VESZPRÉMI EGYETEM
*(25*,.210(= *$='$6È*78'20È1<,.$5
Állattudományi Intézet
ÁLLATTENYÉSZTÉSTANI TANSZÉK
Készült a Veszprémi Egyetem Állattenyésztési Tudományok Doktori Iskola keretében
7pPDYH]HW
Dr. Kovács József professor emeritus
D0H] JD]GDViJWXGomány kandidátusa
.h/g1%g= )$-7È-Ò+Ë=Ï6(57e6(.
1g9(.('e6,5,7086-(//(0= ,1(.
ELEMZÉSE
Készítette
DEÁK TAMÁS
KESZTHELY 2003
.h/g1%g= )$-7È-Ò+Ë=Ï6(57e6(.
1g9(.('e6,5,7086-(//(0= ,1(.
ELEMZÉSE
Írta
DEÁK TAMÁS
Készült a Veszprémi Egyetem Állattenyésztési Tudományok Doktori Iskola keretében
7pPDYH]HW 'U.RYiFV-y]VHISURIHVVRUHPHULWXV D0H] JD]GDViJWXGRPiQ\NDQGLGiWXVD
Elfogadásra javaslom igen / nem
……….
Az értekezést bírálóként elfogadásra javaslom igen / nem
(OV EtUiOy Dr. Csató László egyetemi docens mg. tud. kandidátusa igen / nem
……….
Második bíráló: Dr. Vígh László Ph. D. igen / nem
……….
TARTALOMJEGYZÉK
1. BEVEZETÉS 11
2. SZAKIRODALMI ÁTTEKINTÉS 14
2.1. A sertés növekedése 14
2.2 A sertés növekedésének modellezése 20 2.3. Fejlettség a hizlalás kezdetén 23 2.4. Kiegyenlítettség a hízóállomány fejlettségében 27 2.5. Növekedés -K~VPHQQ\LVpJYiJyiOODWPLQ VpJ
a hús zsírtartalma, zsírsavak 31
2.6. Növekedés -K~VPHQQ\LVpJK~VPLQ VpJ
DK~V]VtUWDUWDOPD]VtUPLQ VpJNROHV]WHULQ 33
3. ANYAG ÉS MÓDSZER 38
3.1.1. A kísérleti állatok tartási, takarmányozási körülményei 38
3.1.2. A növekedési ritmus elemzése 39
3.1.3. Testméretek, testarányok 42
3.1.4. Vágóérték vizsgálatok 43
3.1.5. Biometriai értékelés 44
3.2. A hízóba állítási fejlettség vizsgálati módszere 44 3.3. A hízóállomány kiegyenlítettségének vizsgálati módszere 46
4. EREDMÉNYEK ÉS ÉRTÉKELÉSÜK 52
4.1. Növekedési ritmus jelleP] NHOHP]pVH 52 4.1.1. Hízékonysági tulajdonságok elemzése 52 4.1.1.1. Magyar nagyfehér hússertések hízékonysági
teljesítményeinek elemzése 55
4.1.1.2. Magyar lapálysertések hízékonysági
teljesítményeinek elemzése 65
4.1.1.3. Magyar nagyfehér hússertés x magyar lapálysertés
NHUHV]WH]pV Kt]yVHUWpVHNKt]pNRQ\ViJL
teljesítményeinek elemzése 75
4.1.1.4. H1 hibridsertések hízékonysági
teljesítményeinek elemzése 81
4.1.1.5. H2 hibridsertések hízékonysági
teljesítményeinek elemzése 87
0DQJDOLFD[SLHWUDLQNHUHV]WH]pV
hízósertések hízékonysági teljesítményeinek elemzése 94 4.1.2. Testméretek, testarányváltozások 100 4.1.3. Egyes vágóérték mutatók alakulása a hizlalás alatt 113 4.2. A hizlalás kezdeti fejlettség és a hizlalás alatti növekedés 123
$Kt]yEDiOOtWiVNRULIHMOHWWVpJMHOHQW VpJH 123 4.2.2. A hizlalás alatti növekedés és kiegyenlítettség 132
.O|QE|] Q|YHNHGpVLLQWHQ]LWiV~JHQRWtSXVRNK~ViQDN
zsírtartalma és zsírsavösszetétele 153
4.4 .O|QE|] Q|YHNHGpVLLQWHQ]LWiV~JHQRWtSXVRNK~ViQDN
zsírtartalma és koleszterintartalma 161
5. KÖVETKEZTETÉSEK, JAVASLATOK 169
6. ÚJ KUTATÁSI EREDMÉNYEK 175
7. ÖSSZEFOGLALÁS 178
8. SUMMARY 186
KÖSZÖNETNYILVÁNÍTÁS
9. PUBLIKÁCIÓS JEGYZÉK 194
9.1. Folyóiratban idegen nyelven megjelent cikkek és közlemények 9.2. Folyóiratban magyar nyelven megjelent cikkek és közlemények 9.3. Konferencia kiadványokban megjelent anyagok idegen nyelven 9.4.Konferencia kiadványokban megjelent anyagok magyar nyelven
10. IRODALOMJEGYZÉK 197
11. MELLÉKLETEK 206
KIVONAT
.h/g1%g= )$-7È-Ò+Ë=Ï6(57e6(.1g9(.('e6,5,7086 -(//(0= ,1(.(/(0=e6(
$ V]HU] D NRUV]HU YiJyVHUWpV HO iOOtWiVEDQ UpV]WYHY VHUWpV
genotípusok növekedésdinamikai elemzését végezte. Célja, hogy a hizlalási és vágási teljesítmények kapcsolatát ilyen szempontból jellemezze. A beállítási súly eltéréseinek hatását sokoldalúan feldolgozta. Két fajtatiszta, magyar nagyfehér hússertés és magyar lapálysertés, két keresztezés, magyar nagyfehér hússertés x magyar lapálysertés és mangalica x pietrain, valamint két hibrid, hibrid 1 és hibrid 2 genotípusokkal, összesen 406 hízósertéssel végezte vizsgálatát.
A hízékonysági és vágási paraméterek mellett a testarányváltozásokat szintén megállapította. Kimunkálta az életszakaszonkénti növekedési
NLWHY NHWDV]HUYHVQ|YHNHGpVMHOOHP]pVpUHgVV]HIJJpVYL]VJiODWRNNDO
feltárta a hízékonysági, vágási paraméterek közötti kapcsolatok irányát
pVPpUWpNpW$PXQNiEyONLW QLNKRJ\JHQRWtSXVRQNpQWHOWpU PpUWpN
és irányú kapcsolatok vannak a legfontosabb tulajdonságok
WHNLQWHWpEHQ $ K~VPLQ VpJL YL]VJiODWRNDW NLE YtWHWWH D]
intramuscularis zsír zsírsavösszetételére nézve is. Kiszámította a zsírsavarányok mutatóit. A sertéshús koleszterintartalmának meghatározása is helyet kapott kutatásában. Eredményei azt jelzik, hogy a hajtatott hízóalapanyag-HO iOOtWiV W~O PDJDV EHiOOtWiVL pO V~O\
NHGYH] WOHQYiJiVLPLQ VtWpVWSURGXNiO
ABSTRACT
ANALYSE OF GROWTH DYNAMIC OF DIFFERENT PIG BREEDS
The author analysed the growth dynamic of pig genotypes used in modern fattening to produce slaughtering pig. The aim of the investigation was to characterize the connections of fattening and slaughtering performances in this respect. The effect of differences of starting weight was carried out in several ways. As many as 406 fattening pigs were investigated in Hungarian Large White and Hungarian Landrace breeds, in Hungarian Large White x Hungarian Landrace and Mangalitsa x Pietrain crosses as well as in Hybrid 1 and Hybrid 2 genetic construction. Besides fattening and slaughtering performances the changes of different body regions were also shown.
The exponent of growth in life periods was worked out to characterize the organic growth. Correlations between fattening and slaughtering performances were calculated. It is stated that there were different correlations between the most important parameters. Investigation of meat quality also contained the fatty acid composition of intramuscular
DIE ANALYSE DER ZUNAHMEDYNAMIK VERSCHIEDENE SCHWEINERASSEN
Der Autor hat die Zunahmedynamik der Schweinerasse analysiert, die man benutzt Schlachtschwein zu herstellen. In dieser Rücksicht war das Ziel der Untersuchung, Beziehungen der Mastsleistungen und der Schlachtsleistungen zu charakterisieren. Man hat die Wirkung des Unterscriedes des Anfansgewichts ausgearbeitet. Man hat 406 Stück Mastschweine untersucht in der Rassen Ungarisches Großweiß x Ungarisches Landrace, Mangaliza x Pietrain, und in genetischer Konstruktion Hybrid 1 und Hybrid 2. Sie haben außen der Mastsleitungen und Schlachtsleistungen die Änderungen der verschiedenen Körpergegend auch demonstriert. Der Exponent der Zunahme war in der Lebensperiode ausgearbeitet, die organische Zunahme zu charakterisieren. Die Korrelationen waren zwischen den Mastleistungen und den Schlachtsleistungen erklärt. Man hat bestimmt, dasses verschiedene Korrelationen zwischen den wichtigsten Parametern gab. Die Untersuchung der Fleischqualität hat die Fettsäurezusammensetzung des intramusculären Fettes auch enthaltet.
Die Ergebnisse haben schlehter Schlachtsqualität bezeichnet als, wenn die Herstellung des Grundes der Mästung zu intensiv war (Zu groß Startgewicht).
1. BEVEZETÉS ÉS&e/.,7 =e6
A sikeres sertéshizlalás a mennyiségi teljesítmények mellett határozott
PLQ VpJLN|YHWHOPpQ\HNHWLVWiPDV]W$PLQ VpJMDYtWiVOHKHW VpJHLQHN
kutatása, a biológiai összefüggések elemzése, következtetések levonása
OHKHW VpJHWDGDIDMWD pVWtSXV különbségek biológiai okainak jelzésére.
(]pUW LQGRNROW D QHYHOpVL LG V]DN EHIHMH]WpYHO NH]G G VHUWpVKL]ODOiV IRO\DPDWiEDQ D VHUWpV Q|YHNHGpVpQHN IHMO GpVpQHN ELROyJLDL
változásait nyomon követni. A növekedés a szervezet méret- és súlybeli változását jeOHQWL PtJ IHMO GpVNpQW D KRUPRQiOLV EHIRO\iV DODWW YpJEHPHQ V]|YHWL V]HUYL GLIIHUHQFLiOyGiV KDWiUR]KDWy PHJ $ NpW
folyamat egymással összefonódik és egymástól el nem választható. A
IHODGDW PHJROGiViQDN HOV OpSpVHNpQW D GROJR]DWEDQ V]HUHSO
növekedési ULWPXV MHOOHP] N HOHP]pVH MHO|OKHW PHJ $ NO|QE|]
IDMWiNUD MHOOHP] HOWpU Q|YHNHGpVGLQDPLNDL VDMiWRVViJRN HOHP]pVH D KL]ODOiV HUHGPpQ\HVVpJH V]HPSRQWMiEyO G|QW IRQWRVViJ~ $ NDSRWW HUHGPpQ\HNXJ\DQDNNRUOHKHW YpWHV]LNDV]yEDQIRUJyHOWpU ELROyJLDi sajátosságok céltudatos figyelembe vételét mind a tenyésztési munkában, mind a hizlalási gyakorlatban.
növekedési szakaszokra bontva képet kapjak a hízósertések súlyának, valamint egyéb hizlalási és vágási teljesítményének, vágási
PLQ VpJpQHNUpV]OHWHVDODNXOiViUyO
A sertés súlyának és testméreteinek növekedését az egyes termelési szakaszok elején tapasztalható fejlettség függvényében elemzem.
