SZAKDOLGOZAT
Tóth Zoltán
2014
2
Cerebelláris konvulziók morfológiai és immunhisztokémiai vizsgálata
Tóth Zoltán
Témavezető: Prof. Dr. Mihály András tanszékvezető egyetemi tanár
az orvostudomány doktora
SZTE-ÁOK
Anatómiai, Szövet-és Fejlődéstani Intézet 2014
3
Tartalomjegyzék
1. Bevezetés ... 5
1.1 Az epilepsziák epidemiológiája ... 5
1.2Az epilepsziák etiológiája ... 6
1.3Az epilepsziák csoportosítása ... 8
2. Neuropatológiai elváltozások epilepsziában ... 10
2.1 Hippocampus sclerosis(HS) ... 10
2.2 Idegrendszeri tumorokhoz társuló epilepsziák ... 11
2.3 Cerebellaris atrophia ... 11
3. A cerebellum anatómiája és szövettana ... 13
4. Az akut konvulzív modell ... 14
5. Célkitűzések ... 16
6. Anyag-módszer ... 17
6.1 Kisagyi szemcsesejtek c-fos expressziójának vizsgálata 4-AP konvulziókat követően ... 17
6.2 Középső kisagykar átmetszése műtéti úton ... 17
6.3 Az operált és az áloperált kontroll csoport immunhisztokémiai vizsgálata ... 19
7. Eredmények ... 21
7.1 Az állatok viselkedésének megfigyelése ... 21
7.2 A kisagyi szemcsesejtek c-fos expressziójának immunhisztokémiai vizsgálata ... 23
7.3 A kísérleti állatok viselkedésének megyfigyelése a középső kisagykar lézió után ... 26
7.4 Az operált és áloperált állatok kisagyi szemcsesejtjeinek c-fos expressziója két órával a 4-AP kezelés után ... 27
7.5 Neuronális és szinaptológiai változások vizsgálata a kisagy és a híd területén a műtétet követően ... 30
8. Megbeszélés ... 32
9. Köszönetnyilvánítás ... 37
10. Irodalomjegyzék ... 38
4
Rövidítések jegyzéke
4-AP: 4-aminopyridin
AMPA: α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor ATP: adenozin trifoszfát
EEG: electroencephalographia
EPSP: excitatorikus posztszinaptikus potenciál GABA: gamma-amino vajsav
GFAP: glial fibrillary acidic protein HE: hematoxilin-eosin
HS: hippocampus sclerosis
IPSP: inhibitoros posztszinaptikus potenciál IR: immunreaktív
MCP: middle cerebellar peduncle MRI: mágneses rezonancia képalkotás mRNA: messenger ribonukleinsav NeuN: neuronal nuclear antigen NMDA: N-methyl-D-aspartate receptor PBS: foszfát puffer
PFA: paraformaldehid
TLE: temporális lebeny epilepszia
5
1. Bevezetés
A motoros konvulziók az epilepsziás rohamok és az epilepszia betegség legjellegzetesebb tünetei közé tartoznak. Epilepsziának hívjuk az agykéreg egészének vagy körülírt részének hiperszinkron, kóros kisülését. Ez a világ egyik legrégebben ismert betegsége, amelyet először Hippocrates írt le a Krisztus előtti 5.
században (Adams, 1849). Az epilepsziás kórképek rohamszerűen kezdődnek, eszméletvesztéssel vagy anélkül és egyéb motoros, szenzoros vagy vegetatív tünetekkel járnak. Epilepsziás rohamot okozhat az agyat ért bármely átmeneti vagy tartós ártalom. A rohamok szerint a betegséget két nagy csoportra oszthatjuk:
a. Akut symptomás roham az agy átmeneti kóros állapotának következménye, amely lehet láz, infekció, tumor, alkohol-drogmegvonás, hypoglikémia.
b. Az epilepszia betegség esetén az agyvelő tartós működészavara áll fenn, melynek eredményeképpen spontán ismétlődő rohamok jelentkeznek. A nemzetközileg elfogadott epileptológiai nomenklatúra-rendszer szerint akkor beszélünk epilepszia betegségről, ha a beteg rohamai krónikusan, sztereotip módon, többnyire ugyanazon klinikai tünetek formájában ismétlődnek, minden látható, kiváltó ok nélkül.
1.1 Az epilepsziák epidemiológiája
A betegség prevalenciája 1%, incidenciája 40-70/100000/év, kumulatív incidenciája 5-8%. A világon jelenleg kb. 50 millió ember érintett manifeszt epilepsziában, amely kezelést igényel; mindazonáltal a betegek 30%-a terápia rezisztens. (WHO.
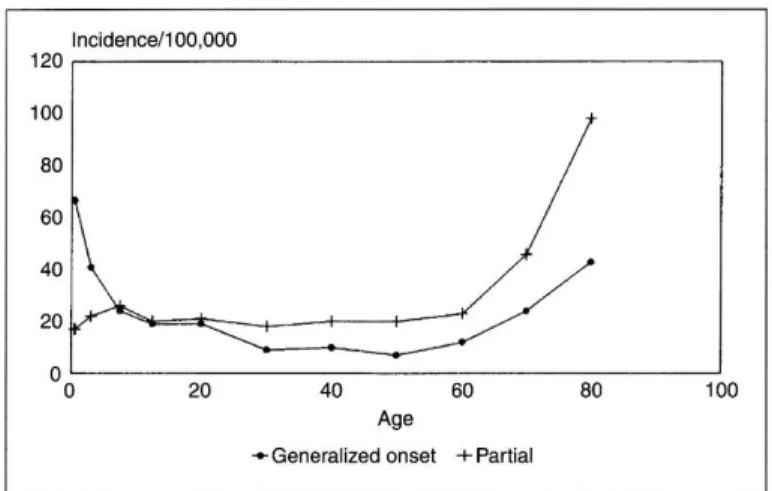
Epilepsy) A betegek csaknem 90%-a a fejlődő országokban él. Becslések szerint Európában 6 millió embernek van aktív epilepsziája, amelynek kezelésére a WHO évi 20 billió eurót fordít. (International Bureau for Epilepsy, WHO) Magyarországon kb. 60-70 ezer diagnosztizált beteg él. Az életkor szempontjából a 60 év felettiek a legveszélyeztetettebbek: ebben a korcsoportban a prevalencia 4- 5%-ra emelkedik (1. ábra).
6
1.2 Az epilepsziák etiológiája
Az epilepsziás roham kialakulásának hátterében egy neuron populáció fokozott excitabilitása, valamint a neuronok spontán és szinkronizált kisülése áll. A fokozott ingerlékenység az EPSP/IPSP arányának felborulását jelenti, amelyet több tényező is előidézhet. Akut symptomás rohamot okozhat koponyatrauma, infekció, fejlődési rendellenességek, intracranialis tumor, agyi keringészavarok, alkoholizmus, alkohol-és drogmegvonás, valamint bizonyos gyógyszerek. Ezek a patogén hatások hajlamosítanak az epilepszia betegség kialakulására (Nikl, 2004). Az inzultust követően egy látens epizód figyelhető meg, amely az agyat ért káros hatástól az első spontán görcs jelentkezéséig tart (Engel, 2005). A látens időszak alatt végbemenő folyamatok közül a legfontosabb a reaktív gliosis, amely az agyat ért káros hatást követő időszak kezdetén protektív hatású, a későbbiekben viszont gátolja az idegrendszer regenerációját (Sofroniew, 2009). Az astrocyták számos strukturális és funkcionális feladatot látnak el (Barres, 2008). Szerepük van a szinaptogenesisben (Christopherson et al., 2005) és direkt szinaptikus kapcsolatban állnak az idegsejtekkel is (Bushong et al., 2002). Az astrocyták aktivációja számos hatás következménye lehet, úgy mint az agyat ért sérülés, vagy a neurotranszmitterek (glutamát, GABA, acetilkolin) fokozott felszabadulása (Halassa and Haydon, 2010). A gliasejtek szabályozzák az extracelluláris tér ionösszetételét, alacsony K+ szintet biztosítva (Oliet et al., 2001, Amedee et al., 1997). A gliasejt diszfunkció
1. ábra: Az epilepszia incidenciája életkorok szerint. Forrás:
Hauser WA. Seizures disorders: the changes with ages.
Epilepsia 1992;33:6-14
7
görcsöket okozhat a K+ koncentráció emelkedése miatt, ami a membrán repolarizációt gátolja így fokozódik a neuronok ingerelhetősége (Traynelis et al., 1988, Sypert et al., 1974). Az intracranialis tumorok szintén okozhatnak epilepsziás rohamokat, mivel a peritumoralis oedema az agy nyomási atrophiáját és reaktív gliosist vált ki, amely csökkenti az intercelluláris teret és fokozza az idegsejtek excitabilitását (Jefferys et al., 1985,1995; Hossman et al., 1982). A manifeszt epilepszia progressziója során megfigyelhető neurodegeneratív változások közül a legfontosabb az excitotoxikus neuron degeneráció, amelynek kialakulásáért a fokozott glutamát felszabadulás, valamint az AMPA és NMDA receptorok fokozott aktivációja, a fokozott Ca2+ influx és az aktiválódott apoptotikus mechanizmusok valamint a mitokondrialis dysfunctio lehet a felelős (Maria Thom, 2004). A megmaradt neuronok excitabilitása fokozódik. Ennek a magyarázata, hogy az epilepsziás roham során fokozódik az ATP igény, amelyet a csökkent vérellátás nem tud biztosítani, így hypoxia és acidosis alakul ki, amely serkenti a neuronok ingerelhetőségét (Duffy et al., 1975, Howse et al., 1974). Ezen folyamatok végeredményben a membrán kóros permeabilitását, az excitatorikus és inhibitorikus neurotranszmitterek arányának felborulását okozzák (2. ábra) és később a manifesztálódott epilepszia progressziójáért is felelősek (Pitkänen et al., 2011, Castillo et al., 1997). Az ismétlődő epilepsziás rohamok összeadódó hatása agyi atrófiához vezet (Szabó et al., 2003). Az agy régiói közül sérülékenység szempontjából a hippocampus, neocortex, thalamus és a cortex cerebelli emelhető ki (Men et al., 2000).
