Research Article 1
Three years of vegetation development worth 30 years of secondary
2
succession in urban-industrial grassland restoration
3
Running head: Urban-industrial grassland restoration 4
5
Anna Kövendi-Jakó1,2, Melinda Halassy1, Anikó Csecserits1,3, Karl Hülber4,5, Katalin 6
Szitár1,3, Thomas Wrbka4, Katalin Török1 7
1 Institute of Ecology and Botany, MTA Centre for Ecological Research, Alkotmány u. 2-4., 8
Vácrátót, 2163 HU 9
2 Department Plant Taxonomy, Ecology, and Theoretical Biology, Eötvös Loránd University, 10
Budapest, Pázmány Péter stny. 1/c, Budapest, 1117 HU 11
3 GINOP Sustainable Ecosystems Group, MTA Centre for Ecological Research, Klebelsberg 12
Kuno u. 3., Tihany, 8237 HU 13
4 Division of Conservation Biology, Vegetation Ecology and Landscape Ecology, Department 14
of Botany and Biodiversity Research, University of Vienna, Rennweg 14., Vienna, 1030 AT 15
5 Vienna Institute for Nature Conservation & Analyses, Giessergasse 6/7, Vienna, 1090 AT 16
Correspondence:
17
1. Anna Kövendi-Jakó, Institute of Ecology and Botany, MTA Centre for Ecological 18
Research, Alkotmány u. 2-4., Vácrátót, 2163 HU 19
Email: kovendi-jako.anna@okologia.mta.hu 20
Funding:
21
The study was funded by the LEGO group and by the National Science Foundation of 22
Hungary (NKFI-OTKA FK127996). AKJ was supported by the ÚNKP-18-3 New National 23
Excellence Program of the Ministry of Human Capacities. ACs was funded by the 24
Hungarian Academy of Sciences (MTA PD-036/2015, PD-019/2016, PD009/2017) and by the 25
GINOP-2.3.2-15-2016-00019 grant. KSz was supported by the GINOP-2.3.2-15-2016-00019 26
grant.
27
Abstract
28
Questions:
29
The restoration of unused urban-industrial areas has largely been neglected despite their great 30
potential for nature conservation purposes. We applied three plant introduction treatments at a 31
highly degraded industrial area to test whether plant material introduced to industrial areas 32
initiate plant assemblages similar to the composition of reference grasslands. We specifically 33
asked (1) Does restoration differ from primary and secondary reference sites in terms of 34
overall species richness and cover three years after plant material introduction? (2) What is 35
the sociability of species of the resulting vegetation at different treatments?
36
Location:
37
Restoration and reference grassland sites with acidic sandy soil in the temperate region of EU, 38
NE Hungary, Nyírség.
39
Methods:
40
As restoration techniques we (i) directly seeded a single dominant species, (ii) applied a 41
commercial seed mixture, and (iii) transferred hay. We compared species composition, cover, 42
species richness and sociability of species of restoration treatments to reference grasslands.
43
Inventories of vascular plant species were made at five randomly placed 2 m x 2 m sampling 44
units per plot.
45
Results:
46
The species composition of seeded restoration plots (by single dominant species and a 47
commercial seed mixture) became similar to reference grasslands. Both type of seeding 48
resulted in similar cover, whereas seeding of commercial seed mixture and hay transfer 49
resulted in similar species richness to reference grasslands. The cover of natural constituents 50
of seeded plots also reached that of reference grasslands, while hay addition resulted in lower 51
cover and higher number of natural constituents than other methods.
52
Conclusions:
53
The introduction of propagules in degraded industrial areas can provide after three years 54
similar communities to those of secondary reference grasslands of 30 years of age. We 55
conclude that investing in the restoration of native grasslands at unused urban-industrial sites 56
can be a great opportunity to enhance biodiversity.
57
Keywords
58
dry grassland, industrial area, hay transfer, multiple reference sites, old-fields, seed sowing, 59
species richness, sociability of species, urban-industrial restoration, vegetation cover 60
61
Introduction
62
In recent decades semi-natural grasslands have become strongly reduced in area and diversity 63
due to the intensification of agricultural production (Bond, 2016; Török, Janišová,Kuzemko, 64
Rūsiņa, and Stevanović, 2018b) and human land use, including urban, industrial and 65
infrastructure expansion that is foreseen to accelerate (Maes et al. 2015). To compensate for 66
this loss of natural areas, restoration must be up-scaled and extended from agricultural and 67
semi-natural areas to urban and industrial sites (Aronson & Alexander, 2013; Hostetler, Allen, 68
and Meurk, 2011; Klaus, 2013; Standish, Hobbs, and Miller, 2013). Maes et al. (2015) 69
recommends every increase of artificial land to be compensated by a 2.2-fold increase of 70
green infrastructure to maintain ecosystem services. Thus, any kind of unused land or vacant 71
lot should be considered for extending the network of green infrastructure that beside the 72
enhancement of biodiversity (Standish et al. 2013; Deák, Hüse, & Tóthmérész, 2016; Hüse, 73
Szabó, Deák, & Tóthmérész, 2016; Anderson & Minor 2017) could greatly increase human 74
wellbeing (Tzoulas et al. 2007).
75
Spontaneous secondary succession can be the most cost-effective way of restoration (Prach &
76
Hobbs, 2008; Kiehl, Kirmer, Donath, Rasran, and Hölzel, 2010; Török, Vida, Deák, Lengyel, 77
and Tóthmérész, 2011) but is not appropriate in cases of missing dispersal agents and 78
processes, depletion of soil seed banks and lack of propagule sources (Bakker & Berendse, 79
1999; Hedberg & Kotowski, 2010). In such cases, artificial introduction of seeds is required 80
(Kiehl et al. 2010; Török et al. 2011). Restoration efforts usually apply seeds of indigenous 81
species aiming to restore (historical) native assemblages as far as it is feasible (Shackelford et 82
al. 2013). Several methods of seed introduction can be used to fit the particular restoration 83
need and abiotic environment. Commercial, low diversity seed mixtures (commonly 84
comprising generalist species) efficiently reduce erosion due to the rapid establishment of a 85
dense vegetation cover compared to hay transfer (Török et al. 2011), but the long-term 86
persistence and ecosystem functioning might be compromised (Kettenring, Mercer, Reinhardt 87
Adams, and Hines, 2014). Collection of local propagules is a viable alternative; however, 88
local seed sourcing is usually difficult in large quantities (Török et al. 2018a) and might lead 89
to poor restoration outcomes due to inbreeding (Broadhurst et al. 2008). The availability of 90
seeds of suitable species in sufficient quantities is a major constraint in restoration (Havens et 91
al. 2015), so the application of a single species can be an option (Piper, Schmidt, and Janzen, 92
2007; Oliveira, Clemente, Nunes, and Correia, 2014). Alternatively, transferring plant 93
material in the form of hay or raked litter can result in higher species richness than seeding 94
(Hedberg & Kotowski, 2010; Kiehl et al. 2010; Török et al. 2011). Since only few papers 95
report on the greening of industrial areas aside from mining sites (Török et al. 2018a), we 96
need further guidance on how to improve the natural state of urban-industrial areas that can 97
contribute to compensate for land degradation.
