Avarlebontási kísérletek dombvidéki kisvízfolyásokon
DOKTORI ÉRTEKEZÉS
Kovács Kata
Témavezető:
Prof. Dr. Padisák Judit, intézetigazgató egyetemi tanár, az MTA doktora Pannon Egyetem, Környezettudományi Intézet,
Limnológia Intézeti Tanszék
Pannon Egyetem
Kémiai és Környezettudományi Doktori Iskola Veszprém
2012
Avarlebontási kísérletek dombvidéki kisvízfolyásokon
Írta:
Kovács Kata
Készült a Pannon Egyetem Kémiai és Környezettudományi Doktori Iskolája keretében Témavezető: Prof. Dr. Padisák Judit
Elfogadásra javaslom (igen / nem)
……….
(aláírás) A jelölt a doktori szigorlaton …... % -ot ért el,
Az értekezést bírálóként elfogadásra javaslom:
Bíráló neve: ……… ………..igen /nem
……….
(aláírás) Bíráló neve: ……… ………..igen /nem
……….
(aláírás) A jelölt az értekezés nyilvános vitáján …...%-ot ért el.
Veszprém,
……….
a Bíráló Bizottság elnöke A doktori (PhD) oklevél minősítése…...
………
Az EDHT elnöke
1
Tartalomjegyzék
Rövidítések jegyzéke ... 3
Ábrák jegyzéke ... 4
Táblák jegyzéke ... 6
Kivonat ... 7
Abstract ... 8
Zusammenfassung ... 9
1 Bevezetés ... 10
1.1 Hazai avarlebontási kísérletek áttekintése ... 14
2 Célkitűzés ... 16
2.1 Avarbomlás és medermorfológia ... 16
2.2 Három, patakparti vegetációt alkotó fafaj avarjának bomlása ... 16
2.3 Avarbomlás hőmérsékletfüggése ... 16
2.4 Saját fejlesztésű terepi eszköz hatékonyságának összevetése a hagyományos módszerekkel ... 16
2.5 Avarbomlás kezdeti szakaszának vizsgálata ... 17
3 Hipotézisek ... 18
3.1 Avarbomlás és medermorfológia ... 18
3.2 Három, patakparti vegetációt alkotó fafaj avarjának bomlása ... 18
3.3 Avarbomlás hőmérsékletfüggése ... 18
3.4 Saját fejlesztésű terepi eszköz hatékonyságának összevetése a hagyományos módszerekkel ... 18
3.5 Avarbomlás kezdeti szakaszának vizsgálata ... 19
4 Anyag és módszer ... 20
4.1 Mintavételi helyszínek ... 20
4.2 A kísérletek felépítése, általános terepi kivitelezése ... 22
4.3 A minták feldolgozása ... 23
4.4 Adatok feldolgozása ... 24
4.5 Alkalmazott statisztikai módszerek ... 24
4.6 Vízkémiai minták feldolgozása ... 25
4.7 Kísérleti módszerek és körülmények egyedi jellegzetességei ... 25
4.7.1 Avarbontás és medermorfológia ... 25
4.7.2 Három, patakparti vegetációt alkotó fafaj avarjának bomlása ... 26
4.7.3 Az avarbomlás hőmérsékletfüggése ... 26
4.7.4 Saját fejlesztésű terepi eszköz hatékonyságának összevetése a hagyományos módszerekkel ... 27
4.7.5 Avarbontás kezdeti szakaszának vizsgálata ... 28
5 Eredmények ... 29
5.1 Avarbomlás és medermorfológia ... 29
5.1.1 Avarbomlás üteme ... 29
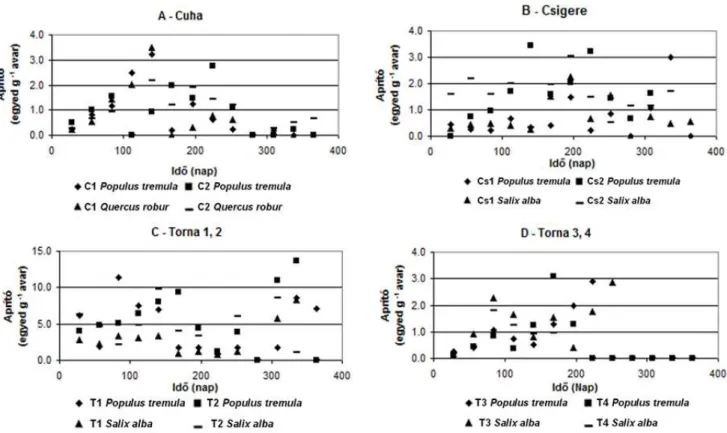
5.1.2 Aprító szervezetek ... 30
5.2 Három, patakparti vegetációt alkotó fafaj avarjának bomlása ... 32
5.2.1 Avarbomlás üteme ... 32
5.2.2 Aprító szervezetek ... 33
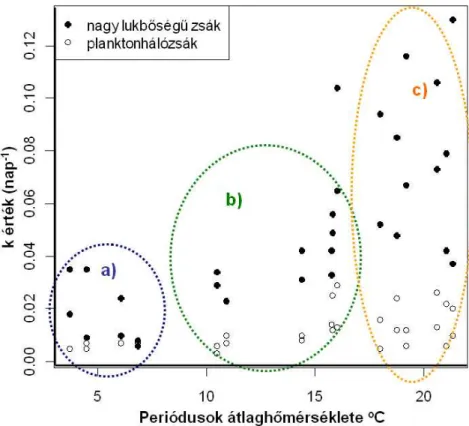
5.3 Avarbomlás hőmérsékletfüggése ... 35
5.3.1 Avarbomlás üteme ... 35
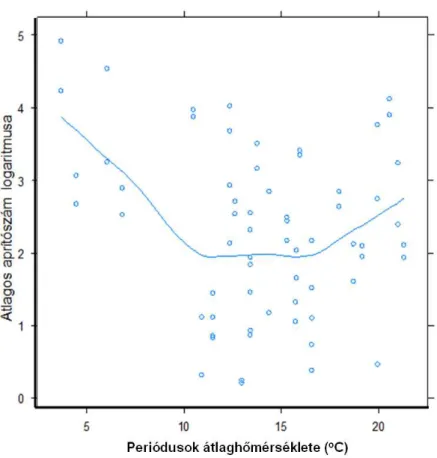
5.3.2 Aprító szervezetek ... 44
2
5.4 Saját fejlesztésű terepi eszköz hatékonyságának összevetése a hagyományos
módszerekkel ... 47
5.4.1 Avarbomlás üteme ... 47
5.4.2 Aprító szervezetek a különböző eszközökben ... 49
5.5 Avarbomlás kezdeti szakaszának vizsgálata ... 51
5.5.1 Avarbomlás ... 51
5.5.2 Aprító szervezetek ... 52
5.6 A patakok vizének fiziko-kémiai paraméterei ... 53
6 Eredmények értékelése ... 54
6.1 Avarbomlás és medermorfológia ... 54
6.2 Három, patakparti vegetációt alkotó fafaj avarjának bomlása ... 55
6.3 Avarbomlás hőmérsékletfüggése ... 58
6.4 Saját fejlesztésű terepi eszköz hatékonyságának összevetése a hagyományos módszerekkel ... 60
6.5 Avarbomlás kezdeti szakaszának vizsgálata ... 61
6.6 A patakok vizének fiziko-kémiai paraméterei ... 64
7 Gyakorlati alkalmazhatóság ... 66
8 Összefoglalás ... 68
9 Köszönetnyilvánítás ... 70
10 Irodalomjegyzék ... 71
11 Tudományos tevékenység adatai ... 81
12 Eredmények tézisszerű összefoglalása ... 84
12.1 Avarbomlás és medermorfológia ... 84
12.2 Három, patakparti vegetációt alkotó fafaj avarjának bomlása... 84
12.3 Avarbomlás hőmérséklet függése ... 85
12.4 Saját fejlesztésű terepi eszköz hatékonyságának összevetése a hagyományos módszerekkel ... 85
12.5 Avarbomlás kezdeti szakaszának vizsgálata ... 86
13 Results of the studies ... 87
13.1 Leaf litter decomposition versus bed morphology ... 87
13.2 Leaf litter decomposition of three tree species from the riparian vegetation ... 87
13.3 Leaf litter decomposition versus temperature... 88
13.4 Newly introduced tool, leaf litter cylinder and its comparison to existing tools . 88 13.5 Examination of the early stages of leaf litter decay ... 89
14 Függelék I. ... 90
3
RÖVIDÍTÉSEK JEGYZÉKE
CPOM: Coarse Particulte Organic Matter (durva szemcsés szerves anyag) FPOM: Fine Particulate Organic Matter (finom szemcsés szerves anyag) WFD: Water Framework Directive
VKI: Víz Keretirányelv MSZ: Magyar Szabvány Avg: Avarage, Átlag
StdDev: Standard Deviation, Szórás DO: Dissolved Oxigen, Oldott oxigén COND: Conductivity, vezetőképesség
CODPS: Chemichal Oxigen Demand, permanganátos kémiai oxigénigény (KOI) PCA: Principal Component Analysis
Qbap: magyarországi víztest típusokra kidolgozott folyóvizek ökológiai állapotának makroszkopikus vízi gerinctelen fajegyüttes alapján történő minősítésére szolgáló index Avar-input: allochton avarterhelés
C1: Cuha-patak első mintavételi helyszínének jelölése C2: Cuha-patak második mintavételi helyszínének jelölése T1: Torna-patak első mintavételi helyszínének jelölése T2: Torna-patak második mintavételi helyszínének jelölése T3: Torna-patak harmadik mintavételi helyszínének jelölése T4: Torna-patak negyedik mintavételi helyszínének jelölése Cs1: Csigere-patak első mintavételi helyszínének jelölése Cs2: Csigere-patak második mintavételi helyszínének jelölése VpS: Veszprémi-séd mintavételi helyszínének jelölése
VzS: Vázsonyi-séd mintavételi helyszínének jelölése Pt: Populus tremula