A görcsök kialakulásáért az afferens pályákon érkező excitáció, a kóros reverberáció, és a serkentő és gátló neuronhálózatok közötti kényes egyensúly felborulása szükséges. A gátló rendszerek kontrollja alól kikerült neuronok képesek spontán, egyidejű aktivációra (Olney et al., 1986).
8
1.3 Az epilepsziák csoportosítása
Az idiopátiás epilepsziák esetében konkrét kóroki tényezőt nem lehet megállapítani, de a genetikai prediszpozíció bizonyított és a poligénes öröklődés valószínűsíthető.
Ezen betegeknél valamilyen központi idegrendszeri fejlődési rendellenesség, esetleg neurotranszmitter-anyagcsere zavar állhat a háttérben. Az epilepsziák leggyakrabban alkalmazott felosztása az EEG kép alapján történik, mely szerint megkülönböztetünk fokális/parciális és generalizált rohamokat. A fokális rohamok az agy valamely körülírt károsodásának eredményeképpen jönnek létre, míg a generalizált rohamok esetén kétoldali motoros jelenségeket és EEG-n szimmetrikus féltekei izgalmi jeleket figyelhetünk meg. (Mumenthaler, Marco: Neurológia, 4.
kiadás)
2. ábra: az akut szimptomatikus epilepszia pathomechanizmusa: a vér-agy gát integritásának felborulása, elektrolit zavarok, vérzés vagy térfoglalás tumorok vagy infectio miatt. A haemosiderinből felszabaduló Fe2+ fokozza a lipid peroxidációt és az arachidonsav kaszkádok aktivációját, ami károsítja a membránt és fokozza a sejtek excitabilitását a fokozott Ca2+ influx által (Carl J Vaughan:Pathophysiology of Acute Symptomatic Seizures)
9
I. Generalizált rohamok
Generalizált rohamok esetében a görcs az egész testre kiterjed. Két alapvető formáját ismerjük: a) primer generalizáció, melynél mindkét hemispherium részt vesz a rohamban; b) szekunder generalizáció, melynél kezdetben a cortex valamely körülírt részében figyelhető meg működészavar, amely később átterjed az egész kéregre. Típusai:
a. Generalizált tónusos-klónusos roham (grand mal): a leggyakoribb rohamforma, amely általában vokalizációval kezdődik, majd a beteg eszméletét veszti, összecsuklik. Ezt követi a tónusos fázis, mely azt jelenti, hogy az izmok testszerte megfeszülnek és átmeneti apnoe társul hozzá. A klónusos fázis alatt az izmok ritmusos rángása figyelhető meg, a száj habzása, enuresis és olykor nyelvharapás mellett.
b. Absence roham (petit mal): genetikailag determinált epilepsziákban fordul elő, főként gyermekkorban. A gyermek tevékenységében megakad, elréved, arca kifejezéstelenné, bambává válik. Motoros tünete általában nincs, a roham 5-10 sec.- ig tart.
c. Myoklónusos roham: genetikusan determinált epilepsziák mellett enkefalopátiás betegeknél is megfigyelhető. Hirtelen kezdődő, gyors végtagizom rángás jellemzi, amely aszimmetrikusan jelentkezik tudatzavar nélkül.
II. Partialis rohamok
1. Simplex partialis rohamok
Tudatzavar nem társul hozzá. Komplex partialis vagy generalizált tónusos-klónusos rohamok bevezető tünete lehet („aura”). Megjelenési formája az érintett agyterülettől függ, mivel bármely lebenyből kiindulhat.
2. Komplex partialis rohamok
Leggyakrabban a temporalis lebenyből indul ki, gyakran simplex partialis vagy psychosensoros élmény vezeti be (szaglási hallucináció, dejà vu, jamais vu).
10
Tudatzavar, elrévedés, homályállapot jellemzi, amelyhez gyakran különféle automatizmusok társulnak (orális automatizmusok, kezek tördelése, matatás).
Fajtái:
csak tudatzavarral jár;
gondolkodászavarral jár;
affektív tünetekkel jár;
pszichoszenzoros rohamok;
pszichomotoros rohamok;
komplex rohamok.
3. Partialis rohamok másodlagos generalizációval. Az agykéreg egy körülírt részének kóros aktivitása ráterjed a cortex egészére.
III. Nem osztályozható rohamok
Ezekben az esetekben a betegséget sem tünettani, sem az EEG kép alapján nem tudjuk besorolni egyik klasszikus formába sem, illetve a háttérben egyéb kiváltó ok nem található.
2. Neuropatológiai elváltozások epilepsziában
2.1 Hippocampus sclerosis (HS)
Azon betegek 50%-ban akikben HS kimutatható a kórelőzményben kimutatható a kora gyermekkori agysérülés. A károsodást követően egy tünetmentes időszak következik, amely után késő gyermekkorban vagy korai felnőttkorban jelentkezik az első spontán görcsroham. Valószínűsíthető, hogy a korai életkorban bekövetkező agysérülés vagy lázas eclampsia lehet az iniciáló faktor, amely celluláris és molekuláris károsodások által olyan elváltozásokat okoz, amelyek később manifeszt epilepsziát okoznak (Ozkara et al., 2005). Szoros kapcsolat mutatható ki a HS és az epilepszia progressziója között, sőt a HS ritkán kóroki tényező is lehet az időskori dementia kialakulásában. Számos MRI vizsgálat kimutatta, hogy
11
krónikus epilepsziás betegekben a hippocampusban jelentős volumencsökkenés figyelhető meg. A HS temporális lebeny epilepsziát (TLE) okozhat, amelyet az is bizonyít, hogy azon TLE-s betegek, akik anterior temporális lobectomián vagy szelektív hippocampectomián esnek át, az esetek nagy részében görcsmentessé válnak (Shaefi et al., 2003). A HS diagnosztikai kritériumai a hippocampus piramis sejtjeinek szignifikáns pusztulása, főleg a CA1 régióban és a reaktív gliosis (Maria Thom; 2004). Ugyanezen folyamatok megfigyelhetők az amygdalában és az entorhinalis cortexben is, amely a hippocampus egyik legfontosabb afferentációját adja (Yilmazer-Hanke et al., 2000). Mivel az entorhinalis cortex reciprok összeköttetésben áll a hippocampusszal, az itt végbemenő strukturális változások elősegíthetik a HS progresszióját. A HS-s betegekben kimutatható továbbá a temporális lebeny laterális részében végbemenő sejtszám csökkenés, amely főleg a II. és a III. rétegben figyelhető meg és jól korrelál a HS progressziójával.
2.2 Epilepsziához társult low-grade tumorok (vagy: idegrendszeri tumorokhoz társuló epilepsziák)
Bármely központi idegrendszeri tumor okozhat epilepsziás rohamokat, főleg a temporális lebenyt érintő astrocytomák, oligodendrogliomák és neuroepitheliomák (Aronica et al., 2001).
2.3 Cerebelláris atrófia
A hippocampalis sclerosis mellett, a leggyakrabban előforduló és régen felismert elváltozás a cerebelláris károsodás (Spielmeyer, 1930., Dam, 1987., Botez et al., 1988). A kisagyi atrófia okai között szerepel a görcs következtében kialakuló celluláris károsodás, a krónikus phenytoin kezelés, a keresztezett cerebralis hemiatrófia miatti cerebelláris degeneráció, az ischaemiás sejtkárosodás a görcs alatt, vagy ezen hatások kombinációja, vagyis a neuropatológiai események szinergizmusa (Ney et al., 1994; Savić and Thorell, 1996; Specht et al., 1997). A kisagyi atrophia mértékét befolyásolja az életkor, a nem, és a fennálló epilepszia időtartama. Megfigyelések szerint a generalizált tónusos-klónusos görcsök száma és intenzitása, valamint a kisagyi atrophia mértéke korrelációt mutat (Hermann et
12
al., 2005). A kisagy sorvadása egyes esetekben, postmortem, makroszkóposan is látható epilepsziások kórboncolása során. A legjellemzőbb elváltozás a foliumok vékonyodása a vermis vagy a hemispheriumok területén, amely megfigyelhető mind az elülső, mind a hátsó lebenynél. A mikroszkópos vizsgálatok kimutatták, hogy a görcsökre leginkább a Purkinje-sejtek érzékenyek. A Purkinje-sejtek 0.01%-os krezil-ibolya festéssel megfigyelhetők és kirajzolódik a teljes dendritfájuk is. Krónikus epilepsziás betegekben a Purkinje-sejtek számának 50%- os csökkenése és kifejezett Bergmann gliosis mutatható ki a kontrollhoz képest ( Crooks et al., 2000). Néhány esetben torpedó-szerű axon duzzadás is megfigyelhető a megmaradó Purkinje- sejteken, ami következményes spontán sejtpusztulás előjele. MRI vizsgálatok eredményei szerint a kisagyi atrófiát drasztikus térfogat csökkenés jellemzi, amelyben kulcsszerepe van a pontocerebelláris pálya degenerációjának (Tan and Urich, 1984). A cerebellumot jellemző térfogat-arányváltozások, melyek a görcsrohamok intenzitásával állnak összefüggésben, arra utalnak, hogy ezek epilepszia alatti sérülések vagy prediszponáló faktorok a másodlagos generalizáció kialakulásában (Szabó et al., 2006). A kisagyi atrófia függ az epilepszia súlyosságától (Lawson et al., 2000;
Sandok et al., 2000; De Marco et al., 2003). Epilepsziás gyermekek egy speciális csoportjánál (epilepszia és hiperaktív viselkedészavar) csökkent frakcionális anisotropia mutatható ki a középső kisagykarnak megfelelően, amely megfelel a pontocerebelláris pályának (Bechtel et al., 2009). Több kutatás számol be olyan kisagyi tumorokról (főleg gangliogliomákról), amelyek konvulzív tünetekkel járnak. Ezen tumorok valamely kisagyi hemispheriumból indulnak és infiltrálják az azonos oldali középső kisagykart és hemifaciális ritmusos izomrángással (Harvey et al., 1996) vagy progresszív myoklónussal (Koh et al., 2010) járnak. Ezen irodalmi adatok alapján feltételezhető, hogy a cortico-ponto-cerebelláris pályáknak és általuk szállított poliszenzoros információnak egyaránt szerepe lehet a kisagyi atrófia patogenezisében és az epilepszia progressziójában. Ezen pályarendszer által közvetített sejtpusztulás jelentős tényező lehet a betegség progressziójára nézve, de nem ad magyarázatot arra a tényre, hogy a sejtpusztulás a kisagy elülső és hátulsó lebenyét egyaránt érinti: mivel ezen afferensek főleg a hátsó lebenybe projiciálnak.