98
The aim of the present study is to test the success of commercial seed mixtures, seeds of a 99
single dominant species and the transfer of hay in restoring a severely disturbed industrial 100
area. We compare the resulting vegetation three years after restoration interventions to those 101
of primary (semi-natural grasslands) and secondary (old-fields) reference grasslands We 102
tested whether plant material introduced to industrial areas initiate plant assemblages similar 103
to the composition of reference grasslands. We specifically asked (1) Does restoration differ 104
from primary and secondary reference sites in terms of overall species richness and cover 105
three years after plant material introduction? (2) What is the sociability of species of the 106
resulting vegetation at different treatments?
107
Methods
108
Study area 109
The study area is located in the Nyírség region (East Hungary, Appendix S1). Annual mean 110
precipitation is 550-600 mm and annual average temperature is 9.8 ºC. The historic landscape 111
was characterised by acidic inland sand dunes (with dune height of 5-10 m) covered with 112
sandy grasslands and dry oak steppe forests on dunes and marshlands in depressions (Boros, 113
1929; Soó, 1939; Papp & Dudás, 1989). These natural vegetation types have been mostly 114
transformed to arable lands, orchards, vineyards and tree plantations, resulting in the loss of 115
natural habitats and the expansion of invasive species (e.g. Asclepias syriaca, Robinia 116
pseudo-acacia, Botta-Dukát, 2008).
117
Restoration site 118
The restoration site (9.8 ha) is located around the factory buildings of the LEGO Group at 119
Nyíregyháza, N-E Hungary in the region of Nyírség (lat 47° 57'N; long 21° 39'E). The factory 120
was established at a former orchard and small farmlands, causing the complete loss of 121
vegetation and damage to the original soil surface by flattening the ground. The remaining 122
soil is compacted sand with low humus, calcium, and nutrient content and very low water 123
holding capacity (Appendix S2). No sandy grasslands occur within a three km radius around 124
the factory (Török et al. 2018a).
125
Reference sites 126
The target sandy grasslands have a high natural value and are protected by the Habitat 127
Directive of the European Union (priority habitat code 6260; Romão, 1996). We selected both 128
open (Festuco vaginatae – Corynephoretum Aszód 1935) and closed sandy grasslands 129
(Potentillo arenariae – Festucetum pseudovinae Soó 1940, Pulsatillo hungaricae - 130
Festucetum rupicolae Borhidi 1996). Open sand grasslands develop on low humus content, 131
loose sandy soils at dune tops with a maximum cover of 75% (Bölöni, Botta‐Dukát, Illyés, 132
and Molnár, 2011). Closed sand grasslands develop on humus-rich sandy soils mostly at the 133
foothills of surrounding mountains or at the lowland. The minimum plant cover is 50 % 134
(Bölöni et al. 2011). The two types have originally formed a mosaic in the landscape, 135
depending on relief, exposure and soil quality. However, both types have very fragmented 136
occurrences in the study area presently. We further refer to grasslands selected at permanent 137
grassland sites as primary reference.
138
Secondary grasslands developed on old-fields by spontaneous succession over more than 30 139
years were used as secondary reference sites (hereafter referred to as secondary reference).
140
These sites created by land abandonment have the potential for the recovery of native 141
temperate grasslands by old-field succession (Cramer, Hobbs, and Standish, 2008). In 142
Central-Eastern Europe, spontaneous succession follows the pathway towards natural 143
vegetation with a rapid development of low diversity, but weedy grasslands in the first ten 144
years (Molnár & Botta-Dukát, 1998, Csecserits & Rédei, 2001; Halassy, 2001; Ruprecht, 145
2006; Csecserits et al. 2011; Albert et al. 2014).
146
Five sites representing the target vegetation were selected as reference for comparison with 147
the restoration site. Primary reference sites representing extensively used, ancient grasslands 148
were selected at three locations (Bátorliget 23 ha, Magy 6.5 ha, Martinka 185 ha). Two of 149
them (Bátorligeti legelő nature conservation area, Martinkai legelő at Hajdúsámson) are in 150
protected areas. The soil of these sites is slightly acidic or neutral sandy soil with low humus, 151
calcium, and nutrient content, and higher nitrogen content (Appendix S2). Secondary 152
reference sites were selected at two locations and included vineyards abandoned in the 1960s 153
(Geszteréd) and in the end of 1980s (Ófehértó). Their soil is slightly acidic adobe sandy with 154
low humus, calcium and nutrient content, and very low water holding capacity (Appendix 155
S2). All reference sites are grazed with sheep or cattle. Both open and closed grasslands were 156
studied at each of the five locations, referred to as primary open (POR), primary closed (PCR) 157
Restoration treatments 159
Around the factory we have seven restoration plots with different plant material introduction 160
methods according to the availability of species at the time of release from construction 161
(Török et al. 2018a). We present here in detail the results of the plant material introduction of 162
2014 at four restoration plots ranging from 1 to 4.5 ha in size (Fig. 1). Since plots of land 163
became available too late in 2013 for applying the plant material introduction, lucerne 164
(Medicago sativa) and rye (Secale cereale) were seeded (20 kg/ha) to reduce erosion by wind 165
and to avoid a vegetation dominated by weeds and invasive species (mainly ragweed, 166
Ambrosia artemisiifolia). All restoration plots were treated by deep soil loosening, ploughing, 167
and seedbed preparation before applying one of the following restoration treatments: 1) 168
introduction of commercial seeds (abbreviation: COM) of Festuca pseudovina as matrix grass 169
species (30 kg/ha) and 26 native generalist forb species (15 kg/ha) produced in the Hungarian 170
Lowland (Sep 2014). 2) Seeding of Festuca rupicola (60 kg/ha) as a single dominant 171
generalist (abbreviation: DOM) harvested by reaping machine in the Hortobágy National Park 172
(Sep 2014). Mulching by cut autumn biomass was applied on seeded plots (COM, DOM) 173
shortly after seeding to control erosion by wind and for weed suppression (9 - 10 bales/ha). 3) 174
Transfer of dried hay (two plots; abbreviation: HAY) obtained from donor sites within a 60 175
km distance from the factory. Early summer hay with a high content of Festuca spp. seeds 176
(23.5 - 26 bales/ha; one bale weighted about 250 kg) was collected in two sites in Jun and in 177
early Jul 2014, which was manually distributed to cover the whole plots by about 5 cm (Jul 178
2014). Bales from a late harvest (from Aug 2014) containing mainly forb seeds (3.5 - 5 179
bales/ha) were collected from a third site, this hay was distributed on top at a 3 cm thickness 180
(Aug 2014). Seed content of hay and mulch was estimated by germination experiment of hay 181
and mulch samples (data not shown). The restoration plots were mown twice yearly since 182
seeding. For more details on the applied restoration treatments see Table 1.
183
Data recording and analyses 184
The success of restoration treatments was assessed by comparing the species composition at 185
restoration plots to those of primary and secondary reference sites. Cover (in percentage) of 186
each vascular plant species was visually estimated in five randomly placed sampling units (2 187
m x 2 m) at each restoration plots (yearly), at the three primary reference sites (in June either 188
in 2015 or in 2016), and at the two secondary reference sites (in June 2017). In this paper we 189
use cover data of restoration plots only from June 2017, i.e. three years after applying the 190
restoration treatments, resulting in n=20 restoration and n=50 reference sampling units 191
altogether. Due to uncertainties in the identification of young vegetative plants, Festuca 192
rupicola, F. pseudovina, and F. valesiaca were merged as Festuca spp. for all analyses. Total 193
cover (i.e. sum of individual species´ cover) and species richness (i.e. number of species) 194
were calculated for each of the 70 sampling units. Nomenclature follows Király (2009).
