the sampling areas.
– All of the sampling areas must be forest communities in which there are not big vertical heterogeneity on the soil surface and the mean cover of the bryophyte species is more than 50%. The shrub layer in the communities is missing or the abundance of it is very small (10%).
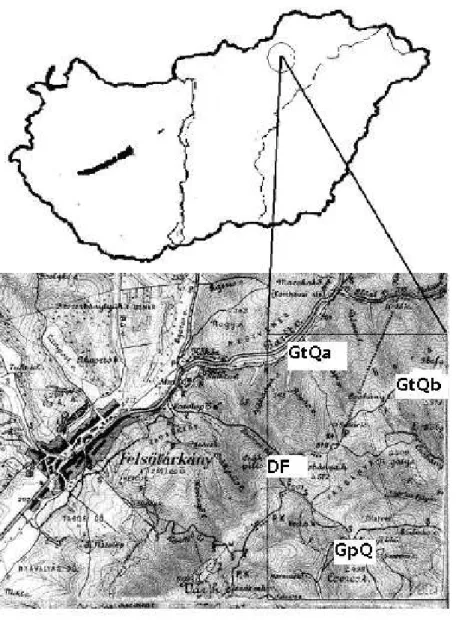
Considering these conditions the following habitats were selected for the study (Fig. 1.).
Genisto tinctoriae-Quercetum petreae Klika 1932 or Luzulo-Quercetum subcarpaticumZólyomi 1955. It is located in th Western-Northwestern slope of kis Vasbánya hegy (GtQa). The slope degree is 40°.
Deschampsio flexuosae-Fagetum silvaticaeSoó 1962 near Felsőtárkány, on the eastern slope of Csák-Pilis (DF). The slope degree is 35°.
Genisto tinctoriae- Quercetum petreae Klika 1932 or Luzulo-Quercetum subcarpaticum Zólyomi 1955. It is located in the North-Western slope of Borzlyuktető, near Bükkzsérc (GtQb). The slope degree is 40°.
Genisto pilosae-Quercetum petreae Zólyomi et al. 1958. In the Csák-pilis- Várhegy range, above the Kövesdi look-out, in Eastern-Southeastern exposition.(GpQ). The slope degree is 35°.
The dominant tree species are Quercus petrea, Quercus pubescens in Genisto pilosae- Quercetum (GpQ) and Genisto tinctoriae-Quercetum communities (GtQa and GtQb), Fagus silvatica and Quercus petrea in Deschampsio-Fagetum (DF). The canopy cover is discontinuous, about 50-70%, which provides mosaic light conditions under the canopy layer and the quantity of fallen leaves is little.
Sampling methods
There were permanent quadrat blocks pointed out in every community (Fig.
2.), they consisted of 4 touching quadrats. They were chosen randomly in every sampling areas. The size of the blocks were 60*60 cm. They were divided into cell network in wich the size of one cell was 3 *3 cm, that means that there were 400 cells of each quadrat blocks. The occuring bryophyte species were recorded in each cell, so the data show the number of the occupied cells by a certain bryophyte species that means the frequency of them. Within each cell the density data of the occuring bryophyte were recorded. Three values were used to weight density of the bryophyte species in the examined cells within a quadrat
1.the cover of the species is less than 30% within the cell,
2. the cover of the species is between 30% and 70% within the cell,
3. the cover of the species is bigger than 70% within the cell.
The names of the bryophyte species was determined by the nomenclature of Smith (1978).
Spatial pattern analysis
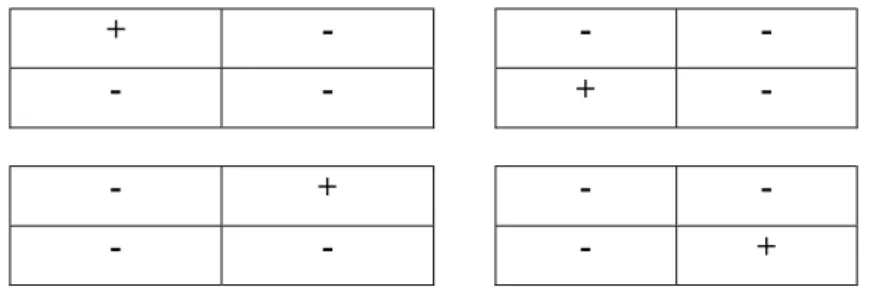
Four-Term Local Quadrat Variance method (4TLQV) was used to detect the scale of pattern of the occuring species (Dale 1999). In quadrat variance methods the variance of differences among blocks of different sizes are calculated and the pattern of the variance estimates are used to determine the scale of pattern (Rosenberg 2003). 4TLQV analysis can be applied for surfaces, so it is the 2D version of Two-Term Local Quadrat Variance (TTLQV).