Felvetődik továbbá a kisagynak az epileptogenesisben betöltött szerepe is (Mink et al., 2003). A számos tény ellenére viszonylag kevés konkrét adat áll a rendelkezésünkre a kisagy epilepsziában betöltött szerepére vonatkozóan.
13
Összefoglalva elmondható, hogy a cerebellumnak az epilepszia betegség kialakulásában és progressziójában is szerepe lehet, és a kisagyi atrophia mértéke összefügg az epilepszia szindróma fennállásának idejével és a rohamok számával (Hagemann et al., 1999).
3. A cerebellum és a középső kisagykar anatómiája és szövettana
A kisagy a hátsó koponyaárokban, a tentorium cerebelli alatt helyezkedik el. Két hemisheriumból és azokat összekötő páratlan vermisből áll. Felszínén harántul futó fissurákat találunk. A két legfontosabb fissura a kisagyat három lebenyre osztja, amelyek megfelelnek a filogenetikai tagolódásnak is. A fissura prima a lobus anteriort és posteriort szeparálja, míg a fissura posterolateralis a lobus posterior és a lobus flocculonodularis között húzódik. A filogenetikai felosztás szerint megkülönböztetünk (1) archicerebellumot (vestibulocerebellum), amely a vestibularis magokkal áll összeköttetésben és az egyensúly fenntartásában játszik szerepet; (2) paleocerebellumot (spinocerebellum), amely az izmok, inak proprioceptív információit dolgozza fel; és (3) neocerebellumot (cerebrocerebellum), amely a neocortex összes lebenye felől poliszenzoros információkat kap. A kisagyat a kisagykarok kapcsolják a környező agyi struktúrákhoz. A pedunculus cerebellaris inferior a nyúltvelőhöz, a pedunculus cerebellaris medius a hídhoz, a pedunculus cerebellaris superior a mesencephalonhoz köti.
A kisagykéreg három rétegre osztható. A külső rétege a stratum moleculare, ahol kevés GABAerg kosársejtet, a Purkinje-sejtek dendritjeit valamint kúszórostokat és parallelrostokat találunk. A stratum ganglionare (stratum purkinjense) a szintén GABAerg Purkinje-sejtek szómáit tartalmazza. A stratum granulosum fő sejtfélesége a szemcsesejt, amely glutamáterg serkentő neuron. Dendritje moharostokkal szinaptizál, axonjuk felszáll a stratum moleculareba, ahol a Purkinje- és a kosársejtekkel kapcsolódik. A granuláris rétegben Golgi sejtek is találhatók, amelyek szintén gátló sejtek. Dendritjük a molekuláris rétegbe, axonjuk a moharostokhoz fut. A kisagykéreghez a kúszó- és moharost rendszeren keresztül diffúz excitáció érkezik, amely először a kisagyi magvakat innerválja. A moharostok a granuláris rétegben végződnek és a parallel rostokon keresztül a
14
Purkinje- és a kosársejteket ingerlik. A kúszórostok közvetlenül a Purkinje-sejtek dendritfáján végződnek. A kisagyi ingerlés Purkinje-sejt excitációt vált ki, amely a kisagyi magvakat gátolja. Mivel ezekhez már korábban diffúz excitáció érkezett a moharost és kúszórost kollaterálisokon keresztül, a Purkinje-sejtek felől érkező gátlás csak a diffúz excitációt alakítja értelmes ingermintázattá (Ito, 1984). A cerebellum legfontosabb feladatai az egyensúly fenntartása, a testhelyzet változtatása az izomtónus szabályozásával, a távolság felismerése. A kisagy sérülésének legfontosabb tünetei, amelyeket kisagyi tünetegyüttesként is ismerünk az izomhypotonia, az ataxia és az intenciós tremor.
A középső kisagykar (middle cerebellar peduncle-MCP, a legnagyobb a három pár kisagykar közül), a kisagyat a híddal köti össze és a neocortex összes lebenyéből származó poliszenzoros információt szállítja a nuclei pontis közvetítésével a kisagykéregbe. A motoros és praemotor áreából származó rostok keresztezettek és többnyire az elülső lebenyben végződnek, míg a parietalis, temporalis lebenyből, a cingularis kéregből és a mesialis temporalis lebenyből származó rostok főleg a hátsó lebenybe projiciálnak és ipsilateralisak (Ito, 1984, Brodal and Bjaalie 1979, Middleton and Strick, 1998, Savić and Thorell, 1998, Schmahmann and Sherman, 1998). A pontocerebellaris pálya a nuclei pontis neuronjaiból indul, és moharostok formájában végződik a stratum granulosumban.
4. Az akut konvulzív modell
Kísérleteinkben 4-AP-t használtunk, amely feszültségfüggő K+ csatornákat blokkol (Ka csatorna és Kv csatorna). Az elhúzódó repolarizáció elnyújtja az akciós potenciál időtartamát, fokozza a Ca2+ beáramlását a preszinaptikus axon végződésekbe (Thesleff, 1980), és ezáltal növeli a transzmitter felszabadulást (Rogawski and Barker, 1983). A megnövekedett transzmitter mennyiség növeli mind a serkentő- mind a gátló posztszinaptikus potenciálokat és kiterjedt görcsöket okoz in vivo (Pasantes-Morales et al., 1987; Szente and Baranyi, 1987; Mihály et al., 1990, 1997, 2000) és in vitro egyaránt (Kuhnt et al., 1983, Brückner and Heinemann, 2000; Marinelli et al., 2000). A 4-aminopyridin gyorsan átjut a vér-agy gáton, bejut a cerebrospinalis folyadékba, majd a vesén keresztül teljesen kiürül
15
(Lemeignan et al., 1984). Sem a vegyület, sem az általa okozott akut konvulzió nem okoz maradandó neurológiai károsodást ezért alkalmas a kísérletes epileptogenesis indukciójára.
A neuronális proto-onkogének az idegsejtek működésének változásaiért felelősek (Dragunow et al., 1989). A c-fos proto-onkogén az indukálható transzkripciós faktorok közé tartozik. A c-fos gént számos extracelluláris tényező aktiválhatja beleértve azon neurotranszmittereket, amelyek serkentik a Ca2+ beáramlást a posztszinaptikus neuronba. A glutamát fokozza a Ca2+ beáramlást és intracelluláris protein kináz kaszkádok által intenzív c-fos expressziót okoz (Szakács et al., 2002).
A c-fos protein a sejtmagba kerül, ahol transzkripciós faktorként más gének működését szabályozza és hatással van a központi idegrendszer fejlődési,
3. ábra: a 4-AP feltételezett hatásmechanizmusa a c-fos expressziót illetően. A 4-AP blokkolja a feszültségfüggő K+ csatornákat, ezáltal fokozza a serkentő neurotranszmitterek felszabadulását. A megnövekedett transzmitter mennyiség növeli a Ca2+ beáramlást a posztszinaptikus neuronba. Ez protein kináz kaszkádok révén a c-fos mRNA expresszióját eredményezi. A fos fehérje a sejtmagba transzlokálódik, ahol a Fos/Jun heterodimereken keresztül a DNS AP-1 szekvenciájához kötődik. Az AP-1 regulátor fehérjék amelyek részt vesznek a felnőtt agy degenerációs és regenerációs folyamatainak szabályozásában (Szakács et al., 2002).
16
adaptációs (Retchkiman et al., 1996) és degenerációs mechanizmusaira epilepsziában is (Herdegen and Waetzig, 2001; Gass et al., 1992). A c-fos fehérje expressziója erős szinaptikus aktivitás eredménye, jól korrelál a glutamát felszabadulással (Labiner et al., 1993) és immunhisztokémiai módszerekkel vizsgálható, ezért alkalmas a neuronális aktivitás vizsgálatára (Hoffmann and Lyo, 2002).
5. Célkitűzések
I. A kisagyi szemcsesejtek c-fos expressziójának vizsgálata 4-AP által keltett akut motoros konvulziókat követően a kisagyi vermis területén (a glutamáterg moharostok itt végződnek).
