DOKTORI (PhD) ÉRTEKEZÉS
Dr. GERGELY LÁSZLÓ
KESZTHELY 2004
VESZPRÉMI EGYETEM
GEORGIKON MEZŐGAZDASÁGTUDOMÁNYI KAR Növényvédelmi Intézet
Programvezető:
Dr. Sáringer Gyula az MTA rendes tagja
Növénytermesztési és Kertészeti Tudományok Doktori Iskola vezetője:
Dr. Gáborjányi Richard az MTA doktora
Témavezető:
Dr. habil. Fischl Géza
a mezőgazdasági tudomány kandidátusa
BURGONYAFAJTÁK REZISZTENCIAVIZSGÁLATA
FITOFTÓRA- (PHYTOPHTHORA INFESTANS [MONT.] DE BARY) FERTŐZÉSSEL SZEMBEN ÉS EGYES KÖRNYEZETI
TÉNYEZŐK HATÁSA A BETEGSÉG-ELLENÁLLÓSÁGRA
Készítette:
Dr. GERGELY LÁSZLÓ
KESZTHELY 2004
BURGONYAFAJTÁK REZISZTENCIAVIZSGÁLATA
FITOFTÓRA- (PHYTOPHTHORA INFESTANS [MONT.] DE BARY) FERTŐZÉSSEL SZEMBEN ÉS EGYES KÖRNYEZETI TÉNYEZŐK HATÁSA A BETEGSÉG-ELLENÁLLÓSÁGRA
Értekezés doktori (PhD) fokozat elnyerése érdekében
a Veszprémi Egyetem Növénytermesztési és Kertészeti Tudományok Doktori Iskolájához tartozóan
Írta:
Dr. GERGELY LÁSZLÓ A jelölt a doktori szigorlaton ………….%-ot ért el.
Keszthely, ………...
………..
a Szigorlati Bizottság elnöke Az értekezést bírálóként elfogadásra javaslom:
Bíráló neve:……….igen/nem
………..
aláírás Bíráló neve:……… ………..igen/nem
………..
aláírás Bíráló neve:……… ……… igen/nem
………..
aláírás A jelölt az értekezés nyilvános vitáján ……….%-ot ért el.
Keszthely, ………
………..
a Bíráló Bizottság elnöke A doktori (PhD) oklevél minősítése: ……….
………..
az EDT elnöke
TARTALOMJEGYZÉK
KIVONATOK ... 5
1. BEVEZETÉS ... 7
1.1. A téma jelentősége és időszerűsége... 7
1.2. A vizsgálatok előzményei ... 8
1.3. Célkitűzések... 8
2. IRODALMI ÁTTEKINTÉS ... 9
2.1. A Phytophthora infestans rövid jellemzése ... 9
2.2. A kórokozó gazdanövényköre és a betegség kialakulása, tünetei..11
2.3. A kórokozó változékonyságának és világméretű elterjedésé- nek (migráció) hatása az integrált növényvédelemre ... 14
2.3.1. A genetikai védekezésre gyakorolt hatások ... 14
2.3.2. A kémiai védekezésre gyakorolt hatások ... 15
2.3.3. Az agrotechnikai védekezésre gyakorolt hatások... 15
2.4. Rezisztenciára nemesítés a burgonyavésszel szemben... 16
2.4.1. Rassz-specifikus (vertikális) rezisztenciára nemesítés... 16
2.4.2. Nem rassz-specifikus (horizontális) rezisztenciára nemesítés .... 18
2.4.3. A lombozat és a gumók fitoftóra-ellenállóságának összefüggése 19 2.4.4. A rezisztenciára nemesítés nem hagyományos módszerei... 20
2.4.5. A rezisztens fajták szerepe az integrált növényvédelemben ... 21
2.5. Környezeti tényezők hatása az ellenálló képességre... 23
3. ANYAG ÉS MÓDSZER ... 26
3.1. A rezisztenciavizsgálatok anyaga és helyszínei... 26
3.2. A rezisztenciavizsgálatok módszerei ... 26
3.2.1. A lombozat fitoftóra-ellenállóságának vizsgálata fóliasátras provokációs kísérletekben ... 26
3.2.2. A lombozat fitoftóra-ellenállóságának vizsgálata szabadföldi provokációs kísérletekben... 29
3.2.3. A gumók fitoftóra-ellenállóságának vizsgálata fajtakísérletekben fellépő természetes fertőzés nyomán ... 32
3.2.4. A gumók horizontális rezisztenciájának vizsgálata laboratóriumban 32 3.2.5. Burgonyagumó-korongok elicitoros vizsgálata. ... 33
3.3. Összefüggés-vizsgálatok... 34
3.4. Meteorológiai megfigyelések ... 34
4. EREDMÉNYEK ... 35
4.1. Burgonyafajták lombozatának fitoftóra-ellenállósága ... 35
4.1.1. Fóliasátras provokációs kísérletek ... 35
4.1.2. Szabadföldi provokációs kísérletek ... 41
4.2. Burgonyafajták gumójának fitoftóra-ellenállósága ... 51
4.2.1. Rezisztenciavizsgálatok fajtakísérletekben, természetes fertőzés nyomán ... 51
4.2.2. Laboratóriumi rezisztenciavizsgálatok (egészgumós mesterséges fertőzés) ... 51
4.2.3. Burgonyagumó-korongok elicitoros vizsgálata ... 52
4.3. A lombozat és a gumók fitoftóra-ellenállóságának kapcsolata..59
4.4. Környezeti tényezők hatása a burgonyavésszel szembeni ellenálló képességre... 59
4.4.1. Fóliasátras provokációs kísérletek ... 59
4.4.2. Szabadföldi provokációs kísérletek ... 61
4.4.3. A gumók rezisztenciavizsgálata fajtakísérletekben, természetes fertőzés nyomán ... 64
4.5. Államilag elismert burgonyafajták fitoftórarezisztenciájának jellemzése ... 65
5. MEGVITATÁS ... 67
5.1. Burgonyafajták fitoftóra-ellenállóságának vizsgálata fajta- és provokációs kísérletekben ... 67
5.1.1. A lombozat rezisztenciavizsgálati módszereinek összehasonlító elemzése ... 67
5.1.2. A gumók rezisztenciavizsgálati módszereinek összehasonlító elemzése ... 69
5.1.3. A lombozat és a gumók fitoftóra-ellenállóságának összefüggése... 70
5.2. Környezeti tényezők hatása a burgonyavésszel szembeni ellenálló képességre ... 71
5.3. Az államilag elismert burgonyafajták rezisztenciaszintjének értékelése és szerepük az integrált és ökológiai termesztésben.. 73
6. ÖSSZEFOGLALÁS... 76
7. ÚJ KUTATÁSI EREDMÉNYEK... 80
8. IRODALOMJEGYZÉK ... 82
KÖSZÖNETNYILVÁNÍTÁS... 94
KIVONAT
BURGONYAFAJTÁK REZISZTENCIAVIZSGÁLATA
FITOFTÓRA- (PHYTOPHTHORA INFESTANS [MONT.] DE BARY) FERTŐZÉSSEL SZEMBEN ÉS EGYES KÖRNYEZETI TÉNYEZŐK
HATÁSA A BETEGSÉG-ELLENÁLLÓSÁGRA
A szerző rezisztenciavizsgálati módszerek továbbfejlesztésével értékelte a burgonyagenotípusok nem rassz-specifikus (horizontális) rezisztenciáját a burgonyavésszel (Phytophthora infestans) szemben, a lombozaton és a gumón.
Az értekezés foglalkozik a rezisztenciavizsgálati módszerek össze- hasonlító elemzésével, a lombozaton és a gumón kifejeződő rezisztencia összefüggés-vizsgálatával, a horizontális rezisztencia tartósságával, az állami- lag elismert burgonyafajták ellenállóságának komplex jellemzésével és egyes környezeti tényezők betegség-ellenállóságra kifejtett hatásaival.
Legfontosabb új tudományos eredmények:
• Megbízható provokációs módszer kidolgozása burgonya-fajtajelöltek horizontális fitoftórarezisztenciájának vizsgálatára a lombozaton.
• Államilag elismert burgonyafajták burgonyavésszel szembeni szántó- földi rezisztenciájának mennyiségi meghatározása a lombozaton három egymást követő évjáratban, a járványgörbe alatti terület (ADPC) átlagai alapján.
• A gumók horizontális fitoftórarezisztenciájának mennyiségi meghatározása digitális képfeldolgozáson alapuló fertőzöttségértékeléssel.
• A gumók elicitorérzékenysége és horizontális fitoftórarezisztenciája között igazolt közepes negatív összefüggéssel (r=–0,56, P=5%) annak megálla- pítása, hogy az elicitoros módszer nem alkalmas rezisztenciavizsgálatok céljára.
• Államilag elismert burgonyafajták komplex, mindkét növényi testtájra kiterjedő horizontális fitoftórarezisztenciájáról mennyiségileg meghatározott adatok első hazai közlése.
• Szabadföldi rezisztenciavizsgálatokban annak igazolása, hogy bizonyos fajták (pl. White Lady, Panda, Raja) szántóföldi rezisztenciája évjárattól függetlenül, változó környezeti feltételek között is következetesen kifeje- ződött, miközben több fajta (pl. Red Scarlett, Ukama, Cicero) érzékenyen reagált az évjárathatásokra.