195
We examined the sociability of species in the restoration plots compared to reference 196
grasslands by merging the social behaviour types defined by Borhidi (1995), based on Grime 197
(1979), in three major categories:1) natural constituents (NC - comprising specialists, 198
competitors, generalists and natural pioneers); 2) disturbance tolerant species (DT, as in 199
Borhidi, 1995); and 3) weeds (W - including introduced cultivated plants, ruderal competitors 200
and adventive competitors). The number of species and cover values belonging to each 201
category of sociability per sampling unit was used for analyses. For more details on the 202
categories of studied species see Appendix S3.
203
Composition of sampling units was compared using non-metric multidimensional scaling 204
(NMDS) and Analysis of Similarity (ANOSIM, Clarke, 1993) based on Bray-Curtis 205
dissimilarity and Sørensen index. Number of dimensions (k) was set to two. For data 206
processing the package “vegan” (Oksanen et al. 2017) of the R 3.3.1 statistical environment 207
(R Core Team, 2016) was applied.
208
We used (generalized) linear mixed effects models (GLMM) implemented in the package 209
“afex” (Singmann, Bolker, Westfall, and Aust, 2018) to compare total cover and species 210
richness among restoration plots (COM, DOM, HAY) and reference sites (PCR, POR, SCR, 211
SOR), used as seven levels of study sites factors in all models. Models were fit to data 212
comprising species richness and total cover of all species of a sampling unit and separately to 213
the three species groups based on sociability types. To consider potential independence of the 214
data of the five sampling units at a site we allowed for a random intercept for each site in each 215
model. For total cover and species richness a Gaussian and Poisson distribution were 216
assumed. For the social behaviour type DT total cover data was transformed by square roots 217
to fulfil the assumptions of normality and homoscedasticity. In case of total cover of the 218
social behaviour type W the model residuals showed heterogeneity of variance for cover data 219
even after square root transformation and, therefore linear mixed-effects (LME) models (Zuur 220
et al. 2009) were applied by using the “nlme” package (Pinheiro et al. 2017) with varIdent 221
variance structure which allowed for different residual spread for each level of the sites 222
variable. For post hoc pairwise comparisons Tukey HSD tests were applied by using 223
“multcomp“ package (Hothorn, Bretz, and Westfall, 2008), with p values adjusted by the 224
method of Benjamini and Hochberg (1995).
225
Results
226
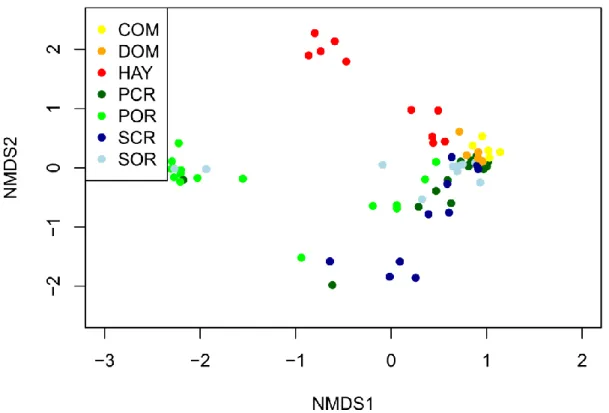
Composition based on multivariate analyses 227
According to the results of NMDS and ANOSIM analysis of cover data, the four reference 228
grassland types proved to have significantly different composition from each other. As for the 229
restoration treatments, the composition of sown restoration plots (COM, DOM) was similar to 230
three reference sites, except for POR (Fig. 2; Table 2). HAY differed significantly from the 231
seeding treatments (COM, DOM) and from all reference sites. Referring to the species 232
composition all study sites differed significantly from each other (except the two secondary 233
reference grasslands (SCR-SOR) based on the results of NMDS and ANOSIM analysis for 234
presence/absence data (Appendix S4; S5).
235
Total cover and species richness 236
Study sites significantly differed (chisq= 51.33, df= 6, p< 0.001) based on the GLMM 237
analysis of total cover. Tukey tests proved significant differences between closed and open 238
types of reference sites (PCR-POR: z= -4.938, p< 0.001; PCR-SOR: z= -5.654, p< 0.001;
239
SCR-POR: z= 3.554, p= 0.007; SCR-SOR: z= -4.374, p< 0.001, Fig. 3a). Concerning 240
restoration treatments, gradually lower cover from the commercial seeding, dominant species 241
seeding, and hay addition was detected, however, not significant. The total vegetation cover 242
of restoration sampling units was similar to that of reference sites, except for significant 243
differences found between COM and SOR (z= -3.624, p= 0.005) and between HAY and PCR 244
(z = -3.930, p= 0.002). Species richness differed significantly among study sites (chisq=
245
14.777, df= 6, p= 0.022) based on the GLMM analysis. According to Tukey test, all 246
restoration treatments resulted in similar species richness to one another, and to the reference 247
sites, except DOM with the lowest species richness per sampling unit, which was significantly 248
different from both closed reference sites (PCR-DOM: z= -3.139, p= 0.027; SCR-DOM: z=
249
2.951, p= 0.047) (Fig. 3b).
250
Sociability of species 251
According to the results of GLMM the total cover of NC species was significantly different 252
among the study sites (chisq= 44.282, df= 6, p< 0.001). Based on Tukey HSD tests, there was 253
a significant difference between closed and open types of reference sites in case of primary 254
grasslands (PCR-POR: z= -3.780, p= 0.003; PCR-SOR: z= -3.243, p= 0.019). Restoration 255
treatments resulted in similar cover of NC compared to reference sites, except for HAY that 256
had significantly lower total cover of NC than closed reference sites (HAY-PCR: z= -6.153, 257
p< 0.001; HAY-SCR: z= -4.394, p< 0.001). HAY and COM (z= -3.602, p= 0.006) as well as 258
HAY and DOM (z= -3.038, p= 0.037) resulted in significant difference in total cover of NC 259
(Fig. 4a). Study sites had significant effect on the species richness of NC species (chisq=
260
28.351, df= 6, p< 0.001). Based on Tukey HSD test the reference sites were similar to each 261
other. HAY resulted in a similar species richness to reference sites, whereas COM and DOM 262
resulted in significantly lower number of NC compared to primary open grasslands (COM- 263
POR: z= -3.513, p= 0.008; DOM-POR: z= -4.104, p< 0.001), primary closed (DOM-PCR: z=
264
-3.416, p= 0.01), and secondary closed grasslands (DOM-SCR: z= -3.096, p= 0.029, Fig. 4b).
265
Study sites had significant effect on the cover of DT species (chisq= 23.152, df= 6, p< 0.001) 266
based on GLMM. The reference sites were not significantly different from each other based 267
on cover of DT species according to the Tukey HSD test. Restoration treatments resulted in 268
similar cover of DT compared to reference sites, except for HAY that resulted in significantly 269
higher total cover than open reference sites (POR-HAY: z= 3.653, p= 0.005; SOR-HAY: z=
270
3.746, p= 0.003, Fig. 4c). Study sites had significant effect on the species richness of DT 271
(chisq= 22.327, df= 6, p= 0.001) based on GLMM. According to Tukey HSD test, 272
significantly higher richness of DT was found for PCR, than POR (POR-PCR: z= -4.129, p<
273
0.001). Restoration treatments resulted in similar species richness of DT compared to 274
reference sites, except for HAY that had significantly higher species richness of this group 275
than POR (z= 3.343, p= 0.014, Fig. 4d).