The chosen and used program package was PASSAGE, Pattern Analysis, Spatial Statistics, and Geographic Exegesis (Rosenberg 2003).
The variance at block size (scale ) b, when comparing the upper left corner (UL) to the other three and y is the actual density value of the block:
VUL
(b)=
b m
j b n
i
2 1
1 2 1
1
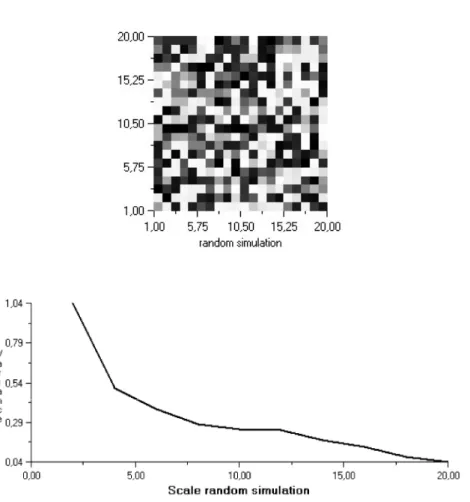
The spatial distribution of the bryophyte species are represented in grid plots in which the lightest colour means the most dense cell (density value 3) and the empty cells are black (Fig. 6a, Fig. 7.). The results of the spatial pattern analysis are represented in variograms (Fig. 6b, Fig.8.)
Results
The occuring species and their ecological features
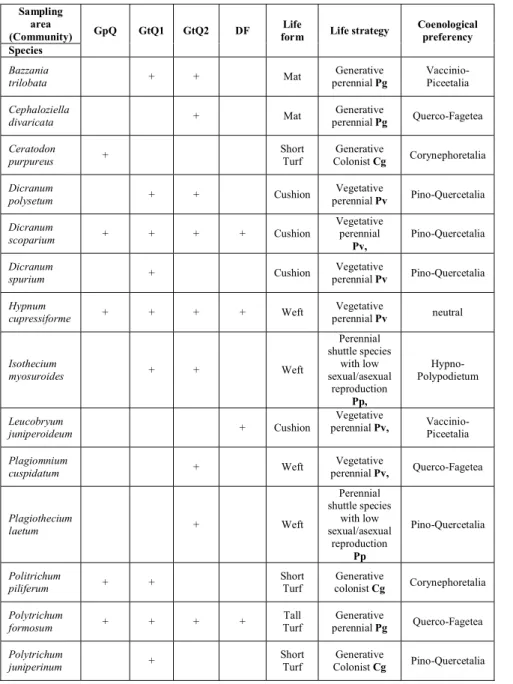
The occurance data of the terricolous bryophyte species are summerized in Table 1. The distribution of the coenological preferency (Orbán& Vajda 1983), the life strategy types (During 1979, During 1990, Kürschner 1999, Orbán 2002) and the life or growth forms of the occuring bryophyte species are summerized in Fig. 4. The most frequent life form of bryophytes are the cushions wefts and short turfs (Fig 4.a) Considering the preferred community types of the species the most bryophytes are Querco-Fagetea and Pino-Quercetalia species in the
examined areas but several species of other coenological preferencies are represented (Fig. 4.b). The distribution of the occuring bryophytes according to life strategies show that in most cases the occuring bryophytes are perennial stayers and within that group the vegetative perennial strategy is represented with the most bryophyte species such as Dicranum species and Leucobryum juniperoideum, Hypnum cupressiforme(Fig. 4.c).