ABSTRACT
TESTING FOR RESISTANCE OF POTATO VARIETIES TO LATE BLIGHT AGENT (PHYTOPHTHORA INFESTANS [MONT.] DE BARY)
AND THE EFFECT OF CERTAIN ENVIRONMENTAL FACTORS ON DISEASE RESISTANCE
The race non-specific (horizontal) resistance of potato genotypes was assessed to late blight agent (Phytophthora infestans) using comparative analysis and development of various resistance testing methods. In spite of the fact that environmental factors may have a significant effect on the horizontal types of resistance, the field resistance of certain registered potato varieties proved to be fairly stable even under altering conditions, expressing in both the foliage and tubers. Therefore these cultivars may play an important role in the integrated and/or organic farming of the near future.
AUSZUG
DIE RESISTENZPRÜFUNG DER KARTOFFELSORTEN GEGEN DEN PHYTOPHTHORA-BEFALL
(PHYTOPHTHORA INFESTANS [MONT.] DE BARY)
UND DIE AUSWIRKUNG EINIGER UMWELTFAKTOREN AUF DIE RESISTENZAUSPRÄGUNG
Der Autor hat Resistenzprüfungsmethoden weiterentwickelt und verglichen, mit denen man die nicht rassenspezifische (horizontale) Resistenz der Kartoffelgenotypen gegen die Kraut- und Knollenfäule (Phytophthora infestans) sowohl beim Laub, als auch beim Knollen sehr zuverlässig ermitteln kann. Trotz des beträchtlichen Einflusses verschiedener Umweltfaktoren auf die horizontalen Resistenzformen gibt es einige staatlich anerkannte Kartoffelsorten, deren Feldresistenz neben der stark schwankenden Auswirkungen der Jahrgänge an beiden Pflanzenteilen eine gewisse Stabilität zeigte. In der Zukunft könnten diese Sorten sehr wichtige Grundlagen für den ökologischen und/oder integrierten Kartoffelanbau sein.
1. BEVEZETÉS 1.1. A téma jelentősége és időszerűsége
A burgonyavész, melynek kórokozója egy sajátos biológiájú mikro- organizmus1, a Phytophthora infestans (Mont.) de Bary, világviszonylatban a burgonya legnagyobb gazdasági kárral fenyegető, legveszélyesebb betegsége. A kórfolyamatra kedvező (mérsékelten meleg és csapadékos) időjárási feltételek mellett, fogékony fajták állományában a termésveszteség elérheti a 80-100%-ot is, a teljes lombvesztés és súlyos gumófertőzés következtében. Ezt igazolta a kórokozó XIX. század közepén lezajlott első világméretű elterjedése (migrációja), amely a feltételezett őshazájából, Közép-Mexikóból indult ki és a növényi betegségek történetében minden idők legdrámaibb társadalmi következményeivel járt Írországban (Írországi éhínség és kivándorlás, 1845-47). O’Brian és Rich (1976) ezekre a tragikus eseményekre emlékeztetve a „történelmi” jelzővel illetik a betegséget.
A burgonyavész első magyarországi megjelenése 1846-ra tehető (Szepes vármegye), amikor Tognio Lajos pesti orvostanár felszólítást kapott a Magyar Királyi Helytartótanácstól a „Magyarhonban mostanában uralkodó burgonyabetegség” tanulmányozására (Husz 1941). Hazánkban a betegség 5-8 évente lép fel járványszerűen, de csaknem minden évben találkozha- tunk az általa kiváltott tünetekkel. Legutóbb 1998-ban okozott érzékeny veszteségeket a burgonya és paradicsom állományokban.
A burgonyavész kiemelkedő jelentőségére utal az a világméretű kutatási- és technológiafejlesztési együttműködés (Global Initiative on Late Blight, GILB), amelyet 1996-ban hívtak életre az integrált védekezési programok összehangolására (CIP 2003). A megoldandó probléma súlyát érzékelteti az a kimutatás, mely szerint a világ burgonyatermésének mint- egy 15%-a esik áldozatul a burgonyavésznek és a védekezés költségei a fejlődő országokban évente 3,25 milliárd USA dollárra becsülhetők (Mackin 1998).
A téma időszerűségét azok a jelzések is alátámasztják, amelyek az 1990-es években Észak-Amerikában és Nyugat-Európában bekövetkezett súlyos fitoftórajárványok kapcsán a védekezési stratégiák újragondolására hívják fel a figyelmet (Fry és Goodwin 1997, Schepers 1998).
1 Bár a Phytophthora infestanst hagyományosan gombaként rendszerezik, sejtszerveződése valójában nagyobb hasonlóságot mutat az algákéhoz és a magasabb rendű növényekéhez, mint a valódi gombákéhoz. Emiatt a legújabb törzsfejlődési rendszerek a kórokozót nem a valódi gombák (Eumycota) törzsébe, hanem egy önálló törzsbe (Oomycota) sorolják be (Hawkworth és mtsai 1995).
1.2. A vizsgálatok előzményei
Hazánkban a fajtaminősítés korábbi gyakorlata szerint a burgonya- vésszel szembeni rezisztenciavizsgálatok az országos fajtakísérletekben fellépő természetes lomb- és gumófertőzéseken alapultak, Hinfner Kálmán (1906-1976) munkássága nyomán. Az utóbbi évtizedekben azonban a meleg és aszályos évjáratok növekvő gyakorisága, valamint a fajtakísérle- tek hatékonyabb vegyszeres növényvédelme miatt ezzel a módszerrel már nem biztosítható a rezisztenciavizsgálatok folyamatossága. A megváltozott körülmények olyan provokációs vizsgálati módszerek kifejlesztését és/vagy adaptálását tették szükségessé, amelyekkel megbízhatóan és folyamato- san értékelhető a burgonyagenotípusok nem rassz-specifikus (horizontális) fitoftórarezisztenciája, a lombozaton és a gumón egyaránt.
A burgonyavész elleni integrált védelem egyik fontos elemével, a fajták ellenálló képességével (genetikai védekezés) 1988 óta foglalkozom, amikor elkezdtem a burgonya rezisztenciavizsgálatait az Országos Mezőgaz- dasági Minősítő Intézet Növénykórtani Osztályán, Budapesten. A burgonya- fajták állami elismeréséhez nélkülözhetetlen rezisztenciavizsgálatok mód- szertani nehézségei (a természetes fertőzések korlátozott felhasználható- sága, a lombozat és a gumók eltérő rezisztenciája, a fertőzési- és fertőzött- ségértékelési módszerekben rejlő bizonytalanságok) nagy kihívást jelen- tettek számomra. Ez arra ösztönzött, hogy 1997-ben jelentkezzem a PATE
„Az integrált növényvédelmi módszerek elméleti alapjai”, és ezen belül a
„Környezeti tényezők hatása a gombabetegségekkel szembeni ellenálló képességre” című doktori programjára, amellyel reményeim szerint hozzá- járulhatok a burgonya integrált növényvédelmének fejlesztéséhez.
1.3. Célkitűzések
Kutató-fejlesztő munkám legfontosabb célkitűzései a következők voltak:
1. Burgonyafajták és -fajtajelöltek burgonyavésszel szembeni horizontális rezisztenciájának vizsgálatára alkalmas mesterséges fertőzési és fertő- zöttségértékelési módszerek kifejlesztése és/vagy adaptálása.
2. A lombozat rezisztenciavizsgálati módszereinek összehasonlító elemzése.
3. A gumók rezisztenciavizsgálati módszereinek összehasonlító elemzése.
4. A lombozat és a gumók rezisztenciája közötti összefüggés vizsgálata.
5. A burgonyavésszel szembeni horizontális rezisztencia tartósságának vizsgálata.
6. Egyes környezeti tényezők hatásának vizsgálata a fajták ellenálló képes- ségére.
7. Államilag elismert burgonyafajták burgonyavésszel szembeni horizontális rezisztenciájának jellemzése mindkét növényi testtájon.
2. IRODALMI ÁTTEKINTÉS 2.1. A Phytophthora infestans rövid jellemzése
A fonalas telepszerveződése alapján hagyományosan gombaként rendszerezett kórokozó heterotallikus szervezet, ivaros szaporodása csak akkor lehetséges, amikor az egymással kompatibilis két párosodási típus (A1 és A2) térben és időben egyszerre van jelen. A valódi gombáktól eltérően vegetatív életciklusára a diploid (2n) állapot jellemző. Ivaros szaporodásakor a meiózis az ivarszervekben (oogónium, anterídium) következik be, így csak az ivarsejtek haploidok. Az ivaros kölcsönhatás végterméke a vastagfalú oospóra (petespóra), amely a talajban áttelelve évekig megőrizheti életképességét. Vegetatív úton micéliummal, sporan- giummal és rajzóspórákkal terjed, mely képletek a terjedést, illetve − ivaros folyamat hiányában − a faj fennmaradását szolgálják. A kétostoros, kettős membránnal határolt rajzók túlélési képessége páratlan az élővilágban: vízi életmódról szárazföldire váltanak (betokozódás sejtfalképzéssel), majd ezt követően áttérnek a gombaszerű, fonalas fejlődésre. A kórokozó alkalmazkodó- képessége is figyelemre méltó, ugyanis életciklusa a külső körülményekhez igazodik: az oospóra bő tápanyagellátás esetén csíratömlővel, kevés tápanyag mellett primer sporangiummal csírázik, ez utóbbi pedig vizes közegben rajzókat fejleszt, míg szárazabb, tápdúsabb környezetben közvetlenül csíratömlőt hajt. A micéliumon képződő sporangiumtartókról sporangiumok fűződnek le (1. és 2. ábra), melyekből vizes közegben, 10 °C körüli hőmérsékleten 6-16 db rajzóspóra szabadul ki. Ha a közegben tápanyag és kevés szabad víz van, a hőmérséklet pedig 25 °C körüli, akkor a sporangium − konídiumként viselkedve − közvetlenül csírázik (Érsek és Bakonyi 1997). Az elsősorban burgonyára specializálódott parazitának, eltérő fiziológiai rasszai révén, nagyfokú a fajtaspecializációja: egy-egy rassza csak bizonyos burgonyafajtákat képes megbetegíteni, másokat viszont nem. A rasszmeghatározást rendszerint úgynevezett differenciáló gazdanövénysorral végzik, melyben a burgonyagenotípusok eltérő fitoftóra- rezisztencia-géneket (domináns R-gének) hordoznak. Flor (1955) gén génnel szembeni (gene-for-gene) elmélete alapján az egyes rasszokat aszerint, hogy melyik R-gén rezisztenciáját képesek feloldani, az illető R-gén számával látták el. Ennek megfelelően az 1-es rassz a recesszív (r-génes), azaz általánosan fogékony fajtán kívül az R1-gént, az 1.2-es rassz pedig az R1- és R2-, valamint az R1, R2-géneket tartalmazó genotípust képes megbetegíteni, és így tovább (Érsek és Bakonyi 1997).