II. A középső kisagykar átmetszése műtéti úton (pontocerebelláris pálya átvágása).
III. A pálya lézió hatásának vizsgálata szemcsesejtek c-fos expressziójára 4-AP konvulziókat követően.
IV. A pálya degenerációjának következtében kialakuló neuronális és szinaptológiai változások vizsgálata immunhisztokémiai módszerekkel a kisagy és a híd területén.
6. Anyag-módszer
6.1 Kisagyi szemcsesejtek c-fos expressziójának vizsgálata 4-AP konvulziókat követően
Kísérleteinkhez 200-220g súlyú hím Wistar patkányokat használtunk (n=18). Az állatokat az Anatómiai Intézet állatházában tartottuk standard körülmények között
17
(világítás: 6:00-18:00-ig, 23oC). A görcskeltőt fiziológiás sóoldatban oldottuk(0.67 mg 4-AP/1ml 0.9% NaCl oldat) és intraperitonealis (i.p.) injekció formájában adagoltuk 4.75 mg/kg egyszeri dózisban (Mihály et al., 1990, 2000). A kontroll állatok csak az oldószert kapták.
Túlélési idő Állatok száma (kontroll)
Állatok száma (kezelt)
1,5 h 3 3
2h 3 3
3h 3 3
A 4-AP beadásától számított 1,5, 2 és 3 óra után az állatokat egy rövid dietiléteres altatás után transcardiálisan perfundáltuk (500 ml 4%-os paraformaldehid (PFA) 0,1 M-os PBS-ben oldva; pH 7.4), majd decapitáltuk. A kivett agyakat 1 órás posztfixálást követően egy éjszakán át 30%-os szukróz oldatban krioprotektáltuk.
Ezt követően 24 µm vastagságú, sagittalis síkú fagyasztott metszeteket készítettünk a vermisből, amelyeken c-fos immunhisztokémiát végeztünk. A c-fos IR sejtmagokat lobulusonként számoltuk (Image Pro Plus 4.5), majd statisztikai számításokat végeztünk (egyutas variancia analízis:ANOVA).
6.2 A középső kisagykar átmetszése műtéti úton
A műtét során 200-230 g súlyú hím Wistar patkányokat használtunk (n=44). Az állatokat két csoportra osztottuk. Az egyik csoport tagjain elvégeztük a cortico- ponto-cerebellaris pálya átmetszését végeztük, míg a másik az áloperált kontroll csoport volt. A műtét első lépéseként az állat altatószert kapott i.p.(80 mg/kg ketamin+20mg/kg pentobarbital+0.01 mg/kg atropin+2 ml 0.9%-os NaCl oldat). Az alvási idő átlagosan 93.35 min. volt. Az elalvás után az állat fejét sztereotaxiás padon rögzítettük majd a bal fül mögött vezetett metszéssel a bőrt megnyitottuk (4.ábra/A). A felületes izmok tompa preparálása után a musculus temporalis alsó szélénél szabaddá tettük a koponyát és azt fogászati fúróval megnyitottuk (4.ábra/B, C, D). A lobus floccularis eltávolítását követően láthatóvá vált a bal oldali középső kisagykar, amelyet elektrocoagulátor segítségével roncsoltunk (4.ábra/E). A
Táblázat 1: a kísérleti állatokat hat csoportra osztottuk a túlélési idők szerint. Minden csoport három állatot tartalmazott.
18
keletkező vérzés csillapítása diathermiával és Spongostan® szivaccsal történt (4.ábra/F). A keletkezett defektust izomlebennyel zártuk, a bőr egyesítéséhez atraumatikus monofil varróanyagot használtunk. A műtétek kivitelezéshez operációs mikroszkópot használtunk. Az áloperált kontroll csoporton ugyanezt a beavatkozást végeztük az agy sértése nélkül. A műtét után két hetes megfigyelési és gyógyulási periódust biztosítottunk. A posztoperatív időszakot követően 2 állat paraffinba beágyazott agyából coronalis síkú metszeteket készítettünk, amelyeken hematoxilin-eosin (HE) festést követően ellenőriztük a lézió helyét (12. ábra).
6.3 Az operált és áloperált kontroll csoportok immunhisztokémiai vizsgálatai
6.3.1 Az első csoportnál (10 műtött, 10 álműtött) a 4-AP beadását követően 2 óra elteltével a korábban leírt módszerrel (az altatott állatok perfundálása 4 %-os
4. ábra: a pedunculus cerebellaris medius műtéti átmetszésének lépései. A, B, C, D: a bőr megnyitása, a lágyrészek felpreparálása és a koponya megnyitása. E: műtéti terület a lézió után (8X-os nagyítás). F: vérzéscsillaptíás Spongostan® szivaccsal.
19
paraformaldehiddel, az agy kivétele, krioprotektálás és fagyasztott metszetek készítése) sagittalis síkú metszeteket készítettünk, külön az operált oldali és külön a nem operált oldali kisagyi haemispheriumokból. Az álműtött kontroll állatok esetében a jobb és a bal oldali féltekékből készültek a metszetek. Ezeken c-fos immunhisztokémiát alkalmaztunk, majd a c-fos IR sejtmagokat Image Pro Plus programmal számoltuk. A statisztikai analízist (ANOVA) követően megvizsgáltuk az általunk végzett műtét hatását a szemcsesejtek c-fos expressziójára.
6.3.2 A második csoport esetében (4 műtött, 4 álműtött) két héttel a műtét után coronalis síkú metszeteken GFAP (gliális fibrilláris savas protein) immunhisztokémiát végeztünk. A GFAP az astrocyták szelektív markere (Wagemann et al., 1995) és alkalmas a műtét utáni reaktív gliosis jelölésére.
6.3.3 A harmadik csoportnál (2 műtött, 2 álműtött) két héttel a műtét után coronalis síkú metszeteken synapsin I immunfestést alkalmaztunk. A synapsin I olyan foszfoprotein, amely a szinaptikus vezikulákhoz kapcsolódik és a neurotranszmitter felszabadulás szabályozásában vesz részt (Orlando et al., 2004). A molekula kapcsolatot létesít a vezikulumok és a mikrotubulusok között (Hilfiker et al., 1999, Petrucci et al., 1987). A cerebellum esetén a synapsin I alkalmas a moharost axonterminálisok jelölésére (Fakuya et al., 1999).
6.3.4 A negyedik csoport (4 műtött, 4 álműtött) esetében két héttel a műtét után coronalis síkú metszeteken NeuN festés történt. A NeuN (neuronal nuclear antigen) olyan általános neuron marker, amely alkalmas a kisagyi szemcsesejtek és a nuclei pontis neuronjainak jelölésére és az esetleges neuronpusztulás értékelésére (Mullen et al., 1992).
6.3.5 Az immunhisztokémia lépései
1. Triton és H2O2 kezelés: a vvt-k peroxidáz hatásának kivédése érdekében 0,5 %- os Triton detergens és 30 %-os H2O2 9:1 arányú keverékét használtuk.
2. Mosás: PBS-ben háromszor 15 percig.
3. Blokkolás: a háttérfestődés kiküszöbölése érdekében (primer szérum nem specifikus helyekre kötődik) a nem specifikus kötőhelyeket olyan antitesttel fedjük le, amelyet szekunder antitest nem ismer fel. Blokkoló oldat: 20 % NPS (normal pig serum) + 0,1 M PBS + 0,03 % Mertiolát (1 órán keresztül).
20
4. Primer antitestek: a megfelelő hígítású antitestekkel egy éjszakán át inkubáltuk a metszeteket rázószerkezeten. Primer antitestek és hígításaik:
anti-c-fos r. 1/2000 (Santa Cruz Biotechnology);
anti-synapsin I r. 1/200 (Abcam);
anti-NeuN m. 1/8000 (Chemicon);
anti-GFAP m. 1/10000 (Daco).
5. Mosás: háromszor 15 percig PBS-ben.
6. Kötő antitestek: megfelelő hígításban 1 órán keresztül. A hígítást kötő antitest hígítóval végeztük. Kötő antitestek:
rabbit (r.) B-GAR, 1/400 hígításban;
mouse (m.) B-BAM, 1/400 hígításban.
7. Mosás: háromszor 15 percig PBS-ben.
8. Harmadlagos antitest STA-PER (Streptavidin-peroxidáz) 1:2000 hígításban. A hígítást tercier hígítóval végeztük. Inkubáció 1 óra.
9. Mosás: kétszer 15 perc PBS-ben majd egyszer 15 perc TBS-ben.
10. Hívás:
Ni-DAB: GFAP, NeuN esetében, az oldat összetétele:
20 ml TBS;
3 ml Ni-ammónium-szulfát;
1 ml DAB (diaminobenzidin);
3,5 µl H2O2.
DAB: c-fos, synapsin I esetében, az oldat összetétele:
20 ml TBS;
1 ml DAB (diaminobenzidin);
21
3,5 µl H2O2.
Az oldatokat szűrőpapíron leszűrtük, a hívási idő 30 perc volt.
11. Mosás: háromszor TBS-sel.
12. Ezután Elvanol (polyvinil-alkohol) oldatból tárgylemezre húzzuk fel a metszeteket.
13. Szárítás egy éjszakán át.
14. Víztelenítés felszálló alkoholsorral (etanol és xylol segítségével) Minden oldatban öt percet tartottuk a metszeteket.