276
Although W species had higher mean total cover in the restored than in the reference 277
grasslands, these differences were not significant based on the LME analyses (chisq= 6.766, 278
df= 6, p= 0.343, Fig. 4e). Concerning species richness of W, the study sites had significant 279
effect (chisq= 28.484, df= 6, p< 0.001). Significant difference of species richness of W was 280
not detected among reference sites and among restoration sites. HAY and COM addition had 281
significantly higher species richness of W than primary reference sites (COM-PCR: z= 3.861, 282
p= 0.002; COM-POR: z= 3.375, p= 0.013; HAY-PCR: z= 3.975, p= 0.001; HAY-POR: z=
283
3.446 p= 0.01, Fig. 4f).
284
Discussion 285
286
Overall cover and species richness 287
Our study proved the difference in total cover of closed versus open grasslands, both in 288
primary and secondary references. Treated samples have intermediate total cover, while only 289
hay additions resulted in lower cover values than primary closed reference. The seeding of 290
commercial seed mixture allowed for a rapid establishment of the seeded species resulting in 291
the highest vegetation cover (85 %) similar as found by Török et al. (2011). Sowing of the 292
dominant grass species resulted more than 70 % average cover by the third growing season, 293
which is in the range reported by other studies applying low-diversity seed mixtures (Török et 294
al. 2010; Vida et al. 2010). This method is more cost-effective than sowing commercial seed 295
mixtures (Török et al. 2011), but at the expense of having less species rich assemblages. Hay 296
transfer, which is often used to start secondary succession (Rasran, Vogt, and Jensen, 2006;
297
Kiehl, Thormann, and Pfadenhauer, 2006), resulted in a slightly lower species cover.
298
Important factors affecting the restoration success when transferring hay are the species 299
composition of donor sites, the timing of the hay collection and hay storage (Rasran et al.
300
2006; Kiehl et al. 2010; Török et al. 2011). In our study, the application of dry, stored hay can 301
be an explanation for lower plant cover, as dry hay usually contains lower amount of viable 302
seeds than that of fresh plant material without storage (Kiehl et al. 2010).
303
Contradicting several studies, the primary and secondary reference grasslands are very similar 304
in species richness (Molnár & Botta-Dukát, 1998; Prach et al. 2016). The colonization of 305
specialist species can take hundreds of years based on these studies, so primary reference 306
grasslands should have higher richness. Fragmented cultivation might have led to specialist 307
species survival at field margins (Hackett & Lawrence 2014) that could contribute to similar 308
species richness at secondary reference sites. Besides, we assume that dispersal by moderate 309
grazing also helps to increase species richness at secondary sites. In the restoration plots 310
species introduction resulted in similar richness to reference grasslands, hay transfer resulting 311
in slightly higher values, however, not significant. The early success of grass species can be 312
contradictory in the long run decreasing the efficiency of further species establishment. Sown 313
grass species (either applied alone or in seed mixtures) can become dominant very fast on the 314
cost of sown or naturally introduced forb species and decrease overall species richness in the 315
long run in restoration sites (Török et al. 2011). The dense cover of dominant species as 316
perennial grasses entails spontaneous colonization of rare species to be very slow even in case 317
of propagule availability in the surroundings (Török et al. 2010; Vida et al. 2010). This 318
limitation for species establishment justifies further interventions including the diversification 319
of the grasslands, like the application of establishment gaps with high-diversity sowing 320
(Valkó et al. 2016). Contrary to several studies (Kiehl et al. 2006; Kiehl et al. 2010; Török et 321
al. 2011), hay transfer resulted in similar species richness to other restoration treatments, 322
which can be explained by the application of dry, stored hay with lower viable seed content.
323
For further restoration projects we suggest using seed-containing hay in combination with 324
single species or seed mixture seeding, preferably from late harvest to increase the number of 325
forbs in the vegetation (Török et al. 2012).
326
Sociability of species 327
The goal in this experiment was to introduce the natural constituents of the target community.
328
Reference sites had higher cover and species richness of natural constituent species. Both 329
types of seedings (dominant species and commercial seed mixture) introduced very few 330
natural constituent species, but these reached a total cover similar to that of the reference 331
grasslands. At the same time, the hay transfer performed slightly higher richness in natural 332
constituents with lower total cover, which supports the idea of using this technique as a 333
complementary treatment to seeding (Török et al. 2012). Despite restoration sites being at an 334
early stage of succession (3rd year), the expected higher cover of disturbance tolerant species 335
and weeds compared to references was not confirmed. The number of weed species was 336
higher with hay transfer and commercial seed mixture than that of primary reference sites.
337
Hay transfer resulted in more open vegetation with gaps, which provide physical space for 338
germination and establishment of weedy species from the soil seed bank. This problem can be 339
detected especially in former agricultural areas, where the original seed bank was replaced by 340
weedy species in the soil seed bank (Bakker & Berendse, 1999). This is less expected in 341
industrial areas, where the seed bank is destroyed, rather than replaced by weedy species. The 342
cover of natural constituents of the restoration plots should be further increased by regular 343
mowing to decrease the total cover and richness of weeds as in other studies (Kiehl et al.
344
2010; Török et al. 2011).
345
Success of restoration 346
We found that the vegetation composition of seeded plots (dominant species and commercial 347
seed mixture) became similar to that of the reference sites based on cover data, except the 348
primary open grassland that also differed from the other reference sites. In contrast, the 349
vegetation composition of the plot with hay addition remained significantly different from all 350
other samples. These results suggest that seeding successfully accelerated succession towards 351
secondary grasslands and primary closed grasslands in three years, compared to secondary 352
reference grasslands developed in more than 30 years. Several investigations (Molnár &
353
Botta-Dukát, 1998; Csecserits & Rédei, 2001; Halassy, 2001; Ruprecht, 2006; Csecserits et 354
al. 2011; Albert et al. 2014) found that spontaneous succession at old-fields in the Pannonian 355
region can develop to semi-natural grasslands on disturbed areas within 10-20 years, however 356
the cover of specialist species remains low (Molnár & Botta-Dukát, 1998; Prach et al. 2016), 357
and the cover of alien species is much higher than in natural grasslands (Csecserits et al.
358
2011; Csecserits et al. 2016). Mitchley, Jongepierova and Fajmon (2012) consider 10-60 359
years insufficient to restore species rich grasslands. Overall, our study supports that this time- 360
consuming succession can be accelerated by seeding.
361
We provide a summary of the advantages and disadvantages of seed introduction methods 362
based on six criteria (Table 3) to help selecting the most appropriate technique in future 363
restoration projects at unused industrial areas or vacant lots. The study demonstrated that 364
there is no single “best method” for restoration; the selection should be based on the particular 365
demand and circumstances (Shackelford et al. 2013; Adams, Hodge, Macgregor, and 366
Sandbrook, 2016). Multiple constraints exist in all restoration projects, but in case of 367
intensively modified landscapes, like urban-industrial sites, efforts needed to handle the 368
constraints can be higher (Arenas, Escudero, Mola, and Casado, 2017). This guidance helps to 369
select appropriate method in future projects, linked to the green infrastructure policy of the 370
European Union (European Commission, 2013). For example, in case of limited seed 371
availability, hay addition can be a good alternative; or if rapid green surface is a priority, 372
commercial seed mixtures are appropriate; low budget can be the reason for single species 373
seeding and still reaching similar target species cover to reference grasslands. It is important 374
however, that the aim to enhance biodiversity should not be compromised (Standish et al.