Sa: Salix alba Qr: Quercus robur
4
ÁBRÁK JEGYZÉKE
1. ábra A kísérlet helyszínei ... 21 2. ábra Mintázási sorozatok időbeli átfedései mind a téli, mind a nyári periódus alatt. A sorozatok az egyes rácsokat jelentik, és az azokon látható kisebb függőleges vonalak az egyes mintavételi alkalmakat jelzik, melyeket könnyen fel lehet vetíteni a felső „vonalzó”
sorra. ... 26 3. ábra Az avarhenger rögzítése a patak alján ... 28 4. ábra Az aprítók relatív mennyisége (egyed g száraz avar-1) a kétféle avarral töltött
zsákokban ... 31 5. ábra Három patakban a három avartípus bomlási üteme (a.: Csigere-patak, b: Torna- patak, c: Veszprémi-séd) ... 32 6. ábra A három patakban (Csigere-patak, Torna-patak, Veszprémi-séd) az aprítók
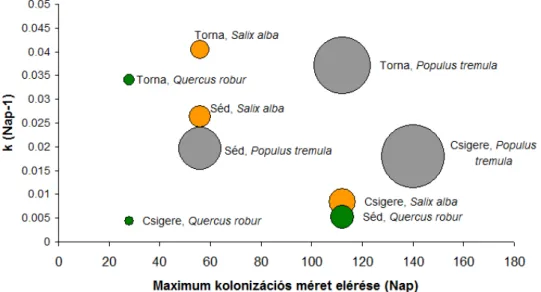
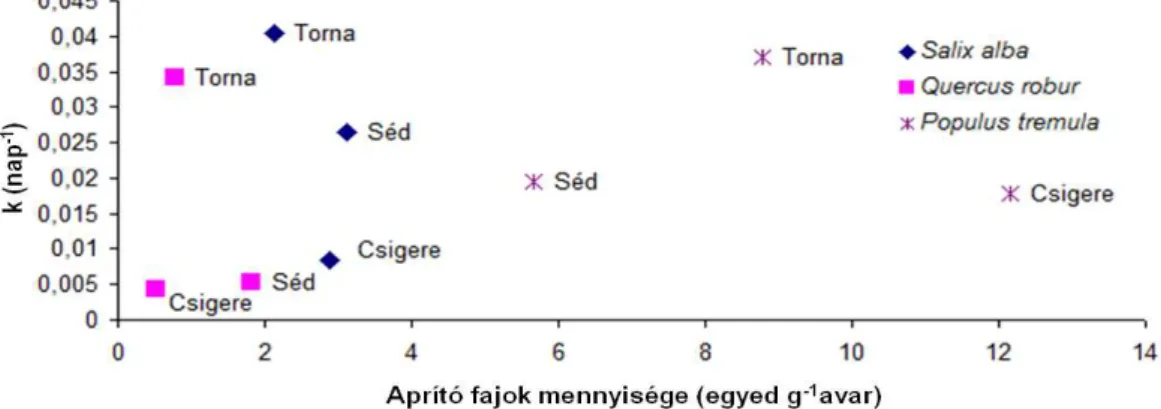
legnagyobb kolonizációjának, az idő és a k együttható összefüggése (A körök területe a kolonizáció mértékével arányos. Narancssárga szín jelöli a Salix alba, a zöld szín a Quercus robur, a szürke szín pedig a Populus tremula avart.) ... 34 7. ábra Az aprítóegyedek átlagos mennyisége (1 g száraz avarra vonatkoztatva) és a k együttható összefüggése) ... 35 8. ábra A k-érték alakulása a hálótípus függvényében (A boxplot ábrán a vastag vízszintes vonal a k-értékek mediánját, a doboz alsó és felső éle az alsó és felső kvartilist, a bajuszok végei pedig az adatok kiugró értékek nélküli tartományát jelölik. A kiugró értékeket kis körök jelzik.) ... 35 9. ábra A k-érték eloszlása a két vizsgálati időszak függvényében (A boxplot ábrán a vastag vízszintes vonal a k-értékek mediánját, a doboz alsó és felső éle az alsó és felső kvartilist, a bajuszok végei pedig az adatok kiugró értékek nélküli tartományát jelölik. A kiugró
értékeket kis körök jelzik.) ... 36 10. ábra A k értékek és a hőmérséklet (az adott rács vízben töltött ideje alatt mért
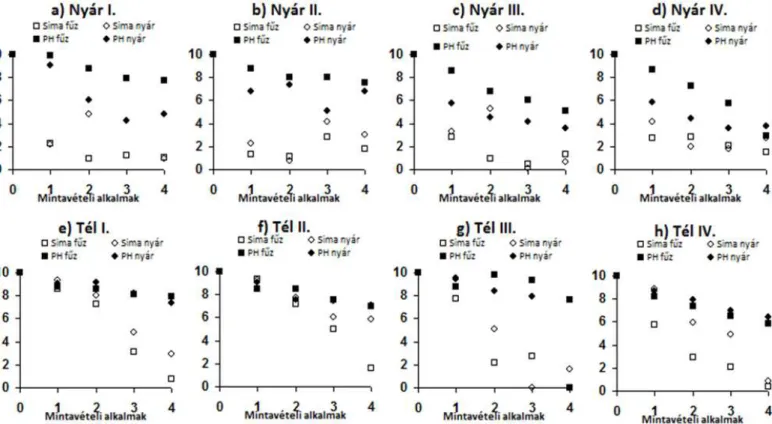
átlaghőmérséklete) alapján készített diagram ... 37 11. ábra A zsákokban talált aprítók átlagos mennyiségének alakulása a zsákok expozíciós ideje alatti átlaghőmérséklet összefüggése. Az y tengelyen az összes aprító egyedszám (n) természetes alapú logaritmusa van ábrázolva, az adatpontokra illesztett vonal a kvadratikus regressziós görbe. ... 38 12. ábra A Csigere-patak nyári és téli időszakában folytatott kísérletek avarcsökkenésének üteme (A mintavételi alkalmak kéthetente történtek. Az üres jelölők minden esetben a sima avarzsákot jelentik, míg a feketék a planktonháló anyagból készült zsákokból történő avarfogyást jelenítik meg. A római számok az egyes időben egymáshoz képest elcsúsztatott mérési periódusokat jelentik, melyek átfedése a 2. ábrán láthatóak.) ... 39 13. ábra A Vázsonyi-séd nyári és téli időszakában folytatott periódus kísérletek
avarcsökkenésének üteme (A mintavételi alkalmak kéthetente történtek. Az üres jelölők minden esetben a sima avarzsákot jelentik, míg a feketék a planktonháló anyagból készült zsákokból történő avarfogyást jelenítik meg. A római számok az egyes időben egymáshoz képest elcsúsztatott mérési periódusokat jelentik, melyek átfedése a 2. ábrán láthatóak.) .... 41 14. ábra A két patakban (Csigere-patak, Vázsony-séd) az aprítók legnagyobb kolonizációs mérete az idő és a k együttható tekintetében ... 43 15. ábra Az aprítóegyedek átlagos mennyisége (1 g száraz avarra vonatkoztatva) és a k együttható tükrében ábrázolva ... 43 16. ábra A két mintavételi helyen talált különböző táplálkozási csoportokba sorolt
makrogerinctelenek aránya a nyári időszak során ... 45 17. ábra A két mintavételi helyen talált különböző táplálkozási csoportokba sorolt
makrogerinctelenek aránya a téli időszak során ... 45
5 18. ábra A k-érték és a hőmérséklet összehasonlítása az aprítók kolonizációs
egyedszámának maximális mennyiségével a Csigere-patakban a két vizsgálati periódus során (Az ábrákon a római számok a kihelyezett rácsra utalnak (lsd. 2. ábra), az arab számok pedig, hogy az adott rácson hányadik mintavételről van szó.) ... 46 19. ábra A k-érték és a hőmérséklet összehasonlítása az aprítók kolonizációs
egyedszámának maximális mennyiségével a Vázsonyi-sédben a két vizsgálati periódus során (Az ábrákon a római számok a kihelyezett rácsra utalnak (lsd. 2. ábra), az arab számok pedig, hogy az adott rácson hányadik mintavételről van szó.) ... 47 20. ábra A Vázsonyi-sédben elhelyezett különböző mintavételi eszközökből történő avar fogyásának üteme ... 48 21. ábra A Vázsonyi-sédben az aprítók legnagyobb kolonizációja, az idő és a k együttható összefüggése (Narancssárga színnel az avarhenger, zöld színnel az avardoboz, szürke
színnel pedig az avarzsák van jelölve) ... 49 22. ábra Az aprító egyedek átlagos mennyisége (ind g-1 száraz avar) a k együttható
tükrében ... 49 23. ábra A teljes egyedszám eloszlása (a), és a táplálkozási csoportok eloszlása (b) az összes terepi eszközben ... 50 24. ábra Az eszközökben található egyedek táplálkozási csoportok szerinti megoszlása ... 50 25. ábra Az eszközökben található egyedek fajszám szerinti megoszlása a; avardoboz, b;
avarhenger, ... 51 26. ábra A két patakban az avarfogyás ütemének tendenciája ... 51 27. ábra A két patakban (Csigere-patak, Vázsonyi-séd) az aprítók legnagyobb
kolonizációja, az idő és a k együttható tükrében ... 52 28. ábra Az aprítók átlagos mennyisége (1 gramm száraz avarra vonatkoztatva) és a k együttható összefüggése ... 53 29. ábra A dolgozatban szereplő összes mintavételkor az egyes patakok helyszíni és
vízkémiai paraméterei alapján készített PCA ... 53 30. ábra Az átlagos k értékek kísérletenként és mederadottságonként ... 56 31. ábra Az első és a második kísérlet átlagos k értékei mederadottságonként (A boxplot ábrán a vastag vízszintes vonal a k-értékek mediánját, a doboz alsó és felső éle az alsó és felső kvartilist, a bajuszok végei pedig az adatok kiugró értékek nélküli tartományát jelölik.