15. Lefedés DPX (depex) segítségével.
7. Eredmények
7.1 Az állatok viselkedésének megfigyelése
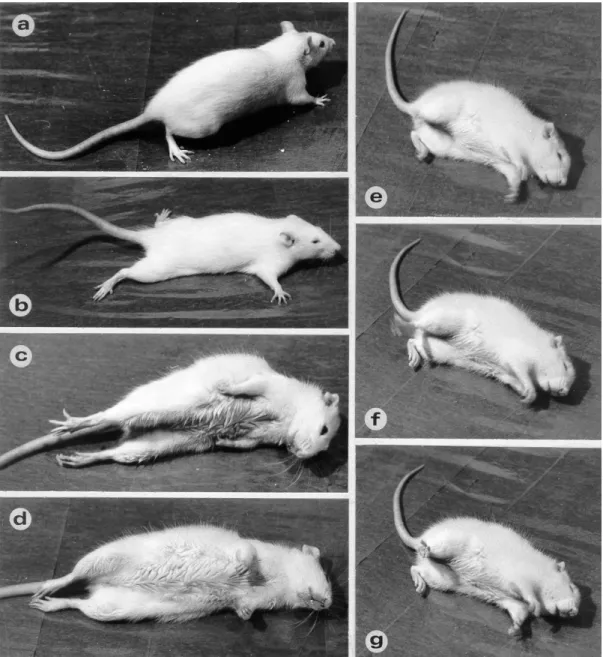
A 4-AP injekciót követő 15 percen belül jellegzetes tünetegyüttes volt megfigyelhető, amely úgy kezdődött, mint fokozott explorátoros tevékenység, majd a rágóizmok és a végtagok izomzatának remegése, végül generalizált tónusos- klónusos görcsök (GTCS) jelentkeztek (5. ábra). A görcstevékenység hirtelen lép fel és egyértelműen kezdődik (5. ábra/C), így lehetséges a latenciaidő pontos mérése (átlagosan 32,53 min).
22
5. ábra: a 4-AP hatásai a kísérleti állatok viselkedésére: b: egyensúlyzavarok, tremor;
c: a GTCS tónusos fázisa; d-g: a GTCS klónusos fázisa Forrás: Mihály et al., 1990)
23
7.2 A kisagyi szemcsesejtek c-fos expressziójának immunhisztokémiai vizsgálata
Str. moleculare
Str. Purkinjense
Str. granulosum
6. ábra: c-fos és parvalbumin kettős festés (20X). A c-fos IR sejtmagokat lobulusonként számoltuk. A nyilak a c-fos IR pozitív sejtmagokat jelölik.
7. ábra: a c-fos IR sejtmagok sűrűségének megoszlása két órával a 4-AP kezelést követően a kisagyi vermis területén. A méréseket lobulusonként végeztük,
(összesítő diagram).
24
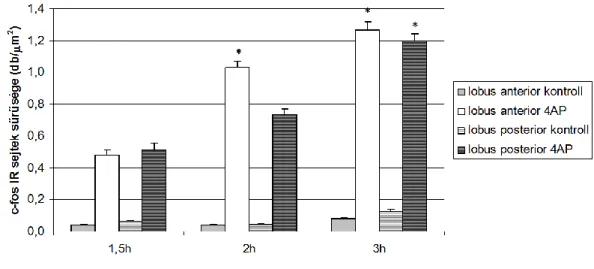
A 4-AP beadást követően a c-fos immunreaktív sejtmagok száma már 1,5 h után szignifikáns különbséget mutatott a kontrollhoz képest, majd folyamatos növekedés után 3 óránál érte el a maximumát (8. ábra).
További vizsgálatainkban a c-fos IR sejtmagokat a kisagyi vermis lobulusainak megfelelően számoltuk. Megpróbáltunk összefüggést keresni a lobus anterior és a lobus posterior és azok lebenykéinek c-fos expressziója és az eltérő afferentáció között. Különbséget találtunk a lobus anterior és a lobus posterior szemcsesejtjeinek aktiválódási dinamikája között (9. ábra).
8. ábra: a c-fos IR sejtmagok száma az idő függvényében a kisagyi stratum granulosum területén. A maximális immunreaktivitás az injekció beadása után 3 órával látható.
25
A lobus anterior és posterior szemcsesejtjeinek c-fos expressziója 1,5 óra elteltével még hasonló mintázatot mutat, azonban 2 és 3 óránál már a lobus anterior neuronjainak aktiválódása szignifikánsan erősebb a hátsó lebenyhez képest. A két lebeny aktiválódási különbségére az eltérő afferentáció elfogadható magyarázatot adhat.
9. ábra: a lobus anterior és lobus posterior szemcsesejtjei eltérő aktiválódási dinamikát mutatnak.
26
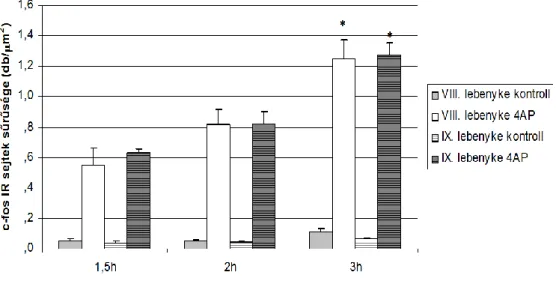
Ebből kiindulva az azonos afferentáció magyarázhatja azt is, hogy az I. és X.
lebenyke idegsejtjeinek nagyon hasonló az aktiválódási mintázata, annak ellenére, hogy eltérő lebenyben helyezkednek el (10. ábra). Megfigyeltük továbbá, hogy a lobus posterioron belül a VIII. és a IX. lebenyke szintén nagyon hasonló c-fos expressziós dinamikát mutat (11. ábra).
7.3 A kísérleti állatok viselkedése a középső kisagykar léziója után
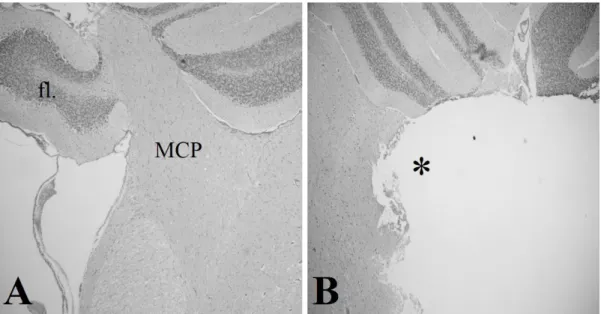
A két hetes megfigyelési időszak alatt nem tapasztaltunk spontán görcsöket és maradandó (szemmel látható) neurológiai góctüneteket. A 4-AP injekciót követő látencia idő átlagosan 41.5 min volt. A 4-AP görcsök morfológiája, a tónusos és klónusos fázisok időtartama és aránya szabad szemmel is észrevehető eltérést mutatott az operált és áloperált állatok között. Ennek pontos vizsgálatához későbbi EEG vizsgálatokat tervezünk. A lézió pontos helyének meghatározása coronalis síkú HE festett metszeteken történt (12. ábra). A nem operált oldalon (A) azonosítani lehet a lobus floccularist (fl.) és a tőle medialisan elhelyezkedő
10. ábra: az I. lebenyke (lobus anterior) és a X. lebenyke (lobus posterior) szemcsesejtjeinek aktivációs mintázata nagyon hasonló, annak ellenére, hogy más lebenyben helyezkednek el. A
jelenséget az azonos afferentációval (vestibularis afferensek) magyarázzuk.
11. ábra: a VIII. és IX. lebenyke hasonló arányban kap afferens rostokat a nuclei pontis felől.
27
pontocerebellaris pályát (MCP). Az operált oldalon (B) ezen struktúra teljes hiányát látjuk (csillag) a floccularis lebennyel együtt.
7.4 Az operált és áloperált állatok kisagyi szemcsesejtjeinek c-fos expressziója két órával a 4-AP kezelés után
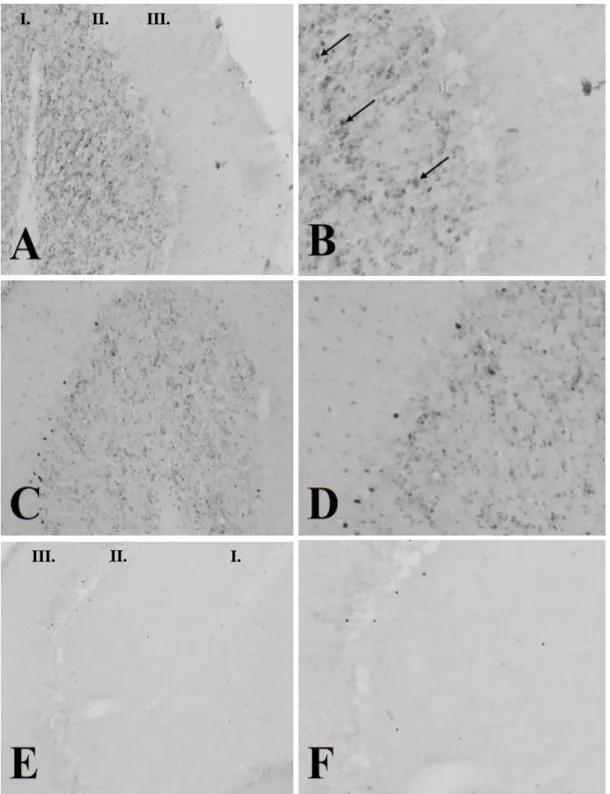
A 4-AP beadása után 2 órával vizsgáltuk a c-fos protein expressziót a kisagyi haemispheriumokban. A kontroll állatok esetében 2 óránál már szignifikáns különbséget tapasztaltunk a kontrollhoz képest, ezért választottuk ezt az időintervallumot. A kisagykéreg szemcsesejtjeinek c-fos expressziója jelentős csökkenést mutatott a nem operált oldalhoz és a kontrollhoz képest egyaránt (13. és 14. ábra).