Éppen úgy, ahogy a születési súly (fejlettség) életre szóló kihatásait vizsgálják, tisztázni kívánom, hogy a hizlalás kezdeti súly (fejlettség) milyen hatást gyakorol a hízékonysági paraméterekre. A hízóbaállítás kori fejlettség befolyása ugyanis igen összetett módon érvényesülhet a hízékonysági eredmények és a vágási K~VPLQ VpJ DODNtWiViEDQ $
mutatók sokszor ellentmondásos eredményekben juthatnak érvényre.
A növekedési, súlygyarapodási paraméterek fajtánkénti,
JHQRWtSXVRQNpQWL HOWpUpVHLQHN YL]VJiODWiQ W~O D YiJiVL pV K~VPLQ VpJL
tulajdonságokat is részletesen értékelem. Ennek a kérdésnek a
WLV]Wi]iViUD D J\DNRUODWEDQ HOOHQ U]|WW YiJypUWpN pV K~VPLQ VpJL pUWpNHN WDQXOPiQ\R]iViQ W~OPHQ HQ QDSMDLQN KXPiQHJpV]VpJJ\pEHQ
fontos zsírsavak és koleszterin vizsgálatára is kiterjesztettem kutatóunkámat. Az említett analízisek ~MV]HU VpJpW D] DGMD KRJ\ D
dolgozatban a sertéshús (karaj) intramuscularis zsírjának jellemzése szerepel, a már többször analizált hátszalonnából nyert zsír vizsgálata helyett.
.XWDWyPXQNiPDN|YHWNH] WpPDN|U|NYL]VJiODWiUDWHUMHGNL
.O|QE|] VHUWpVJHQRWtSXVRNQ|YHNHGpVpYHO|VV]HIJJ V]DNDV]RVViJ
elemzése, a szakaszok hosszának és a növekedés sebességének
megállapítása. A testsúlynövekedéssel együtt járó testarányváltozások genotípusonkénti jellegzetességeinek ábrázolása. Ultrahangos mérések segítségével a vágási paraméterek hizlalás alatti változásának bemutatása.
A hízóbaállítási (80 napos) fejlettség és a hizlalás alatti növekedés,
YDODPLQW D NO|QE|] Kt]pNRQ\ViJL pV YiJiVL SDUDPpWHUHN
kapcsolatainak vizsgálata.
A hízóállomány hízóbaállításkori és hizlalás alatti kiegyenlítettsége közötti kapcsolatok elemzése.
.O|QE|] Q|YHNHGpVGLQDPLNiM~ VHUWpVJHQRWtSXVRN K~ViQDN ]VtUVDY|VV]HWpWHOHpVDK~VPLQ VpJYL]VJiODWD
(OWpU Q|YHNHGpVL WHPHW IHOPXWDWy VHUWpVJHQRWtSXVRN K~ViQDN
koleszterintartalma pVDK~VPLQ VpJYL]VJiODWD
2. SZAKIRODALMI ÁTTEKINTÉS
2.1. A sertés növekedése
A sertések növekedése folyamatos, de nem egyenletes SMIDTH (1995) megállapítja, hogy a sertések életkorának változását a növekedés gyorsaságának, intenzitásának változása kíséri. A növekedés
J\RUVDViJiW OHJHJ\V]HU EEHQ D QDSL iWODJRV WHVWW|PHJ J\DUDSRGiVVDO OHKHW MHOOHPH]QL (] D V]OHWpVW O NH]GYH Q|YHNV]LN PDMG VWDJQiO LOOHWYH NpV EE NLV PpUWpNEHQ IRNR]DWRVDQ FV|NNHQ (] D] LYDUpUpV LGHMpUH WHKHW DPHO\ IDMWiQNpQW pVgenotípusonként változóan 70-90 kg-os testtömegnél következik be. A növekedés során a súlygyarapodás
|VV]HWpWHOHLVYiOWR]LN$]pOHWNRUHO UHKDODGWiYDOIRNR]DWRVDQFV|NNHQ D Yt] D] iVYiQ\L DQ\DJ pV D IHKpUMH WDUWDORP GH iOODQGyDQ Q D
szervezet zsírtartalmának aránya. A növekedési intenzitás (súlygyarapodás) csökkenése két okkal magyarázható. Az egyik a
JHQHWLNDL DODS DPHO\ HOV GOHJHVHQ PHJKDWiUR]]D D Q|YHNHGpV
sebességét, a másik ok pedig, hogy a súlygyarapodás összetétele megváltozik.
A genetikai alaS MHOHQW VpJpUH KtYMD IHO D ILJ\HOPHWCSATÓ nyomán HORN (2000), amikor megállapítja, hogy a nemesítés során kialakított
HOWpU JHQRWtSXV~ iOORPiQ\RNDW IDMWiN KLEULGYRQDODN D Q|YHNHGpV LQWHQ]LWiViUD pV WDUWDPiUD YRQDWNR]y MHOHQW V NO|QEVpJHN MHOOHP]LN
amelyek meghatározzák az állományok növekedési kapacitását. A
JHQHWLNDL PHJKDWiUR]RWWViJ MHOHQW V D Q|YHNHGpVL HUpO\ iWODJRV QDSL V~O\J\DUDSRGiVUD YRQDWNR]y |U|N|OKHW VpJL pUWpNV]iPD pV N|]|WW YiOWR]LN $ NO|QE|] JHQRWtSXVRNUD MHOOHP] HOWpU
hústermelési kapacitás meghatározza a fehérje beépítési képességet
minden életkorban és testtömeg tartományban. A fehérjeépítés és a
V]tQK~VQ|YHNHGpVN|]|WWLLJHQHU V|VV]HIJJpVUHKtYMDIHODILJ\HOPHW
PETTIGREW (QQHN D NDSFVRODWQDN PHJIHOHO HQ 1 g fehérje növekedés 2,55 g zsírmentes színhúsnövekedést eredményez. De a növekedés fehérjeigényének kielégítése céljából tudnunk kell, hogy hogyan alakul a fehérjeépítés, vagy színhúsgyarapodás a növekedés szakaszaiban. A növekedés ütemének becsléséhez növekedési görbék
HOHP]pVpW MDYDVROMD D]RN MHOOHJ]HWHV HONO|QO PHUHGHNVpJL
szakaszainak megállapításával egyetemben. DOORNENBAL és TONG (1981), SUSENBETH és KEITEL (1987) nyomán HOLLIS (1993) eredményei alapján a testsúly és testfehérje között megközeOtW HQ D]
DOiEELNDSFVRODWEHFVOKHW
Testsúly (kg) 22 38 65 85 110
Testfehérje (kg) 3,9 6,2 10,6 13,0 16,6
A testsúly és az izomtömeg közötti kapcsolat állandónak és
SiUKX]DPRVQDN WHNLQWKHW NOVAKOFSKI és CUSHER nyomán HOLLIS (1993) szerint irányszámként megadható, hogy egy190 napos állat testének 40 %-a izomállomány.
háromhetenként testsúlymérlegeléseket, valamint ultrahangos szalonna-
pV L]RPYDVWDJViJ PpUpVHNHW WDUW FpOV]HU QHN 8J\DQDNNRU D
metabolizálható energia /ME/ szükséglet megállapításához naponkénti takartmányfogyasztás-mérések elvégzését javasolja. A növekedés ütemének változása természetesen a táplálóanyag szükségletben is viOWR]iVRNDWLGp]HO
CAMPBELL (1997) megállapítja, hogy a sertések aminosav szükséglete gyorsan változik 63 és 110 napos kor között. Az esszenciális aminosavak közül a lizinszükséglet kielégítése – különösen a növekedés maximumánál – 63 és 110 napos kor között lényeges szempont, mert 85 és 112 nap között a súlygyarapodás ütemében már csökkenés mutatkozik. Az energiaszükséglet tekintetében 63 és 98
QDSRV NRU N|]|WW PpUKHW D OHJQDJ\REE PpUWpN Q|YHNHGpV
Hozzáteszi, hogy csoportos tartásban az extra energiaellátás nagyobb növekedést eredményez. Hangsúlyozza a 70 és 110 kg-os súlyhatárok közötti életszakasz növekedésdinamikai vizsgálatát, mert véleménye
V]HULQW D PRGHUQ IDMWiNQiO D WiSOiOyDQ\DJ V]NVpJHW OpSFV ]HWHVHEEHQ
változik, mint azt korábban tapasztaOWiN 5iDGiVXO D NO|QE|]
genotípusok genetikai kapacitását is figyelembe kell venni. CLOSE
XJ\DQH]WD]DODSHOYHWKHO\H]LHO WpUEHDPLNRUNLMHOHQWLKRJ\ D QDJ\ Q|YHNHGpVL HUHM iOODWRN QDJ\REE PHQQ\LVpJ V]tQK~VW NpSHVHN
beépíteni, de nagyobb az aminosav felvételük is. Ha egy állat már elérte
D V]tQK~VpStW JHQHWLNDL NpSHVVpJpQHN PD[LPXPiW D WRYiEEL
súlygyarapodás megnövekedett zsírlerakódást eredményez, ami csökkenti a színhús %-ot. Tehát fontosnak tekinti a genotípusokra
MHOOHP] JHQHWLNDLV]tQK~VWHUPHO NpSHVVpJPHJiOODStWiViW
KOVÁCS NLV pV QDJ\ Q|YHNHGpVL HUpO\ Kt]yVHUWpVHN Kt]ODOiVL
eredményeit hasonlította össze. Megállapítja, hogy azonos vágósúlyban a növekedés dinamikáját a vágási életkor kifejezetten mutatja. Ennek
PHJIHOHO HQ D QDJ\ Q|YHNHGpVL HUpO\ iOODWRN NRUiEEDQ HOpULN D
vágósúlyt és így 30-40 napos eltérés is tapasztalható a vágási életkorban. Úgy találta 105 kg-LJ W|UWpQ HJ\HGL KL]ODOiVEDQ KRJ\ D
takarmányfogyasztás és takarmányértékesítés kapcsolatának szorossága mérséNO GLN D NLV Q|YHNHGpVL LQWHQ]LWiV KDWiViUD $ YiJypUWpN
alakításában a takarmányfogyasztás meghatározó a növekedési intenzitástól függetlenül. Értékelésében kimutatta, hogy a hizlalás kezdeti, nagyobb növekedési intenzitás jobb hízékonysági mutatókat eredPpQ\H] GH D YiJiVL SDUDPpWHUHNEHQ NHGYH] WOHQ LUiQ\~
YiOWR]iVRNDWRNR]$JHQHWLNDLKDWiVMHOHQW VpJpUHKtYMDIHODILJ\HOPHW
a növekedési erély vonatkozásában az NRC (l998). Ennek alapján a hízók képesek kompenzálni az energia-felvételüket, tehát a takarPiQ\IRJ\DV]WiVXNDW D WDNDUPiQ\ HQHUJLDWDUWDOPiWyO IJJ HQ pV
így a genetikai alapban meghatározott növekedési erély ad libitum
WDNDUPiQ\R]iV VRUiQ NO|QE|] HQHUJLDWDUWDOP~ WDNDUPiQ\RN HVHWpQ
bizonyos határok között érvényre jut. CUNHA (1977) a sertés táplálóanyag-V]NVpJOHWpW PHJKDWiUR]y WpQ\H] N N|]|WW HPOtWL D
gyorsabb növekedést. A növekedés intenzitását célzó tenyésztés
Ugyanezt az alapelvet hangsúlyozza WEBB (1997) amikor leszögezi, hogy a takarmányozási módszert az állat genetikai képességéhez kell igazítani az egész termelési folyamatban. A modern sertésfajták és genotípusok a színhúsépítés maximális arányát korában érik el, mint az
MHOOHP] YROW pYYHO NRUiEEDQ ËJ\ NO|Q|VHn a növekedés korai stádiumában fennáll az alultakarmányozás veszélye. Az intenzívebb
V~O\J\DUDSRGiV V]HUHSpUH YLOiJtWDQDN Ui pUWpNHO PXQNiMXNEDQ &2/(
pV 9$5/(< gWH]HU VHUWpVUH NLWHUMHG DGDWHOHP]pVN DODSMiQ
megállapítják, hogy a választástól a hL]yEDiOOtWiVLJ WHUMHG LG V]DNEDQ
100 grammos napi súlygyarapodás többlet a hizlalásban 147 g/nap többlet súlygyarapodást jelentett. A 3-400 g/napos súlygyarapodás különbségek már közel 12 nappal rövidebb vágási életkort eredményezhetnek.