1. ábra A Phytophthora infestans sporangiumtartói és sporangiumai burgonyalevélen (40 x-es nagyítás)2
2. ábra A Phytophthora infestans sporangiumtartói és citrom alakú sporangiumai (500 x-os nagyítás)
2 Az értekezés valamennyi felvételét a szerző készítette.
2.2. A kórokozó gazdanövényköre és a betegség kialakulása, tünetei A P. infestansnak szűk a gazdanövényköre: a burgonyán és a Solanum- nemzetség más fajain kívül csak a paradicsomot képes megbetegíteni.
Újabban az Egyesült Államokban üvegházban nevelt petúnián (Petunia x hybrida Hort.) is megbetegedést okozott (Deahl és Fravel 2003).
Fertőzésre hajlamosító, hűvös és csapadékos időjárásban a burgonyavész járványos fertőzése állandó fenyegetést jelent. A fertőzés legtöbbször légáramlatokkal érkező sporangiumokkal történik, melyek a növény hajtásain, levelén lévő vízcseppben közvetlenül (csíratömlővel) vagy közvetve (rajzóspórákkal) csíráznak. A közvetlen (konídiumszerű) csírázás hőmérsékleti optimuma 24 °C, míg a rajzóspóraképződésé 12-13 °C-ra tehető (Mizubuti és Fry 1998). Ullrich (1962) szerint a sporangiumok kialaku- lásához 8 °C-ot meghaladó léghőmérséklet és legalább 8 órás telített pára- tartalom szükséges. Ahol az ivaros kölcsönhatás előfordul, ott a talajban áttelelő oospórák is elsődleges fertőzési források lehetnek. Az egyik évről a másikra, illetve a nagy földrajzi távolságokra való terjedés leggyakrabban micéliummal fertőzött gumóval történik. Enyhébb telű vidékeken az árva- kelésű burgonyák és a hulladékburgonya-halmok tekinthetők a legfontosabb elsődleges fertőzési forrásoknak.
A betegség tünetei a lombozaton: többnyire a levéllemez széléről vagy csúcsáról kiinduló, szabálytalan alakú, nedves időben vízzel átitatott, barnuló és gyorsan terjedő foltok, melyek később beszáradnak. A foltokat esetenként sárga, klorotikus udvar szegélyezi (3. ábra). A levélfonákon, a foltok szélén nedves, párás időben fehér színű penészgyep képződik, amely a sporangiumtartók és sporangiumok tömegéből áll (4. ábra). A levélnyélen és a száron is megjelenhetnek a betegség tünetei szabálytalan alakú barna foltok formájában (5. ábra), melyeken szintén sporulálhat a gomba. A sporangiumok széllel vagy vízzel (eső vagy öntözővíz) terjednek növényről növényre, illetve a talajon keresztül a gumókra és a gyökérzetre.
A gumók sérüléseken, rügyeken (szemeken), paraszemölcsökön vagy a sztólókon keresztül fertőződhetnek. A gumófertőzés tünetei: a gumóhéjon kissé besüppedő, kemény tapintatú, ólomszürke foltok képződnek, melyek alatt a gumószövetek vörösesbarna, barna szárazkorhadása látható (6. ábra).
Nem megfelelő tároláskor a betegség gyorsan terjed a héjtól a belső szövetekbe és gumóról gumóra, ami súlyos rothadási veszteségekhez vezet. Gyakoriak a másodlagos fertőzések (Fusarium spp., Erwinia spp.) nyomán kialakuló vegyes rothadások is.
3. ábra A burgonyavész kezdeti tünetei a lombozaton
4. ábra Burgonyvész okozta sporuláló foltok a lombozaton
5. ábra A burgonya fitoftórás szárfertőzésének tünetei
6. ábra A burgonyagumó fitoftórás szárazkorhadásának tünetei
2.3. A kórokozó változékonyságának és világméretű elterjedésének (migráció) hatása az integrált növényvédelemre
2.3.1. A genetikai védekezésre gyakorolt hatások
A P. infestansnak az 1970-es évek végén bekövetkezett második nagy elterjedése (migrációja) olyan lényeges változásokat eredményezett világszerte a helyi populációkban, amelyek a rezisztenciára nemesítési stratégiák újragondolását tették szükségessé (Spielman és mtsai 1991, Zarzycka és Sobkowiak 1997a). A régi, egyszerű rasszokból álló populációt egy összetett virulenciájú, komplex rasszokból álló populáció váltotta föl, amely gyakran együtt járt az agresszivitás erősödésével (Peters és mtsai 1998, Miller és mtsai 1998, Bohár és mtsai 1999). Németországban és Hollandiában az 1980-as évek végére gyakorlatilag megduplázódott az egy izolátumra jutó virulenciagének száma az 1970-es évekhez viszonyítva, a különböző virulenciagén-kombinációjú rasszok száma pedig megtöbbszörö- ződött (Spielman és mtsai 1991). Hasonló folyamat játszódott le Magyar- országon is az 1960-as évekből származó (Mudich 1965) és a legutóbbi vizsgálatok (Érsek 2001) tanúsága szerint. Mivel a komplex virulenciájú rasszok megjelenése időben egybeesik az A2 párosodási típus európai elterjedésével, kézenfekvő volt az ivaros folyamatokban keresni a változások magyarázatát. Fry és munkatársai (1993) szerint azonban molekuláris markereken (izozimmintázat, DNS fingerprint) alapuló elemzések nem támasztják alá ezt a feltételezést, jóllehet az ivaros rekombináció révén létrejövő genetikai változásokra már van európai példa (Drenth és mtsai 1994, Sujkowski és mtsai 1994). A rezisztenciagének (R-gének) szelekciós nyomása sem érvényesülhetett, hiszen a termesztett burgonyafajták több- sége vagy egyáltalán nem tartalmaz R-géneket, vagy csak a kisebb szám- mal jelzett (R1-R4) géneket hordozzák különböző kombinációkban. Miután az ivartalan ciklus és a mutációk szerepét is kizárták a lehetséges okok közül, arra a következtetésre jutottak, hogy az új populációk kialakulásában a kórokozó migrációja („gene-flow”) játszotta a legfontosabb szerepet (Fry és mtsai 1993). Természetesen a behurcolt populációk összetétele azóta is állandó változásnak van kitéve az ivaros és ivartalan rekombináció és a mutáció révén.
Az összetett virulenciaspektrumú, rezisztenciatörő rasszok fokozatos terjedésével egyre nyilvánvalóbbá vált a vertikális rezisztencia sebezhető- sége (Mackay és mtsai 1992, Peters és mtsai 1998, Gergely és mtsai 2003). Emiatt a burgonyavésszel szembeni rezisztenciára nemesítésben stratégiaváltás következett be az elmúlt néhány évtizedben a nem rassz- specifikus (horizontális) rezisztenciatípusokat részesítve előnyben.
2.3.2. A kémiai védekezésre gyakorolt hatások
Napjainkban a vegyszeres védelemnek meghatározó szerepe van a burgonyavész elleni küzdelemben. A nem megfelelő fungicidhasználat azonban felgyorsíthatja a kórokozó populációiban kialakuló szerreziszten- ciát. Az oospórás gombák ellen széleskörűen alkalmazott fenilamidok közül főként a metalaxilra rezisztens P. infestans izolátumok fellépéséről jelentek meg közlemények (Davidse és mtsai 1989, Bakonyi és Érsek 1997, McLeod és mtsai 2001). Gisi és Cohen (1996) szerint a fenilamidrezisztencia kialakulása nincs összefüggésben sem az izolátumok párosodási típusával, sem a rasszkarakterrel és a rezisztens izolátumok száma jelentősen változik évről évre, sőt a tenyészidőszakon belül is. A rezisztens törzsek kiszelektálódásában a helytelen fungicidhasználat a meghatározó, de föld- rajzi elterjedésükben a migráció is fontos szerepet játszhat. A feltételezések szerint Észak-Mexikóból az Egyesült Államokba behurcolt, és ott 1994-ben súlyos járványt okozó metalaxilrezisztens P. infestans rassz (US-8) fellépése nyomán elementáris igény mutatkozott új, hatékony fungicid-hatóanyagok iránt (Mayton és mtsai 2001).
Hanson és Shattock (1998) szerint az oospóraképződés a közepes szántóföldi rezisztenciájú fajták lombozatában volt a leggyakoribb, ami a kiegészítő vegyszeres védelem jelentőségére hívja fel a figyelmet. Ráadásul az ilyen genotípusok lombján, bár mérsékelten, de hosszantartóan sporulálhat a kórokozó, ami megnöveli a gumófertőzések veszélyét.