The frequency distribution of the species in the communities
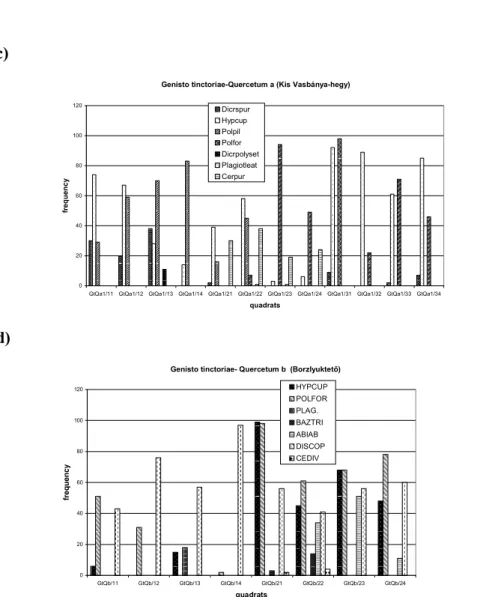
The distribution of the frequency data of the occuring bryophyte species can be seen in Fig 5. In this case frequency means the number of the occupied cells of a bryophyte species in the quadrats. One of the characteristic features of the terricolous bryophyte vegetation in the examined communities that the number of the bryophyte species is low, but the moss cover is above 50% on the soil in every community. The chosen forest communities developped on radiolarian bedrock type are similar considering the discontinuous canopy structure, the abscence of the shrub layer, the poor vascular plant abundance and the strongly acidic rubbling soil. But they differ in the exposition, the dominant tree species and in the canopy height (Kun 2000, Borhidi 2003, Pénzes-Kónya 2003, Pénzes- Kónya 2004). The occurance and frequency data of the bryophyte species differed in the four communities (Table 1. Fig. 3.) The species which occured only one or two cells in the quadrats (Plagiomnium cuspidatum, Polytrichum juniperinum) are not represented in the frequency diagrams (Fig 5.). The previ- ous quantitative analysis of these data show a relationship between the spatial terricolous bryophyte species composition of the acidofil forest community and the type of the dominant bryophyte species appering with great abundance there.
The importance of the dominant species is growing with that fact that these communities are poor in vascular plant species and the bryophyte species diversity is low as well.(Table 1. ). The dominant bryophyte species are:
Polytrichum piliferum and Ceratodon purpureusin Genisto pilosae-Quercetum community, Hypnum cupressiforme, Dicranum spurium and Dicranum polysetum in the Genisto tinctoriae-Quercetum at Kis Vasbánya hegy (GtQb), Polytrichum formosum, Hypnum cupressiforme and Dicranum scoparium at Borzlyuktető (GtQb).
The results of the fine scale spatial pattern analysis
Analysing the spatial pattern of these bryophyte species provides additional information about the spatial processes changing the bryophyte species composition on the soil surface. The spatial density data of 14 bryophyte species were recorded in 8 blocks of quadrats (There were four quadrats in each blocks) in four community stands (Fig. 1.). As one block consists of 400 cells, 3200 cells
were examined for each species. As they were permanent plots, the data were recorded in several years. The temporal changes in the spatial pattern of the bryophyte species are not discussed here, some results have been published (Pénzesné-Kónya 2003). In order to show the differences between the random spatial pattern and the real patchy pattern a grid plot of a matrix from random simulation and the variogram of the simulated data can be seen on Figure 6. The grid plots and the results of the spatial pattern analysis (4TTLQV, see methods, Fig. 3.) of 8 bryophyte species can be seen on Fig. 7. and 8.
The variograms of the 4TLQV analysis carry two kinds of information about the spatial structure of a species. One of them is the locality or the scale of the variance maximum or variance peak. In the case of the random pattern (Fig.
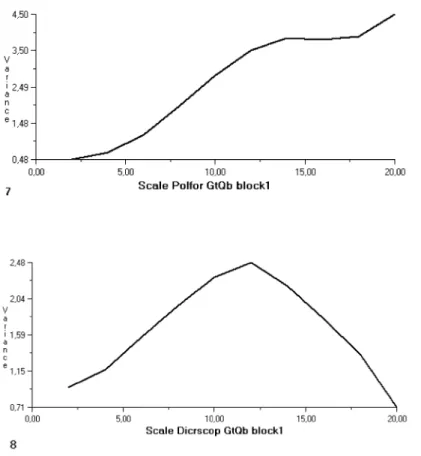
6.a) every density value (from 1 to 3) can occur in every cell with the same probability. The maximum variance value is about 1 (Fig. 6 b) is at scale 1 and then variance values are going down close to zero. In the case of a real pattern the variance is growing until a scale value and then it is falling again. Therefore the locality of the variance peak or maximum is at a bigger scale, but in many cases this scale is more than the maximum scale of the quadrat block (it is 20) and the variance may increase almost monotonically in some cases (Fig. 8.). The other information which is an important factor in the spatial structure is the value of the variance maximum. The higher is the variance maximum, the more dense, contrast patches can be observed in pattern. The grid plots (Fig. 7) and the variograms (Fig. 8.) of Ceratodon purpureus (CerpurGpQ,) Polytrichum piliferum (Polpil GpQ and Polpil GtQa) Dicranum scoparium(Dicrscop DF and Dicrscop GtQb), Leucobryum juniperoideum (Leucjun DF), Hypnum cupressiforme (Hypcup GtQa) Polytrichum formosum (Polfor GtQb).