Az intenzívebb növekedés a takarmányozási módszerek átgondolására
|V]W|Q|](':$5'6V]HULQWDPRGHUQJHQRWtSXVRNUDMHOOHP]
gyors növekedés következménye, hogy nehéz meghatározni, és optimálisan kielégíteni a táplálóanyag-szükségletet. Így a fiatal állatokat a kondtFLyMXNQDN PHJIHOHO HQ WDNDUPiQ\R]]iN (QQHN
következtében könnyen lecsökkenhet az izomépítés és megnövekedhet
D]VtUpStWpVWHPH'HDV]tQK~VWHUPHOpVUHW|UWpQ V]HOHNFLyHUHGPpQ\H
a kis étvágy is lehet, ami pedig meggátolja a sertést a növekedés ütemének fenntartásában. HENRY (2000) kísérletében megállapította,
KRJ\ NO|QE|] HQHUJLDWDUWDOP~ WDNDUPiQ\RN HWHWpVHNRU D] DODFVRQ\
energiatartalom hatását csak az egyedileg tartott sertések tudták kompenzálni több takarmány felvételével. 18 és 40 kg között a takarmány alacsony energiatartalma miatti takarmányfelvétel növekedés 40 %-os is lehet. Csoportos tartásban azonban ez nem
következik be, így a sertések nem képesek a genetikailag meghatározott súlygyarapodás huzamos fenntartására.
A takarmányozási módszer k|UOWHNLQW PHJYiODV]WiViUD KtYMD IHO D
figyelmet KUTAS nyomán SZABÓ (1984). Az izomszövetet alkotó izomrostok száma kizárólag a méhen belüli életben növekszik (hyperplasia), ez a folyamat takarmányozással nem befolyásolható. Az izom méretének növekedése a IHMO GpVVHO SiUKX]DPRVDQ YpJEHPHQ L]RPURVW PHJQDJ\REERGiVQDN K\SHUWURSK\D N|V]|QKHW DPHO\
takarmányozással nagymértékben alakítható. Így fontossá válik az optimális takarmány és fehérje ellátás, amelyek az izomrost
NLIHMO GpVpQHN IHOWpWHOpW NpSH]LN A túlzott takarmányfelvétel gyors
Q|YHNHGpVWHUHGPpQ\H] pVQHPiOOUHQGHONH]pVUHHOpJLG DPD[LPiOLV L]RPPpUHW NLIHMO GpVpKH] $ W~O NHYpV WDNDUPiQ\ YLV]RQW ODVV~EE
növekedést és kisebb húsarányt eredményez. Ugyanakkor ENSMINGER és PARKER (1984) szerint a növekedés kapcsolatban van a korral, a napi takarmányfelvétellel, a napi súlygyarapodással és a takarmányértékesítéssel. Alapelvként fogalmazzák meg, hogy a táplálóanyag-szükséglet megállapításának alapja a növekedés matematikai görbéje legyen, de a fajták közötti, a növekedés során mutatkozó különbségek megállapítása is fontos szempontot képvisel.
az egyéb mutatókban is módosulásokat eredményez. A BRANDON RESEARCH CENTRE kutatói (1996) összehasonlító adatértékelésükben megállapították, hogy a nagyobb súlygyarapodásra irányuló szelekció növekedést okoz a hasított felek hosszúságában is.
Ennek mértéke yorkshire sertések vonatkozásában átlagosan 2,9 %-ot
WHWW NL ),('/(5 pV PWVDL LV D V]HOHNFLy MHOHQW VpJpW
hangsúlyozzák adatelemzésük alapján a zsírépítés és a vágóérték tekintetében. Nagyfehér hússertések 30-100 kg közötti ivadékvizsgálatának eredményeit értékelve az alábbi korrelációs együtthatókat kapták:
Karajkereszt-
metszet Színhús % Hátszalonna vastagság Átlagos napi
súlygyarapodás -0,17 +0,29 +0,04
Karajkeresztmetszet +0,56 -0,47
Színhús % -0,66
2.2. A sertés növekedésének modellezése
A növekedésEHQ /$:5(1&( pV PWVD V]HULQW NpW HU KDW HOOHQWpWHVHQ $] HJ\LN D J\RUVtWy HU D PiVLN D] HOOHQV~O\R]y HU
amely lefékezi a szervezet növekedését, valamint befolyásolja a takarmány felvételt azért, hogy az lépést tudjon tartani a test növekedésével. A kifejlett kori súly felé közeledve a növekedés lelassul, aminek a takarmányfelvétel-csökkenés a következménye. A
Q|YHNHGpVW OHtUy HJ\HQOHW OHKHW Yp WHV]L D Q|YHNHGpVL V]DNDV]RN
Q\RPRQ N|YHWpVpW YDJ\ EL]RQ\RV V]DNDV]RNEDQ D N|YHWNH]
növekedési intervDOOXPRNUDYRQDWNR]yHO UHMHO]pVHNPHJKR]DWDOiW
A növekedés folyamatát a BRODY-féle képlet alkalmazásával írják le
(]D]HJ\HQO VpJ
W=Aekt , ahol
: D]pO V~O\WHUPpV]HWHVORJDULWPXVD W LG
A= a W természetes logaritmusa, ha t=0,
k= konstans, amely a folyamatos relatív növekedési arányt mutatja.
BLACK nyomán MOUGHAN és mtsai (1995) szerint dinamikus
PDWHPDWLNDL PRGHOO DONDOPDV YDODPHO\ LG EHOL YiOWR]iV OHtUiViUD $ PRGHOOEHQ HOYpJ]HWW V]iPtWiVL P YHOHWHN PHJKDWiUR]RWW LG V]DNRNUD
vonatkoznak, amHO\HNQHN D YpJV iOODSRWD OHV] D N|YHWNH]
LQWHUYDOOXP N|YHWNH] iOODSRWD $ VHUWpV Q|YHNHGpVL PRGHOOHNEHQ D OHJJ\DNUDEEDQ KDV]QiOW LG V]DNDV] D] HJ\ QDS GH OHKHW HQQpO W|EE YDJ\ NHYHVHEE LV D YL]VJiOW WXODMGRQViJWyO IJJ HQ $] LG V]DNRN
nagysága – melyekben például egy meghatározott állandó növekedési sebesség érvényes – a vizsgált mutatók változásának a mértéke alapján határozható meg. A növekedési sebesség jellemzésével kapcsolatban
növekedése a csontváz növekedésének befejeztével az ivarérettség elérése idején nagymértékben lelassul.
BLACK (1999) a sertés növekedésének jellemzésére dinamikus matematikai modell használatát tDUWMD FpOV]HU QHN ( PyGV]HU
segítségével a takarmányfogyasztás, a takarmány értékesítés
HO UHMHO]pVpW OiWMD PHJYDOyVtWKDWyQDN YDODPLQW D] VHJtWKHW D
táplálóanyag-szükséglet meghatározásában is. POND és MANER (1984) szerint a szerves növekedést leíró függYpQ\ DODNMiW MHOHQW VHQ EHIRO\iVROMD D JHQHWLNDL KiWWpU pV D N|UQ\H]HW KDWiVD (WW O IJJHWOHQO
már a BRODY (1966) által meghatározott „S” alakú görbe használatát tartják alkalmasnak a növekedés jellemzésére. Ezt támasztja alá KAKUK és SCHMIDT (1988) amikor megállapítják, hogy a testsúly változása a fogamzástól a teljes, kifejlett kori testtömeg eléréséig egy
Ä6´ DODN~ ~J\QHYH]HWW V]LJPRLG J|UEpYHO MHOOHPH]KHW (] D J|UEH D
kezdeti állapottól viszonylag lassú emelkedés után egyre meredekebbé válik, majd az ivarérés után ellaposodik. A kifejlett kori testtömeget és
D ILDWDONRUL IHMO GpV WHPpW D JHQHWLNDL NRQVWUXNFLy KDWiUR]]D PHJ GH D]|U|NOHWHVDODSPHJQ\LOYiQXOiViWNOV WpQ\H] NLVEHIRO\iVROMiN
KOVÁCS nyomán KOVÁCS (1984) felhívja a figyelmet arra, hogy a sertés növekedése az élete folyamán nem egyenletes, ezért a növekedés elemzésekor jelezni kell az értékelés határértékeit. Megállapítja, hogy a
Q|YHNHGpVUH KDWy WpQ\H] N N|]O MHOHQW V D IDMWDEHOL KRYDWDUWR]iV
Ennek alapján vizsgálati szemponttá váOLN D NO|QE|] JHQRWtSXVRNUD MHOOHP] pV D] D]RN N|]|WW PXWDWNR]y Q|YHNHGpVL ULWPXV HOWpUpVHN
megállapítása.
2.3. Fejlettség a hizlalás kezdetén
Számos esetben vitatott kérdés, hogy a hajtatott malacnevelés
HUHGPpQ\HNpQW MHOHQWNH] HU WHOMHV Q|YHNHGpV Piként érvényesül a
KL]ODOiV HUHGPpQ\HVVpJpQHN DODNtWiViEDQ )HOW Q pO V~O\NO|QEVpJHNNHO Kt]yED iOOtWRWW HJ\HGHN WHOMHVtWPpQ\PXWDWyLQDN
N|UOWHNLQW pUWpNHOpVH PLQG D WHV]WiOORPiVRN PLQG D J\DNRUODWL
sertéshizlalás szempontjából hasznos tájékoztatást nyújt a gazdaságos termeléshez.
6]iPRV NO|QOHJHV PLQ VpJ PDODFWiSV]HU WDOiOKDWy D NHUHVNHGHOPL
forgalomban, melyek alkalmazásával, a szokvány abrakkeverékkel
QHYHOWPDODFRNURYiViUDD]RQRVpOHWNRUUDV]HPEHW Q V~O\NO|QEVpJHN
alakulnak ki az egyedek között. Természetesen az egyedi, biológiai
NO|QEVpJHNQ|YHNHGpVLHUpO\KDVRQOypO V~O\-differenciákat idéznek
HO D PDODFQHYHOpV VRUiQ .XWDWyPXQNiP VRUiQ H MHOHQVpJ WRYiEEL
kihatásának elemezésére is sor került mind a hizlalás alatti, mind a vágáskori teljesítményekkel összefüggésben.