2.3.3. Az agrotechnikai védekezésre gyakorolt hatások
A P. infestans A2 párosodási típusának széleskörű elterjedése meg- teremtette a lehetőségét az ivaros szaporodás általánosabbá válásának. Az ivaros kölcsönhatásból származó kitartó spóraalak (oospóra) a talajban évekig életképes maradhat, növelve ezzel az elsődleges fertőzési források számát (Zarzycka és Sobkowiak 1997b, Schepers 1998). Ez a kedvezőtlen folyamat pedig végsősoron a vetésváltás növényvédelmi hatékonyságának csökkenéséhez vezethet. Hanson és Shattock (1998) A1 és A2 párosodási típusú izolátumokkal fertőzött, eltérő fogékonyságú burgonyafajták levelé- ben vizsgálta az oospóraképződést. A nagyon fogékony fajták (Home Guard, Bintje) lombja a fertőzést követően hamar elpusztult, így ezekben az oospóraképződés elmaradt. A közepes szántóföldi rezisztenciájú fajták levelében és szárában gyakori oospóraképződést észleltek, a legellenállóbb fajtából (Stirling) viszont nem lehetett kitartó spóraalakot kimutatni. Az oospórás áttelelés alapvetően megváltoztathatja a betegség járványtani helyzetét és előrejelzését is a korai fertőzések lehetősége miatt (Strömberg és mtsai 1999).
2.4. Rezisztenciára nemesítés a burgonyavésszel szemben 2.4.1. Rassz-specifikus (vertikális) rezisztenciára nemesítés
A burgonyavésszel szembeni rezisztenciára nemesítés rassz-specifikus útja az 1910-es években kezdődött (Jellis 1995), miután 1909-ben Salaman felfigyelt a kultúrburgonya (Solanum tuberosum L.) és egy vad rokon faj (S. demissum Lindl.) keresztezéséből származó fajhibridekben a fertőzés nyomán bekövetkező hiperszenzitív (túlérzékenységi) válaszreakcióra (HR).
Ennek a védekezési reakciónak a lényege a fertőzési hely körül kialakuló, korlátozott kiterjedésű, gyors szövetelhalás, amellyel a gazdanövény lokalizálja a kórokozót. Később a fent említett interspecifikus hibridizációval előállított burgonyafajtákkal igazolták a P. infestans nagyfokú fajtaspeciali- zációját, amely szerint a kórokozó populációi eltérő fiziológiai rasszokból állnak és egy-egy rassz csak bizonyos fajtákat képes megbetegíteni, másokat viszont nem (Király és mtsai 1970). A HR-en alapuló rezisztenciát a Solanum nemzetség vad fajaiban, elsősorban a S. demissumban és a S. stoloniferumban lévő egyedi és domináns rezisztenciagének (R-gének) határozzák meg. Emiatt a rassz-specifikus vagy vertikális rezisztenciát főgénes (R-génes) ellenállóságként is tárgyalják, utalva a mono- vagy oligo- génes öröklésmenetre. A Solanum nemzetségben eddig felkutatott legalább 11 R-gén közül 9-et a hexaploid mexikói vad fajban, a S. demissumban azonosítottak, amely a legnépszerűbb rezisztenciaforrássá vált, mivel 336 európai burgonyafajta pedigréjében szerepel (Darsow és mtsai 1998).
Németországban Broili és Müller a S. tuberosum x S. demissum kereszte- zésből származó fajhibrideket „W-rasszok”-nak nevezte el, amelyekből számos rezisztens fajtát állítottak elő, mint pl. a Sandnudelt és az Aquila-t.
Főként az utóbbi fajtát azután Európa-szerte használták szülőpartnerként a nemesítők (Erwin és mtsai 1983). A rassz-specifikus rezisztencia sebez- hetőségére az 1932. évi fitoftórajárvány mutatott rá elsőként, amikor az addig ellenálló „W-rasszok” is megbetegedtek (Ross 1979). A rezisztenciatörő rasszt (amely az R1-gén által szabályozott védelmet feloldotta) később 1-es rasszként jelölték. Az a tény, hogy ezt követően nem gyorsult föl a kórokozó fiziológiai specializációja új, virulens rasszokat eredményezve, részben azzal magyarázható, hogy a rezisztens (R-génes) fajták nem tudtak elterjedni a köztermesztésben sem Európában, sem Észak-Amerikában (Erwin és mtsai 1983). Emiatt az R-gének nem fejthettek ki szelekciós nyomást a kórokozó helyi populációira. A fogyasztói konzervativizmus ugyanis mindenekelőtt a kiváló étkezési minőséget honorálja, ezért egyes népszerű fajták akár 50-100 évig (!) is jelentős arányt képviselhetnek a fajtaszerkezetben – a betegségek iránti fogékonyságuk ellenére (pl. a Bintje fajta Hollandiában, vagy a Russet Burbank fajta az Egyesült Államokban). A P. infestans eltérő virulenciájú változatainak helyi kialakulását az a körülmény is hátráltatta, hogy az 1980-as évekig Közép-Mexikón kívül az ivaros
rekombináció nem járulhatott hozzá a változékonyság növeléséhez az A2 törzs hiányában. A burgonya növényegészségügyi helyzete ezt követően, a kórokozó 2. világméretű elterjedése nyomán változott meg drámaian, amikor komplex virulenciájú, A2 törzset képviselő és helyenként metalaxil- rezisztens rasszokat hurcoltak be Mexikóból Észak-Amerikába és Európába (Spielman és mtsai 1991, Goodwin és mtsai 1994, Zarzycka és Sobkowiak 1997a). A szokatlan hevességgel támadó és súlyos károkat okozó fitoftóra- járványok tették nyilvánvalóvá a vertikális ellenállóság legnagyobb hátrányát, nevezetesen azt, hogy az egy (vagy néhány) rasszal szemben fennálló védelem bizonytalan időtartamú és azt könnyen áttörhetik a kórokozó helyben kialakuló és/vagy migráció útján behurcolt virulens rasszai.
Magyarországon a burgonyavész elleni rezisztenciára nemesítés II.
világháború utáni első eredménye 1960-ban a Somogyi sárga fajta volt (nemesítő: Barsy Sarolta), melynek rezisztenciája lehetővé tette gombaölő- szermentes termesztését. A fajta a magyar nemesítésű Margit és a német Edelgard keresztezéséből származott, mely utóbbi valószínűleg a „W-rasz- szok”-ra vezethető vissza (Lönhárd és mtsai 2003). Később, 1967-68-tól a Sárvári István vezette keszthelyi nemesítőcsoport indított R-génes bázison nemesítési programot azzal az elképzeléssel, hogy több R-gén kombinálása egy genotípusban nemcsak a rassz-specifikus, hanem a horizontális ellen- állóságot is fokozza. Darsow és munkatársainak (1993) vizsgálatai egyér- telműen igazolták, hogy a Solanum tuberosum ssp. andigena, mint termesz- tett kultúrváltozat, valamint a S. demissum és S. stoloniferum vad fajok a horizontális ellenállóság szempontjából is értékes rezisztenciaforrásoknak tekinthetők. A hazai rezisztenciára nemesítés széles genetikai bázison, 10 vad burgonyafaj (közöttük a S. acaule, S. demissum, S. stoloniferum, S. andige- num és S. spectabile) bevonásával kezdődött és 1978-ra már olyan ”extrém”
fitoftórarezisztenciájú fajhibrideket eredményezett, amelyek az akkor ismert R-géneken kívül a S. spectabileból származó két komplementer gént is hordoztak (Sárvári 1978).
A több mint 40 éve elkezdett nagyszabású nemesítési program késői hozadékának tekinthető annak a piros héjú, új burgonyafajtának a sikeres szereplése egy angliai fitoftóra-provokációs kísérletben (Northumberland), amelyet a néhai Sárvári István fia által fémjelzett Sarpo társaság állított elő.
Az ökológiai termesztés számára is ígéretes fajta nagyfokú szántóföldi rezisztenciájával tűnt ki, miközben a különböző fogékonyságú kontrollfajták (Nicola, Bintje, Santé, Cara) jelentős lombveszteséget szenvedtek el (Anonym 2001). Örvendetes módon a keszthelyi nemesítési hagyományok napjainkban sem szakadtak meg, hiszen a Veszprémi Egyetem Burgonya- kutatási Központjában a komplex betegség-ellenállóságon túl az új fajták étkezési és feldolgozási minőségének javítását is kiemelt nemesítési célként tűzték ki (Polgár Zs., szóbeli közlés).