Considering the variance peaks real pattern can be realised within the scale of the quadrat size (within scale 20) in the case of Dicranum scoparium (Fig. 8.4.:
Vmax. is at scale 13 , Fig. 8.7: Vmax. is at scale 12), Hypnum cupressiforme (Fig. 8.5.: Vmax. is at scale 12) Leucobryum juniperoideum (Fig. 8.3.: Vmax is at scale 8.). In the variogram of Ceratodon purpureus, Polytrichum formosum and Polytrichum piliferum at GtQa (Fig. 8.6. Genisto tinctoriae-Quercetum at Kis Vasbánya hegy) the scale of the real pattern that is the locality of the variance peak is out of the quadrat, so it appears at a bigger scale. The pattern of Polytrichum piliferumis close to the random pattern (see Fig. 7a) in the quadrat of Genisto pilosae-Quercetum community considering the locality of the variance peaks and the value of the variance maximum as well (Fig. 8.2). The value of the variance maximum was relatively high in the case of Dicranum scoparium in both habitats (Fig.8.3 Dicrscop DF: Vmax=5.33, Fig.8.7 Dicrscop GtQb Vmax=2.48), Polytrichum piliferum in Genisto tinctoriae-Quercetum (Fig.8.2 Polpil GtQa Vmax= 10.48) and Polytrichum formosum in Genisto tinctoriae-Quercetum at Borzlyuktető (Fig.8.8 PolforGtQb Vmax=4.5).
Discussion
The bryophyte layer of some types of temperate forest communities may have a greater production than the herb layer living on the soil under the canopy shade patches. If the effect of limiting factors for the distribution of bryophyte species decrease, such as litter production, shading by shrubs, trees and herbs (Longton 1995) the abundance and the production of bryophytes may increase on the soil surface. The extended bryophyte cover consisting of species representing different life forms, life strategy types on the rocky soil surface have dynamic influence for it trapping inorganic and organic materials from the wind, producing organic matter, moderating the extreme temperature changes on the soil. Bryophytes can interact in these forest communities directly with invertebrates providing habitats for them (Varga 1990). The scale and intensity of the spatial pattern of the bryophyte species can be examined in fine scale spatial pattern analysis using quadrat blocks but considering the results it is worth to choose bigger blocks (e.g. 1*1 m in Genisto pilosae-Quercetum) as well to detect the scale of pattern more effectively. In the case of the species Ceratodon purpureus, Polytrichum piliferum in Genisto pilosae-Quercetum community the scale of the real spatial pattern was bigger than the size of the quadrat block. If we examine the life forms of these species and the low maxi- mum variance value of their variograms (Table 1. Fig. 8.) a relationship can be found between the life forms of the occuring bryophyte species and the value of the variance peaks of their variograms using Four-Term Local Quadrat Variance (4TLQV) method. The bryophyte species having small turf life forms (Table 1.) can be characterized with slow variance changes in spatial pattern at a growing scale if the soil surface is homogenous (there are not big rocks on the surface).
The results of 4TLQV analysis can be well compared to other works and it will be used to compare the results of the same quadrat including the spatial pattern of the same bryophyte species in several years to see temporal changes as well.
The results of the spatial pattern analysis show that in the open, acidofil forest communities like Genisto pilosae-Quercetum the spatial pattern of the dominant bryophytes with generative colonist strategy type and short turf life form such as Polytrichum piliferum (see Table 1) is similar with the random spatial pattern and the scale of pattern is out of the quadrat size. It is probably determined by the spatial pattern of the surface and the light mosaic of the open canopy structure (Pénzes Kónya 2003). The dominancy of Polytrichumspecies may have relation with the special lamellae of leaves in the wich decrease not only the water deficiency of the plant but the CO2 loss as well which is an important factor of the photosynthetic response of bryophytes (Marschall &
Proctor 2004). Ceratodon purpureus is known as a colonist species on nitrate defficient soils (Seppelt & Hancock 1991). The real patchy pattern can be recognized within the given quadrat size at the bryophytes of generative and
vegetative perennial strategy type and having tall turf and cushion growth form.