A kiugró értékeket kis körök jelzik.) ... 57 32. ábra Az aprítók átlagos egyedszámának logtranszfolmárt értékei a medertípusok szerint (A boxplot ábrán a vastag vízszintes vonal a k-értékek mediánját, a doboz alsó és felső éle az alsó és felső kvartilist, a bajuszok végei pedig az adatok kiugró értékek nélküli
tartományát jelölik. A kiugró értékeket kis körök jelzik.) ... 58 33. ábra Az átlagos k értékek a harmadik és az ötödik kísérletben az avarzsákok lyukbősége szerint (A boxplot ábrán a vastag vízszintes vonal a k-értékek mediánját, a doboz alsó és felső éle az alsó és felső kvartilist, a bajuszok végei pedig az adatok kiugró értékek nélküli tartományát jelölik. A kiugró értékeket kis körök jelzik.) ... 62 34. ábra A dolgozatban szereplő összes kísérlet k értékei a kísérlet időtartalmának tükrében ... 63
6
TÁBLÁK JEGYZÉKE
1. táblázat Az egyes kísérletek helyszíneinek megjelölése ... 20 2. táblázat Az egyes kísérletekben használt terepi eszközök (Ø: a zsákok anyagának
szembősége)... 22 3. táblázat Az egyes kísérleti helyszíneken kihelyezett kísérleti eszközök mennyisége (db) avartípusonként (Pt: Populus tremula, Sa: Salix alba, Qr: Quercus robur) ... 23 4. táblázat A kísérletek hossza (napban), és a mintavételek gyakorisága (napban) (Az 5.
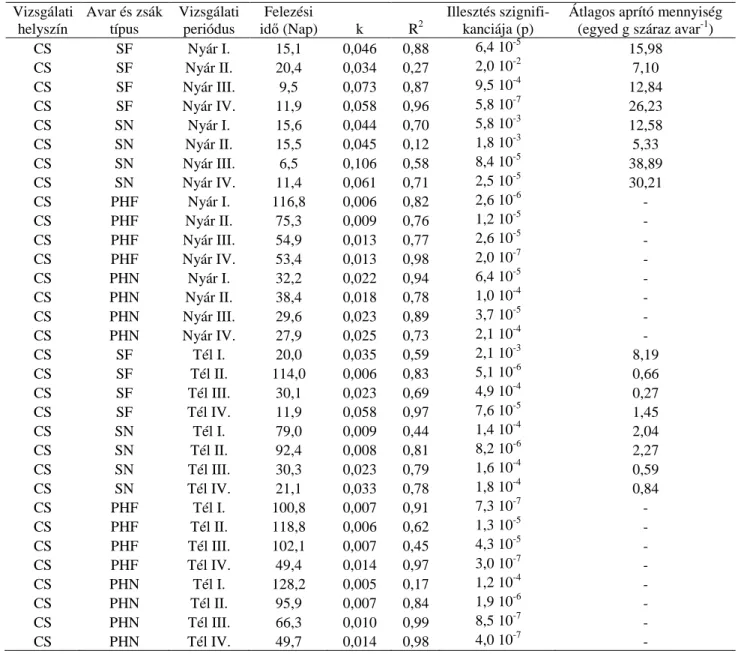
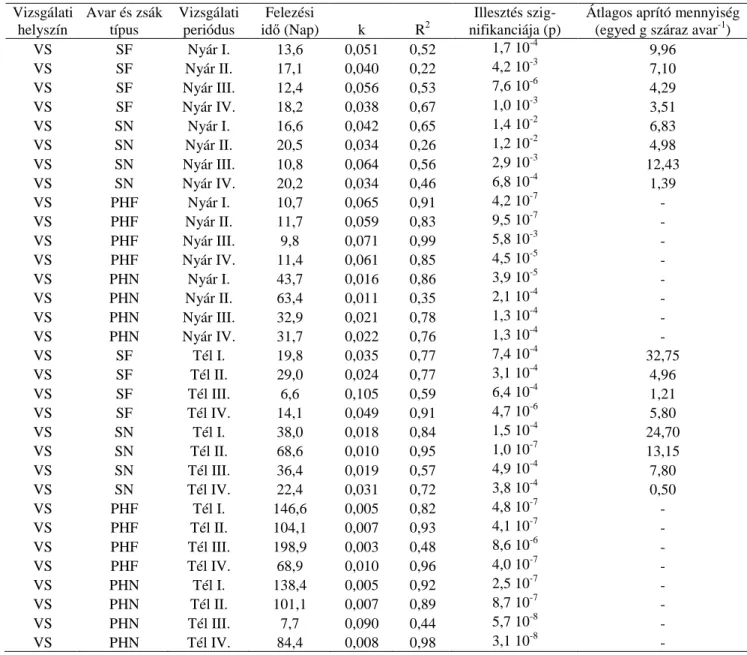
kísérletben a vizsgálni kívánt cél érdekében a mintavételi gyakoriság nem volt konstans.).. 23 5. táblázat A vizsgálati helyszínek főbb jellemzői ... 25 6. táblázat A nyolc mintavételi helyen az avarfogyás néhány mutatója (Felezési idő (nap):
avar felezési ideje, k: az exponenciális illesztésből számolt avarbomlási együttható, R2: az exponenciális görbe illeszkedésének mértéke, p: az illeszkedés szignifikanciája, Átlagos aprító mennyiség: az avarzsákban az egy gram avarra jutó aprító makrogerinctelenek átlagos egyedszáma) ... 30 7. táblázat A három patak (Csigere-patak, Torna-patak, Veszprémi-séd) avarfogyási
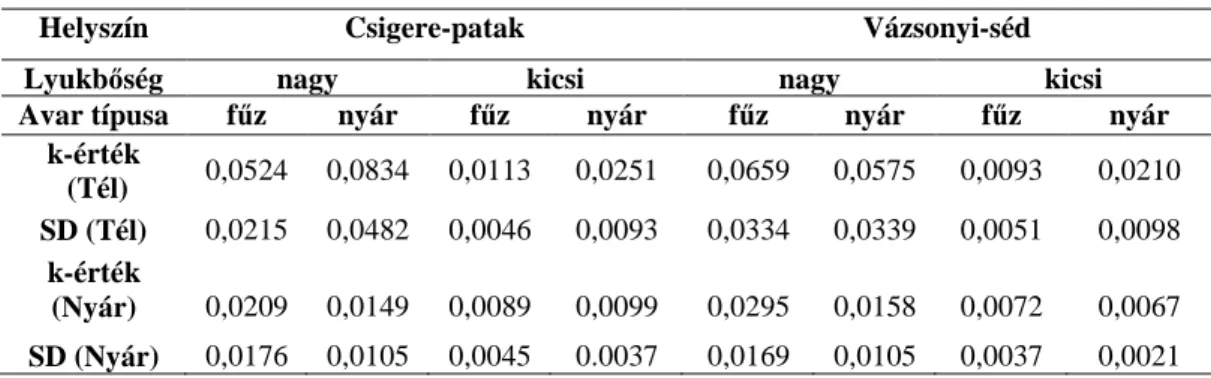
ütemének néhány mutatója (Felezési idő (nap): avar felezési ideje, k: az exponenciális illesztésből számolt avarbomlási együttható, R2: az exponenciális görbe illeszkedésének mértéke, p: az illeszkedés szignifikanciája, Átlagos aprító mennyiség: az avarzsákban az egy gram avarra jutó aprító makrogerinctelenek átlagos egyedszáma) ... 33 8. táblázat A különböző avarzsák típusok átlagos k-értékei és azok szórása a téli vizsgálat során (A nagy lyukbőség: 3mm-t, míg a kicsi lyukbőség 100µm-t jelent.) ... 36 9. táblázat A zsákok lyukbősége, a mintavételi helyek és a hőmérséklet interakciós
vizsgálata többváltozós lineáris modellel ... 38 10. táblázat A Csigere-patakban a nyári és a téli időszak alatt az avarfogyás néhány
mutatója (Rövidítések: CS: Csigere-patak, SF: sima zsák fűzavarral töltve, SN: sima zsák nyár avarral töltve, PHF: planktonhálózsák fűzavarral töltve, PHN: planktonhálózsák nyáravarral töltve, Római számok: periódusok jelölése, az „aprítók átlagos mennyisége”
című oszlopban a planktonháló zsák (PH) mintáinak sorában nem látható számszerű adat, mert azokba a makrogerinctelen szervezetek nem tudnak bejutni. ... 40 11. táblázat A Csigere-patakban a nyári és a téli időszak alatt az avarfogyás néhány
mutatója (Rövidítések: VS: Vázsonyi-séd, SF: sima zsák fűzavarral töltva, SN: sima zsák nyár avarral töltve, PHF: planktonhálózsák fűzavarral töltve, PHN: planktonhálózsák nyáravarral töltve, Római számok: periódusok jelölése, az „Átlagos aprítófaj Mennyiség”
című oszlopban a planktonháló zsák (PH) mintáinak sorában nem látható számszerű adat, mert azokba a makrogerinctelen szervezetek nem tudnak bejutni. ... 42 12. táblázat Az aprítók mennyisége, a mintavételi helyek és a hőmérséklet interakciós vizsgálata többváltozós lineáris modellel (log (apr. átl. rel): az aprítók átlagos relatív
mennyiségének természetes alapú logaritmusa) ... 43 13. táblázat A terepi eszközökben történő avarbomlás néhány mutatója (Felezési idő (nap):
avar felezési ideje, k: az exponenciális illesztésből számolt avarbomlási együttható, R2: az exponenciális görbe illeszkedésének mértéke, p: az illeszkedés szignifikanciája, Átlagos aprító mennyiség: az avarzsákban az egy gram avarra jutó aprító makrogerinctelenek átlagos egyedszáma) ... 48 14. táblázat Az avarfogyás ütemének néhány mutatója (Felezési idő (nap): avar felezési ideje, k: az exponenciális illesztésből számolt avarbomlási együttható, R2: az exponenciális görbe illeszkedésének mértéke, p: az illeszkedés szignifikanciája, Átlagos aprító mennyiség:
az avarzsákban az egy gram avarra jutó aprító makrogerinctelenek átlagos egyedszáma) .... 52
7
KIVONAT
Avarlebontási kísérletek dombvidéki kisvízfolyásokon
A doktori értekezésben bemutatott kutatások célkitűzése az volt, hogy információval szolgáljon a magyarországi dombvidéki kisvízfolyásokban zajló avarlebontási folyamatokról. A kutatások elsősorban a bomlási ráták megállapítása és a vízi makrogerinctelen szervezetek aprító tevékenységének meghatározása érdekében történtek.