2.4.2 Nem rassz-specifikus (horizontális) rezisztenciára nemesítés
Amikor a burgonyavészjárvány végigsöpört Észak-Amerikán és Európán a XIX. század közepén, a termesztett burgonyafajták többsége nagyon fogékony volt (mint pl. a katasztrofális írországi járvány idején a köztermesztést uraló Lumper fajta). Néhány azonban túlélte a pusztító epidémiát, ezek voltak azok a genotípusok, melyekről később megállapí- tották, hogy nem rassz-specifikus (horizontális) ellenállósággal bírnak (Erwin és mtsai 1983). Egészen a HR-alapú rezisztencia felfedezéséig (1900-as évek eleje) gyakorlatilag erre a rezisztenciatípusra nemesítettek és csak a Solanum nemzetségben fellelt domináns rezisztenciagének (R-gének) változtatták meg a nemesítési stratégiát. Ennek alapvetően két oka volt:
egyrészt az R-génes ellenállóság a HR révén teljes lomb- és gumó- védelemmel kecsegtetett, hiszen sporuláció hiányában nem képződött fertőző anyag a lombozaton; másrészt a monogénes, domináns öröklés- menet viszonylag gyors eredményt ígért a nemesítői munkában. Ezzel az erős motivációval indult el a rassz-specifikus rezisztenciára nemesítés arra a több mint 50 éves útjára, melynek során a kultúrburgonya számos értékes fajtájába építették be a S. demissum rezisztenciagénjeit hagyomá- nyos, visszakeresztezéses nemesítéssel (Toxopeus 1956, Swiezynski és mtsai 1993, Micheletto és mtsai 1999). Ezt az ígéretes folyamatot szakították meg a P. infestans populációiban világszerte lezajlott és még ma is tartó lényeges változások (virulenciaspektrum bővülése, A2 törzs elterjedése, fittebb és/vagy agresszívebb izolátumok fellépése), amelyek ismételten stratégiaváltásra kényszerítették a nemesítőket, ezúttal a nem rassz-specifikus rezisztenciatípusokat részesítve előnyben. Bár ezek a rezisztenciaformák nem nyújtanak teljes védelmet, de mivel minden rasszal szemben kifejeződnek, a horizontális rezisztencia kifejezést is használják meg- nevezésükre. A szakirodalomban előforduló egyéb szinonim elnevezések:
általános-, részleges- vagy szántóföldi rezisztencia. A horizontális rezisztencia nem a kórokozó lokalizációjára és elpusztítására irányul, hanem sokkal inkább az inkubációs időszak meghosszabbodásában, a léziók számának és/vagy kiterjedésének csökkenésében, valamint a sporuláció mérséklődé- sében nyilvánul meg. Mindezek a tényezők együttesen a járvány gyengülését eredményezik. Mivel valamennyi rassz korlátozott mértékű sporulációra képes a horizontális rezisztenciájú genotípusok lombozatán, nincs szelekciós nyomás a kórokozóra, következésképp a védelem tartósnak ígérkezik, s ez a rezisztenciára nemesítés végső célja. A nem rassz-specifikus ellenállóságnak van néhány, főként nemesítéstechnikai és növényvédelmi szempontból hátrányos tulajdonsága:
1. Mennyiségi jellegként viselkedik, poligénes öröklődésű, ezért nehezebb a recesszív rezisztenciagének (r-gének) azonosítása és a kívánatos számú r-gén egyetlen genotípusban való egyesítése.
Emellett számolni kell nemkívánatos, kapcsolt tulajdonságok megjelené- sével is (pl. késői érés, minőségromlás);
2. A szántóföldi rezisztencia a kifejlett növényeken vizsgálható, ezért kérdé- ses a korai szelekció alkalmazhatósága a nemesítésben;
3. A P. infestans korlátozott mértékű, de hosszantartó sporulációja a lombo- zaton megnöveli a gumófertőzés lehetőségét. Emiatt legalább közepes mértékű horizontális rezisztencia kívánatos a lombon és a gumókon egyaránt, ami a részben független öröklődés miatt nehezen kombinálható;
4. A horizontális rezisztenciaformákra a környezeti tényezők nagyobb hatást gyakorolnak, mint a vertikális rezisztenciára.
Számos mexikói vad burgonyafaj fontos rezisztenciaforrásnak tekinthető a nem rassz-specifikus rezisztenciára nemesítésben, mint pl. a Solanum andigenum, S. phureja, S. demissum és a S. stoloniferum, melyek közül az utóbbi két faj R-géneket is hordoz (Király és mtsai 1970). Rivera-Pena (1992) a S. demissumban, S. verrucosumban, S. ispetalumban és a S. brachicarpumban igazolt horizontális rezisztenciát a fitoftórával szemben a mexikói Toluca völgyben. Ugyanitt 12 burgonyafajta és nemesítési klón szántóföldi rezisztenciáját értékelve az 575049-es számú klón, valamint a Michoacan, az Ireri és a Nortena fajták bizonyultak a legellenállóbbnak (Flores-Gutierrez 1997). Colon és munkatársai (1993) a burgonyavésszel szemben tartós rezisztenciát hordozó genotípusok után kutatva a kultúrfajjal jól keresztezhető, nem mexikói eredetű, 14 diploid vad faj többségében részleges-, néhány fajban pedig immunitásszintű ellenállóságot mutattak ki.
Annak ellenére, hogy a nem rassz-specifikus rezisztenciára nemesítés módszertanilag bonyolultabb és időigényesebb, mégis a tartósabb, kevésbé erodálódó ellenállóság reményében előnyt élvez napjaink nemesítési programjaiban (Bhattacharyya és Singh 1986, Darsow 1989, Mackay és mtsai 1992, Colon és mtsai 1995, Inglis és mtsai 1996, Oberhagemann és mtsai 1999).
2.4.3. A lombozat és a gumók fitoftóra-ellenállóságának összefüggése Az utóbbi néhány évtizedben, amióta a megbízhatóbbnak és tartó- sabbnak ítélt nem rassz-specifikus rezisztencia kialakításán fáradoznak a nemesítők, a lombozat mellett a gumók fitoftóra-ellenállósága is fölértékelő- dött. Ezt az a körülmény idézte elő, hogy a részleges ellenállóságot mutató fajták lombján a kórokozó bármely rassza korlátozott mértékű sporulációra képes. Bár a léziók száma és/vagy kiterjedése, illetve a spóratermelés intenzitása lényegesen kisebb, mint a fogékony genotípusokon, de a hosszabb időtartamú sporuláció alatt képződhet annyi fertőző anyag (sporangium), ami eső- vagy öntözővízzel a talajba mosódva gumófertőzést okozhat. Következésképpen a lombvesztésből és a gumórothadásból eredő károk mérséklése céljából arra kell törekedni, hogy az új fajták lombozata és gumója egyaránt legalább közepes rezisztenciaszintet mutasson.
Persze ezt a nemesítési célt sokkal könnyebb megfogalmazni, mint meg- valósítani. Számos vizsgálati eredmény alapján ugyanis a burgonyanövény különböző testtájain észlelt fogékonyság/rezisztencia között nincs szoros összefüggés (Erwin és mtsai 1983, Arora és mtsai 1985, Cristinzio és Testa 1999, Gergely 1999, 2000). Ez nem számít ritka jelenségnek, hiszen pl. a napraforgó (Helianthus annuus L.) – Sclerotinia sclerotiorum gazda-parazita kapcsolatban a szártő- és a tányérrothadással szembeni rezisztencia hasonló- képpen nagymértékű függetlenséget mutat (Vear és Tourvieille de Labrouche 1984, Békési és mtsai 2000, Gergely és Birtáné 2002).
Deahl és Sinden (1973) amikor R1-gént hordozó burgonyafajták lomb- ját és gumóját fertőzte a P. infestans 0-ás rasszával, az inkompatibilis kapcsolattól várt hiperszenzitív nekrózisok megjelentek ugyan a lombozaton, de a gumók – genotípustól függően – kisebb-nagyobb mértékben meg- betegedtek. A szakirodalomban egymásnak ellentmondó megállapítások is előfordulnak a lomb és a gumók ellenállóságának kapcsolatáról. Piotrowski (1975) szerint a lomb és a gumók fitoftórarezisztenciájának együttes elő- fordulása gyakoribb jelenség, mint ennek az ellenkezője. Ugyanakkor Collins és munkatársai (1999) interspecifikus keresztezésből származó diploid, hasadó burgonyapopulációban a két eltérő testtáj ellenállósága között negatív összefüggést mutattak ki. Mindenesetre a felmerülő ellentmondások az eltérő növényi részek genetikai különbségeire éppúgy visszavezethetők, mint a nem kellően standardizált értékelési módszerekre (Érsek és Bakonyi 1997).
2.4.4. A rezisztenciára nemesítés nem hagyományos módszerei
A nem hagyományos nemesítési módszerek közül a betegség- ellenálló (köztük a fitoftórarezisztens) alapanyagok előállítására is alkalmas eljárás a szomatikus hibridizáció, különösen azokban az esetekben, amikor a rezisztenciaforrásként felhasznált vad faj ivarosan nem keresztezhető a kultúrburgonyával. Sczerbakowa és munkatársai (2000) egy gumót nem fejlesztő, hexaploid vad faj, a Solanum nigrum L. fitoftórarezisztenciáját vitték át a kultúrfaj diploid hibridjeibe protoplasztfúzió segítségével. Ugyanezzel a módszerrel sikerült a Solanum circaeifolium subsp. circaeifolium Bitter faj fitoftóra- és fonálféreg- (Globodera pallida Pa2 és Pa3) ellenállóságát átül- tetni a kultúrburgonyába (Mattheij és mtsai 1992).
Napjainkban már egyre szélesebb körben használják fel az in vitro eljárásokat a burgonyanemesítésben, így pl. különösen nagy jelentőséget tulajdonítanak a burgonyavész elleni horizontális rezisztencia egyes komponenseire irányuló korai szelekciós módszereknek. A Nemzetközi Burgonyakutató Központban (CIP, Peru) molekuláris markerek segítségé- vel gyorsítják föl a horizontális fitoftórarezisztencia kialakításának szelek- ciós munkáit (Trognitz és mtsai 2000). Oberhagemann és munkatársai (1999) interspecifikus hibridizációból származó diploid burgonyaklónokban
a nem rassz-specifikus rezisztenciát meghatározó QTL-ek (mennyiségi tulajdonságot kódoló lókuszok) genetikai elemzése és DNS-alapú markerek szelekciós célra való felhasználásának vizsgálatakor azt tapasztalták, hogy a lombozat nagyfokú ellenállósága kapcsoltan öröklődik a mérsékelt gumórezisztenciával, a késői éréssel és a nagyobb vigorral. Érsek és Bakonyi (1997) szerint nagyszámú citoplazmatikus és nukleáris DNS- markerhez juthatunk olyan módszerekkel, mint az RFLP- és a PCR-analízis, továbbá az izozimelemzés is jól használható molekuláris markereket ered- ményez a P. infestans biológiájának alaposabb feltárásához.