$ NpUGpVN|U YL]VJiODWD QHP ~M NHOHW GH D NRUV]HU VHUWpVKL]ODOiVKR]
NDSFVROyGyDQPHJOHKHW VHQPpUVpNHOWV]iP~N|]OHPpQ\MHOHQLNPHJH
csoportban ugyanezek az értékek 606 g, 1001 g, 1,76 kg voltak. A beállítási testtömeg 21 és 37 kg között változott de ez az eltérés nem okozott szignifikáns különbséget a napi testtömeggyarapodásban.
SPRYSL és mtsai (1999) kísérletében a hízóbaállítási súlyok 21,30 és 29,84 kg között változtak. A kisebb súllyal induló sertések az átlagos 120 kg-EDQ W|UWpQ YiJiVLJ QDSRW D QDJ\REE V~O\~DN QDSRW
tölt|WWHNDKt]yLVWiOOyEDQ$]HO EELHVHWEHQJD]XWyEELEDQJ
napi testtömeggyarapodást mértek. A hízlalási napok száma gazdasági
V]HPSRQWEyOLVLJHQMHOHQW VWpQ\H] H]pUWpUWpNHOpVHPLQGHQKt]ODOiVL
kísérlet alapjául szolgál. NEVES és mtsai (1998) V]iUD]iUX HO iOOtWiV
céljára hizlalt, olyan sertéseket vontak kísérletükbe, amelyek súlya a hízlalás befejezésekor 130 kg volt, de kora 12 és 15 hónap között
YiOWR]RWW 0HJiOODStWRWWiN KRJ\ D] HOWpU Kt]ODOiV YpJL pOHWNRU
szignifikáns különbséget okozott a hússzínben, de nem tapasztaltak ilyen eltérést a hús kémhatásában és víztartó képességében.
7HUPpV]HWHVHQD]DGRWWV~O\LJWHUMHG Kt]ODOiVLLG pVDYiJiVLPXWDWyN D JHQRWtSXVRN N|]|WW MHOHQW V GLIIHUHQFLiOWViJRW PXWDWQDN 6]iPRV
kísérlet igazolja azonban, hogy egy-egy fajtán belül is heterogenitás tapasztalható a termelési paraméterek tekintetében. Ez a jelenség
ILJ\HOKHW PHJBEREK és mtsai (1982) közleményében, amelyben
DGDWRNDW LVPHUWHWQHN NO|QE|] IDMWiM~ VHUWpVHN N|]SRQWL
teljesítményvizsgálatának összehasonlításáról. Elemzésükben egy genotípuson belül 13 napos eltérést mutattak ki a vágáskori életkorban, amelyhez 34 g napi tömeggyarapodási és 170 g takarmányértékesítési különbség tartozott. A fenti, vágáskori életkorkülönbség esetében 3 mm-es átlagos hátszalonna vastagság, 3,4 cm2-es karajkeresztmetszet,
0,9%-os értékes húsrész % és 1,52%-os fehéráru % különbség számítható. Nyilvánvaló, hogy azonos fajtán belül, standard takarmányozási és tartási viszonyok között, esetleg nemek szerint is csoportosítva a populációt, a termelési mutatók kiegyenlítetlenségét egyedi biológiai, genetikai eltérések okozzák.
KOVÁCS és mtsa (1978) hízósertések hízékonysági és vágási teljesítményeinek alakulását elemezték életkor, illetve súly szerinti vizsgálat során. KíVpUOHWNEHQ D] HOV iUWiQ\FVRSRUWRW SRQWRVDQ
napos korban, 29,61 kg-os átlagos testsúllyal, a második ártánycsoportot pontosan 30 kg-os testsúllyal, de átlagosan 83 napos életkorral állították hízóba. A beállításkor pontosan 83 napos korú csoportban a sertések 169 napos életkorban érték el a 105,26 kg-os végsúlyt, míg a pontosan 30 kg-os beállítási súlyú csoport sertései 162,82 nap alatt produkáltak 101,24 kg-os testtömeget. Az azonos korban beállított sertések hizlalás alatti súlygyarapodása 869,85 g, takarmányértékesítése 3.20 kg volt, míg az azonos súlyban beállított sertéscsoport 899,64 g hizlalás alatti napi testtömeggyarapodást és 3,07 kg takarmányértékesítést ért el. BEREK és mtsai (1977) munkájukban ugyancsak 30 kg-ban hízóba állított sertések hízékonysági és vágási adatait dolgozták fel 102 kg súlyhatár eléréséig. Ad libitum etetés
de rávilágít a hízóbaállításkori fejlettség, a beállított állomány
HJ\|QWHW VpJpQHNIRQWRVViJiUDLV
KOVÁCS D KL]ODOiV HUHGPpQ\HVVpJpW PHJKDWiUR]y WpQ\H] N
között az állatok egyedi eltéréseire hívja fel a figyelmet. Véleménye szerint e sajátosságok megnyilvánulnak a növekedési erély,
WDNDUPiQ\pUWpNHVtW NpSHVVpJ pWNHVVpJ WHNLQWHWpEHQ ËJ\ D] HJ\HGL NO|QEVpJHNE O V]iUPD]y NLHJ\HQOtWHWOHQVpJ D VHUWpVKL]ODOiV HJ\LN I
gondja. A növekedési erély variabilitása 10%-os is lehet, még alomtestvérek esetében is, amely átlag 180 napos, azonos végsúlyú vágásnál 18 napos életkorkülönbséget jelent. Az átlagos 110 kg-os hízlalási végsúlynál 20,40 kg-RV NO|QEVpJHN LV DGyGWDN D] HOWpU
növekedési erély eredményeként, még az azonos hízóbaállításkori fejlettség ellenére is.
SCHANDL és mtsai (1961) javaslata szerint a hizlalás elején az egy
IDONiEDNHUO VOG NOHKHW OHJD]RQRVpOHWNRU~DNOHJ\HQHN$KL]ODOiV
végén az egyedek átlagos napi súlygyarapodását – az összehasonlíthatóság érdekében – mindig ugyanazon súlyhatárok között kell megállapítani, például: 30-130 kg, 40-140 kg, 30-140 kg, 40-150 kg súlyhatárok között. Adataik alapján már hízóba állításkor, 21- NJ pO W|PHJQpO J PtJ -40 kg-nál 430 g átlagos napi súlygyarapodás jellemezte a vizsgálatba vont sertéseket.
Napjainkban a Sertés Teljesítményvizsgálati Kódexben foglaltak
DODSMiQ QDSRV pOHWNRUEDQ NH]G GQHN D Kt]pNRQ\ViJL WHV]WHN 8J\DQDNNRU D] ]HPL KL]ODOiVL J\DNRUODW LV HOV VRUEDQ D] pOHWNRUW
veszik alapul a hízóba állításnál. A tapasztalat viszont azt mutatja, hogy
D]RQRV NRUEDQ MHOHQW V V~O\GLIIHUHQFLD MHOOHP]L D VHUWpVHNHW DPHO\HN
súlya akár 25-45 kg is lehet. BECKER és mtsai (1966) szerint egy 36,30 kg-os sertés elvárható napi tömeggyarapodása 750 g, takarmányértékesítése 2,80 kg, de egy 45,40 kg-os egyednél ugyanezen mutatók 820g illetve 3,10 kg.
$ PDODFNRUL HOWpU V~O\J\DUDSRGiV HUHGPpQ\HNpQW MHOHQWNH]
hízóbaállításkori nagyobb testtömeg és a hízlalás alatti testtömegnövekedés intenzitása közötti kapcsolatokat REICHENBACH (2001) vizsgálta. Azt állapította meg, hogy a 30-112 kg közötti súlyhatáron belül a növekedés-kompenzáló képesség csak egyes állatoknál következik be.
)HOWpWHOH]KHW KRJ\DN|]HOD]RQRVNRUEDQMHOHQWNH] HJ\HGLHOWpUpVHN D Kt]ODOiV NpV EEL V]DNDV]iUD D YiJiVL PXWDWyNUD LV NLKatnak.
Munkámban éppen ezért a fontosabb hízlalási paraméterek és a
EHiOOtWiVNRUL IHMOHWWVpJ NO|QE|] JHQRWtSXVRNRQ EHOOL |VV]HIJJpVHLW HOHPH]WHP 0LYHO D Kt]yDODSDQ\DJ V~O\D HJ\V]HU HQ PHJKDWiUR]KDWy
ezért e sajátosság és a hízlalási és vágási teljesítmények közötti
|VV]HIJJpVHNIHQRWtSXVRVNRUUHOiFLyNNDOYDOyHOOHQ U]pVHW|UWpQWPHJ
WHVWW|PHJUH YRQDWNR]y HJ\|QWHW VpJ PHOOHWWa hizlalás során további fontos szempont, hogy az állatok hizodalmassági és vágási tulajdonságaikban egységes képet mutassanak. Részleteiben ez azt
MHOHQWL KRJ\ D Kt]yEDiOOtWiV LGHMpUH N|]HO D]RQRV IHMOHWWVpJ
(egységesen 80 napos korban jelenleg 30-35 kg pO V~O\~ iOORPiQ\
NLDODNtWiViUDW|UHNHGQHNDPLNpV EEPDJDVV]LQWHQGHHJ\VpJHVHQIRJ J\DUDSRGQL pV WHUYH]KHW HQ HJ\|QWHW PLQ VpJ YpJWHUPpNHW IRJ V]ROJiOWDWQL$NRUV]HU VHUWpVIDMWiNJHQHWLNDLNpSHVVpJHLNDODSMiQFVDN PHJN|]HOtW HQ WHOMHVtWLN H]HNHW a feltételeket még optimális tartási és
WDNDUPiQ\R]iVL N|UOPpQ\HN N|]|WW LV 0LQG D WHQ\pV]W L PLQG D
termelési gyakorlat azonban sok esetben az optimális eredmény elérése
KHO\HWWDOHKHW OHJQDJ\REEWHUPpNW|PHJHO iOOtWiViUDW|UHNV]LN
A maximális terPHOpV LQNiEE |NRQyPLDL PHJN|]HOtWpVW WHV] OHKHW Yp
míg a biológiai szervezetek esetében az egyensúlyi helyzet fenntartása
NHUOKHW HO WpUEH (]W D] DODSHOYHW WiPDV]WMD DOi FEKETE (1995)
PHJiOODStWiVD PHO\ V]HULQW XJ\DQ D PDODFRNDW DG OLELWXP FpOV]HU
takarmányozni genetikai képességeik kiaknázása érdekében, de a
KR]DPQ|YHO N pV WDNDUPiQ\t]HVtW N iOWDO HO LGp]HWW W~O J\RUV Q|YHNHGpV NHUOHQG $] LJpQ\ V]HULQWL WDNDUPiQ\R]iVVDO NLKDV]QiOKDWy D ILDWDO iOODWRNUD MHOOHP] QDJ\REE UHODWtY Q|YHNHGpVL
erély és eEE O IDNDGyDQ D MREE WDNDUPiQ\pUWpNHVtW NpSHVVpJ $
takarmányfelvétel korlátozását állategészségügyi okok indokolhatják,
GH D PiVLN YpJOHWNpQW D NO|QOHJHV KR]DPQ|YHO NNHO NLHJpV]tWHWW
tápszerek etetése hajtatott malacneveléshez vezet. A hátrány hízókorbaQ MHOHQWNH]LN $ VOG N -75 kg-RV pO V~O\EDQ HOYHV]WLN
étvágyukat, tömeggyarapodásuk lelassul, takarmányértékesítésük
URPOLN 8J\DQH]HQ QHYHOpVL HOY IHOLVPHUpVpUH pV LG WiOOyViJiUD XWDO
hogy már KERTÉSZ (1963) is figyelmeztet a hajtatott nevelés hátrányaira. Leszögezi, hogy célunk a genetikai alap növekedési
HUpO\pQHN K~VWHUPHO NpSHVVpJpQHN NLKDV]QiOiVD D OHKHW OHJFVHNpO\HEE PpUWpN HO]VtURVRGiVVDO $] DGRWW JHQRWtSXVUD MHOOHP]
intenzitást meghaladó, túl gyors tömeggyarapodás miatt korán jelentkezik a mértéken felüli elzsírosodás.