12. ábra: a coronalis síkú metszeteken a MCP és a lobus floccularis kiesése látható (B) az ép oldalhoz képest (A).
(5X-ös nagyítás)
28
**
***
13. ábra: az operált oldali IR sejtek sűrűsége szgnifikánsan csökken az egészséges oldalhoz és az áloperált kontrollhoz
képest is ( p<0,001).
29
14. ábra: az operált oldali haemispheriumban (E 20X, F 40X) a szemcsesejtek c-fos expressziója jelentősen csökken a nem operált oldalihoz (C 20X, D 40X) és a kontrollhoz (A 20X, B 40X) képest
egyaránt. A nyilak a szemcsesejteket jelölik. I.: stratum granulosum; II.: stratum Purkinjense; III.
stratum moleculare
I. II. III.
I.
II.
III.
30
7.5 Neuronális és szinaptológiai változások vizsgálata a kisagy és a híd területén a műtétet követően
7.5.1 GFAP immunhisztokémia
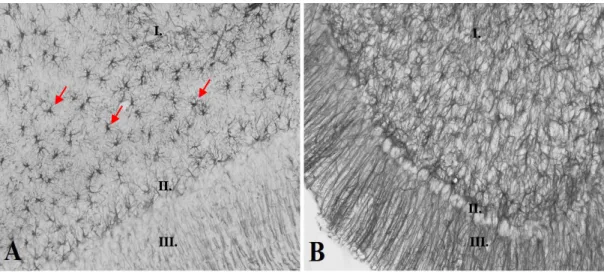
A műtét után két héttel készített GFAP festés intenzív Bergmann gliosist mutatott a kisagyban, főleg az operált oldalon (15. ábra). A híd területén szintén gliosist találtunk a basis pontisnak megfelelően. (16. ábra).
15. ábra: GFAP immunhiszkokémia intenzív gliosist mutatott (B) a kontrollhoz képest (A 20X) a kisagy területén a műtét után két héttel. I.: stratum granulosum, II.:
stratum Purkinjense, III.: stratum moleculare (20X-os nagyítás)
16. ábra: a híd területén a nuclei pontisnak megfelelően reaktív gliosist látunk az operált oldalon (C,F) a kontrollhoz (A,D) és a nem operált oldalhoz képest (B,E).
(A,B,C: 10X-es nagyítás; D,E,F: 20X-os nagyítás) III.
I.
II.
I.
II.
III.
31
7.5.2 Synapsin I immunhisztokémia
A synapsin I immunreaktivitás jelentős csökkenését tapasztaltuk a stratum granulosum és a stratum moleculare területén egyaránt (főként a laesio környezetében), ami arra utal, hogy a pontocerebellaris pálya átmetszésének következtében a moharost axon terminálisok degenerálódnak és a kisagyi glomerulusok összeesnek (17. ábra)
7.5.3 A pontocerebellaris pálya laesiojának hatása a nuclei pontis neuronjaira (NeuN festés)
A kisagyi stratum granulosumban nem találtunk jelentős eltérést az operált és az áloperált állatok NeuN-nel festett metszetei között. A basis pontis coronalis síkú metszetein eltérés mutatkozott az operált és nem operált oldal között. A nuclei pontis idegsejtjeinek immunreaktivitása csökken az operált oldalon a nem operált oldalhoz képest. Az eredmény kvantitatív értékelése, vagyis a NeuN immunreaktív sejtek számolása nem történt meg. A mérések a munka következő fázisában lesznek elvégezve.
17. ábra: a synapsin I immunreaktivitás jelentős csökkenése figyelhető meg az operált oldalon (B) a kontrollhoz képest (A). I.:stratum granulosum, II.: stratum Purkinjense,
III.: stratum moleculareS. (10X-es nagyítás) II. I.
III.
I. II. III.
32
8. Megbeszélés
A c-fos protein expressziójának kimutatása számos előző közlemény tárgya (Szakács et al., 2001; Kovács et al., 2003; Mihály et al., 2005). A konvulzió során a c-fos expresszió jellegzetes dinamikát mutat: a neocortexben és a striatumban a 4- AP kezelést követő első órában maximális (Kovács et al., 2003; Mihály et al., 2005). Az mRNS hasonló dinamika szerint termelődik: a neocortexben a c-fos mRNS-szint szignifikáns növekedése a kezelést követően, 1 órával maximális (Mihály et al., 2005). A c-fos expresszió ionotrop glutamátreceptor antagonistákkal gátolható, ami az ionotrop (főleg az NMDA) glutamátreceptorok szerepét támasztja alá (Szakács et al., 2003).
18. ábra: a nuclei pontis neuronjainak NeuN immunreaktivitása csökken az operált oldalon (nyíl)
33
Kísérleteink célja volt, hogy megvizsgáljuk a 4-AP által kiváltott akut konvulziók kisagyi hatásait. Miután a cerebelláris neuronkörök egyik fő transzmittere a glutaminsav (Ito, 1984), feltételezhető volt a kisagykéreg c-fos expressziójának növekedése. A glutaminsav az afferensek (moharostok és kúszórostok) fő transzmittere. A neuronok közül a szemcsesejtek működnek glutamáttal. Vagyis a cortex cerebelli minden rétegében sok glutamáterg szinapszis található. A masszív glutamáterg afferentáció ellenére azt találtuk, hogy a kisagykéreg c-fos expressziója csak lassan nő: maximumát 3 órával a 4-AP kezelés után érte el. Ez a dinamika lényegesen különbözik a neocorticalis c-fos expresszió dinamikájától: a neocortexben 1 óra elegendő a maximális c-fos expresszió kialakulásához. Ez az időérték minden neocorticalis laminában egyforma: jóllehet az egyes neocortex rétegek afferentációja eltérő (Mihály et al., 2005). A kisagykéreg glutamáterg neuronja a kisagyi szemcsesejt. A kisagyi szemcsesejtek a szintén (túlnyomórészt) glutamáterg moharostok közvetítésével kapnak afferentációt (Ito, 1984). A szemcsesejtek és a kisagyi glomerulusok igen jelentős számban vannak jelen a kisagykéregben: becslések szerint 1 mm3 szövetben 3 millió szemcsesejt és 600,000 glomerulus található (Carpenter, 1991). Nem meglepő tehát, hogy a jelentős c-fos expresszió a kéreg stratum granulosumában mérhető. Kimutattuk, hogy a szemcsesejtek c-fos expressziója a konvulzáló állatokban már 1.5 óra után szignifikáns különbséget mutatott a kontrollhoz képest, de a c-fos immunreaktív sejtmagok száma csak 3 órával a 4-AP kezelés után érte el a maximális értéket. A granuláris réteg c-fos tartalmú sejtjeinek többsége szemcsesejt. A szemcsesejtek c- fos expressziójának lassú dinamikája a gátló neuronok hatásával magyarázható, vagyis a sajátos aktiválódási dinamika magyarázata a kisagykéreg belső neuronköreinek sajátosságaiban rejlik. Az excitatorikus neuronok között nincsenek belső reverberációt segítő szinaptikus kapcsolatok. Ehelyett a glomeruláris szinapszisok vonatkozásában, a Golgi-neuronok felől jövő GABAerg gátlással kell számolnunk. A Golgi-neuron axonja preszinaptikus gátlás formájában szabályozza/gátolja a szemcsesejtek aktiválódását; amely gátlási forma igen hatékony. Miután a Golgi-neuront is a moharost aktiválja, itt egy gátló visszacsatolás érvényesül (Ito, 1984). Ennek alapján kijelenthetjük, hogy az akut konvulziók során a szemcsesejtek működését a moharostok a Golgi-neuronok révén szabályozzák. Miután a Golgi-neuronok becsült száma 1 mm3 szövetben mindössze 50, ez a preszinaptikus gátlás nagy excitatorikus bemenettel ellensúlyozható.
34
Amikor a szemcsesejteket bármelyik afferens pályán át érkező excitáció működteti, akkor valószínűleg moharost szinkronizáció szükséges ahhoz, hogy nagyszámú szemcsesejt egyszerre aktiválódjon, és c-fos expresszió indukálódjon. A moharost szinkronizációt további kísérletekben kell bizonyítani.
A vermis lebenykéinek vizsgálata során megfigyeltük, hogy a különböző lebenyek és lebenykék eltérő aktivációs dinamikát mutatnak. A vermis elülső lebenye a spinocerebellumhoz tartozik. Emellett cuneocerebellaris, cochlearis és vestibulocerebellaris afferenseket is kap (I. lobulus). A hátsó lebeny nagy része a cerebrocerebellumhoz tartozik, kisebb része reticulocerebellaris, cuneocerebellaris (IX. lebenyke) és vestibulocerebellaris afferentációt kap (X. lebenyke). Az I. és a X.
lebenyke a vestibulocerebellum részei, annak ellenére, hogy az egyik a lobus anterior, a másik a lobus flocculonodularis része. Ez a maygarázata annak, hogy a két lobulus c-fos expressziója igen hasonló dinamikát mutat. Irodalmi adatok szerint a lobus posterioron belül a pyramis (VIII.) és az uvula (IX.) hasonló arányban kap afferenseket a nuclei pontis felől (Zguczynski et al., 2010). Ennek megfelelően, c-fos expressziójuk is hasonló dinamikájú. Miután immunhisztokémiai méréseink szerint a kisagykéreg c-fos expressziója az afferentációval mutat korrelációt, elvégeztük az egyik pályarendszer műtéti roncsolását. A MCP roncsolása után kapott eredmények azt bizonyították, hogy az afferensek roncsolása szignifikáns módon csökkenti a granuláris réteg c-fos expresszióját, alátámasztva azt az feltevést, hogy a szemcsesejtek c-fos expresszióját a moharost szinapszisokon érkező (esetleg szinkronizált) excitáció okozza. A szemcsesejt aktiváció idején ECoG segítségével a neocortex felett agykérgi nagy frekvenciájú tüskesorozatok figyelhetők meg (Mihály et al., 2005).