Az öröklött ellenálló képességen kívül a szisztemikus szerzett reziszten- cia kialakításával is csökkenthetők a betegségek okozta kártételek, amelyben szerepet játszhat a növények előzetes fertőzése, vegyszeres előkezelése vagy mechanikai sérülések kiváltása (Goodman és mtsai 1991). Strömberg (1995) a P. infestansszal és a burgonyára nem patogén P. cryptogea fajjal előfertőzött, különböző horizontális rezisztenciájú burgonyafajtákban indukált szisztemikus szerzett rezisztenciát, amely a legellenállóbbnak ismert fajtában (Matilda) volt a legintenzívebb, 50-60%-kal csökkentve a burgo- nyavész másodlagos fertőzéséből származó léziók kifejlődését. Cohen és munkatársai (1991) telítetlen zsírsavakkal váltottak ki ismeretlen mechaniz- musú szisztemikus szerzett ellenállóságot a P. infestansszal szemben burgonyában. Abad és munkatársai (1997) transzgénikus burgonyanövények- ben indukáltak szisztemikus szerzett rezisztenciát, amely azonban csak a P. infestans US-1 rasszával szemben nyújtott kielégítő védelmet, viszont az US-8 rasszal szemben nem, s ráadásul a növények fogékonynak bizonyultak a burgonya X vírusra (PVX) és a baktériumos lágyrothadásra (Erwinia spp.) is.
A rezisztenciára nemesítés legújabb vívmányai a géntechnológiához fűződnek. Wu és munkatársai (1995) az Aspergillus niger gombafajból izo- lált glükózoxidáz-gént felhasználva állítottak elő transzgénikus burgonya- növényeket. Ennek az enzimnek a működése során – a D-glükóz oxidáció- ját katalizálva – hidrogénperoxid (H2O2) is képződik, melynek – egyéb reaktív oxigénfajtákkal együtt – fontos szerepet tulajdonítanak a növények korai védekezési folyamataiban. A szerzők azt tapasztalták, hogy a transz- formáns gumók ellenállósága nagyobb mértékű volt mind a baktériumos (Erwinia carotovora), mind a gombás (P. infestans, Verticillium sp.) fertőzé- sekkel szemben.
2.4.5. A rezisztens fajták szerepe az integrált növényvédelemben
A túlzott genetikai egyöntetűség nemcsak az élettelen környezeti tényezőkkel, hanem a kórokozók támadásával szemben is sebezhetőbbé teszi termesztett növényeinket. Így például a Bipolaris maydis (syn.
Helminthosporium maydis) T-rassza azért tudott súlyos járványt kiváltani 1970-ben az Egyesült Államokban, mert a termesztésben lévő kukorica- hibridek többsége a fogékonyságot meghatározó cms-T citoplazmát hordozta
anyai szülővonalában (Barabás 1979). Az 1845/46-os hírhedt burgonyavész- járvány idején Írországban egyetlen burgonyafajta (Lumper) uralta a közter- mesztést, amelynek nagymértékű fogékonysága társadalmi katasztrófához vezetett (Érsek és Bakonyi 1997). Ezek az események az egynemű fajtaszerkezet növényegészségügyi veszélyeire hívják föl a figyelmet. Amint a fajtaösszetételben megjelennek a betegség-ellenálló genotípusok, a járványok veszítenek pusztító erejükből és a gazdasági károk mérséklődnek.
Más szóval a járványelhárítás leghatékonyabb eszköze a rezisztens növény- fajták termesztése. Jelenleg Írországban a burgonya termőterületének meghatározó részén legkevesebb 10 fajtát termesztenek, melyek között fitoftórarezisztensek is vannak (G.R.Mackay,szóbeli közlés). A magasabb szintű genetikai védelem és a sokat fejlődött kémiai védekezés együttesen megfelelő biztonságot nyújt. Mindenesetre a burgonyavész kórokozójának rendkívüli genetikai változékonysága – amely mind a rezisztenciára nemesítés, mind pedig a vegyszeres és agrotechnikai védekezés hatékonyságát csökkentette – az utóbbi néhány évtizedben nyilvánvalóvá tette, hogy önmagában is kielégítő hatékonyságú növényvédelmi módszer nincs a betegséggel szemben. A hosszabb távon is eredményes, vagyis tartós és megbízható védelem csak egy minden részletében kidolgozott integrált védekezési rendszertől remélhető (Érsek és Bakonyi 1997, Grunwald és mtsai 2000). Ennek sikere azon is múlik, hogy mennyire ellenállóak a termesztett burgonyafajták. Grunwald és munkatársai (2000) előrejelzésre alapozott fungicidhasználatot és nagyfokú szántóföldi rezisztenciával bíró fajtákat (pl. Rosita, Nortena) ajánlanak a burgonyavész elleni integrált védekezési rendszerbe, a mexikói Toluca-völgyben. A nem rassz-specifikus (horizontális) rezisztenciájú burgonyafajták fokozatos térnyerése fontos szerepet játszhat a növényvédőszerfelhasználás csökkentésében is. Fry (1978) a burgonyafajták horizontális rezisztenciáját mennyiségileg, a fungicid- kezelések csökkentésének mértékében határozta meg, amely szerint a leg- ellenállóbb és a legfogékonyabb klón szántóföldi rezisztenciája közti különbséget egyenértékűnek ítélte 0,7 kg/ha mankozeb hatóanyag heten- kénti kijuttatásával. Clayton és Shattock (1995) kísérleteiben a magasszintű szántóföldi rezisztenciát mutató fajták állományában 20-80%-kal lehetett csökkenteni a burgonyavész elleni fungicidkezelések számát a fogékony fajtákhoz képest. Shtienberg és munkatársai (1999) a fajták rezisztenciáján és a specifikus fungicideken alapuló integrált védekezési módszer haté- konyságát hasonlították össze a burgonyavész és az alternáriás betegség elleni hagyományos védekezéssel. Az integrált védekezést reprezentáló kezelésekben a gumótermés vagy nem tért el a hagyományos kezelésekétől, vagy szignifikánsan nagyobb volt azokénál. Wallace (1993) értékelése alapján az integrált növényvédelem (IPM) amellett, hogy hatékonynak és gazdaságosnak bizonyult, kiemelkedő eredményeket ért el világszerte a növényvédőszerhasználat csökkentésében, az élelmiszerbiztonságban, vala- mint a vízkészlet és az élővilág megóvásában. Szerepének felértékelődését
jelzi, hogy az Egyesült Államokban 1992-ben társadalmi összefogással megalakult a mezőgazdaság és élelmiszeripar, a környezetvédő szervezetek és a kormány részvételével a Nemzeti Koalíció az Integrált Növényvédelemért (NCIPM) nevű szerveződés, melynek fő célja a környezetbarát növény- védelmi módszerek fejlesztése és támogatása. Mivel a burgonyavész tekinthető a világ élelmiszeripari termelését sújtó egyik legköltségesebb biotikus tényezőnek, 1996-ban létrehoztak egy globális kutatási és technoló- giafejlesztési együttműködést (Global Initiative on Late Blight, GILB) a betegség okozta károk mérséklésére (CIP 2003). A GILB fontos feladatának tartja a tartós fitoftórarezisztenciát mutató burgonyafajták termesztésének ösztönzését.
2.5. Környezeti tényezők hatása az ellenálló képességre
A környezeti tényezők nemcsak a burgonyafajták növekedésére és fejlődésére hatnak, hanem különböző mértékben befolyásolhatják a rezisz- tenciagének kifejeződését is. Bizonyos rezisztenciaformákra (mint pl. a főgénes fitoftórarezisztencia vagy a PVX és PVY-immunitás) kevésbé hatnak, ugyanakkor más, főként mennyiségi (poligénes) öröklődésű rezisztencia- típusokra (pl. horizontális fitoftórarezisztencia) nagyobb befolyást gyakorolnak.
Az élettelen (abiotikus) környezeti tényezők közül a hőmérséklet és a vízellátás jelentősen befolyásolhatja a növények ellenálló képességét. Klement és munkatársai (2003) szerint a növények baktériumokkal szemben meg- nyilvánuló általános, nem-specifikus ellenálló képessége szélsőségesen alacsony hőmérsékleten, 10 °C alatt csak gyengén vagy már egyáltalán nem működik. S mivel az általános védekezési mechanizmus a növény fehérje- szintézisének függvénye, az élettanilag idős levelekben (az érés közeled- tével) már nem nyújt kielégítő védelmet. Stewart és munkatársai (1993) a termesztőközeg nedvességtartalmának hatását vizsgálták különböző rezisztenciaszintű burgonyafajták gumóinak fitoftóra iránti fogékonyságára, üvegházi tenyészedényes kísérletben. A háromféle kezelés (száraz, nyirkos, nedves) közül – a várakozással ellentétben – a virágzás időszakától szárazon tartott közegből származó gumók szignifikánsan fogékonyabbaknak bizo- nyultak, mint a másik két kezelésben neveltek. A vízellátottsági szint a fogékony fajta (Bintje) gumófertőzöttségére kisebb hatást gyakorolt, mint az ellenállóbb fajtákéra. A szerzők szerint többféle magyarázat lehetséges a gumók megnövekedett fogékonyságára: (1) a gumó felületén élő antagonista szervezetek hatását gátolta a nedvességhiány, versenyelőnybe hozva a kórokozót, (2) a vízhiány okozta stressz a gumószövetek rezisztenciájának csökkenéséhez vezetett, (3) a szárazság valamilyen módon mérsékelte a gumóhéjnak, mint mechanikus gátnak a hatékonyságát a fertőzéssel szemben.