.|]HO IpO pYV]i]DG HOWHOWpYHO DPLNRU D] HJpV] PH] JD]GDViJUD
trendként állapítható meg a kemizálás visszaszorulása és a természetes
PyGRQ HO iOOtWRWW ELRWHUPpNHN WpUKyGtWiVD PHJIRQWRODQGy D ELROyJLDL
határok túlzott feszítése. VADÁNÉ (l999) szerint a biogazdálkodás
IHOWpWHOH D] DQWLELRWLNXPRN pV KR]DPIRNR]yN PHOO ]pVH RÁCZNÉ (2000) állatvédelmi szempontokra hívja fel a figyelmet. A hizlaldába való betelepítésnél különös tekintettel kell lenni a kor és fejlettség kapcsolatiUD D Kt]yFVRSRUWRN NLDODNtWiViQiO (OWpU IHMOHWWVpJ iOODWRN QDJ\ FVRSRUWRNED KHO\H]pVH VWUHVV]RUNpQW KDW DPHO\ YpJV VRURQ
termeléscsökkenést eredményez. WITTMANN (2000) a sertéshizlalást
EHIRO\iVROy WpQ\H] N N|]|WW D] XWyQHYHOW PDODFRN IHMOHWWVpJpW D] HOV
helyen említi. JAKSA (1994) kísérletében fehér hússertések hízlalásának eredményeit vizsgálta. Kísérletében még azonos beállításkori fejlettség ellenére is (90 napos korban az ártányok
Az életnapi, vagy nettó tömeggyarapodás és az átlagos hátszalonna vastagság kapcsolatát elemezték TRAN és mtsai (1993) magyar nagyfehér húVVHUWpVHNNHO YpJ]HWW NtVpUOHWNEHQ .O|QE|]
JD]GDViJRNEDQ D] HO EEL NpW SDUDPpWHU N|]|WW
+0,21 korrelációt számítottak. JAKSA, TRAN és mtsai fentebb közölt vizsgálati eredményei is ösztönzést adtak jelen tanulmány elkészítésére, a hizlalás kezdeti testsúly és az egyéb hízékonysági és vágási paraméterek közötti kapcsolatrendszer elemzésére.
JAKSA (1993) magyar lapálysertésekkel és magyar nagyfehér
K~VVHUWpVHNNHO YpJ]HWW NtVpUOHWH DODSMiQ IHOYHWKHW DQQDN OHKHW VpJH KRJ\ D] pO W|PHJvagy hasított tömeg helyett az értékes húsrészek
W|PHJpW KDV]QiOMXN YDODPHO\ pUWpNPpU WXODMGRQViJ
megfogalmazásakor. Ha ezzel a sajátossággal végezzük a szelekciót,
DNNRU D]RNDW D] HJ\HGHNHW YiORJDWMXN NL PHO\HN QDSRV pO W|PHJH
az átlag felett van és H] D] pO W|PHJ D V]HOHNFLyV V]LQWHN HPHOpVHNRU Q|YHNV]LN (] D]W MHOHQWL KRJ\ D PDODFNRUL HU WHOMHV IHMO GpVL HUpO\
kihasználása - egy meghatározott fejlettségi állapot eléréséig -
|VV]KDQJEDQ YDQ D K~VWHUPHOpV JD]GDViJRVViJiW NLIHMH] PXWDWyN
alakulásával.
STANDAL ÉS VANGEN (1986) a növekedésre irányuló szelekció kidolgozásakor úgy találták, hogy a korrelációs válasz rendszerint pozitív a takarmányértékesítésre, aminek okát abban látták, hogy a
J\RUVDEEDQ IHMO G VHUWpV MREEDQ KDV]QRVtW WALACH ÉS ZEBROWSKA ODSiO\ NDQVOG NNHO YpJH]WHN WDNDUPiQ\R]iVL
kísérletet. Magas, alacsony, majd újra magas fehérje tartalmú
takarmány etetésekor azt tapasztalták, hogy a második fázis lemaradását nemhogy behozták az állatok, hanem a harmadik fázisban nagyobb életQDSL V~O\J\DUDSRGiVW SURGXNiOWDN PLQW D] HOV Ii]LVEDQ
illetve szignifikánsan több nitrogént építettek testállományukba. Tehát
D ILDWDO iOODWRNUD MHOOHP] QDJ\ Q|YHNHGpVL HUpO\ PpJ MHOHQW V
fejlettségbeli különbségek esetén is képes kompenzálni a lemaradást.
Mindezen ismeret ösztönzésére célom az volt, hogy a hízóbaállításnál
D]RQRV NRU~ pV XJ\DQDNNRU MHOHQW V IHMOHWWVpJEHOL D]D] pO V~O\
NO|QEVpJHNQHNDKDWiViWHOHPH]]HPDKL]ODOiVLLG KRVV]iUDYDODPLQW
egyes hízékonysági és vágási paraméterek alakulására.
2.5. Növekedés – K~VPHQQ\LVpJ YiJyiOODW PLQ VpJ D K~V
zsírtartalma, zsírsavak
$YiJRWWVHUWpVHNPLQ VtWpVpWKRVV]~LG QNHUHV]WODIHKpUiUXDUiQQ\DO MHOOHPH]WpN .pV EE D] pUWpNHV K~VUpV]HN PHQQ\LVpJpW LV ILJ\HOHPEH YHWWpN D PLQ VtWpVNRU $ K~VW DQQDN PLQ VpJL NULWpULXPDLW MHOHQW VHQ NLE YtWYH ILQRPDEE IL]LNDL NpPLDL SDUDPpWHUHNNHO LV MHOOHP]LN
napjainkban: hússzín, létartó képesség, kémhatás, elektromos
YH]HW NpSHVVpJ pVSCHWÖRER (2000) szerint RFN (red, firm, non exudative), PFN (pale, firm, non exudative), PSE, RSE (red, soft,
A behatóbb tájékozódás érdekében a karajból nyert zsír - annak
PHQQ\LVpJpQ W~OPHQ HQ- zsírsavösszetételét vizsgáltam egyes vágási
PLQ VpJHW MHO] PXWDWyNNDO NDSFVRODWRVDQ .tVpUOHWHPEHQ DEEyO D PiV NXWDWyN iOWDO PiU N|]|OW HUHGPpQ\HNE O LQGXOWDP NL PLV]HULQW D
genotípus és a kor határozottan módosítja a kérdéses beltartalmi értékeket. A korral összefüggésben CSAPÓ és mtsai (1999) néhány Magyarországon tenyésztett fajta és genetikai konstrukció szalonnájának zsírsavösszetételét mutatták be tanulmányukban. A nagy hústermelésre szelektált genotípusok hasított féltestének kisebb a
]VtUWDUWDOPD PLQW D PpUVpNHOWHEE K~VWHUPHOpVVHO UHQGHONH] IDMWiN
esetében, ami a zsírsavak összetételében is megmutatkozik (LAWRENCE és FOWLER nyomán WOOD és mtsai 1983).
A zsír és izomszövet zsírsavösszetétele és így a zsírok halmazállapota a korral változik. Az izomszövetben az intramuscularis trigliceridek zsírsavösszetétele a zsírok halmazállapotát jobban befolyásolja, mint a membránalkotó foszfolipidek zsírsavösszetétele (LAWRENCE és FOWLER 1997). Napjainkban jól ismert, hogy az n-3 zsírsavak
HVV]HQFLiOLVDN D QRUPiOLV Q|YHNHGpVKH] pV IHMO GpVKH] YDODPLQW N|]SRQWL V]HUHSHW MiWV]DQDN D N|YHWNH] EHWHJVpJHN PHJHO ]pVpEHQ
szívkoszorúér betegség, szívaritmia, magas vérnyomás, cukorbetegség, ízületi gyulladás, gyulladásos és immunbetegségek (KANG és LEAF
D PDJ]DW UHWLQD pV KiPIHMO GpVH +2))0$1 pV PWVDL OpJ] V]HUYL EHWHJVpJHN 785(. pV PWVDL $ W|EEV]|U|VHQ WHOtWHWOHQ ]VtUVDYDN HJpV]VpJUH J\DNRUROW KDWiViYDO D] DOiEEL V]HU] N LV
foglalkoznak (WISEMAN 1997, SIMOPOULOS 1999). Tekintettel
DUUD KRJ\ D NRUV]HU YiJyVHUWpV HO iOOtWiV PLQG QDJ\REE KDQJV~O\W
KHO\H]DK~VPLQ VpJLN|YHWHOPpQ\HNUpV]OHWHVHEEPHJLVPHUpVpUHH]pUW
kutatásomban a karajból kinyert zsír zsírsavösszetételét, a zsírsavak
HJ\PiVKR] YDOy DUiQ\iW pV D]RN QpKiQ\ K~VPLQ VpJL PXWDWyYDO YDOy
kapcsolatát is elemeztem.