375
2013). In our case the applied restoration methods significantly contributed to increase 376
species richness, cover and similarity towards reference grasslands, but little significant 377
difference was detected among the restoration treatments.
378
Conclusions 379
The potential of unused urban-industrial areas for enhancing biodiversity is widely ignored in 380
the world, although their restoration has advantages for conservation (Klaus, 2013; Deák et al.
381
2016; Hüse et al. 2016) and green infrastructure development (Hostetler et al. 2011; Deák et 382
al. 2016; Hüse et al. 2016). Instead of creating intensively managed, species poor green areas, 383
these non-built-up lands could be used to restore more self-sustainable, native biodiversity 384
refuges (Török et al. 2018a) that provide additional ecosystem services, like pollination 385
(Kovács-Hostyánszki et al. 2017) or amenity value (Martens, Gutscher, and Bauer, 2011). We 386
tested the role of restoration treatments in enhancing the species cover, richness, naturalness 387
in terms of sociability of species and similarity to reference grassland composition and 388
provide guidance on how to choose best method for a given situation. The success of our 389
grassland restoration project is encouraging regarding the difficulties of urban-industrial area 390
restoration, like dispersal (limitation of available propagule and dispersal agents), biotic 391
(competition) and abiotic (soil, microclimate) constraints (Klaus, 2013). In three years the 392
restored vegetation reached a state similar to that of old-fields by spontaneous succession of 393
more than 30 years. We conclude that the re-creation of native grasslands by assisted 394
introduction of species at industrial areas provides a great opportunity to enhance biodiversity 395
in a relatively short period of time and thus contribute to the development of green 396
infrastructure in Europe (Standish et al. 2013; Liquete et al. 2015).
397
Acknowledgments 398
The authors thank the LEGO group for providing the infrastructure and the Hortobágy 399
National Park Directorate for providing plant material for restoration. We thank Csaba 400
Szigetvári, who helped to find appropriate reference sites Gardening work has been carried 401
out by the Deep Forest Ltd. We thank students and Barbara Lhotsky for help in field work.
402
Author contributions 403
ACs, AKJ, KT, MH conceived and designed the study; ACs, AKJ, MH, KT did collections 404
and other field work; AKJ, KSz, KH made statistical analyses; AKJ, MH, KH, KSz, KT, TW 405
wrote and edited the paper. All authors contributed critically to the drafts and gave final 406
approval for publication.
407
Data accessibility 408
Data are available from ZENODO: https://zenodo.org/record/1227269 (Kövendi-Jakó, 2018) 409
References 410
Adams, W. M., Hodge, I. D., Macgregor, N.A., & Sandbrook, L. (2016). Creating restoration 411
landscapes: partnerships in large-scale conservation in the UK. Ecology and Society, 21, 1.
412
https://doi.org/10.5751/ES-08498-210301 413
Albert, Á. J., Kelemen, A., Valkó, O., Miglécz, T., Csecserits, A., Rédei, T., … Török, P.
414
(2014). Secondary succession in sandy old‐fields: a promising example of spontaneous 415
grassland recovery. Applied Vegetation Science, 17, 214-
416
224.https://doi.org/10.1111/avsc.12068 417
Anderson, E.C., & Minor, E.S. (2017). Vacant lots: An underexplored resource for ecological 418
and social benefits in cities. Urban Forestry & Urban Greening, 21, 146-152.
419
https://doi.org/10.1016/j.ufug.2016.11.015 420
Arenas, J. M., Escudero, A., Mola, I., & Casado, M. A. (2017). Roadsides: an opportunity for 421
biodiversity conservation. Applied Vegetation Science, 20, 527–537.
422
https://doi.org/10.1111/avsc.12328 423
Aronson, J., & Alexander, S. (2013). Ecosystem restoration is now a global priority: time to 424
roll up our sleeves. Restoration Ecology, 21, 293–296. https://doi.org/10.1111/rec.12011 425
Aszód, L. (1935). Adatok a nyírségi vegetáció ökológiájához és szociológiájához [Data on the 426
vegetation ecology and sociology of Nyírség]. Tiscia, 1, 75–107 (in Hungarian).
427
Bakker, J. P., & Berendse, F. (1999). Constraints in the restoration of ecological diversity in 428
grassland and heathland communities. Trends in Ecology & Evolution, 14, 63–68.
429
https://doi.org/10.1016/S0169-5347(98)01544-4 430
Benjamini, Y., & Hochberg, Y. (1995). Controlling the false discovery rate: a practical and 431
powerful approach to multiple testing. Journal of the Royal Statistical Society, Series B 432
(Methodological), 289–300.
433
Bond, W. J. (2016). Ancient grasslands at risk. Science, 351, 120–122. doi:
434
10.1126/science.aad5132 435
Borhidi, A. (1995). Social behaviour types, the naturalness and relative ecological indicator 436
values of the higher plants in the Hungarian flora. Acta Botanica Hungarica, 39, 97–181.
437
Borhidi, A. (1996). Critical revision of the Hungarian plant communities. Janus Pannonius 438
University, Pécs, HU.
439
Boros, Á. (1929). Die Flora und die Planzengeographen verhältnisse des Nyírség [in 440
German]. - Mathematischer und Naturwissenschaftlicher Anzeiger der Ungarischen 441
Akademie der Wissenschaften XLVI. Sonderabdruck, 48–59.
442
Botta-Dukát, Z. (2008). Invasion of alien species to Hungarian (semi-)natural habitats. Acta 443
Botanica Hungarica, 50, 219–227. https://doi.org/10.1556/ABot.50.2008.Suppl.11 444
Bölöni, J., Botta‐Dukát, Z., Illyés, E., & Molnár, Z. (2011). Hungarian landscape types:
445
classification of landscapes based on the relative cover of (semi‐) natural habitats. Applied 446
Vegetation Science, 14, 537-546. https://doi.org/10.1111/j.1654-109X.2011.01139.x 447
Broadhurst, L. M., Lowe, A., Coates, D. J., Cunningham, S. A., McDonald, M., Vesk, P. A., 448
& Yates, C. (2008). Seed supply for broadscale restoration: maximizing evolutionary 449
potential. Evolutionary Applications, 1, 587–597. https://doi.org/10.1111/j.1752- 450
4571.2008.00045.x 451
Clarke, K. R. (1993). Non-parametric multivariate analysis of changes in community 452
structure. Australian Journal of Ecology, 18, 117–143. https://doi.org/10.1111/j.1442- 453
9993.1993.tb00438.x 454
Cramer, V., Hobbs, R., & Standish, R. (2008). What’s new about old fields? Land 455
abandonment and ecosystem assembly. Trends in Ecology & Evolution, 23, 104–112.
456
https://doi.org/10.1016/j.tree.2007.10.005 457
Csecserits, A., & Rédei, T. (2001). Secondary succession on sandy old-fields in Hungary.