Ezekre vonatkozóan tudományos eredmények a következők:
(1) A szerző nyolc patakszakaszon egyéves kísérleti periódusban vizsgálta az avarlebontás ütemét, és ebben a folyamatban a vízi aprító makrogerinctelenek szerepét. A vizsgálat helyszínéül kiválasztott patakszakaszok természetes és erősen módosított kategóriába sorolhatók. Az eredmények alapján megállapítható, hogy a természetes mederrel és part menti vegetációval rendelkező patakszakaszokon a makrogerinctelen fauna mennyiségi és minőségi viszonyai megfelelőek. Ezzel szemben az erősen módosított mederben ugyan gyors a lebontás, de ez nem az aprító szervezeteknek köszönhető, hanem a vízfolyás alacsony visszatartó képességének, aminek egyik oka a meder degradáltsága.
(2) A szerző kísérletei során három fafaj avarját használta (jellemzően egy helyszínen kétféle avart használt, azokat, melyek a part menti fás szárú vegetációban a legjellemzőbbek). A használt avarféleségek eltérő bomlási ütemének vizsgálatára a szerző külön kísérletet végzett három hasonló patakszakaszon. Az eredmények szerint leggyorsabb fogyású a Populus tremula avarja, ennél kicsit lassabban fogyott a Salix alba, és jóval lassabban a Quercus robur avar.
(3) Mivel Magyarországon még semmilyen összefüggésben nem vizsgálták az avarfogyást kisvízfolyásokban, így a szerző megvizsgálta az avarfogyás ütemének hőmérsékletfüggését is. Erre azért is szükség volt, mert a hőmérséklet nem csak a bomlási folyamatra hat, hanem az aprítók egyedszámára is, így közvetett módon is hatással lehet a lebontásra. A szerző eredményei a k-értéket (bomlási mutató) befolyásoló tényezők vizsgálata során azt mutatják, hogy a téli periódus alatt az aprítók száma és a hőmérséklet is szignifikánsan növeli a bomlás sebességét (k-érték). A nyári periódus alatt viszont csak az aprítók száma növelte szignifikánsan a lebontás sebességét (k-érték).
(4) A szerző a tapasztalatai alapján egy újfajta, avarfogyás vizsgálatára alkalmas terepi eszközt fejlesztett ki, melyhez hasonló a szakirodalomban nem ismert. A fejlesztés szükséges, és jelentős, mivel az irodalomban általában durva mederanyagú patakokban (vagy állóvízben) történt vizsgálatokat lehet találni, szemben a magyarországi, már dombvidékeken is jelentős hordalékot szállító patakokkal (mely sokszor betemeti az objektumokat). Az új eszköz (avarhenger) elsősorban a kisodródást tartja vissza, és célirányosan a makrogerinctelenek aprító tevékenységének felmérésére alkalmas.
(5) A szerző vizsgálta a vízben történő avarbomlás kezdeti szakaszát rövidtávú kísérletekkel, melyek során a kezdeti kioldódás mértékét mérte fel. A szerző eredményei szerint az avarok előkezelése (pl. tömegállandóságig való szárítás), mely minden kísérletben azonos volt, a jelentős kezdeti kimosódás (első 24-48 óra) nem torzította a hosszú időlefutású kísérletek eredményét.
8
ABSTRACT
Leaf litter decomposition experiments in low order streams
The aims of the research discussed in this PhD thesis were to provide information about the leaf litter processes in Hungarian low order streams. The research was focused on determination of leaf litter decomposition rates and examination of shredding activities of macroinvertebrates.
The new results are the following:
(1) In a 12 months research eight sampling sites were examined in terms of decay of leaf litter decomposition and role of macroinvertebrates in the related shredding processes. Both natural and strongly modified water bodies were among the sampling sites which enabled the author to compare the relation between leaf litter decomposition and streambed morphology. Results show that the quantity and quality of macroinvertebrate fauna is sufficient at sites with natural water body and riparian vegetation resulting sufficient leaf litter decomposition. On the other hand leaf litter decomposition is fast in heavily modified water bodies but this is a result of the bed morphology and high velocity and low leaf- retraining capability of the water body and not the result of a sufficient shredding population.
(2) The author used three species of leafs – two at each location – those which are the most typical in the riparian vegetation. To determine the dependency of the research on the leaf species a separate research was conducted at three sites with similar water bodies. The results show that highest decay is measured for Populus tremula, decomposition is somewhat slower for Salix alba and the slowest for Quercus robur.
(3) As leaf litter decomposition was not yet studied in Hungary the correlation of decay-rate and temperature was also studies. This was a necessity not only as temperature can have a direct effect on the decomposition but it also has an indirect effect via the quantity and species of shredder invertebrates. Results show that during winter both higher temperature and higher number of shredders are increasing the k-value of the decay while during summer the speed of decay correlates only with number of shredders but not with the temperature.
(4) During the research a new field tool was also developed to examine leaf litter decomposition which tool is not known yet in the literature: leaf litter cylinder. While existing literature examines leaf litter decay in stream with stable flow and stream and lake beds with rough sediments and rocks, Hungarian streams have volatile flow and carry large amount of fine sediments resulting high level of drift-away of leaf litter and tools being covered frequently by sediment. The new tool, the leaf litter cylinder, is primarily applicable to eliminate drift-away effect and examine pure impact of shredder activities of macroinvertebrates.
(5) The early phase of leaf litter decay was also examined with shorter experiments to determine the impact of initial dissolution. Dissolution process is important at the initial stage as the impact is depending on the pre-treatment of leaf litter even if same species are used. It was shown that the pretreatment (drying the leaf-litter to steady dry mass) and the resulting initial dissolution were not influencing the result of the long duration experiments.
9
ZUSAMMENFASSUNG
Laubabbruch Experimente in Hügelland Bächen Die neuen Ergebnisse für die Wissenschaft sind folgende:
Das Ziel der Dissertation war es, Informationen über die Laubabbruch Prozesse in ungarischen kleinen Hügelland Bächen mitteilen. Die Forschung konzentrierte auf Abbruchraten und auf die Schredder-Tätigkeit von aquatischen Makroinvertebraten.
Die wissenschaftlichen Ergebnisse sind die folgenden:
(1) Der Autor hat die Abbruchraten und die Rolle der aquatischen Makroinvertebraten auf acht Abschnitten durch ein Jahr lange experimentelle Periode untersucht. Die Test Standörter sind natürliche und stark modifizierte, so der Autor konnte die Phasendifferenz des Laubabbruchs in den verschiedenen Bachbettmorphologien messen. Die Ergebnisse deuten, dass die Abschnitte mit natürlichem Bachbett und Küstenvegetation angemessene Qualität und Menge von Flora haben. Im Gegensatz dazu hat das stark modifizierte Bachbett schnellen Abbruch, aber das ist nicht durch die Schredder-Organisationen, sondern duch die geringe Speicherfähigkeit der Bachbettmorphologin verursacht.