Dicranum species andLeucobryum juniperoideumare succesful dominant speci- es occuring with high frequency or cover values in the acidofil forest communities which have more closed canopy structure (Genisto tinctoriae- Quercetum and Deschampsio-Fagetum communities). The similarity of the two genus can be found not only in their similar spatial pattern, growth form, strategy type and dominant character but in their vegetative reproductive features: the species of both genera have „deciduous leaves” which are asexual vegetative shoots (Newton & Mishler 1994).
The spatial structure of the bryophyte vegetation living on the soil is an important character of the acidofil forest community and the fine scale pattern of the dominant species can be examined with two dimensional spatial pattern analysis. The results of it can be compared with the spatial data of other species or other areas and the spatio-temporal changes can be detected with the results of the quadrat variance methods such as 4TLQV analysis as well.
Acknowledgements
I would like to thank S. Orbán for his field observations and his advices in determining the species, Mrs Erzsébet Nagy for making field work together with me, the collegues in the Department of Botany of Eszterházy College for offering useful publications.
Bibliography
ALPERT, P. 1985: Distribution quantifyed by microtopography in an assemblage of saxicolous mosses. Vegetatio,64, pp. 131–139.
BORHIDI, A. 2003: Plant Communities in Hungary(In Hungarian) Akadémiai Kiadó, Budapest. 610 p
BOROS, Á. 1968: Bryogeographie und Bryoflora Ungarns. Akadémiai Kiadó, Buda- pest.
CARPENTER, S. R. & CHANEY, J. E. 1983: Scale of spatial pattern: four methods compared. Vegetatio 53, 153–160.
DALE, M. R. T., DIXON, M., FORTIN, M. J., LEGENDRE, P., MYERS, D. E. AND ROSENBERG, M. S. 2002: Conceptual and mathematical relationships among methods for spatial analysis. Ecography25: 558–577.
DALE, M. R. T. 1999: Spatial Pattern Analysis in Plant Ecology. Cambridge University Press, 325 pp
DOBOLYI, K., SZABÓ, L., SZERDAHELYI, T.,SZUJKÓ-LACZA, J. 1981: Data to the Genisto-pilosae Quercetumand the flora of the Bükk mountains.Studia Bot.
Hung. 15: 77–90.
DURING, H. J. 1979: Life Strategies of Bryophytes: a preliminary review. Lindbergia5:
2–18.
DURING, H. J., LLORET, F. 1996: Permanent grid studies in bryophyte communities.
J. Hattori Bot. Lab.No. 79. pp 1–45.
DURING, H. J. 1990: Clonal Growth Patterns Among Bryophytes. Clonal Growth in Plants: regulation and function, pp. 153–176.
DURING, H. J. 1992: Ecological classification of bryophytes and lichens. In: Bates, J.W.& Farmer, A. M. (eds.) (1992): Bryophytes and lichens in a changing environment.pp 1–31. Clarendon Press, Oxford, 404 pp
KOVÁCS, M. 1975: Beziehung zwischen Vegetation und Boden. Die Vegetation Ungarischen Landschaften. Band 6. Akadémiai Kiadó, Budapest.
KUN, A. 2000: Összehasonlító vizsgálatok a hárshegyi homokkő növénytakaróján.
Comparative examinations on the vegetation of the sandstone areas on Hárshegy (in Hungarian) Tilia Vol. IX.pp. 60–127.
KÜRSCHNER, H. 1999: Life strategies of epiphytic bryophytes in Mediterranean Pinus woodlands and Platanus orientalis alluvial forests of Turkey. Cryptogamie, Bryologie, 20 (1): 17–33.
LESS, N., HORVÁTH, F., LENDVAI, G. & MATUS, G. 1991:A Hór-völgy környéké- nek (Déli-Bükk) vegetációja (in Hungarian). The Vegetation of the surroundings of Hór valley (Southern Bükk mts.)Bot. Közlem. 76. 1–2: 21–28.
LIEBHOLD, A. M., GUREVITCH, J. 2002: Integrating the statistical analysis of spatial data in ecology. Ecography25: 553–557.
LONGTON, R. E.(1992): The role of bryophytes and lichens in terrestrial ecosystems. In Bates, W., J.(edited): Bryophytes and Lichens in a Changing Environment.