458
Applied Vegetation Science, 4, 63–74. https://doi.org/10.1111/j.1654-109X.2001.tb00235.x 459
Csecserits, A., Czúcz, B., Halassy, M., Kröel-Dulay, G., Rédei, T., Szabó, R., ... Török, K.
460
(2011). Regeneration of sandy old-fields in the forest steppe region of Hungary. Plant 461
Biosystems, 145, 715-729. https://doi.org/10.1080/11263504.2011.601340 462
Csecserits, A., Botta-Dukát, Z., Kröel-Dulay, G., Lhotsky, B., Ónodi, G., Rédei, T., ...
463
Halassy, M. (2016). Tree plantations are hot-spots of plant invasion in a landscape with 464
heterogeneous land-use. Agriculture, Ecosystems & Environment, 226, 88-98.
465
https://doi.org/10.1016/j.agee.2016.03.024 466
Deák, B., Hüse, B., & Tóthmérész, B. (2016). Grassland vegetation in urban habitats–testing 467
ecological theories. Tuexenia, 36, 379-393. doi: 10.14471/2016.36.017 468
European Commission (2013). Building a Green Infrastructure for Europe. European Union.
469
Grime, J. P. (1979). Plant strategies and vegetation processes. Chichester, UK: J. Wiley and 470
Sons.
471
Hackett, M., & Lawrence, A. (2014). Multifunctional role of field margins in arable farming.
472
CEA report. Cambridge Environmental Assessments.
473
Halassy, M. (2001). Possible role of the seed bank in the restoration of open sand grassland in 474
old fields. Community Ecology, 2, 101–108. https://doi.org/10.1556/ComEc.2.2001.1.11 475
Havens, K., Vitt, P., Still, S., Kramer, A.T., Fant, J. B., & Schatz, K. (2015). Seed sourcing 476
for restoration in an era of climate change. Natural Areas Journal, 35, 122–133.
477
https://doi.org/10.3375/043.035.0116 478
Hedberg, P., & Kotowski, W. (2010). New nature by sowing? The current state of species 479
introduction in grassland restoration, and the road ahead. Journal for Nature Conservation, 480
18, 304–308. https://doi.org/10.1016/j.jnc.2010.01.003 481
Hostetler, M., Allen, W., & Meurk, C. (2011). Conserving urban biodiversity? Creating green 482
infrastructure is only the first step. Landscape and Urban Planning, 100, 369–371.
483
https://doi.org/10.1016/j.landurbplan.2011.01.011 484
Hothorn, T., Bretz, F., & Westfall, P. (2008). Simultaneous inference in general parametric 485
models. Biometrical Journal, 50, 346–363. https://doi.org/10.1002/bimj.200810425 486
Hüse, B., Szabó, S., Deák, B., & Tóthmérész, B. (2016). Mapping an ecological network of 487
green habitat patches and their role in maintaining urban biodiversity in and around Debrecen 488
city (Eastern Hungary). Land Use Policy, 57, 574-581. doi:10.1016/j.landusepol.2016.06.026 489
Kettenring, K. M., Mercer, K. L., Reinhardt Adams, C., & Hines, J. (2014). Editor’s choice:
490
Application of genetic diversity–ecosystem function research to ecological restoration.
491
Journal of Applied Ecology, 51, 339–348. https://doi.org/10.1111/1365-2664.12202 492
Kiehl, K., Thormann, A., & Pfadenhauer, J. (2006). Evaluation of initial restoration measures 493
during the restoration of calcareous grasslands on former arable fields. Restoration Ecology, 494
14, 148–156. https://doi.org/10.1111/j.1526-100X.2006.00115.x 495
Kiehl, K., Kirmer, A., Donath, T. W., Rasran, L., & Hölzel, N. (2010). Species introduction in 496
restoration projects–Evaluation of different techniques for the establishment of semi-natural 497
grasslands in Central and Northwestern Europe. Basic and Applied Ecology, 11, 285–299.
498
https://doi.org/10.1016/j.baae.2009.12.004 499
Király, G. (Eds.) (2009). Új magyar füvészkönyv. Magyarország hajtásos növényei.
500
Határozókulcs. Ábrák [New Hungarian herbal. Vascular plants of Hungary. Identification 501
keys]. Jósvafő, HU: Aggteleki Nemzeti Park Igazgatóság.
502
Klaus, V. H. (2013). Urban grassland restoration: A neglected opportunity for biodiversity 503
conservation. Restoration Ecology, 21, 665–669. https://doi.org/10.1111/rec.12051 504
Kovács-Hostyánszki, A., Espíndola, A., Vanbergen, A. J., Settele, J., Kremen, C., & Dicks, L.
505
V. (2017). Ecological intensification to mitigate impacts of conventional intensive land use on 506
pollinators and pollination. Ecology Letters, 20, 673-689. https://doi.org/10.1111/ele.12762 507
Liquete, C., Kleeschulte, S., Dige, G., Maes, J., Grizzetti, B., Olah, B., & Zulian, G. (2015).
508
Mapping green infrastructure based on ecosystem services and ecological networks: A Pan- 509
European case study. Environmental Science & Policy, 54, 268–280.
510
https://doi.org/10.1016/j.envsci.2015.07.009 511
Maes, J., Barbosa, A., Baranzelli, C., Zulian, G., e Silva, F. B., Vandecasteele, I., … Lavalle, 512
C. (2015). More green infrastructure is required to maintain ecosystem services under current 513
trends in land-use change in Europe. Landscape Ecology, 30, 517–534.
514
https://doi.org/10.1007/s10980-014-0083-2 515
Martens, D., Gutscher, H., & Bauer, N. (2011). Walking in ‘wild’ and ‘tended’ urban forests:
516
the impact on psychological well-being. Journal of Environmental Psychology, 31, 36–44.
517
https://doi.org/10.1016/j.jenvp.2010.11.001 518
Mitchley, J., Jongepierová, I., & Fajmon, K. (2012). Regional seed mixtures for the 519
re‐creation of species‐rich meadows in the White Carpathian Mountains: results of a 10‐yr 520
experiment. Applied Vegetation Science, 15, 253-263. https://doi.org/10.1111/j.1654- 521
109X.2012.01183.x 522
Molnár, Z., & Botta-Dukát, Z. (1998). Improved space-for-time substitution for hypothesis 523
generation: secondary grasslands with documented site history in SE-Hungary.
524
Phytocoenologia, 28, 1–29.
525
Oksanen, J., Blanchet, F. G., Friendly, M., Kindt, R., Legendre, P., McGlinn, D., … Wagner, 526
H. H. (2017, August 24). Vegan: Community Ecology Package. R package version 2.4-4.
527
Retrieved from https://CRAN.R-project.org/package=vegan 528
Oliveira, G., Clemente, A., Nunes, A., & Correia, O. (2014). Suitability and limitations of 529
native species for seed mixtures to revegetate degraded areas. Applied Vegetation Science, 17, 530
726–736. https://doi.org/10.1111/avsc.12099 531
Papp, L., & Dudás, M. (1989). Adatok a Közép-, a Dél-Nyírség és környékének botanikai 532
értékeiről [Data of botanical values of Middle-, South-Nyírség and surroundings] [in 533
Hungarian]. Calandrella II/2.