(2) Der Autor benutzte das Laub von drei Baumsorten und prüfte auch die verschiedene Abbruchraten der Baumsorten. Die Ergebnisse zeigen, dass Populus tremula den schnellsten Abbruch hat, ein wenig langsamer Salix alba, und viel langsamer Quercus robur.
(3) Da Laubabbruch in ungarichen Kleinwasserflüssen noch nicht gemessen würde, hat der Autor auch die Temperaturabhängigkeit des Laubabbruchs gemessen. Das was auch notwendig, denn die Temperatur nicht nur den Abbruch, sondern auch die Nummer der Schredder beeinflusst. Die Untersuchung der k-Wert (Decay-Index) zeigt, dass sie in der Winterzeit durch die Anzahl der Schredder und die Temperatur erheblich erhöht wurde.
Während der Sommermonate, hat jedoch nur die Anzahl der Schredder die k-Wert deutlich erhöht.
(4) Der Autor hat durch die Erfahrung eine neue Methode entwickelt, die in der Literatur nicht bekannt ist. Die Entwicklung ist notwendig und wichtig, denn die Literatur enthält im allgemeinen Forschungen in Seen, aber nicht in ungarischen Hügelland Bächen, die auch erheblichen Sediment haben (der oft die Objekte verschütten). Das neue Gerät (Laubzylinder) reduziert das Treiben und ist gezielt auf die Forschung von Makro- Invertebraten.
(5) Der Autor untersuchte auch die Anfangsphase des Laubabbruchs im Wasser mit kurzfristigen Experimenten, und beurteilte die anfängliche Auflösungsgeschwindigkeit. Die Auflösung ist wichtig in der ersten Zeit, da sie ist abhängig von der Behandlung des Laubs.
Der Autor zeigte, dass während der Vorbehandlung des Laubs (zB Trocknung bis zur Gewichtskonstanz), die das gleiche in jedem Experiment war, wurden die Ergebnisse durch ein signifikantes anfängliches Treiben (ersten 24-48 Stunden) nicht verzerrt.
10
1 Bevezetés
Az élővizekben a természetes eredetű szerves anyagok két fő forrásból származnak: (1) autochton forrásból, azaz a vízben történő fotoszintetizáló termelési folyamatokból és az (2) allochton forrásból, melynek eredete a környező erdő (Dobson & Frid 1998).
Az évenként bekövetkező lombhullással a partmenti növényzet biztosítja az alsóbbrendű patakok allochton szerves anyag igényét, amely elsődleges energiaforrás a kisvízfolyásokban. A behulló avar mennyisége az egyes erdőkben és patakok mentén jelentősen eltérhetnek. Az avarmennyiség több tényezőtől függ, pl. éghajlat, növényzet, talaj típus, fák kora (Hernandez et al. 1992), és a patak morfológiai jellemzőitől.
A behulló szerves anyag jelenthet leveleket és levél töredékeket, virágos részeket, kéregdarabokat, fadarabokat (ágakat és gallyakat), tobozokat, csonthéjas terméseket, gyümölcsöket és egyéb növényi részeket (Benfield 1997).
A behulló szervesanyag összetétele a lombhullással, a növényzet típusától és elhelyezkedésétől függően változik. Általában a levelek alkotják a legnagyobb hányadot, amelynek 41-98%-a lombhullással kerül a patakba (Abelho & Graça 1998; Benson &
Pearson 1993; Abelho & Graça 1996; Grigg & Mulligan 1999), ezért az avarlebomlás fontos része az ökoszisztéma összefüggéseinek, valamint vizsgálatával közelebb kerülhetünk a biodiverzitással kapcsolatos ismereteink kiegészítéséhez.
A biológiai sokféleség csökkenésének oka globális léptékben az erőteljes humán hatások melyek az ököszisztémákat érik (Sala et al. 2000; Dudgeon et al. 2006). Ennek a csökkenő tendenciának az egyik áttételes következménye az egyedsűrűségek csökkenése, mely jelenség önmagában kihat az ökoszisztéma folyamatokra, mint pl. a növénytermesztésre, tápanyag-átalakulásokra, és a lebomlásra is (Loreau et al. 2001; Srivastava & Vellend 2005).
Az elsődleges termelési folyamatok gyakran pozitívan befolyásolják a növényzet sokféleségét gyepes és egyéb környezetben (Hooper et al. 2005; van Ruijven & Berendse 2005; Balvanera et al. 2006), és néhány kísérlet azt sugallja, hogy bizonyos helyzetekben a biológiai sokféleség hatása az ökológiai folyamatokra riválizálhat a környezeti (rendelkezésre álló tápanyag) és a biotikus faktorokkal (predációs nyomás) is (Wojdak 2005).
Az erdei és ez egyéb vízi ökoszisztémákban az avarbomlás az egyik központi folyamat (Gartner & Cardon 2004) és ennek a folyamatnak a lefolyása függ a biológiai sokféleségtől is (Gessner et al. 1999; Moore 2004).
Ez a rendszer magában foglalja erdei patakokat, ahol fák tartanak árnyékot, így erősen korlátozva az elsődleges termelést, viszont bőségesen kínálnak egyéb forrásokat a táplálékhálózatoknak az avarral (Wallace et al. 1997).
Mivel a különböző szervezetek felhasználják az avart (Hieber & Gessner 2002; Moore 2004), a bomlási folyamat hatással lehet a biológiai sokféleségre több szinten is, beleértve a fajok gazdagságát mikrobiális közösségekben (Dang et al. 2005; Tiunov & Scheu 2005;
Duarte et al. 2006), detritivóra gerinctelen faunában (Jonsson & Malmqvist 2000; Ruesink
& Srivastava 2001), és növényi avarban is (Scherer-Lorenzen et al. 2007).
Több kísérlet is rámutatott arra, hogy jelentős az összefüggés az avar sokszínűsége (fafajok gazdagsága az adott területen) és a bomlási gyorsaság között (Swan and Palmer 2004;
Lecerf et al. 2007).
Összességében ezek az eredmények arra utalnak, hogy (1) az avar sokszínűsége fontos tényező lehet a bomlási folyamatokban, és ez fontos jele lehet (2) a diverzitás hatásának a bonyolult mögöttes mechanizmusok megértésében (Schindler & Gessner 2009).
A vízfolyásokban az avarterhelés feldolgozása négy szakaszból áll: (1) kioldás, (2) mikrobiális kolonizáció, a (3) gerinctelen szervezetek aprítása és a (4) fizikai kopás (Abelho 2001; Gessner et al. 2003).
11 Kioldódás tulajdonképpen az oldható anyagok abiotikus úton való távozása elsősorban a levéllemezből (pl. fenol, szénhidrátok és aminosavak). Ez a folyamat a vízbe merülést követően javarészt az első 24-48 óra alatt megtörténik, és esetenként akár 30%-os tömegveszteséget is jelenthet. Gessner és Schwoerbel (1989) kimutatta, hogy nem figyelhető meg ilyen gyors kimosódás veszteség, ha friss, és nem előre szárított avart használnak.
Mivel a lombhullató erdők avarja a táplálékhálózatok nagy jelentőségű energiaforrása (Fisher & Likens 1973), foglalkozni kell az elsődleges lebontó szervezetekkel is, a mikrobiális gombákkal. Gombákat, különösen a vízi Hyphomycetes fajokat tekintik elsődlegesen kolonizáló lebontóknak a patakokban (Suberkropp 1992). A mikrobiális bomlás mértékét a patakokban az abiotikus tényezők, mint hőmérséklet, áramlás, a fizikai kopás (Petersen & Cummins 1974), pH (Chamier 1987), nitrát és foszfát koncentráció (Pozo 1993), valamint a biotikus tényezők, mint az avar minősége (Melillo et al. 1984) határozza meg. A mikrobiális bomlás után vagy azzal egyidőben megindul a makrogerinctelenek kolonizációja is (Anderson & Sedell 1979). Az avar bomlása, az egymást követő funkcionális csoportok kolonizációja tükrözi a biológiai közösségek és a közvetlen környezet kölcsönhatását (Frankland 1998). A mikroorganizmusok megtelepedése a leveleken (Graça 1993, 2001) felgyorsítja az avar mineralizációját, javítja a táplálék minőségét és feldolgozhatóságát az aprítók számára (Gessner et al. 1999). Az aprító szervezetek az avart kisebb részekre darabolják, ami hat a gombák fogyasztására is, és kompetíciót is eredményez, hiszen nem mindegy, hogy először a gombák vagy a makrogerinctelenek kolonizálnak. A korai gombakolonizáció jelentős mennyiségű értékes energiatartalmat von ki a levelekből az aprító makrogerinctelen szervezetek elől (Bärlocer 1992; Suberkropp 1998). A lebontó gombák mennyiségét az avaron található ergoszterol mennyiségével szokás meghatározni. Az ergoszterol egy elsődleges szterol, mely a sejtmembránok felépítő anyaga a fonalas gombákban (Axelsson et al. 1995), de hiányzik az edényes növények és állatok szervezetéből (Newell 1992), ezért az ergoszterol tartalom meghatározását széles körben használják a gomba biomassza becslésére, mert erős korrelációt mutat a gomba biomasszával (Gessner & Chauvet 1997).
Az aprítók a gombák tevékenységével párhuzamosan, vagy valamelyest később az aprító makrogerinctelen szervezetek apró darabokra szaggatják elsősorban a levéllemezek érhálózata közötti részeket. Mivel a fák leveleinek kémiai összetétele más és más, ezért némelyik avar típus könnyebben aprítható és emészthető, mint mások. A kémiai összetétel meghatározza a fizikai kopásnak való ellenállóképességet is, melyet igen nehéz természetes körülmények között az aprító szervezetek tevékenységétől különválasztani.