Clarendon Press. Oxford.
LONGTON, R. E. & MILES, C. J. 1982: Studies on the reproductive biology of mosses.
J. Hattori Bot. Lab. 52: 219–240.
MARSCHALL, M. & PROCTOR, M. C. F. 2004: Are Bryophytes Shade Plants?
Photosynthetic Light Responses and Proportions of Chlorophyll a, Chlorophyll b and Total Carotenoids. Annals of Botany94: 593–603.
NEWTON, A. E., MISHLER, B. D. 1994: Asexual reproduction in mosses. J. Hattori Bot. Lab. No. 76: 127–145.
ÓDOR, P. & STANDOVÁR, T. 2001: Richness of bryophyte vegetation in near-natural and managed beech stands: the effects of management-induced differences in dead wood. Ecological Bulletins49: 219–229.
ÓDOR, P., STANDOVÁR, T. 2002: Substrate specifity and community structure of bryophyte vegetation in a near-natural montane beech forest. Community Ecology 3 (1): 39–49. Akadémiai Kiadó, Budapest.
OECHEL, W. C., VAN CLEVE, K. 1986: The role of bryophytes in nutrient cycling in the taiga. In Forest Ecosystems in the Alaskan taiga (eds K. Van Cleve, F. S.
Chapin III, P. W. Flanagan, L. A.Viereck and C. T. Dyrness) pp. 121–37. Sprin- ger, New York.
ORBÁN, S., VAJDA, L. 1983:Magyarország mohaflórájának kézikönyve. Manual of the bryophytes of Hungary(in Hungarian) Akadémiai Kiadó, Budapest
ORBÁN, S1984:A magyarországi mohák stratégiái és T, W, R értékei. Life strategies and T W R values of Hungarian bryophytes (in Hungarian) Acta Acad. Paed.
Agriensis17: 755–765.
ORBÁN, S. 1989: Analysis of some plant communities based on bryophyte layer. Acta Acad. Paed. Agriensis19/9: 197–209.
ORBÁN, S. 2002: A löszfalak moháinak életstratégiái. The life strategies of the bryophytes living on loess walls (in Hungarian) Salamon Albert Éva (szerk.) 2002. Magyar botanikai kutatások az ezredfordulón.Hungarian botanical studies at the end of the 20th century(in Hungarian) Tanulmányok Borhidi Attila 70.
születésnapja tiszteletére. pp 581–587.
PÉNZESNÉ KÓNYA, E., ORBÁN S. 2000:A Bükk-hegység radiolarit alapkőzetű terü- leteinek mohaflórája II. The Bryophyte flora of the areas of the Bükk mountains on radiolarian rocks Kitaibelia Vol. 5 No. 1. pp. 125–130.
PÉNZES KÓNYA, E. 2003: The Effect of Animal Disturbance on the Spatial Pattern and Dynamics of Leucobryum juniperoideum (Brid.) C. Muell. Acta Acad. Paed.
Agriensis, Sectio Biologiae XXIV. pp. 201–213.
PÉNZES-KÓNYA, E. 2004: Comparative Analysis of the Bryophyte Vegetation in Acidophilous Forest Communities in the Bükk Mts (NE Hungary). Acta Bot.
Hung. 46 (3–4), pp 373–384
PÓCS, T. 1967: Néhány adat hazánk mohaflórájához. A few records to the Hungarian bryoflora (in Hungarian) Acta Acad. Paed. Agriensis, n. 5,419–421.
PROCTOR, M. C. F. 2000: Physiological Ecology In: Shaw, A.J.& Goffinet, B. (eds):
Bryophyte Biologypp 225–247. Cambridge University Press
ROSENBERG, M. S. 2001. PASSAGE. Pattern Analysis, Spatial Statistics and Geographic Exegesis. Version 3.0 Department of Biology, Arizona State Univer- sity, Tempe, AZ.
SEPPELT, R. D., HANCOCK, R. J. 1991: Growth responses of Ceratodon purpureusin culture. J. Hattori Bot. Lab. No. 69: 133–145.
SIMON, T. 1977: Vegetationsunterschungen in Zempléner Gebirge. Die Vegetation Ungarischen Landschaften. Band 7. Akadémiai Kiadó, Budapest.
SMITH, A. J. E. 1978:The Moss Flora of Britain and Ireland. Cambridge University Press.
SÖDERSTRÖM, L., HERBEN, T. 1997: Dynamics of bryophyte metapopulations. Adv.