534
Pinheiro, J., Bates, D., DebRoy, S., Sarkar, D., Heisterkamp, S., Van Willigen, B., &
535
Maintainer, R. (2017, January 22). Package ‘nlme’. Linear and nonlinear mixed effects 536
models, 3-1. R package version 3.1-128. Retrieved from https://cran.r- 537
project.org/web/packages/nlme/index.html 538
Piper, J. K., Schmidt, E. S., & Janzen, A. J. (2007). Effects of species richness on resident and 539
target species components in a prairie restoration. Restoration Ecology, 15, 189–198.
540
https://doi.org/10.1111/j.1526-100X.2007.00203.x 541
Prach, K., & Hobbs, R. J. (2008). Spontaneous succession versus technical reclamation in the 542
restoration of disturbed sites. Restoration Ecology, 16, 363–366.
543
https://doi.org/10.1111/j.1526-100X.2008.00412.x 544
Prach, K., Tichý, L., Lencová, K., Adámek, M., Koutecký, T., Sádlo, J., … Řehounková, K.
545
(2016). Does succession run towards potential natural vegetation? An analysis across seres.
546
Journal of Vegetation Science, 27, 515–523. https://doi.org/10.1111/jvs.12383 547
R Core Team (2016). R: A language and environment for statistical computing. R Foundation 548
for Statistical Computing, Vienna, Austria. Retrieved from https://www.R-project.org/.
549
Rasran, L., Vogt, K., & Jensen, K. (2006). Seed content and conservation evaluation of hay 550
material of fen grasslands. Journal for Nature Conservation, 14, 34–45.
551
https://doi.org/10.1016/j.jnc.2005.08.002 552
Romão, C. (1996). Interpretation manual of European Union habitats. European Commision.
553
Ruprecht, E. (2006). Successfully recovered grassland: a promising example from Romanian 554
old-fields. Restoration Ecology, 14, 473–480. https://doi.org/10.1111/j.1526- 555
100X.2006.00155.x 556
Shackelford, N., Hobbs, R., Burgar, J. M., Erickson, T. E., Fontaine, J. B., Laliberte, E., … 557
Standish, R. (2013). Primed for change: Developing ecological restoration for the 21st 558
century. Restoration Ecology, 21, 297–304. https://doi.org/10.1111/rec.12012 559
Singmann, H., Bolker, B., Westfall, J., & Aust, F. (2018, April 4). Afex: Analysis of Factorial 560
Experiments. R package version 0.20-1. Retrieved from https://CRAN.R- 561
project.org/package=afex 562
Soó, R. (1939). A Nyírség természeti kincsei [Nyírség natural assets] [in Hungarian].
563
Budapest, HU: Merkantil-nyomda.
564
Soó, R. (1940). Vergangenheit und Gegenwart der pannonischen Flora und Vegetation. Nova 565
Acta Leopold, 9, 1–49.
566
Standish, R., Hobbs, R., & Miller, J. R. (2013). Improving city life: options for ecological 567
restoration in urban landscapes and how these might influence interactions between people 568
and nature. Landscape Ecology, 28, 1213–1221. https://doi.org/10.1007/s10980-012-9752-1 569
Török, K., Csecserits, A., Somodi, I., Kövendi‐Jakó, A., Halász, K., Rédei, T., & Halassy, M.
570
(2018a). Restoration prioritization for industrial area applying multiple potential natural 571
vegetation modeling. Restoration Ecology, 26, 476-488. https://doi.org/10.1111/rec.12584 572
Török, P., Deák, B., Vida, E., Valkó, O., Lengyel, S., & Tóthmérész, B. (2010). Restoring 573
grassland biodiversity: Sowing low-diversity seed mixtures can lead to rapid favourable 574
changes. Biological Conservation, 143, 806–812.
575
https://doi.org/10.1016/j.biocon.2009.12.024 576
Török, P., Vida, E., Deák, B., Lengyel, S., & Tóthmérész, B. (2011). Grassland restoration on 577
former croplands in Europe: an assessment of applicability of techniques and costs.
578
Biodiversity and Conservation, 20, 2311–2332. https://doi.org/10.1007/s10531-011-9992-4 579
Török, P., Miglécz, T., Valkó, O., Kelemen, A., Tóth, K., Lengyel, S. & Tóthmérész, B.
580
(2012). Fast restoration of grassland vegetation by a combination of seed mixture sowing and 581
low-diversity hay transfer. Ecological Engineering 44, 133–138.
582
https://doi.org/10.1016/j.ecoleng.2012.03.010 583
Török, P., Janišová,M., Kuzemko, A., Rūsiņa, S.,& Stevanović, Z. D. (2018b). Grasslands, 584
their threats and management in Eastern Europe. In: Squires, V.R., Dengler, J., Feng, H., Hua, 585
L. (Eds.) Grasslands of the world: diversity, management and conservation. Boca Raton, US:
586
CRC Press.
587
Tzoulas, K., Korpela, K., Venn, S., Yli-Pelkonen, V., Kaz´mierczak, A., Niemela, J., &
588
James, P. (2007). Promoting ecosystem and human health in urban areas using Green 589
Infrastructure: a literature review. Landscape and urban planning, 81, 167–178.
590
https://doi.org/10.1016/j.landurbplan.2007.02.001 591
Valkó, O., Deák, B., Török, P., Kirmer, A., Tischew, S., Kelemen, A., … Tóthmérész, B.
592
(2016). High-diversity sowing in establishment windows: a promising new tool for enhancing 593
grassland biodiversity. Tuexenia, 36, 359–378. http://dx.doi.org/10.14471/2016.36.020 594
Vida, E., Valkó, O., Kelemen, A., Török, P., Deák, B., Miglécz, T., … Tóthmérész, B. (2010).
595
Early vegetation development after grassland restoration by sowing low-diversity seed 596
mixtures in former sunflower and cereal fields. Acta Biologica Hungarica, 61, 226–235.
597
https://doi.org/10.1556/ABiol.61.2010.Suppl.22 598
Zuur, A. F. (Ed.) 2009. Mixed effects models and extensions in ecology with R, Statistics for 599
biology and health. New York, US: Springer.
600 601
Table 1. Summary of restoration treatments applied. For details on seeding rates per species 602
see Török et al. (2018a). Quantity of grass and forb species by hay transfer are derived from 603
germination experiment (data not shown). One-third of the bales used for hay transfer or 604
mulch were sampled before distribution in 2014. Roughly cleaned samples of hay and mulch 605
were measured and germinated under controlled conditions (temperature: 16±0.3°C; dew 606
point:12±0.2g/m3; humidity: 84±0.7%) on clean construction sand in trays at the 607
experimental area of the National Botanical Garden, Vácrátót. Germinated adult species were 608
identified at species level. Germination data (number of specimen and species) from hay or 609
mulch samples were used to determine the seed content of species of hay and mulch; and to 610
estimate introduced seed quantity (kg/ha) to restoration plots (* seeds added by hay transfer or 611
mulch), by using thousand seed weight data.