Az, hogy egy aprító makrozoobentosz fajt mely funkcionális táplálkozási csoportba soroljuk (aprító-, aktív és passzív szűrő, törmelékevő-, ragadozó-, gyűjtögető-, élősködő-, legelő szervezetek), attól függ, hogy mely táplálkozási formát részesíti előnyben. Így, ha valamely fajt pl. aprítónak tekintünk, az nem jelenti azt, hogy más módon nem tud táplálkozni, csak az egyéb táplálkozási módok jóval kevésbé hatékonyak számára. Ez megmutatkozik a faj mennyiségi viszonyaiban, így az ökoszisztéma „egészségének” megítélése szempontjából lényeges. Tehát az aprítók elsősorban aprítanak, a felhalmozódott CPOM kupacokban jó búvóhelyre, védett élőhelyre és bőséges táplálékra találhatnak. Ahhoz, hogy a CPOM néhol felhalmozódjon, szükséges hogy a víztér változatos legyen, hogy az eltérő áramlási viszonyokkal rendelkező kisebb-nagyobb kanyarokban, kövek mögött az áramlás lerakhassa a leveleket. Az ilyen helyekre az aprítók után betelepülnek a ragadozók is a táplálékfajok tömegessége miatt, a könnyebb préda reményében.
Ahol a legfőbb energiaforrás az avar, ott elsősorban az aprító szervezeteknek kell dominálnia a vízi makrogerinctelen faunában. Tehát mennyiségük jelentős, hiszen éppen a
12 bőséges avarbehullást eredményező part menti vegetáció miatt a víztér árnyékolt, a dús diatóma vegetáció kialakulása fénylimitált, tehát a legelő szervezetek nem válhatnak dominánsá a faunában. Mivel a hegyvidéki patakok általában ultra-oligotrófok, így a jelentősebb vegetáció kialakulását a tápanyaglimitáció is gátolja. Ezáltal az aprítóknak csak a külső avar szolgál táplálékként. Mivel ezeken a szakaszon aprítódik fel a sok szerves anyag, amelyek apró darabkák formájában sodródnak tova, itt még sem az aktív, sem a passzív szűrögető szervezetek mennyisége nem lehet jelentős mennyiségű. A ragadozó fajok éppen ezért elsősorban az aprító fajokat fogyasztják.
A bomlási folyamat hatással lehet a biodiverzitás változására a különböző szinteken: a fajok gazdagságára a mikrobiális közösségekben (Duarte et al. 2006), detrivora gerinctelenek összetételére (Schädler & Brandl 2005) és a növényi maradványok bomlási dinamikájára, annak oxigén fogyasztási viszonyaitól függően (Kominoski et al. 2007).
Az egy élőhelyen található vízi makrogerinctelen szervezeteket korábban nagyobb taxonómiai csoportokba sorolva vonták össze, amit felváltott az ún. táplálkozási csoportokba (pl. aprító, aktív és passzív szűrő, törmelékevő, ragadozó, gyűjtögető, élősködő és legelő szervezetek) sorolás. A makrogerinctelen funkcionális csoportok olyan változók, melynek vizsgálata - a hagyományos biomonitoring mellett - egyre inkább elfogadott (Bunn 1995) az ökológiai állapot értékelése esetében. A funkcionális mutatók használata fontos előrelépés, hiszen közvetlenül mérhető paramétereiből az ökoszisztémák működésére, és annak sérülékenységére is lehet következtetni (Gessner & Chauvet 2002; Brooks et al. 2002).
Nagyon fontos az ökoszisztémák működése szempontjából az élőhely heterogenitása, mivel befolyásolja az ökológiai folyamatokat valamennyi szerveződési szinten, beleértve a viselkedést, populációdinamikát, a fajok közötti interakciókat-, és a közösség szerkezetét (Free 2009; Gascón et al. 2008; Brown 2007). Az Európai Unió Víz Keretirányelve (WFD 2000) is hangsúlyozza, hogy a patakmedrek módosításának egyik következménye a degradálódó, elszegényedő növény- és állatvilág, ami egyszerűsíti az anyag- és energia- áramlási hálót. Így a folyamatok sérülékenyebbé válhatnak, ami által az adott víztest1 nem éri el a jó állapotot vagy potenciált. Az utóbbi években fontossá vált a medermódosítások ökológiai hatásainak vizsgálata. Például a természetes vízjárás helyreállítása is fontos lépés lehet a zavartalan állapotú ökoszisztéma működésében (Muehlbauer et al. 2009). A helyreállítás fontos eleme lehet a part menti vegetáció újbóli létrehozása is (Harding et al.
2006).
A nagyjából érintetlen parti régióval rendelkező természetes jellegű vízfolyásokba szignifikánsan magasabb avarmennyiség (avar -input) jut, továbbá az avarbontási képességük az akkumulációs zónában akár kétszer nagyobb is lehet, mint a módosított patakoké (Watson & Barmuta 2010). Mivel a vízi aprító makrogerinctelen szervezeteknek kulcsszerepük van az avarbomlás folyamatában (Bärlocher 1985), a módosított szakaszokon az avar -input és annak megtartása (tovasodródás) is befolyásolja a táplálékbőséget, tehát az aprító tevékenység aktivitását.
A medermódosításokkal az élővilág strukturális és funkcionális jellemzői is összefüggésben állnak. Például Stenger-Kovács és munkatársai (2006) valamint Kovács és munkatársai (2008) arról számolnak be, hogy a mederbeli duzzasztások hogyan befolyásolják a diatóma közösségek faji összetételét. Mindkét cikk rámutat a vízfolyásokon létrehozott tározók fel-, és alvízi hatására a diatóma közösségekre. Moulton és munkatársainak (2010) tanulmánya
1 A víztest a VKI szerint a vízgazdálkodás alapegysége. Olyan különálló és jelentős elem, mely tulajdonságaiban bizonyos mértékig homogén. Ez a VKI ideája. Valójában ilyen víztest nincs, vagy alig van.
13 szerint az avar bomlását a városi (tehát módosított) patakokban más fogyasztók, halak és ebihalak végzik az aprító makrogerinctelenek helyett. Lorenz és munkatársai (2009) módosított víztestek makrozoobentosz közösségeit vizsgálták helyreállítás után, és felhívták a figyelmet arra, hogy a mederrehabilitációs intézkedéseket körültekintően kell megtervezni és kivitelezni, hogy azok valóban elősegítsék a diverz makrogerinctelen fauna kifejlődését.
Friberg és munkatársai (2009) megállapították, hogy viszonylag gyengék a megfogható összefüggések a jelenleg használt medermorfológiai intézkedések és makrogerinctelen fauna értékelési szempontjai között, és sürgetik a szisztematikus, megalapozott biológiai értékelés szempontjainak kidolgozását a mederágyi változtatások hatásának felmérésére.
A hegyi patakokban végbemenő avarbontás a nemzetközi irodalomban jól kutatott területnek számít (Hawkins et al. 1982; Golladay et al. 1989; Stout et al. 1993; Stone & Wallace 1998), de az alacsonyabb területeken folyó síkvidéki patakok kevésbé kutatottak (Benke et al.
1984; Kedzierski & Smock 2001).
Az avarbontást jelentősen befolyásolja a patakot övező területek hasznosítási formája is (Mulholland & Lenat 1992), mely hatással van a patakok makrogerinctelen élőhelyeinek összetételére és a szerves anyag feldolgozásra (Hax & Golladay 1998; Kedzierski & Smock 2001). Mindkettőt jelentősen módosíthatja a patakok medermorfológiája. Az allochton eredetű durva szemcsés szerves anyagok (CPOM) primer energiaforrást képeznek erdős területen futó patakokban (Webster & Benfield 1986; Minshall 1996; Giller 1998), ezért az aprítók elsődleges kapcsolatalkotók a patak tápanyagellátottsága és a környező táj között. Az aprítók alakítják át CPOM-ot finom szemcséjű szerves anyaggá (FPOM), amely aztán a táplálékhálózat többi tagjának biztosít táplálkékot (pl. a gyűjtögetőknek és a szűrögetőknek;
Cummins et al. 1989; Cuffney et al. 1990). Ennek eredményeként a patak vízgyűjtő területén lévő növényzet változása módosítja a minőségi és mennyiségi allochton CPOM bemenetek ütemét, ami erőteljes hatással lehet a közösség szerkezetére és ökológiai funkciójára (Golladay et al. 1987; Smock et al. 1989; Bilby & Ward 1991). Figyelembe véve a patakok és az őket környező szárazföldi táj közötti szoros kapcsolatot, arra következtethetünk, hogy ezek a hatások befolyásolhatják a víz minőségét még a nagyobb patakok és folyók esetében is, mivel a kisvízfolyások erősen meghatározzák, hogy a magasabb rendű folyókban mennyi táplálék áll rendelkezésre a különböző táplálkozású makrogerinctelenek számára.