A megvilágítás (nappalhosszúság) nemcsak az érést befolyásolja, hanem az ellenálló képességre is hat. Így például a hosszúnappalos körül- mények között nagyfokú szántóföldi rezisztenciát mutató burgonyafajtáknak
rövidnappalos környezetben megnőhet a burgonyavész iránti fogékonysága (Beukema és van der Zaag 1990). Umaerus (1959) szerint nyilvánvaló pozitív összefüggés van a Solanum tuberosum eredetű fajták burgonya- vésszel szembeni szántóföldi rezisztenciája és a levelek peroxidázaktivitása között. Mivel az enzimaktivitás a napi megvilágítás függvényében változik, ugyanazon fajták (pl. Alpha, Sebago) hosszúnappalos megvilágításban – a peroxidázszint emelkedésével – nagyfokú szántóföldi rezisztenciát mutattak, rövidnappalos körülmények között viszont (pl. Mexikóban) fogékonynak bizonyultak. Darsow és munkatársai (1988) szerint a növények kora és a különböző környezeti tényezők (megvilágítás, vízellátás, stb.) kisebb-nagyobb mértékben befolyásolták a Solanum demissum és a Solanum stoloniferum származékok leveleinek nem rassz-specifikus fitoftórarezisztenciáját üvegházi kísérletekben. A hajlamosító környezeti tényezők hirtelen megváltozása csökkentette a vizsgálat megbízhatóságát, ezért a szerzők többéves értékelést tartanak szükségesnek a szántóföldi rezisztencia pontos meg- határozásához.
A tápanyagellátás és a betegségek iránti fogékonyság összefüggésé- nek vizsgálatakor arra a megállapításra jutottak (Moeller és mtsai 1999, Zadoks és Pennings 1999), hogy a növények nagyadagú N-táplálása fokoz- hatja a burgonyavész okozta megbetegedések súlyosságát és ezáltal a termésveszteségeket. Mudich (1981) savanyú homoktalajon beállított kísér- leteiben (Mariettapuszta, Vadaspuszta) a 100 kg/ha hatóanyagtartalmú alap- trágyadózisnál (N:P:K = 1:1:1) állapította meg a legnagyobb ellenállóságot a lombozat és a gumók fitoftórás megbetegedésével szemben, viszont a 300 kg/ha-os adagnál már a fogékonyság növekedését tapasztalta. Erdélyi (1983) különböző összetételű levéltrágyák hatását vizsgálta a lomb és a gumók burgonyavész-rezisztenciájára és arra a megállapításra jutott, hogy a mikroelemeket nagyobb koncentrációban tartalmazó készítmények (pl.
Plantán, H-92 E) növelték a legnagyobb mértékben az ellenállóságot.
A biotikus környezeti elemek közül Darsow és Wulfert (1989) a vírusfertőzések (PVX, PVY, PVM, PLRV) hatását vizsgálták néhány burgonya- fajta lombjának burgonyavésszel szembeni szántóföldi rezisztenciájára. A vírusfertőzések általában mérsékelt hatást gyakoroltak a fitoftóra-ellenálló- ságra, amelynek szintje inkább növekedett a vírusmentes növényekéhez képest, de a kölcsönhatást sok tényező befolyásolta (fajta, évjárat, vírusfaj, stb.). Egyéb vizsgálati eredmények is azt támasztják alá, hogy egyes burgonyapatogén vírusokkal (pl. PVX, PVY) való előfertőzés csökkentette a növények fitoftórás megbetegedésének súlyosságát (Hossain és mtsai 1998, Li és mtsai 1999). Umaerus (1959) a burgonya X vírussal fertőzött növények- ben 1,6 - 1,8-szor nagyobb peroxidázaktivitást mért, mint az azonos anya- gumóból származó vírusmentes növényekben. A nagyobb peroxidázaktivitás fokozta a fitoftórával szembeni szántóföldi rezisztenciát, ami magyarázatot ad arra, hogy az elsődleges vírusfertőzésen átesett növényeken miért nem képes a P. infestans súlyos megbetegedést kiváltani.
Connolly és munkatársai (1995) az eltérő fogékonyságú burgonyafajták közötti szomszédhatást elemezték a lombozat fitoftóra-ellenállóságának értékelésére szántóföldi kísérletben. Megállapították, hogy egy adott geno- típus fertőzöttségének mértékét befolyásolja a szomszédos genotípusok fogékonysága vagy rezisztenciája. Így például súlyosan fertőzött, fogékony növények szomszédságában a rezisztens változatok a vártnál nagyobb mértékben betegedtek meg és fordítva: rezisztens szomszédok hatására a fogékony fajták fertőzöttsége mérsékeltebbnek bizonyult. Mindenesetre a kölcsönhatások varianciaanalízise nem tárt fel szignifikáns különbségeket, tehát a genotípusok relatív fogékonysági rangsora nem változott meg.
Az előzőekben hivatkozott vizsgálati eredmények kétséget kizáróan igazolják, hogy a környezeti tényezők jelentősen módosíthatják a növények betegségekkel szembeni ellenálló képességét.
3. ANYAG ÉS MÓDSZER 3.1. A rezisztenciavizsgálatok anyaga és helyszínei
Valamennyi vizsgálat anyagát az Országos Mezőgazdasági Minősítő Intézethez (OMMI) fajtaelismerés céljából bejelentett burgonya-fajtajelöltek és/vagy az államilag elismert burgonyafajták képezték. A horizontális rezisztencia mértékét meghatározó vizsgálatokban a felhasznált fertőző anyag minden esetben hazai Phytophthora infestans izolátumok keverékéből állt és legkevesebb 7 virulenciafaktort tartalmazott, megfelelően reprezen- tálva a honi populációk összetett rasszkarakterét (1. táblázat). Az összetett virulenciájú fertőző anyag használatát azok a vizsgálatok indokolják, amelyek a legutóbbi években feltárták a P. infestans magyarországi populá- ciójának rasszösszetételét. Ezek szerint a hazai izolátumok döntő többsége komplex virulenciájú és legalább 4-7, vad Solanum fajokból származó rezisz- tenciagén (R-gén) letörésére képes a burgonyában (Bohár és mtsai 1999, Érsek 2001).
A lombozat fitoftóra-ellenállóságának vizsgálatát az OMMI Fajtakiter- mesztő Állomásán, Monorierdőn (fóliasátras teszt), valamint az OMMI Növénykórtani Állomásán, Röjtökmuzsajon (szabadföldi teszt) végeztük, külön erre a célra szolgáló provokációs kísérletekben.
A gumók fitoftórás megbetegedésének mértékét a fajtakísérletekben előforduló természetes fertőzések nyomán állapítottuk meg Röjtökmuzsajon azokban az évjáratokban, amelyekben a késői lombfertőzésből származó inokulum, valamint az ősz eleji csapadék mennyisége értékelhető mértékű gumófertőzést indukált. Mivel a spontán gumófertőzésekhez szükséges környezeti feltételek 10 évből csak 3 évben teljesültek, egyéb laboratóriumi módszereket is be kellett vonni a rezisztenciavizsgálatokba (egészgumós mesterséges fertőzés, gumókorongok elicitoros kezelése).
3.2. A rezisztenciavizsgálatok módszerei
3.2.1. A lombozat fitoftóra-ellenállóságának vizsgálata fóliasátras provokációs kísérletekben
A hazai fajtaminősítés jelenlegi gyakorlata szerint a burgonyafajta- jelöltek gazdasági értékének meghatározása 3 vizsgálati év és 6 termőhely kisparcellás fajtakísérleti eredményein alapszik. Mivel az ún. teljesítménykí- sérletek átlagos (üzemi) vegyszeres védelemben részesülnek, alkalmatla- nok a gombás lombbetegségek (köztük a burgonyavész) elleni rezisztencia vizsgálatára. Ezért szükségessé vált egy olyan provokációs kísérlet kidol- gozása, amellyel nagyobb számú genotípus lombozatának ellenállósága viszonylag rövid idő alatt (kb. 50 nap), kevés költséggel, de megbízhatóan vizsgálható. Erre a célra a fóliasátor alatti provokációs kísérlet őszi időszakban
(szeptember-október) való alkalmazását választottuk, amely optimálisan ki- elégíti a kórfolyamat hőmérsékleti (15-20 °C) és relatív páratartalom (95-100%) iránti igényeit.
Az ún. egésznövény-fertőzési módszer leírása a következő: a vizsgá- landó genotípusok 1. szaporítási fokú gumóiból 60-60 db rügydugványt vágtunk ki, amelyeket a rügynyugalom vegyszeres megszakítása (0,1 %-os gibberellinsav (GA3) + 1%-os tiokarbamid oldatában 10 perces áztatás, majd csapvizes lemosás) után fóliasátorban kialakított tőzegágyba ültettük ki (30 növény/parcella), kétismétléses, véletlenblokk-elrendezésű, mikroparcellás kísérletbe (7. ábra). Fertőző anyagként a P. infestans két hazai izolátumának (H-K/91, H-3.1/93) 1:1 arányú keverékét használtuk, amelyek komplex rassz- karakterűek és legalább 7 R-gén letörésére képesek (1. táblázat). Mivel az izolátumok mindkét (A1 és A2) párosodási típust képviselik, az ivaros kölcsön- hatásból eredő potenciális veszélyek (oospóraképződés) miatt a kísérleti anyagokat és a termesztőközeget a vizsgálatok végeztével megsemmisítettük.