612 613
614
Restoration plot COM DOM HAY1 HAY2
area (ha) 4.5 2.6 1 1.7
Preparatory plant
Timing 2013 autumn 2014 spring 2013 autumn 2013 autumn Preparatory plant
(kg/ha)
20 20 20 20
Hay transfer
Timing 2014 summer 2014 summer
Grass (bale) 26 40
Forb (bale) 5 6
Seeding
Timing 2014 autumn 2014 autumn
Dominant grass Festuca
pseudovina
Festuca rupicola
grass (kg/ha) 30 60 1.769* 0.717*
forb (kg/ha) 15 0.093* 0.08*
Mulching
Timing 2014 autumn 2014 autumn
Mulch (bale) 42 26
grass (kg/ha) 0.022* 0.007*
forb (kg/ha) 0.005* 0.132*
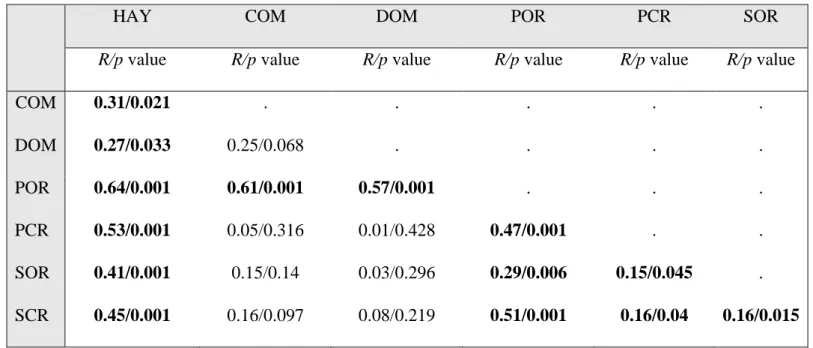
Table 2. Results of ANOSIM analyses. Composition of reference sites and restoration plots compared using ANOSIM analyses based on cover 615
data. Significant differences are given in bold. Abbreviations: primary closed reference (PCR), primary open reference (POR), secondary closed 616
reference (SCR), secondary open reference (SOR), commercial seed mixture (COM), seeds of a single dominant species (DOM), hay transfer 617
(HAY).
618
HAY COM DOM POR PCR SOR
R/p value R/p value R/p value R/p value R/p value R/p value
COM 0.31/0.021 . . . . .
DOM 0.27/0.033 0.25/0.068 . . . .
POR 0.64/0.001 0.61/0.001 0.57/0.001 . . .
PCR 0.53/0.001 0.05/0.316 0.01/0.428 0.47/0.001 . .
SOR 0.41/0.001 0.15/0.14 0.03/0.296 0.29/0.006 0.15/0.045 . SCR 0.45/0.001 0.16/0.097 0.08/0.219 0.51/0.001 0.16/0.04 0.16/0.015
Table 3. Guidance helping to select the most appropriate restoration treatment regarding six 619
priorities (naturalness = appropriate ratio of sociability of species, total cover, species 620
richness) or constraints (treatment effort, cost effectiveness, shortage of local propagules).
621
Treatments are assessed as highly appropriate/effective (+++), moderately 622
appropriate/effective (++) or less appropriate/effective (+) concerning six selection criteria.
623
Assessment was based on results of this study (see Fig. 3 and 4) supplemented by expert 624
knowledge. (* Lower treatment effort is considered more effective; mulching after seeding 625
increases efforts in commercial seed mixture and sowing of dominant species.) 626
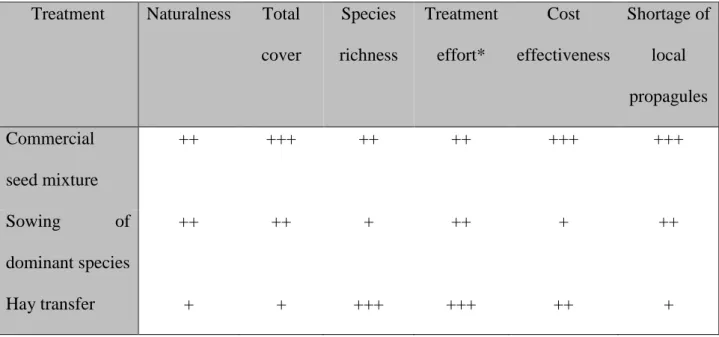
Treatment Naturalness Total cover
Species richness
Treatment effort*
Cost effectiveness
Shortage of local propagules Commercial
seed mixture
++ +++ ++ ++ +++ +++
Sowing of
dominant species
++ ++ + ++ + ++
Hay transfer + + +++ +++ ++ +
627
Figures 628
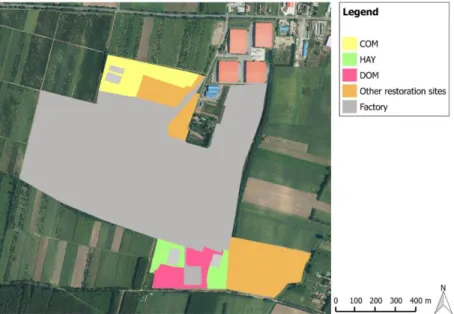
Figure 1. Map of three applied plant introduction treatments at an unused urban-industrial 629
area in Nyírség, NE Hungary. Restoration treatments were the following: commercial seed 630
mixture (COM); seeds of a single dominant species (DOM), hay transfer (HAY). Species is 631
listed in Appendix S3.
632
633 634
Figure 2. Non-Metric Multidimensional Scaling (NMDS) of the composition of three 635
restoration treatments (COM, DOM, HAY) and four reference types (PCR, POR, SCR, SOR) 636
based on cover data by using of Bray-Curtis dissimilarity. Abbreviations: primary closed 637
reference (PCR), primary open reference (POR), secondary closed reference (SCR), 638
secondary open reference (SOR), commercial seed mixture (COM), seeds of a single 639
dominant species (DOM), hay transfer (HAY).
640
641 642 643
Figure 3. Total cover (A) and species richness (B) of restoration plots and reference sites.
644
Figures represent the distribution of total cover/species richness data based on the minimum, 645
first quartile, median, third quartile, and maximum. Total cover (i.e. sum of individual 646
species´ cover) and species richness (i.e. number of species) were calculated for each of the 647
70 sampling units. Significant differences among study sites are indicated by lower case 648
letters. Abbreviations: primary closed reference (PCR), primary open reference (POR), 649
secondary closed reference (SCR), secondary open reference (SOR), commercial seed 650
mixture (COM), seeds of a single dominant species (DOM), hay transfer (HAY).
651
652 653
Figure 4. Total cover (A,C,E) and species richness (B,D,F) of sociability of species in 654
restoration plots and reference sites. Figures represent the distribution of total cover/species 655
richness data based on the minimum, first quartile, median, third quartile, and maximum.
656
Total cover (i.e. sum of individual species´ cover) and species richness (i.e. number of 657
species) were calculated for each of the 70 sampling units. Merged Borhidi (1995) categories:
658
1) natural constituents (NC); 2) disturbance tolerant species (DT); and 3) weeds (W).
659
Significant differences among study sites are indicated by lower case letters. Abbreviations:
660
primary closed reference (PCR), primary open reference (POR), secondary closed reference 661
(SCR), secondary open reference (SOR), commercial seed mixture (COM), seeds of a single 662
dominant species (DOM), hay transfer (HAY).
663
664
Supporting information 665
Additional supporting information may be found in the online version of this article:
666
Appendix S1. Map of study sites.
667
Appendix S2. Basic soil properties of the restoration and reference sites.
668
Appendix S3. List of plant species, their life form and sociability of species.
669
Appendix S4. Results of NMDS analysis based on presence/absence data.
670
Appendix S5. Results of ANOSIM analysis with species richness.
671