Az erdészeti munkálatok jellemzően a part menti és a vízgyűjtőn elterülő erdőket is érintik, de jelentős lehet az útépítés és a mezőgazdasági termelés hatása is. Ezek a tevékenységek egyaránt érintik az abiotikus és a biotikus folyamatokat is a patakokban. Például parti növényzet híján nő a víz hőmérséklete (Brown & Krygier 1970; Webster & Waide 1982), csökken a szerves anyagok beáramlása és a szerves anyag visszatartás (Newbold et al. 1980;
Golladay et al. 1989; Smock et al. 1989), valamint nő a patak szervetlen tápanyag koncentrációja (McClurkin et al. 1985; Swank et al. 1988).
Utak és patak kereszteződésénél az üledék mennyisége növekedhet, ami szintén nem természetes terhelési forrás (McClurkin et al. 1985; Swank 1988). A fokozott fény és hőmérsékleti viszonyok, valamint a többlet tápanyag-koncentráció növelheti az alga biomasszát, ami megváltoztatja az őshonos makrogerinctelen fajegyüttesek összetételét (Likens et al. 1970; Wallace & Gurtz 1986; Bilby & Bisson 1992). Mindezen folyamatok következményeképp csökken az élőhelydiverzitás és a makrogerinctelenek sokszínűsége (Hynes 1975; Richardson 1985). Pusztán a part menti sávok megfelelő gondozásával nemcsak ezeket a problémákat lehet kiküszöbölni, hanem csökken az erózió és a tápanyag- lefolyás is (Newbold et al. 1980; Gowns & Davis 1991; Ormerod et al. 1993).
A patak szerkezetét és energiaforrását érő módosulások miatt változások következhetnek be a makrogerinctelen közösség szerkezetében és funkcióiban (Golladay et al. 1987; Smock et
14 al. 1989; Bilby & Ward 1991). A fő allochton energiaforrást biztosító fák hiánya miatt kisebb lesz a szerves táplálékforrás (Webster & Benfield 1986; Maloney & Lamberti 1995;
Giller 1998, Goodman et al. 2006).
1.1 Hazai avarlebontási kísérletek áttekintése
Az előző, bevezetés fejezetben áttekintettem egyúttal a nemzetközi irodalomnak azon részét, mely jobban érinti jelen értekezés témáját. Az alábbiakban a hazai szerzők munkáját kívánom röviden összegezni, az avarlebontás területét szélesebben értelmezve, mint ahogyan az értekezés témája megkívánja.
A funkcionális csoportok használata nemzetközi szinten széles körben elfogadott (Moog 2002), ugyanakkor Magyarországon csak csekély számú irodalommal rendelkezik az a terület, mely a növényi szerves anyag lebontása folyamán a makrogerinctelenek tevékenységét (tehát a funkcionális csoportok közül az aprító makrogerinctelenekét) is vizsgálja. Az alábbiakban rövid áttekintés következik arról, hogy országunkban eddig milyen tanulmányok születtek vízi dekompozíciós folyamatok terén.
A vizsgálatok elsősorban a Fertőben történtek, de volt már hazai laboratóriumi vizsgálat, és egy-egy a Gemenc és a Kis-Balaton területén. A Kis-Balatonban történt az avas nád levele és szára bomlásának megfigyelése in situ körülmények között, melynek előzményeként a laboratóriumi vizsgálattal a szerzők igazolták, hogy huminanyagok is képződnek (V.-Balogh et. al. 2001). Ezzel bebizonyították, hogy a Balaton Kesztelyi-medencéje szerves anyag (huminanyag) terhelésének egy része a nád bomlásból eredeztethető, mely a Zala folyó közvetítésével éri el a Balatont. Varga (2001, 2003) a Fertőben a nád gyöktörzsének bomlását figyelte meg 1997 és 2000 között, mely időtartam alatt 41 makrogerinctelen taxont mutatott ki. A domináns taxonok Diptera, Isopoda és Gastropoda csoportba tartoztak, melyek közül csak az Isopoda tekinthető kizárólagosan aprítónak. A kísérlet szerint a növény anyagának (nád-rizóma) szerkezete és kémiai összetétele, valamint a környezeti tényezők határozzák meg a makrogerinctelen közösség tevékenységét a Fertőben (Varga, 2003).
A Fertőben előzőleg más vizsgálatok is folytak, elsősorban szintén náddal, de e vizsgálatok során nem figyelték meg a makrogerinctelenek tevékenységét. Ezek a vizsgálatok elsősorban a tó nádas parti zónájában folytak, nem a nyílt víz felőli részen (Dinka 2001). A szerző szerint a nádas nagy változékonyságot mutat térben és időben is (Dinka 1993), így a vizsgálatban a zsákok helyének kiválasztása nagyon körültekintően folyt. Ezekben a tanulmányokban megállapították, hogy a kimosódás az avarzsákokból 50% körül van az első három hónapban (Dinka & Szeglet 1999; Dinka 2002) és a mikrobiális közösség oxigénfogyasztása nagyjából 50%-kal emelkedett az első négy hónap során (Dinka 2001).
Ezeket az eredményeket összevetették a szerzők saját előző kísérleteikkel, melyek szintén a helyszínen folytak, és hasonló eredményekre jutottak (Dinka 1999; Varga 2001). Mindezek mellett még azt is megállapították, hogy a nád levele a rizómához képest több könnyebben bomló hemicellulózt tartalmaz (Dinka 1999).
Szabó (2000; 2003) vizsgálta laboratóriumi körülmények között a tetrazólium csökkentését enzimkinetikai vizsgálattal a lebomló nádban, hogy így gyűjtsön információt a mikrobiális lebontók tevékenységéről. Ezek a kísérletek igazolták azt, hogy növekszik a nyári időszak alatt a holt rizómában a mikrobiális bomlás (Dinka & Szabó 2002).
A nádban tápanyagszint-változás vizsgálatok is folytak hazánkban szintén a Fertőben. E vizsgálat folyamán szintén avarzsákos módszert használva a tömegveszteséget, hemicellulóz, cellulóz, lignin, C, N, S és P koncentrációjának változásait követték nyomon (Dinka et al. 2004; Ágoston-Szabó et al. 2006). E vizsgálatban is alátámasztották a szerzők
15 egy korábbi saját eredményüket, miszerint a nád levelének alacsonyabb rosttartalma (ezáltal eltérő kémiai összetétele) miatt háromszor gyorsabban bomlik, mint a rizóma (Dinka 1986).
A fent említett szerzők nemcsak a nád bomlását vizsgálták, hanem a keskenylevelű gyékényét (Thypa angustifolia) is, a Fehér-tóban (sekély tó a Fertő-Hansági Nemzeti Park területén). Kétféle lyukbőségű (1 mm és 0,05 mm) zsákot töltöttek meg gyékénnyel, de ezek között nem találtak különbséget az avarbomlás gyorsaságában (Ágoston-Szabó & Dinka, 2005). Ugyanezen szerzők összehasonlították a nád és a gyékény bomlásának gyorsaságát is, és megállapították, hogy a nád rizómája bomlott lassabban (Ágoston-Szabó & Dinka 2008).
Egy, a Gemenc területén folytatott vizsgálat (Ágoston-Szabó et. al. 2011) helyszínét tekintve újszerű, elsősorban állóvízi kísérletnek tekinthető, mivel holtágon végezték. A bomló fűz avar C, N, P és S tartalmát mérték, valamint becsülték a gomba biomasszát ergoszterol koncentráció alapján.
A kis patakok allochton szerves anyag terhelésének becsléséről egy felmérés született (Vass et al 2010) ezidáig, dombvidéken futó két patakon.
A Torna-patakban 2010 október 4-én bekövetkezett vörösiszap katasztrófa után a Pannon Egyetem munkatársai vizsgálták a mikrobiális bomlást az újraéledő patakban (Üveges et al.
2011), melynek során 26 Hyphomycetes fajt tudtak kimutatni már 5 hónappal a katasztrófa után (Vass et al. 2011). A gombák elsősorban a Salix alba, Quercus cerris, Fagus sylvatica és Populus nigra fák avarját kolonizálták a legaktívabban. Ezen kívül vizsgálták a kocsányos tölgy (Quercus robur), a rezgő nyár (Populus tremula) és a fehér fűz (Salix alba) lebomlási ütemét is (Kucserka et. al 2011). E vizsgálat eredményei szerint a Torna-patak károsodott szakaszán (ahol a vizsgálat ideje alatt nem voltak makrogerinctelen szervezetek) az avarféleségek bomlása nem volt exponenciális. Az első 56 napban alacsony ütemű avartömeg veszteség volt kimutatható. Ergoszterol mennyisége a katasztrófa után felére csökkent.
A rövid áttekintésből kiderül, hogy a hazai szerzők publikációi nem olyan szempontból vizsgálják a vízterekben a szerves anyag bomlást, mint az jelen dolgozatban történik, mivel a hazai vizsgálatok ezidáig elsősorban állóvizekben történtek, míg jelen dolgozatban a kísérletek csak kisvízfolyásokon folytak. Az aprító makrogerinctelenek tevékenységére is csak egy vizsgálat folyt a Fertőben (Varga 2003), így azt lehet mondani, hogy a jelen dolgozat témájának vizsgálata hiánypótló.
16
2 Célkit ű zés
2.1 Avarbomlás és medermorfológia
A kutatás egyik célja, hogy felmérjem a természetes jellegű patakmedrekben és a módosított patakmedrekben folyó avarbomlás ütemét és jellegét. Vizsgálatomban az elsősorban az aprító makrogerinctelen szervezetek tevékenységére és az avarlebontás ütemének összefüggéseire összpontosítottam az egyéves kísérleti időszak alatt. Célom volt az aprító makrogerinctelen denzitás és az avar bomlási rátái közötti összefüggések megkeresése, melyek segítségével következtetni tudtam az ismert medermódosítás hatásaira a makrozoobentosz összetételére. Ezen kívül meg szerettem volna állapítani az összefüggést az aprító makrogerinctelen szervezetek denzitása és az avarbomlási ráta között.