Ez a hiperaktivitás a diencephalonra és a striatumra is átterjed és regisztrálható (Kovács et al., 2003). Úgy gondoljuk, hogy az akut konvulziók során a kisagykéregre is az agykérgi neuronok hiperszinkron aktivációja tevődik át, a tractus corticopontinus – nuclei pontis – tractus pontocerebellaris révén, amely a középső kisagykaron keresztül ingerli a kisagykéreg neuronjait. Az operált állatok kisagyának egészséges oldalán is szignifikánsan csökken a c-fos expresszió a kontrollhoz képest, amely arra utal, hogy a cortico-ponto-cerebellaris pályán érkező excitáció valamilyen arányban kereszteződik a kisagyban is. Ezt a mechanizmust látszik alátámasztani az a megfigyelésünk, miszerint az MCP roncsolása megnöveli
35
a generalizált tónusos-klónusos görcsök látenciáját. Az aktiválódás mechanizmusát későbbi kísérletek során, elektrofiziológiai kísérletekben szeretnénk bizonyítani. Ez a kisagyi aktiváció krónikus epilepsziákban felelős lehet a kórboncolásokban látott kisagyi atrófiáért (Ney et al., 1994). MRI-vel végzett vizsgálatokkal kimutatták a középső kisagykar károsodását krónikus epilepsziás betegekben (Okamoto et al, 2003). A krónikus epilepszia progressziója során megjelenő keresztezett cerebellaris atrophia során a kisagyi károsodás a cortico-ponto-cereballaris pálya közvetítésével történik (Tan and Urich, 1984). A leszálló cortico-cerebellaris rostok funkció mellett a kisagykéregből a thalamuson át a neocortexbe felszálló pályák hatásait is leírták, utóbbiak inhibitoros hatást fejtenek ki a cortex cerebri aktivitására (Middleton and Strick, 1998). Ezen megfigyelések arra engednek követeztetni, hogy nem csupán a cortex tudja befolyásolni a kisagykéreg működését és fokozni a cerebellaris atrophia mértékét epilepszia során, hanem a kisagykéreg is befolyásolja a kéreg működését és szerepe lehet az epilepszia pathomechanizmusában, progressziójában vagy a konvulziók szabályozásában.
További vizsgálataink során arra kerestük a választ, hogy vajon a középső kisagykar léziójának milyen hosszú távú következményei lehetnek a kisagyi és hídi neuronhálózatokra. Régóta ismert, hogy krónikus epilepsziás betegekben gyakran fordul elő kisagyi atrophia, gliosis és sejtpusztulás, ami leginkább a Purkinje- sejteket érinti (Crooks et al., 2000). A neocortex – híd – kisagy – neocortex neuronkörök szerepe még nem tisztázott. Az egyik elmélet szerint az agykérgi neuronok hiperszinkron kisülése és ennek az aktivitásnak áttevődése a kisagykéregre a kisagyi neuronális hiperaktivitás révén sejtpusztulást okoz (Savić and Thorell, 1996). A másik teória szerint a hiperaktivitás miatt akut ödéma jön létre amit a serkentő neurotranszmitterek által nyitva tartott csatornákon keresztül beáramló ionok és víz okoznak (Seitelberger et al., 1990). Vizsgálataink szerint a kisagykéreg hiperaktivitása elhúzódó folyamat, amely egyaránt kedvező a glutamát neurotoxicitás és az elhúzódó agyduzzadás számára. Így mindkét mechanizmussal számolnunk kell a hosszan tartó epilepszia betegség során kialakuló cerebelláris atrófia szempontjából.
Másrészt, a MCP roncsolása bizonyította, hogy az afferensek pusztulása következményes gliózist (ami kisagyi atrófiában is megfigyelhető) okoz. A gliózis mellett jelentős deafferentáció is bekövetkezik, amit a synapsin I immunreaktivitás
36
csökkenése bizonyított. A kisagykéreg atrófiája feltehetően együtt jár a nuclei pontis átrendeződésével és retrográd degeneráció révén, pusztulásával. Ezek a degeneratív változások megszakítják a neocortex – cerebellaris cortex neuronköröket és feltehetően mélyreható módon befolyásolják az epileptogenezist.
Első megfigyelésünk, hogy a műtétet követően folyamatosan emelkedik a kisagykéregben és a hídban az astrocyták száma. A Bergmann-gliosis krónikus epilepsziás betegekben is megfigyelhető, amely a középső kisagykar károsodásának a következménye is lehet, vagy éppen az excitotoxicus neuronpusztulás ütemét fokozza. Az astrocyták segítik a neuronok tápanyaghoz jutását, az anyagcsere végtermékek eliminálását, befolyásolják a neurotranszmissziót a neurotranszmitterek visszavétele és lebontása által. Pusztulásuk fokozza az idegsejtek érzékenységét és fokozza az apoptosisra való hajlamukat egyaránt (Borges et al., 2003). A hídban kialakuló gliosis magyarázata a retrográd neuronpusztulás lehet, amely az axonok átmetszésének következménye. Az axonátmetszés másik következménye az anterográd degeneráció, amely a moharost- terminálisok degenerációját okozza. A folyamatot jelzi a synapsin-I immunreaktivitás nagyfokú csökkenése a stratum granulosumban (Kopniczky et al., 2005). Ezeket a folyamatokat kíséri a reaktív gliosis a híd és a kisagy területén egyaránt. Ezek alapján elmondható, hogy a középső kisagykarnak szerepe van az epilepszia betegség progressziójában. További vizsgálatainkban arra a kérdésre keressük a választ, hogy a kisagyi károsodást valóban a pályarendszeren érkező fokozott excitáció okozza és a betegség előrehaladásának a következménye a kisagykar károsodása vagy éppen ez okozza a kisagyi atrophiát. Ehhez krónikus epilepszia modellre van szükségünk, amelyet operált állatok pilokarpinos kezelésével tudunk elérni.
37
9. Köszönetnyilvánítás
Köszönetemet fejezem ki Dr. Mihály András professzor Úrnak, aki a TDK munkám kezdete óta mentorálta a tevékenységemet és lehetővé tette számomra, hogy megírjam a diplomamunkámat.
Köszönettel tartozom Dr. Kopniczky Zsolt adjunktus Úrnak (Idegsebészeti Klinika, SZTE, ÁOK) a kísérleti állatok operációiban nyújtott igen hasznos gyakorlati tanácsaiért.
Köszönöm Krisztinné Péva Beáta egyetemi tanársegédnőnek (Anatómiai Intézet) az ábrák és diagramok szerkesztésében nyújtott segítségét.
Hálával tartozom Kara Mónikának, Kobolák Andreának és Papp Gabriellának (Anatómiai Intézet) a kísérletekben végzett asszisztensi munkájukért és segítségükért.
A kutatás az Európai Unió és Magyarország támogatásával, az Európai Szociális Alap társfinanszírozásával a TÁMOP 4.2.4A/2-11-1-2012-0001 azonosító számú
„Nemzeti Kiválóság Program-Hazai hallgatói, illetve kutatói személyi támogatást biztosító rendszer kidolgozása és működtetése konvergencia program” című kiemelt projekt keretei között valósult meg.
38
10. Irodalomjegyzék
Adams F. On the sacred disease. In: The genuine works of Hippocrates.
Vol II. London: Sydenham Society;1849:831-58.
Amedee T, Robert A, Coles JA. Potassium homeostasis and glial energy metabolism.
Glia 1997;21:46–55.
Aronica E, Yankaya B, Jansen GH, Leenstra S, van Veelen CW, Gorter JA, Troost D. Ionotropic and metabotropic glutamate receptor protein in glioneuronal tumours from patients withintractable epilepsy.
Neuropathol Appl Neurobiol 2001;27:223–37.
Barres BA (2008) The mystery and magic of glia: a perspective on their roles Bechtel, N., Kobel, M., Penner, K.I., Klarhöfer, M., Scheffler, K., Opwis, K., et al.
Decreased fractional anisotropy in the middle cerebellar peduncle in children with epilepsy and/or attention deficit/hyperactivity disorder: A preliminary study.
Epilepsy and Behavior 2009;15:294-98.
Borges K, Gearing M, McDermott DL, Smith AB, Almonte AG, Wainer BH, Dingledine R. Neuronal and glial pathological changes during epileptogenesis in the mouse pilocarpine model.
Exp Neurol 2003;Jul;182:21-34.
Botez MI, Ezzedine A, Vezina JL. Cerebellar atrophy in epileptic patients.
Neurol. Sci. 1988;15:299-303.
Brodal P, Bjaalie JG. Salient anatomic features of the cortico-ponto-cerebellar pathway.
Brain Res 1979;114:227 – 49.
Brückner C, Heinemann U. Effects of standard anticonvulsant drugs on different patterns of epileptiform discharges induced by 4-aminopyridine in combined entorhinal cortex-hippocampal slices.
Brain Res 2000;859:15-20.
Bushong EA, Martone ME, Jones YZ, Ellisman MH Protoplasmic astrocytes in CA1 stratum radiatum occupy separate anatomical domains.