Bryol. 6. : 205–240.
VARGA, J., NAÁR, Z.,DOBOLYI, CS. 2002: Selective feeding of collembolan species Tomocerus longicornis (Müll.) and Orchesella cincta (L.) on moss inhabiting fungi. Pedobiologia46, 526–538.
VOJTKÓ, A. 2001: A Bükk hegység flórája. The flora of the Bükk mountains (in Hungarian) Sorbus Kiadó, Eger 340 pp.
YAMAGUCHI, T. 1993: A Revision of the Genus Leucobryum (Musci) in Asia. J.
Hattori Bot. Lab. No. 73: 1–123.
GpQ- Genisto-pilosae Quercetum (Kövesdi kilátó) GtQa-Genisto tinctoriae-Quercetum (Kis Vasbánya hegy) GtQb- Genisto tinctoriae-Quercetum (Borzlyuktető) DF- Deschampsio-Fagetum (Csák-pilis)
Fig 1. The localities of the sampling areas in the Bükk mountainsin Hungary
c3a1b1 c3 c3 c3a1 a2c2f1 a2f2 k1f1c1b1 b2a1c1
c2a1f1 c1f1a1 c3a1 c3a1 c2a2 a3f1 a2k1f1 k2f1
c3f1 c3k1 c3f1k1 c3f1 c1 a2b2 a2c1 a1c1f1
a2c2 a2c2 a2c2 a1c2 c3b1 b1c1 c2 k2f1
a3c1 a3c1 a3c1 a3c1b1 c3f1 c1b2 b2k1 c1k1
c3a1 c3a2 a2c2 a2c2 c2f1 c2f1 c2f1 c2f1
c3 c3 c3 c3 c2a1 c1a1 c2a2 c2a2
c3 c3 c3 c3 c3 a1c3 c3 c3a1
c3k1 c3 c3 c3 c3k1 c3k1f1 c3 c3a1
c3 c3f1 c3 c3k1 c3k1 c3 c3 c3a1
a= Dicranum scoparium, b=Hypnum cupressiforme, c=Polytrichum piliferum, f=
Polytrichum formosum, k= Dicranum polysetum
Fig. 2. One of the sample quadrats with field data. The numbers mean the actual weighted density, the letters mean the name of the bryophyte species. The quadrat blocks
consisted of four such quadrats.
+ - - -
- - + -
- + - -
- - - +
Fig 3. The potential block comparisons for a 4TLQV analysis
Table 1. The occurance, life forms, life strategies and coenological preferencies of the bryophyte species on the soil surface in the examined communities
Sampling area
(Community) GpQ GtQ1 GtQ2 DF Life
form Life strategy Coenological preferency Species
Bazzania
trilobata + + Mat Generative
perennial Pg Vaccinio- Piceetalia Cephaloziella
divaricata + Mat Generative
perennial Pg Querco-Fagetea Ceratodon
purpureus + Short
Turf
Generative
Colonist Cg Corynephoretalia Dicranum
polysetum + + Cushion Vegetative
perennial Pv Pino-Quercetalia Dicranum
scoparium + + + + Cushion Vegetative
perennial Pv,
Pino-Quercetalia
Dicranum
spurium + Cushion Vegetative
perennial Pv Pino-Quercetalia Hypnum
cupressiforme + + + + Weft Vegetative
perennial Pv neutral
Isothecium
myosuroides + + Weft
Perennial shuttle species
with low sexual/asexual
reproduction Pp,
Hypno- Polypodietum
Leucobryum
juniperoideum + Cushion
Vegetative
perennial Pv, Vaccinio- Piceetalia Plagiomnium
cuspidatum + Weft Vegetative
perennial Pv, Querco-Fagetea
Plagiothecium
laetum + Weft
Perennial shuttle species
with low sexual/asexual
reproduction Pp
Pino-Quercetalia
Politrichum
piliferum + + Short
Turf Generative
colonist Cg Corynephoretalia Polytrichum
formosum + + + + Tall
Turf
Generative
perennial Pg Querco-Fagetea Polytrichum
juniperinum + Short
Turf Generative
Colonist Cg Pino-Quercetalia
a Mat
Weft Short turf Tall turf Cushion
b
Pg Pv
Pp Cg
c cosmopolitan Festuco brometea Corynephoretalia Fagetalia
Festucetalia-valesiacae Acerion pseudoplatani Ctenidio-Polypodietum Querco-Fagetea
Vaccinio-piceetalia
Pp= Perennial shuttle species,
Pg = generative perennials Pv=vegetative perennials Cg= generative colonists
Fig.4.The percent distribution the occuring bryophyte species on the basis of the a) growth form, b) life strategy types, c) coenosystematic categories.