2.6. Növekedés – K~VPHQQ\LVpJ K~VPLQ VpJ D K~V ]VtUWDUWDOPD ]VtUPLQ VpJNROHV]WHULQ
$ K~VLSDU pV D IRJ\DV]WyL SLDF D PDJDV PLQ VpJL N|YHWHOPpQ\HNHQ
kívül különOHJHV LJpQ\HNHW LV WiPDV]W D WHQ\pV]W NNHO WHUPHO NNHO
szemben. Ezek az elvárások folyamatos kutatásra ösztönöznek, amelynek során a sertés hústermelésének vizsgálata újabb és újabb
V]HPSRQWRNNDOE YO
A kísérletekben vizsgált fajtákat, genotípusokat olyan mennyiségi és
PLQ VpJL SDUDPpWHUHN V]HULQW pUWpNHOLN KRJ\ D V]HUpQ\HEE
teljesítményre képes fajták háttérbe szorulnak. A mennyiségi mutatók
Q|YHNHGpVpYHO GH D PLQ VpJ MDYXOiViYDO LV HJ\WW MiUW EL]RQ\RV NiURV
tulajdonságok megjelenése vagy hatványozoWW HO IRUGXOiVD -HOOHP]
SpOGDNpQW HPOtWKHW D QDJ\ K~VWHUPHOpVUH NpSHV IDMWiN VWUHVV]pU]pNHQ\VpJH pV J\DNRUL URVV] K~VPLQ VpJH 0HJROGiVNpQW
lehHW VpJ DGyGLN RO\DQ pUWpNHV K~VNpV]tWPpQ\HN PLQW SO D 6HUUDQR VRQNDHO iOOtWiViUD$IDMWDVWUHVV]PHQWHVVpJHPLDWWV]NVpJHVQHNOiWMiN NRUV]HU WHQ\pV]WpVVHO D IDMWD OpWV]iPiQDN HPHOpVpW $ V]HUpQ\
szaporasági mutatók miatt az embrió-átültetés alkalmazását hangsúlyozzák. BODÓ és MIHÓK (2002) a magyar, különleges, speciális termékeket hungarikum néven említik. Véleményük szerint a
SLDF PHJKyGtWiViUD NpSHVHN D NLW Q Q\HUVDQ\DJRW V]ROJiOWDWy KDJ\RPiQ\RV IDMWiN (QQHN QpJ\ DODSIHOWpWHOHNpQW D PHJIHOHO
léts]iPRW D My PLQ VpJHW D WXGRPiQ\RV PHJDODSR]RWWViJRW pV D
piacteremtést jelölik meg. A tudományos megalapozottságot
iOODWpOHWWDQL JHQHWLNDL WHQ\pV]W L pV K~VPLQ VpJL YL]VJiODWRNNDO OHKHW PHJWHUHPWHQL$K~VPLQ VpJLYL]VJiODWRNQDNNO|Q|VMHOHQW VpJNYDn a mangalica esetében, hiszen a fajta termelési paraméterei elmaradnak a
QDJ\IHKpUDODSiO\pVHJ\pENRUV]HU IDMWiNPHQQ\LVpJLPXWDWyLWyO
$PLQ VpJWpPDN|UpQHNIRQWRVWHUOHWpWNpSH]LNDKXPiQ-egészségügyi vonatkozások. Ebben a tekintetben a sertéshús egyik legfontosabb alkotórésze a zsír- és koleszterintartalom. A zsírtartalom a hús
SRUKDQ\yVViJiW pV t]OHWHVVpJpW MHOHQW VHQ EHIRO\iVROMD tJ\ PLQW D]
t]DQ\DJRN OHJI EE KRUGR]yMiW D K~V PLQ VtWpVHNRU D OHJW|EE HVHWEHQ
vizsgálják. Éppen ezért a húsipar bizonyos területein a kiváló hústermelést nyújtó fajtáknál a 60 %-nál nagyobb színhússzázalékot és a túl alacsony izmon belüli, intramuscularis zsírtartalmat nem kedvelik.
CUNHA (1977) szerint a telített zsírsavak és a koleszterin együttesen fejtik ki káros hatásukat az érrendszerre. A koleszterinmetabolizmus jellemzésekor HUSVÉTH (1994) megállapítja, hogy az
DUWHULRV]NOHUy]LV pUHOPHV]HVHGpV IRNR]RWWDEE HO IRUGXOiViQDN RND D
vérplazma koleszterinszintjének növekedése, majd a koleszterin artériák falába t|UWpQ OHUDNyGiVD$WDNDUPiQ\HUHGHW YDJ\DPiMEDQ NpS] G NROHV]WHULQE O D PiMEDQ HSHVDYDN NpS] GQHN +869e7+
1994), vagy továbbszállítódva a szteránvázas hormonok szintézisében
pVDYpUHUHNEHOV V]HUNH]HWpQHNNLDODNtWiViEDQYHV]QHNUpV]W58'$6
és FRENYÓ (1995) szerint a nagy koleszterintartalmú lipoprotein –
PLQW D NROHV]WHULQ HOV GOHJHV V]iOOtWyGiVL IRUPiMD QHJDWtY YLVV]DFVDWROiVVDO |QPDJD V]DEiO\R]]D D VHMWHNEH W|UWpQ EHMXWiViW +D H]DIRO\DPDWKLEiVDQP N|GLNDVHMWHNEHQ]VtUFVHSSHFVNpNYiOQDNki.
$YpUHUHNIDOiEDQH]D]VtUV]DSRUXODWYH]HWD]pUV] NOHWKH]DPHO\QHN
következménye a vérerek rugalmasságának csökkenése, majd az érrendszeri megbetegedések. A koleszterin által okozott humán betegségek tehát a hibás koleszterin-anyagcsere következményei is lehetnek. CSAPÓ (1999) megállapítja, hogy a koleszterin naponta felvett mennyisége nem haladhatja meg a 300 mg-ot.
SZABÓ és mtsai (2001) sertésfajták vérplazmájának koleszterinszintjét
PpUWpN ÈWODJRV pUWpNNpQW PPROOLWHU YROW MHOOHP] GH D
legalacsonyabb szintet mangalica-keresztezésben mérték. CSAPÓ és mtsai (2000) mangalica, magyar nagyfehér x magyar lapály és mangalica x duroc fajtatiszta és keresztezett sertések hátszalonnájának
A fogyasztói piac nagyobb százalékban a sovány, kis zsírtartalmú húsokat kedveli, de egyes termékek – kolbász és szalámifélék -
HO iOOtWiViKR] D QDJ\REE ]VtUWDUWDOP~ K~VRNDW NHUHVL D K~VLSDU
SCHWÖRER és mtsai (1989) úgy találták, hoJ\D]HJ\HVK~VPLQ VpJL
mutatók - íz, állag, csepegési veszteség – és a hús intramuscularis zsírtartalma között szoros kapcsolat van. A fajták között különbségek
MHOOHP] HN PLQG D] LQWUDPXVFXODULV ]VtUWDUWDORP PLQG D] pU]pNV]HUYL
vizsgálatok eredményében. A hússertések genetikai javítása az intramuscularis zsírtartalom csökkenéséhez vezetett, amelynek
|U|NO GKHW VpJH PDJDV pUWpN EiU H] D] pUWpN IDMWiQNpQW YiOWR]LN
Tapasztalataik alapján, a szelekció során a karaj intramuscularis zsírtartalmának csökkenésével javul a takarmányértékesítés, növekedésével viszont javul a napi súlygyarapodás. Ezt a megállapítást támasztják alá SCHWÖRER és mtsai (1999) kísérleti eredményei, melyekben közlik, hogy a BLUP tenyészértékbecslési módszerrel növelték a karaj intramuscularis zsírtartalmát. 1991 és 1998 között a genetikai növekedés mértéke a nagyfehéreknél 0,3%-os, a lapályoknál 0,2%-os volt. Ez együtt járt 18 grammos, illetve 25 grammos napi tömeggyarapodás növekedéssel. A takarmányértékesítés tekintetében 0,1 kg javulás mutatkozott.
A húsipar igényei folyamatosan változhatnak a vágósertések
PLQ VpJpYHO NDSFVRODWEDQ QRKD DODSN|YHWHOPpQ\NpQW IRJDOPD]]iN
meg a magas színhús %-ot, amelyhez alacsony fehéráru arány tartozik.
$ NO|QE|] WHUPpNHN HOWpU ]VtUWDUWDOP~ DODSDQyagot igényelnek,
WHKiW QpKiQ\ WHUPpN HO iOOtWiViKR] D QDJ\ ]VtUWDUWDOP~ K~V YiOLN V]NVpJHVVp $] RN D QDJ\ ]VtUWDUWDORP PHOOHWW D ]VtU PLQ VpJpEHQ LV NHUHVHQG :22' pV (16(5 V]HULQW D JHQHWLNDL KDWiV
befolyásolja az állati test zsírtartalmát és a vastagabb hátszalonna jobb
]VtUPLQ VpJJHO WiUVXO $ QDJ\IHKpUHNHW pV ODSiO\RNDW YpNRQ\DEE GH NHPpQ\HEE tJ\ URVV]DEE PLQ VpJ ]VtU MHOOHP]L .2/67$'
kísérlete alapján az javasolja, hogy a fajták közötti különbségek
PHJiOODStWiViW FpOV]HU D ILDWDO állatok (25-85 kg) esetében elvégezni.
10 és 105 kg között ugyanis a fajták között nagyobb eltérések
PXWDWNR]QDN D NOV ]VtUGHSyN WHNLQWHWpEHQ PLQW D EHOV ]VtUUDNWiUDN
vonatkozásában.
Húsipari méréseit felhasználva CSAPÓ (1999) a sertéskaraj zsírtartalmát 7%, koleszterintartalmát 53,6 mg/100 g értékben
KDWiUR]]D PHJ )HOYHWL D OHKHW VpJpW KRJ\ D K~V NROHV]WHULQWDUWDOPiW JHQHWLNDL WpQ\H] N LV EHIRO\iVROKDWMiN $ NHYpVEp QHPHVtWHWW
zsírosodásra hajlamos fajtáknál az izmon belüli nagyobb zsírtartalmat vLV]RQW HJ\pUWHOP HQ EL]RQ\tWRWWQDN OiWMD %2/(0$1' pV PWVDL PHJiOODStWRWWiNKRJ\DKRVV]~KiWL]RPEDQOpY ]VtUNROHV]WHULQ
tartalma és a genetikai alap között nincs összefüggés. Kísérletükben inkább a takarmányozás hatását hangsúlyozzák, mert a takarmányba adagolt koleszterin-kiegészítéssel a születés után 8, 20, és 24 héttel az idegszövet tömegében történt növekedést tapasztaltak.
3. ANYAG ÉS MÓDSZER
3.1.1.A kísérleti állatok tartási, takarmányozási körülményei
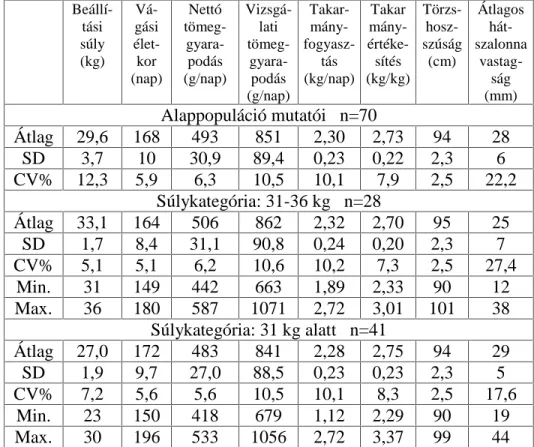
A keszthelyi hízékonyságvizsgáló állomáson 406 hízósertés hízékonysági adatait vontam vizsgálatba. A vegyesivarú, kísérleti állományban, amelyben a vizsgálati populációkat fele-fele ivararány jellemezte:
- 175 magyar nagyfehér hússertés (a továbbiakban (MNF) - 151 magyar lapálysertés, a továbbiakban (ML)
- 31 magyar nagyfehér hússertés x magyar lapálysertés keresztezés (a továbbiakban MNF x ML)
- 20 hibridsertés 1 a (továbbiakban H1) - 19 hibridsertés 2 a (továbbiakban H2)
- és 10 mangalica x pietrain keresztezés (a továbbiakban M x P) szerepel.
A vizsgálat 1998 és 2001 év között zajlott. Az állatokat az ország
NO|QE|] UpV]HLQ ]HPHO VHUWpVWHOHSHNU O V]iOOtWRWWiN D
hízékonyságvizsgáló állomásra, átlagosan 70 napos korban. A hizlalás
NH]GHWHNRU D VHUWpVHN iWODJRV pOHWNRUD QDS YROW GH LG N|]EHQ D
Sertés Teljesítményvizsgálati Kódexben a hizlalás kezdeti életkort 80 napra módosították. Ez az oka a vizsgálati létszám helyenkénti, hizlalás kezdeti változásának (80 napos életkor: MNF: n=163, ML: n=141, 90 napos életkor: MNF: n=175, ML: n=151). Az állatok hizlalása 105 kg átlagos testtömeg eléréséig tartott.