J Neurosci 2002;22:183–92.
Carpenter MB (1991) Core Text of Neuroanatomy. Williams&Wilkins.
39
Castejón OJ, Fuller E, Dailey ME. Localization of synapsin-I and PSD-95 in developing postnatal rat cerebellar cortex.
Develop Brain Res 2004;150:25-32.
Castillo J, Davalos A, Noya M. Progression of ischaemic stroke and excitotoxic aminoacids.
Lancet 1997;349:79-83.
Christopherson KS, Ullian EM, Stokes CC, Mullowney CE, Hell JW, Agah A, Lawler J, Mosher DF, Bornstein P, Barres BA. Thrombospondins are astrocyte- secreted proteins that promote CNS synaptogenesis.
Cell 2005;120:421–33.
Crooks R, Mitchell T, Thom M. Patterns of cerebellar atrophy in patients with chronic epilepsy: a quantitative neuropathological study.
Epilepsy Res 2000;41:63–73.
Dam M, Wolf P, Janz D, Dreifus FE. Neuropathology of the cerebellum. Advances in Epileptology 1987;16:15-20.
De Marco FA, Ghizoni E, Kobayashi E, Li LM, Cendes F. Cerebellar volume and long-term use of phenytoin.
Seizure 2003;12:312-15.
Dragunow M, Currie RW, Faull RLM, Robertson HA Jansen K. Immediate-early- genes, kindling and long-term potentiation.
Neurosci Behav Rev 1989;24:301-13.
Duffy TE, Howse DC, Plum F. Cerebral energy metabolism during experimental status epilepticus. J Neurochem 1975;24:925–34.
Engel J. Jr. Introduction to temporal lobe epilepsy.
Epilepsy Res 1996;26:141-50.
Fukaya M, Hiroshi U, Yamauchi Y, Inoue Y, Watanabe M, Distinct spatiotemporal expression of mRNAs for the PSD-95/SAP90 protein family in the mouse brain.
Neurosci Res 1999;33:111 –18.
Gass P, Herdegen T, Bravos R, Kiessling M. Induction of immediate early gene encoded protein sin the rat hippocampus after bicuculline-induced seizures:
Differential expression of KROX-24, fos and jun proteins.
Neurosci 1992;48:315-324.
Hagemann G., Everitt AD, Lemieux L, Krakow K, Free SL, Kendall BE, Stevens JM, Duncan JS, Shorvon SD. Volumetric MRI of cerebellar atrophy in a prospective study of a community based population with epilepsy. The 7th Scientific Meeting & Exhibition of the International Society for Magnetic Resonance in Medicine in Philadelphia, Pennsylvania, U.S.A., 24-28 May 1999.
40
Halassa MM, Haydon PG Integrated brain circuits: astrocytic networks modulate neuronal activity and behavior.
Annu Rev Physiol 2010;72:335–55.
Harvey, AS., Jayakar, P., Duchowny, M., Resnick, T., Resnick, T., Prats, A. et al.
Hemifacial seizures and cerebellar ganglioglioma: an epilepsy syndrome of infancy with seizures of cerebellar origin.
Ann Neurol. 1996; 40(1):91-98.
Herdegen T, Waetzig V. AP-1 proteins in the adult brain: Facts and fiction about effectors of neuroprotection and neurodegeneration.
Oncogene 2001;20:2424-37.
Hauser WA. Seizures disorders: the changes with ages.
Epilepsia 1992;33:6-14.
Hermann BP, Bayless K, Hansen R, Parrish J, Seidenberg M. Cerebellar atrophy in temporal lobe epilepsy.
Epilepsy & Behavior 2005;7:279-87.
Hilfiker S, Pieribone VA, Czernik AJ, Kao HT, Augustine GJ, Greengard P.
Synapsins as regulators of neurotransmitter release.
Biol Sci 1999;354:269–79.
Hoffmann GE, Lyo D. Anatomical markers of activity in neuroendocrine systems:
are we all ’fos-ed out’?
J Neuroendocr 2002;14:259-68.
Hossman KA, Niebuhr I, Tamura M. Local cerebral flow and glucose consumption of rats with experimental gliomas.
J Cereb Blood Flow Metab 1982;2:25–32.
Howse DC, Caronna JJ, Duffy TE, Plum F. Cerebral energy metabolism, pH, and blood flow during seizures in the cat.
Am J Physiol 1974;227:1444–51.
International Bureau for Epilepsy, WHO. Epilepsy in the WHO European region:
fostering epilepsy care in Europe. http://www.ibe-epilepsy.org/downloads/EURO Report 160510.pdf (2010)
Ito M. The Cerebellum and Neural Control.
Raven Press New York 1984;pp:14–15.
Jefferys JG, Haas HL. Synchronized bursting of CA1 hippocampal pyramidal cells in the absence of synaptic transmission.
Nature 1982;300:448–50.
41
Jefferys JG. Nonsynaptic modulation of neuronal activity in the brain: electric currents and extracellular ions.
Physiol Rev 1995;75:689–23.
Koh, N.K., Lim, C.B., Hwang, H., Park, D.J., Chae, H.J., Kim, J.K. et al.
Cerebellum can be a possible generator of progressive myoclonus.
Journal of Child Neurology 2010; 25:728
Kopniczky Zs, Dobó E, Borbély S, Világi I, Détári L, Krisztin-Péva B, Bagosi A, Molnár E, Mihály A. Entorhinal cortex lesions rearrange afferents, glutamate receptors, increase seizure latency and supress seizure-induced c-fos expression in the hippocampus of the adult rat.
J Neurochem 2005;95:111-24.
Kovács A, Mihály A, Komáromi A, Gyengési E, Szente M, Weiczner R, Krisztin- Péva B, Szabo Gy, Telegdy Gy. Seizure, neurotransmitter release, and gene expression are closely related in the striatum of 4-aminopyridine-treated rats.
Epilepsy Res 2003;55:117-29.
Kuhnt U, Mihály A, Joo F. Stimulation-dependent calcium binding site in the guinea pig hippocampal slice: an electrophysiological and electron microscope study.
Brain Res 1983;279:19-30.
Labiner DM, Butler LS, Cao Z, Hosford DA, Shin C, McNamara JO. Induction of c-fos mRNA by kindled seizures: complex relationship with neuronal burst firing.
J Neurosci 1993;1:744-51.
Lawson JA, Vogrin S, Bleasel AF, Cook MJ, Bye AM. Cerebral and cerebellar volume reduction in children with intractable epilepsy.
Epilepsia 2000;41:1456-62.
Lemeignan M, Millart H, Lamiable D, Molgo J, Lechat P.Evaluation of 4- aminopyridine and 3,4-diaminopyridine penetrability into cerebrospinal fluid in anesthetized rats.
Brain Res 1984;304:166-69.
Marinelli S, Gatta F, Sagratella S. Effects of GYKI 52466 and some 2,3-
benzodiazepine derivates on hippocampal in vitro basal neuronal excitability and 4- aminopyridine epileptic activity.
Eur J Pharmacol 2000;391:75-80.
Men S, Lee DH, Barron JR, Munoz DG. Selective neuronal necrosis associated with status epilepticus: MR findings.
AJNR 2000;21:1837–40.
42
Middleton FA, Strick PL. Cerebellar output: motor and cognitive channels. Trends Cogn Sci 1988;2:348–54.
Mihály A, Bencsik K, Solymosi T. Naltrexone potentiates 4-aminopyridine seizures in the rat.
J Neural Transmiss 1990;79:59-67.
Mihály A, Shihab-Eldeen A, Owunwanne A, Gopinath S, Ayesha A, Mathew M.
Acute 4-aminopyridine seizures increase the regional cerebral blood flow in the thalamus and neocortex, but not in the entire allocortex of the mouse brain.
Acta Physiol 2000;87:43-52.
Mihály A, Borbély S, Világi I, Détári L, Weiczner R, Zádor Z, Krisztin-Péva B, Bagosi A, KOpniczky Z, Zádor E. Neocortical c-fos mRNA transcription in repeated, breaf, acute seizures: Is c-fos a coincidence detector?
Internat J Mol Med 2005;15:481-86.
Mihály M, Szente Zs, Dubravcsik B, Boda E, Király T, Nagy A, Domonkos.
Parvalbumin- and calbindin-containing neurons express c-fos protein in primary and secondary (mirror) epileptic foci of the rat neocortex.
Brain Res 1997;761:135-45.
Mink, J.W., Caruso, P.A., Pomeroy, S.L. Progressive myoclonus in a child with a deep cerebellar mass.
Neurology. 2003;61(6):829-831.
Mullen RJ, Buck CR, Smith AM. NeuN, a neuronal specific nuclear protein in vertebrates.
Development 1992;116:201-11.
Mumenthaler, Marco:Neurológia, 4. kiadás Medicina Könyvkiadó Rt. 1996.
Ney GC, Lantos G, Barr WB, Schaul N. Cerebellar atrophy in patients with long term phenytoin exposure and epilepsy.
Arch Neurol 1 9 9 4 ; 51:767 – 71.
Nikl J. Questions of epileptogenesis and prevention in symptomatic epilepsies.
Clin Neurosci 2004;57:164-73.
Okamoto K, Tokiguchi S, Furusawa T, Ishikava K, Quardery FA, Shinbo S, Sasai K. MR Features of deseases involving bilateral middle cerebellar peduncles.
American J Neurorad 2003;24:1946-54.
Oliet SH, Piet R, Poulain DA Control of glutamate clearance and synaptic efficacy by glial coverage of neurons.
Science 2001;292:923–26.