Az izolátumokat egész gumón (OMMI, Budapest) illetve paradicsomnövények lombozatán (Heves megyei NTSZ, Eger) tartottuk fenn és fogékony burgonyafajták lombján szaporítottuk fel (8. ábra). A mesterséges fertőzést 3x104 sporangium/ml sűrűségű vizes szuszpenzió lombra permetezésével végeztük a növények 10-14 lombleveles korában, a késő délutáni órákban (30 ml/parcella). A milliliterenkénti sporangiumszámot Bürker-kamra segít- ségével határoztuk meg. A fertőzöttség mértékét 1-10-es skálájú bonitálás- sal állapítottuk meg az inokulációt követő 14-18. napon, fajtánként meg- becsülve a megbetegedett levélfelület nagyságát az összes lombfelülethez képest. A fertőzöttség mértékét a fertőzött lombfelület %-ában adtuk meg (Gergely 1999).
A genotípusokat fitoftórás lombfertőzöttségük alapján 5 fogékonysági osztályba soroltuk be. A rezisztenciavizsgálatainkban általánosan használt módszer szerint a besorolás az egyes fajták fertőzöttségének a mindenkori kísérleti átlagtól való eltérésén alapszik. Így a kísérleti átlagot 100%-nak tekintve, egy adott genotípus rezisztens (1), ha fertőzöttsége a kísérleti átlag értékének 25%-át nem haladja meg, mérsékelten rezisztens (2), ha fertőzöttsége a kísérleti átlag értékének 25-75%-a, közepesen fogékony (3), ha fertőzöttsége a kísérleti átlag értékének 75-125%-a, közepesnél fogékonyabb (4), ha fertőzöttsége a kísérleti átlag értékének 125-175%-a és nagyon fogékony (5), ha a fertőzöttsége meghaladja a kísérleti átlag 175%-át. Azokban az esetekben, amikor a fertőzöttség kísérleti átlaga nagyobb 50%-nál, a besoroláskor a tényleges fertőzöttség százalékos értékei a rezisztencia/fogékonyság következő fokozatainak felelnek meg:
(1) 0-20% = rezisztens,
(2) 21-40% = mérsékelten rezisztens, (3) 41-60% = közepesen fogékony, (4) 61-80% = közepesnél fogékonyabb,
(5) 81-100% = nagyon fogékony (Hinfner és Homonnay 1966).
7. ábra Fóliasátor alatti rezisztenciavizsgálat (Monorierdő)
8. ábra A Phytophthora infestans fenntartása egész gumón
A lombfertőzöttség vizsgálatakor fajtánként értékeltük a P. infestans sporulációjának mértékét is, három kategóriát alkalmazva:
(1) gyenge: gyér, elszórt sporangiumtartó-megjelenés,
(2) közepes: mérsékelt intenzitású sporangiumtartó-képződés, (3) erős: dús, egybefüggő sporangiumtartó-gyep kialakulása.
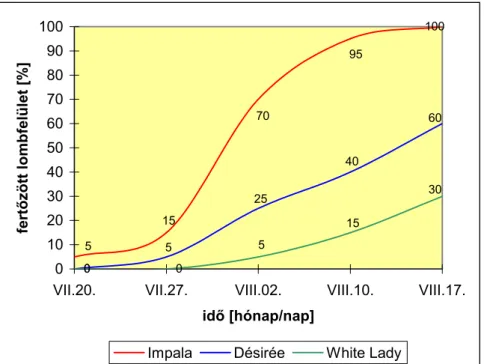
A több mint 10 éves rezisztenciavizsgálati eredményeink birtokában összehasonlítottuk néhány hazai és külföldi burgonyafajta (n=18) egymástól távoli évjáratokból származó fitoftórás lombfertőzöttségét a horizontális rezisz- tencia tartósságának megítéléséhez. Az összehasonlíthatóságot az tette lehetővé, hogy a fogékony (Cleopatra) és a közepesen fogékony (Désirée) standard fajták a teljes vizsgálati időszakban szerepeltek a provokációs kísérletekben.
A legutóbbi 5 éves időszakban (1998-2002) 60 burgonya-fajtajelölt és államilag elismert fajta lombozatának horizontális rezisztenciáját vizsgáltuk a leírt módon.
3.2.2. A lombozat fitoftóra-ellenállóságának vizsgálata szabadföldi provokációs kísérletekben
A 3.2.1. pontban leírt fóliasátras provokációs vizsgálat, amellyel jól elkülöníthetők a genotípusok burgonyavésszel szembeni főbb viselkedési típusai (fogékony, átmeneti, rezisztens), kevésbé alkalmas a mennyiségi jellegű szántóföldi rezisztencia mérésére. Ezért ennek a horizontális rezisztenciatípusnak a pontosabb meghatározására a Shaner és Finney (1977) által leírt ADPC (Area under the Disease Progress Curve) módszert alkalmaztuk. Ez az vizsgálati módszer a betegség lombozaton való kifejlő- désének és terjedésének ütemét a járványgörbe alatti terület nagyságával méri az integrálszámítás elve alapján.
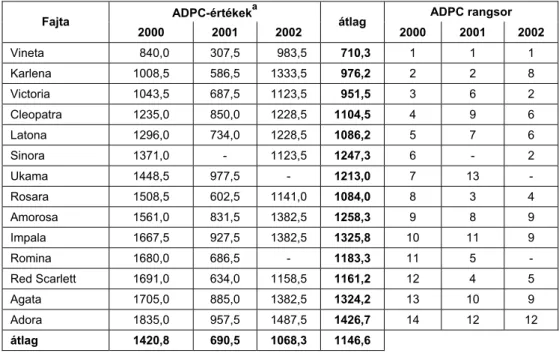
Három éven át (2000-2002) 36 államilag elismert burgonyafajta lombozatának szántóföldi rezisztenciáját vizsgáltuk fungicidmentes, kétismét- léses, mikroparcellás kísérletekben (10 növény/parcella). A vizsgálandó genotípusok blokkjait a vírusellenálló Százszorszép fajtával ültettük körbe (belső fertőzési forrás), amelyet mesterségesen fertőztünk a P. infestans 3 hazai izolátumának (H-K/91, 2001/04, 2001/21/3) 1:1:1 arányú keverékével (1. táblázat). Az inokulációt a lombzáródás után 1-2 héttel végeztük el a növényállomány előzetes beöntözését követően, 2-3 x 104 sporangium/ml sűrűségű vizes szuszpenzió lombra permetezésével, az esti órákban. A kísérletekben 3 eltérő fogékonyságú kontroll fajtát szerepeltettünk: a White Lady-t (1=rezisztens), a Désirée-t (3=közepesen fogékony), és az Impalát (5=nagyon fogékony). A lombozat fertőzöttségét 5-8 naponkénti bonitálással állapítottuk meg (fertőzött lombfelület %) az első tünetek fellépésétől a vegetáció végéig, évjárattól függően 4-6 alkalommal.
A szántóföldi rezisztencia mértékének kifejezésére használt ADPC- értékeket a következő képlet alapján számoltuk ki:
n
ADPC= ∑ [(Xi+1 + Xi)/2] [ti+1 - ti] ahol
i = 1
xi = a fertőzöttség mértéke az i-edik megfigyeléskor, ti = az időpont (napokban) az i-edik megfigyeléskor,
n = a megfigyelések száma (Shaner és Finney 1977).
A nagy ADPC-értékek gyors betegségkifejlődésre, azaz fogékonyságra utalnak (meredek járványgörbe), míg a kisebb értékek a szántóföldi reziszten- cia különböző fokozatait jelzik (lassú kórfolyamat, elnyújtott járványgörbe).
A genotípusok fogékonysági osztályokba való besorolását a legfogé- konyabb fajtához viszonyított relatív ADPC-értékek (RADPC) alapján vé- geztük el az alábbiak szerint (Jenkins és Jones 2003):
(1) = 0,00 - 0,20 (rezisztens)
(2) = 0,21 - 0,40 (mérsékelten rezisztens) (3) = 0,41 - 0,60 (közepesen fogékony) (4) = 0,61 - 0,80 (közepesnél fogékonyabb) (5) = 0,81 - 1,00 (nagyon fogékony)
1. táblázat
A rezisztenciavizsgálatokban felhasznált Phytophthora infestans izolátumok jellemzése
(Bohár és mtsai 1999, Nagy és mtsai 2003)
Izolátum Virulenciatípus Párosodási típus Gazdanövény H-K/91 1.2.3.4.7.10.11 A2 paradicsom H-3.1/93 1.3.4.7.(8).10.11 A1 burgonya
2001/04 nem vizsgált A2 burgonya
2001/21/3 nem vizsgált A1 burgonya
A szabadföldi provokációs kísérletek legfontosabb agronómiai adatait a 2. táblázat tartalmazza.
2. táblázat
A szabadföldi provokációs kísérletek fontosabb agronómiai adatai (Röjtökmuzsaj)
Agrotechnikai elem/művelet 2000 2001 2002
Elővetemény Őszi búza Kukorica Kukorica
Tápanyagellátás - szervestrágya (t/ha) 40 (1999. 08. 09.)
- műtrágya NPK (kg/ha) 102-63-240 164-105-85 39-39-63 Talajfertőtlenítés (kg/ha) Thimet 10G (15) Counter 5G (25) Ültetési idő (hónap/nap) 04. 18 04. 27 05. 03 Gumócsávázás (l/ha) Ültetéssel egymenetben, Persite (2,0 - 2,5) Gyomirtás (kg, l/ha) Patoran 50 WP (3,0) + Dual Gold (1,5), preemergensen Lombvédelem Fungicides és inszekticides kezelés nem volt
Öntözés (mm) 20 100 160
Talajtani jellemzők
Talajtípus Öntés réti talaj (karbonátos)
pH (KCl) 7,1
KA 43
Összes só (m/m%) 0,0
Szénsavas mész (m/m%) 3,0
Humusz (m/m%) 2,72