a)
Genisto pilosae-Quercetum
0 20 40 60 80 100 120
GpQ11 GpQ12 GpQ13 GpQ14 Gpq21 GpQ22 GpQ23 GpQ24 GpQ31 GpQ32 GpQ33 GpQ34
quadrats
DISCOP
CERPUR
POLPIL
b)
Deschampsio-Fagetum
0 10 20 30 40 50 60 70 80 90 100
DF1199 DF1299 DF1399 DF1499 DF2199 DF2299 DF2399 DF2499 DF3199 DF3299 DF3399 DF3499
quadrats
HYPCUP POLFOR DISCOP LEJUN
c)
Genisto tinctoriae-Quercetum a (Kis Vasbánya-hegy)
0 20 40 60 80 100 120
GtQa1/11 GtQa1/12 GtQa1/13 GtQa1/14 GtQa1/21 GtQa1/22 GtQa1/23 GtQa1/24 GtQa1/31 GtQa1/32 GtQa1/33 GtQa1/34 quadrats
Dicrspur Hypcup Polpil Polfor Dicrpolyset Plagiotleat Cerpur
d)
Genisto tinctoriae- Quercetum b (Borzlyuktető)
0 20 40 60 80 100 120
GtQb/11 GtQb/12 GtQb/13 GtQb/14 GtQb/21 GtQb/22 GtQb/23 GtQb/24
quadrats
HYPCUP POLFOR PLAG.
BAZTRI ABIAB DISCOP CEDIV
Fig. 5. The absolute frequency spectra of the bryophyte species occuring in the quadrats of the different communities: a) Genisto pilosae-Quercetum (GpQ), b) Deschampsio- Fagetum (DF), c) Genisto tinctoriae-Quercetum a at Borzlyuktető (GtQa), d) Genisto
tinctoriae-Quecetum b at Kis Vasbánya-hegy (GtQb).
Cerpur = Ceratodon purpureus, Polpil = Polytrichum piliferum, Polfor = Polytrichum formosum, Dicrscop = Dicranum scoparium, Leucjun = Leucobryum juniperoideum, Hypnum = Hypnum cupressiforme Dicrspur = Dicranum spurium, Dicrpolyset = Dicranum polysetum, Plagiotlet = Plagiothecium laetum, Cediv = Cephaloziella divaricata, Baztri = Bazzania trilobata, Abiab= Abietinella abietina
Figure 6. The results of a 4TLQV analysis in the case of a random pattern simulation.
The plot (Fig. 6.a) represents the spatial structure of the distribution pattern where the cells with the highest density are white and the empty cells (with density value 0) are black. There are no patches in the random pattern. The variogram (Fig. 6 b) of 4TLQV analysis refers to no patches : the locality of the maximum value is at the smallest scale.
Fig. 7. The plot maps made on the basis of the quadrat data of the analysed bryophyte species in different communities. The lightest grid cells (white) mean the greatest density
of the species. 1. Ceratodon purpureus in Genisto pilosae-Quercetum 2.Polytrichum piliferum in Genisto pilosae-Quercetum 3.Dicranum scoparium in Deschampsio-
Fagetum 4. Hypnum cupressiforme Genistotinctoriae-Quercetum (GtQa) 5.
Polytrichum piliferum inGenisto tinctoriae-Quercetum (GtQa) 6.Leucobryum juniperoideumin Deschampsio-Fagetum 7.Dicranum scopariumin Genisto tinctoriae- Quercetum (GtQb) 8. Polytrichum formosumin Genisto tinctoriae-Quercetum 9. Plot of
a random pattern simulation
Fig. 8. The 4TLQV variograms of the dominant bryophyte species in the different communities. On the horizontal axis the scale is represented while on the vertical one the variance can be seen. The locality of the variance peak refers to the scale of spatial
pattern. The names of the bryophytes are:
Cerpur = Ceratodon purpureus, Polpil = Polytrichum piliferum, Polfor = Polytrichum formosum, Dicrscop = Dicranum scoparium, Leucjun = Leucobryum juniperoideum, Hypnum = Hypnum cupressiforme