A hízósertések takarmányozása hízékonyságvizsgálati, granulált keveréktakarmánnyal történt, melynek egységes összetételét 12,33 MJ/kg metabolizálható energiatartalom (ME) és 18,61% nyersfehérje tartalom jellemezte. A takarmány adagolása ad libitum, szárazabrak
|QHWHW E O D Yt] LWDWiVD V]RSyNiV |QLWDWyEyO W|UWpQW $ VHUWpVHk hizlalása klimatizált istállóban folyt, félig rácspadozatú, egyedi ketrecekben. A testsúlynövekedés és a takarmányfogyasztás mérésére 10 naponként került sor.
3.1.2 A növekedési ritmus elemzése
Az állatok növekedésének matematikai leírását a VERHULST- BRODY-féle (JÁNOSSY és mtsai 1966) növekedési függvény teszi
OHKHW Yp DPHO\HW %52'< D] iOODWL WHVW Q|YHNHGpVL IRO\DPDWDLUD DONDOPD]WD 69È% V]HULQW D ELROyJLiEDQ iOWDOiEDQ D IHMO GpV-
Q|YHNHGpV NH]GHWL V]DNDV]D H[SRQHQFLiOLV IJJYpQQ\HO MHOOHPH]KHW (NNRU D IJJHWOHQ YiOWR]y ; W|EEQ\LUH D] LG D YL]VJiOW iOODW NRUD YDJ\ DIHMO GpVHJ\PiV XWiQN|YHWNH] V]DNDV]DLDIJJ YiOWR]y<
pedig a mért tulajdonság. Általános képletként az:
Y=abx (b>0) vagy az:
Y=aecx egyenlet, ahol az ec=b írható fel.
%$5726V]HULQWDV]HUYHVQ|YHNHGpVUHMHOOHP] KRJ\D]HJ\LN PHQQ\LVpJL WXODMGRQViJQDN SO V~O\ WHVWPpUHW < IJJ YiOWR]y D PiVLN PHQQ\LVpJKH] SO LG K|] ; IJJHWOHQ YiOWR]y YLV]RQ\tWRWW
változási sebessége arányos D] < IJJ YiOWR]yYDO $ V]HUYHV
növekedés egyenletét:
Y=cekt |VV]HIJJpVEHQ KDWiUR]]D PHJ KD D IJJHWOHQ YiOWR]y D] LG
Megállapítja, hogy a szerves növekedés egyenlete egy exponenciális trend.
Az Y=cekt egyenletben:
- k= a pillanatnyi, relatív növekedés mértéke, - e = a természetes logaritmus (ln) alapszáma, - Y= a mért súly (testtömeg),
- c= kezdeti súly -W LG < FKDW
$ V]HUYHV Q|YHNHGpV HJ\HQOHWpEHQ V]HUHSO F pV N SDUDPpWHUHNHW D OHJNLVHEE QpJ\]HWHN HOYH DODSMiQ PHJKDWiUR]YD D N|YHWNH]
normálegyenlet-rendszert kapjuk:
ORJ< QORJFNORJHW
WORJ< ORJFWNORJHW2
$ PyGV]HUEHQ IRJODOWDN V]HULQW HO EE D] DGDWRNDW ORJDULWPXVRV
diagramon ábrázoltam, hogy a növekedési szakaszok megállapíthatók legyenek. A diagram vízszintes tengelyén D] HJ\HQO N|] LG KHO\H]NHGLN HO D IJJ OHJHV WHQJHO\HQ SHGLJ D WHVWV~O\ -es alapú
ORJDULWPXVD (]HN XWiQ D SRQWGLDJUDP PHJN|]HOtW HQ HJ\ HJ\HQHVUH HV SRQWMDLW |VV]HN|W|WWHP pV PLQW Q|YHNHGpVL V]DNDV]RNDW NH]HOWHP
A számítógépes feldolgozásban a szakaszokra külön-külön elvégeztem a szerves növekedés egyenletének meghatározását. A munka
befejezésekor a k Q|YHNHGpVL NLWHY NHW KDVRQOtWRWWDP |VV]H (QQHN
folyamatában az egyes növekedési szakaszokra kiszámított exponenciális függvények egyenleteiben láthatók azok a súlyhatárok, amelyeknél a növekedés intenzitása (sebessége) megváltozik. A növekedési görbe töréspontjainak az x tengelyre (életkor) való
YHWtWpVpYHO SHGLJ D] pOHWNRUQDN PHJIHOHO HQ LV PHJKDWiUR]KDWyN D Q|YHNHGpVHOWpU LQWHQ]LWiV~V]DNDV]DLQDN határai.
$ Q|YHNHGpV V]DNDV]RVViJiQDN Q\RPRQ N|YHWpVpW D YL]VJiODWL LG V]DN
dekádjai átlagos napi súlygyarapodásának és a hizlalás idején felrakott
WHVWW|PHJ HJ\HV GHNiGRNUD HV -os megoszlásának ábrázolását is elvégeztem. Ezeken a diagramokon a vonaldiagram mindig a testsúlynövekedést az oszlopdiagram az adott paramétert szemléleti.
A növekedésdinamika takarmányozáselméleti vonatkozásában felmerülhetnek olyan kérdések, amelyek az egyes növekedési
V]DNDV]RNUD MHOOHP] QDSL WDNDUPiQ\IRJ\DV]WiV QDSL V~O\gyarapodás és a takarmányértékesítés közötti kapcsolatokat tisztázzák. Bonyolítja
H]W D WpPDN|UW KD JHQRWtSXVRNNpQW HOWpU Q|YHNHGpVL WHPHW WDSDV]WDOXQNKLV]HQD]]HPLKL]ODOiVLJ\DNRUODWPHJOHKHW VHQiOODQGy
határok közé szorítja a hízótápok beltartalmi értékeit. Az alapul
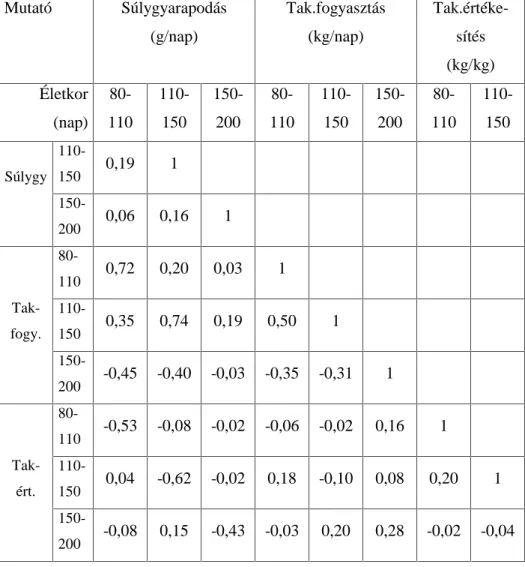
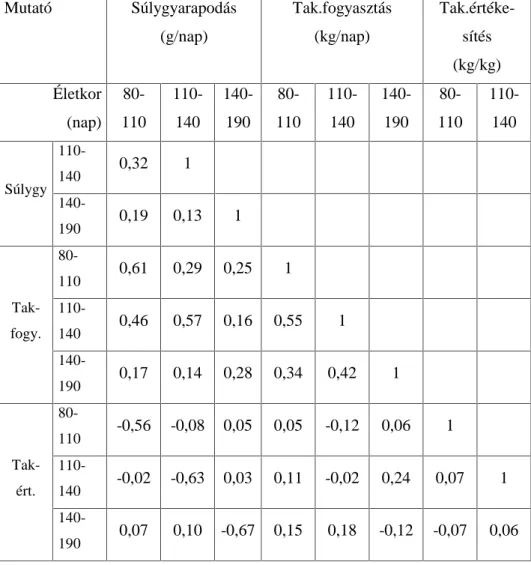
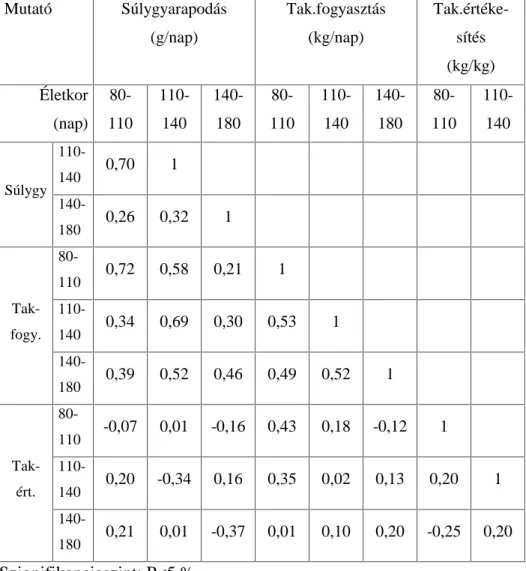
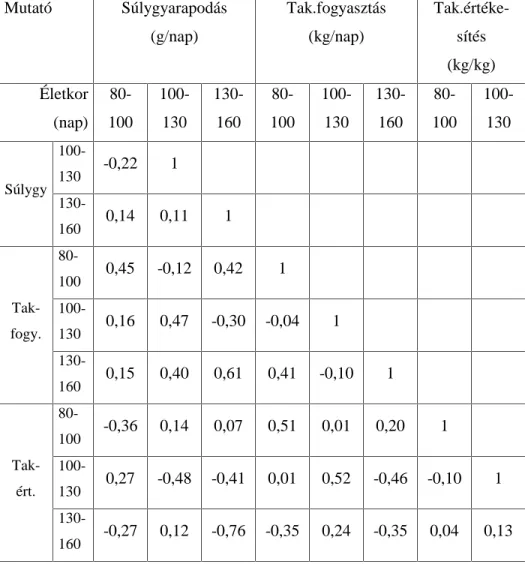
.RUUHOiFLyDQDOt]LVVHO DUUD LJ\HNH]WHP YiODV]W DGQL KRJ\ D] HOWpU
genotípusokra kiszámított növekedési szakaszok hízékonysági paraméterei (napi súlygyarapodás, takarmányfogyasztás,
WDNDUPiQ\pUWpNHVtWpV N|]|WW PLO\HQ LUiQ\~ pV HU VVpJ NDSFVRODWRN
mutatkoznak. A korrelációs mátrixok értékei fajtánként és genotípusonként mutatják a hízósertések növekedési ütemét az
DONDOPD]RWW IRO\DPDWRVDQ D]RQRV |VV]HWpWHO DEUDNNHYHUpkkel való takarmányozás körülményei között..
3.1.3. Testméretek, testarányok
$ V~O\Q|YHNHGpV PpUpVpQ W~OPHQ HQ D] HJ\HV JHQRWtSXVRNEDQ
testméret-IHOYpWHOH]pVHNNHOU|J]tWHWWHPDI EEWHVWPpUHWHNYiOWR]iViW$
QDSRVEHiOOtWiVLNRUUDONH]G G HQDKDYLV]ületésnapokon és a vágás
HO WWLQDSRQDKt]yVHUWpVHNHQD]DOiEELWHVWPpUHWHNHWYHWWHPIHO
-Magassági méretek: marmagasság, farbúbmagasság -Hosszúsági méretek: testhosszúság, törzshosszúság
-Szélességi méretek: vállszélesség, mellkasszélesség, farszélesség -Mélységi méret: mellkasmélység
-Körméret: övméret.
A mérésekhez Lydtin-IpOH PpU ERWRW pV PpU V]DODJRW KDV]QiOWDP $
kilenc testméretre vonatkozó, mért adathalmazból számított átlagértékek (leíró statisztika) bemutatása mellett, egyedenként, az adott havi születésnapon mért marmagasság százalékában fejeztem ki az összes többi, azonos napon mért testméretet (KERTÉSZ 1963, NAGY 1996). E módszer segítségével jobban bemutathatók és
pU]pNHOKHW ND]HOWpU JHQRWtSXVRNQ|YHNHGpVDODWWLWHVWDUiQ\YiOWR]iVDL
(relatív testméretek), mint a tényleges adatok értékelésével. Azért, hogy
PpJ V]HPEHW Q EEp WHJ\HP D WHVWPpUHWL YiOWR]iVRNDW D PDUPDJDVViJ V]i]DOpNiEDQNLIHMH]HWWW|EELWHVWPpUHWE ONLYiODV]WRWWDPDPLQLPXPpV
maximum értékeket felmutató genotípusokat. A kiválasztás feltétele,
KRJ\ OHJDOiEE NHWW -KiURP PpUpVL LG SRQW KDYL V]OHWpVQDS DONDOPiYDO PXWDWNR]]DQDN V]pOV pUWpNHN 7HUPpV]HWHVHQ HJ\QpO W|EE SRSXOiFLy LV PXWDWKDW V]pOV pUWpNHW (]HNQHN D SRSXOiFLyNQDN D
testarány változásait oszlopdiagramok szemléltetiN$]iEUiNIHOW Q HQ
jelzik, hogy melyik genotípus testarány változásai a legnagyobb vagy
OHJNLVHEEPpUWpN HND]DGRWWWHVWPpUHWWHNLQWHWpEHQ
3.1.4. Vágóérték vizsgálatok
A testméret-IHOYpWHOH]pVHNNHO HJ\ LG EHQ 62120$5. XOWUDKDQJRV
készülékkel (SONOMARK Kft. Esztergom) hátszalonna vastagság és
NDUDMiWPpU PpUpVHNHW YpJH]WHP D] (8523 h679 YL]VJiODWQDN PHJIHOHO HQ6HUWpV7HOMHVtWPpQ\YL]VJiODWL.yGH[
A hasított sertés testösszetételének meghatározásához az állatok vágása és darabolása a Sertés Hízékonyságvizsgálati Kódexben foglaltak alapján történt. Az értékes húsrészek megállapításának módszere az
HOV UHQG K~VRN V~O\iQDN PpUOHJHOpVH D NDUDMNHUHV]WPHWV]HW WHUOHW
A vágóérték paramétereinek hizlalás alatti változását az ultrahangos készülékkel mért adatokat felhasználva, grafikonnal szemléltetem. A
NRUV]HU KL]ODOiVEDQLVPHUWMHOHQVpJKRJ\D]pUWpNHVK~VUpV]HNDUiQ\D
és a fehéráru aránya körülbelül 150 kg-os testtömegnél keresztezi egymást (KOVÁCS nyomán KOVÁCS 1984, CSATÓ nyomán HORN), tehát tulajdonképpen a zsírépítés üteme ekkor haladja meg az
HOV UHQG K~VRN J\DUDSRGiViQDN WHPpW .tVpUOHWL HUHGPpQ\HLPHW
felhasználva, viszont azt kívántam megállapítani, hogy milyen
pO V~O\QiO pV pOHWNRUEDQ D NO|QE|] JHQRWtSXVRNUD YRQDWNR]WDWYD D
hizlalás folyamatában hogyan változik a színhús százalék (az ÜSTV-
QDNPHJIHOHO HQNJ-RVWHVWV~OO\DONH]G G HQMHOH]YHDNDUDMiWPpU
és az ultrahangosNpV]OpNNpWpUWpNpE OV]iPtWRWWiWODJRVKiWV]DORQQD YDVWDJViJ JUDILNRQMD ËJ\ NpSHW NDSKDWXQN D] HOWpU JHQRWtSXVRN QpKiQ\YiJypUWpNHWMHO] SDUDPpWHUpQHNKL]ODOiVDODWWLDODNXOiViUyO
3.1.5. Biometriai értékelés
A leíróstatisztikai értékelésben az átlag, a szórás, a variációs koefficiens (CV %) a minimum és maximum értékek kiszámítása történt. A elvégzett és bemutatott összefüggés-vizsgálatok statisztikai
PHJEt]KDWyViJL V]LQWMH HJ\VpJHVHQ 3 $ V]iPtWiVL P YHOHWHNKH]
MICROSOFT OFFICE EXCEL 97 számítógépes programot használtam.
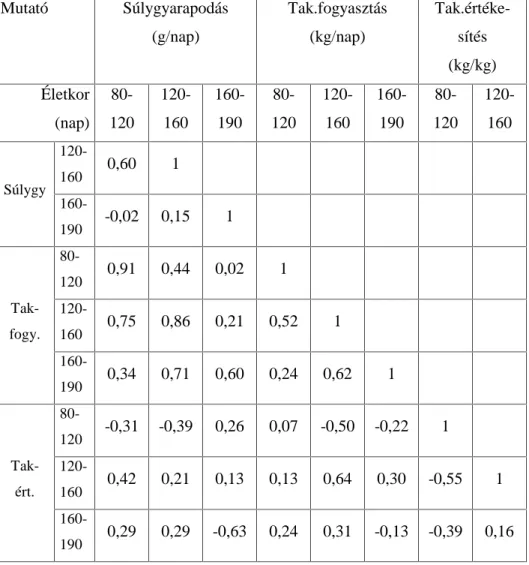
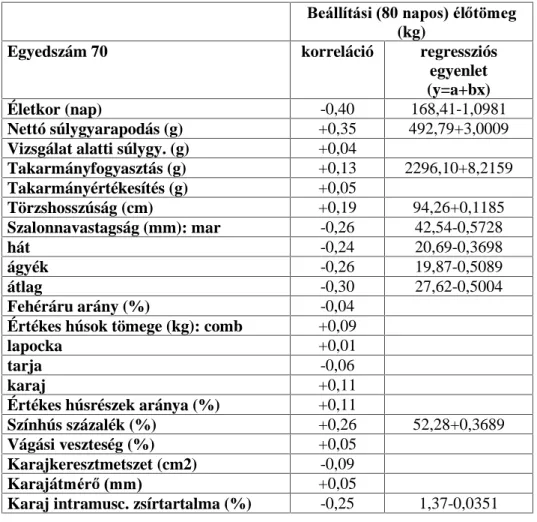
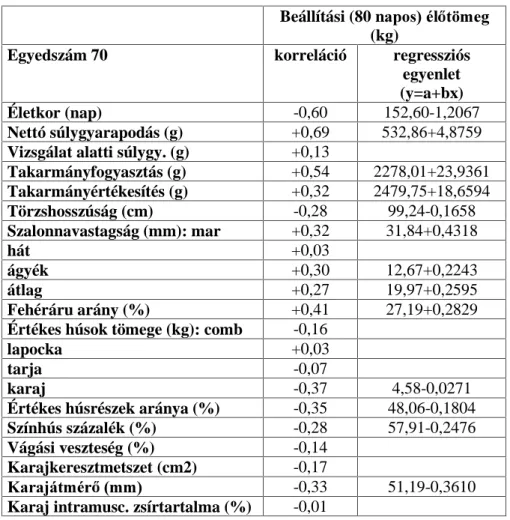
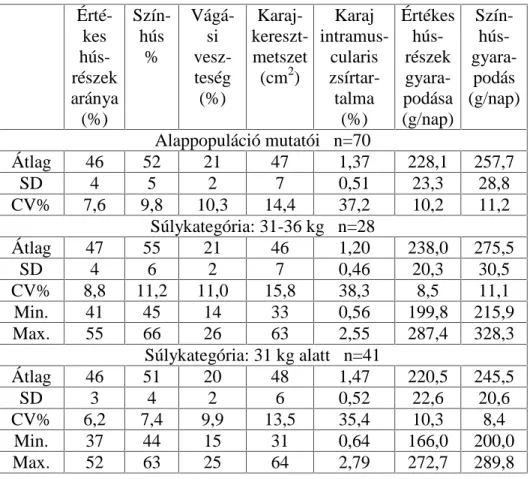
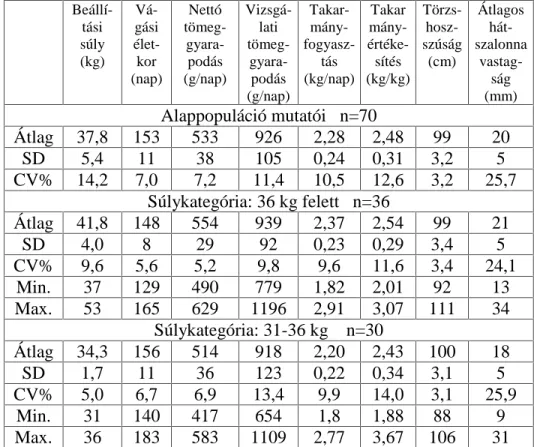
3.2. A hízóba állítási fejlettség vizsgálati módszere
A Magyarországon legelterjedtebb fajta a magyar nagyfehér hússertés, vegyes ivarú 70 egyedének, valamint a magyar lapálysertés ugyancsak
70 egyedének hízóbaállításkori ( QDSRV pOHWNRU pO V~O\D YDODPLQW D
fontosabb hízlalási és vágási mutatók közötti összefüggést vizsgáltam.
Magyarországon ugyanis a teljesítményvizsgálatokat azonos életkorban kezdik (80 napos életkorban). A vizsgált egyedek folyamatosan és párhuzamosan NHUOWHN D WHV]WiOORPiVUD NO|QE|] WHQ\pV]HWHN
állományaiból. Az állatok ad libitum fogyasztották a sertés
Kt]pNRQ\ViJYL]VJiOy iOORPiVRNRQ HWHWHWW V]DEYiQ\ V]HULQWL |VV]HWpWHO
abrakkeveréket, teljesen azonos, egyedi elhelyezési körülmények között. A súlygyarapodás és takarmányfogyasztás mérése 10 napos megosztásban történt a 105 kg-os vágási súly eléréséig.
A vágóérték meghatározása végett a tesztállomás vágóhídján végeztem a méréseket ugyancsak a teljesítményvizsgálati kódex darabolási
HO tUiVDLV]HULQW(QQHNNHUHWpEHQV]iPRVYiJypUWpNHWMHO] WXODMGRQViJ
(pl.: szalonnavastagsági méretek, értékes húsrészek mennyisége,
K~VPLQ VpJ WHVWPpUHWL DGDWRN NDUDMiWPpU NDUDM ]VtUWDUWDOPD VWE
szintén meghatározásra került.
A karaj zsírtartalmát a 13-14. hátcVLJRO\D IHOHWWL UpV]E O YHWW V]HOHWE O
nyert mintából, laboratóriumi, extrakciós úton állapítottam meg a 6830- as Magyar Szabvány szerint (MSZ 6830). A karaj vastagságát a levágás