2.2 Három, patakparti vegetációt alkotó fafaj avarjának bomlása
A lombhullató és elegyes erdőkben hazánkban 3-5 fafaj a legdominánsabb a hegyvidékeken, míg a dombvidéki területekre érve a zonalitás miatt a fafajok száma növekszik. Mivel minden fafaj levelének kémiai összetétele más, e strukturális és kémiai különbségek meghatározzák a fizikai aprózódásnak való ellenállást és az aprítók számára az emészthetőséget is. A fizikai aprózódásnak való kitettség a sodrási viszonyokkal változik, míg a sodrási viszonyok a medereséssel állnak összefüggésben. Célom az volt, hogy megállapítsam, a három fafaj avarja közül melyik áll ellen legjobban a fizikai aprózódásnak és a makrogerinctelen szervezetek aprító hatásának, valamint hogy megtudjam, milyen mértékben különbözik a három fafaj avarjának bomlási üteme a természetes és a mesterséges patakszakaszokon.
2.3 Avarbomlás hőmérsékletfüggése
A harmadik nagy kísérleti periódusnak az volt a célja, hogy az avarlebontás sebességét avarzsákos módszerrel tanulmányozzam a közel természetes állapotú Vázsonyi-séd és az enyhén módosított jellegű Csigere-patak egy-egy kiválasztott pontján, egy nyári és egy téli vizsgálati időszak alatt. A kutatás során kíváncsi voltam az avarlebontási ráta, az aprító szervezetek mennyisége és a hőmérséklet összefüggéseire, valamint az aprítók avartípus preferenciájára. Mindezek mellett ebben a kísérletben alkalmaztam először két lyukbőségű avarzsákot, melyekkel az volt a célom, hogy a kisebb lyukbőségű zsákokból kizárjam a makrogerincteleneket, ezáltal a mikrobiális bomlás mértékét kívántam becsülni a makrogerinctelen szervezetek tevékenysége mellett és nélküle.
2.4 Saját fejlesztésű terepi eszköz hatékonyságának összevetése a hagyományos módszerekkel
Az irodalomban a legelterjedtebb terepi avarlebontást vizsgáló eszköz az avarzsák, de emellett használatos még az avardoboz is (Graça et al. 2005). A zsákok nagy hátránya, hogy a felaprított és a mechanikai aprózódás által darabolódott avar könnyen elsodródik, míg a dobozokban ez benne marad. Viszont a doboz nagy hátránya az, hogy a sodródó aljzatalkotó szemcsék a dobozban könnyen felhalmozódnak. Ezeken a hatásokon elgondolkozva fejlesztettem ki az avarhengert, melynek ötlete igen egyszerű. A henger hálóval lefedett két végén a szervezetek ugyanolyan könnyen bejutnak, mint a hagyományos eszközökbe.
Miután azonban a sodrásra merőlegesen áll a henger palástja, így abban nem tud lerakódni a hordalékanyag, és az aprózódott avar sem tud kisodródni. E terepi módszerek összehasonlítása volt a célom ezzel a kísérleti periódussal. Szándékomban állt
17 összehasonlítani az avarlebontási ráta alakulását és a makrogerinctelen szervezetek eszközpreferenciáját egy referencia eszközben (planktonhálóból készült zsák, melybe az aprítók nem tudnak bejutni), egy klasszikus avarzsákban (melynek lyukbősége alapján bejuthatnak az aprító szervezetek, de ki is sodródhat az avar), az avardobozban (mely könnyen telítődik aljzatszemcsékkel), és az avarhengerben (melynek használata az irodalomból eddig nem ismert). Kíváncsi voltam továbbá arra is, hogy függ-e az avarlebontási ráta az eszköz típusától.
2.5 Avarbomlás kezdeti szakaszának vizsgálata
Folytattam két rövidtávú avarbomlási kísérletet olyan helyszíneken, ahol már előzőleg folytak hosszútávú kísérletek is. Ezzel a kísérlettel az volt a célom, hogy felmérjem a kezdeti aprózódás ütemét, valamint pontosabban szerettem volna megbecsülni azt, hogy mikor történik a makrozoobentosz szervezetek kezdeti betelepülése.
18
3 Hipotézisek
3.1 Avarbomlás és medermorfológia
Munkahipotézisem az volt, hogy a módosított patakszakaszokon szegény és/vagy alacsony a makrogerinctelen denzitás, mert egyrészt az avar-input nem elégséges és azt a kevés avart is azonnal elmossa a kibetonozott trapézmederben futó víz, másrészt a homogén élettér miatt a makrozoobentosz fauna amúgy is szegényes. Ezen fő okok miatt, az ilyen egyszerű folyamatok - avar bomlása a víztérben - nem teljesek.
3.2 Három, patakparti vegetációt alkotó fafaj avarjának bomlása
Egy eltérő áramlású, természetes lefutású mederszakaszon a sodrás kisebb, mint egy egyenes, homogén mederben. A három fafaj egyike, a kocsányos tölgy (Quercus robur) jellemzően előfordul hegy- és dombvidékeken is. A rezgő nyár (Populus tremula) és a fehér fűz (Salix alba) pedig a dombvidéki patakszakaszok part menti fái, egyrészt a természetes hullámtereken, de alkalmazzák őket szántóföldeken part menti deflációs erdősáv kialakítására is, tehát mindhárom jellegzetesen előfordul dombvidéki patakjaink partján.
Hipotézisem szerint leglassabb fogyási ütemmel a kocsányos tölgy avarja, leggyorsabban pedig a fehér fűz avarja fogy majd. Előzetesen azt az eredményt vártam, hogy az aprító makrozoobentosz szervezetek a kocsányos tölgy avarját fogják a legkisebb mértékben kolonizálni.
3.3 Avarbomlás hőmérsékletfüggése
Az avarbomlás ütemét természetesen jelentősen meghatározza a víz hőmérséklete is (a sok egyéb tényező mellett), hiszen az hatással van a mikrobiális bomlás gyorsaságára és a makrogerinctelenek egyedszámára is. Hipotézisem az volt, hogy a nyári időszakban egyértelműen és jól kimutathatóan gyorsabb az avarok bomlása. Feltételezésem szerint a nyári magasabb hőmérséklet mellett az avar gyorsabb fogyásának oka lesz a megnövekedett aprító szám is, valamint, hogy mivel bőven áll majd rendelkezésre táplálék számukra, tényleg csak azt az avarféleséget fogják fogyasztani, amelyik előnyösebb számukra.
Hipotézisem szerint mind télen, mind nyáron a makrogerinctelenek által is bontott avar fogyása jóval gyorsabb lesz, mintha csak a mikrogombák végzik.
3.4 Saját fejlesztésű terepi eszköz hatékonyságának összevetése a hagyományos módszerekkel
Egyik fő terepi tapasztalatom az volt, hogy egy-egy zivatar után pár órára jelentősen megnő a hordalék mennyisége a patakokban, melyek lerakódnak az avarzsákokra, ezáltal befolyásolják a bennük lévő avar bomlását. Ez a probléma vezetett az avarhenger kitalálásához. Hipotézisem szerint az avarhenger palástja azáltal, hogy merőlegesen helyezkedik el a sodrás tengelyére, megakadályozza azt, hogy a szediment rárakódjon a kihelyezett avarra. Feltételezésem szerint az avarhengerben kiegyenlítettebben folyhat az avarbomlás, a természeteses viszonyokhoz hasonlóbb körülmények között. Ezzel párhuzamosan a többi terepi eszköz használata kapcsán a hipotézis az volt, hogy az avardoboz a legkevésbé célszerű eszköz dombvidékeken az avarbomlás vizsgálatára, ennél praktikusabb használhatóságú az avarzsák, és a legjobb használhatóságú az avarhenger lesz, tehát ebben a sorrendben illeszkedik majd a legjobban az avarok exponenciális fogyási görbéi a klasszikus görbelefutáshoz.
19 3.5 Avarbomlás kezdeti szakaszának vizsgálata
Az avar fogyását általában hosszabb időtávon érdemes vizsgálni, ha a bomlás ütemére és a funkcionális táplálkozási csoportok összetételére vagyunk kíváncsiak az adott víztérben.
Azonban az avar behullásától fogva már kis idő elteltével is jelentős változások állhatnak elő, pl. a víz alacsonyabb hőmérséklete miatt előálló aprózódás, vagy a levéllemezből való szerves anyag kioldódás. Feltételeztem, hogy a kezdeti sűrűbb mintavételezésnek köszönhetően a bomlási ütemek nagyon fognak hasonlítani majd az ideális exponenciális görbéhez. Valamint feltételeztem, hogy az első két hétben már jelentős lesz az aprítók kolonizációja.
20
4 Anyag és módszer
4.1 Mintavételi helyszínek
Az öt fő kísérletsorozatom öt kisvízfolyás tíz helyszínén folyt. A kijelölt patakszakaszok többsége természetes jellegű. Az egyes szakaszokon több kísérlet is folyt (1. táblázat), de nem egy időben. Az alábbiakban, általánosságban bemutatom a kísérleti helyszíneket.
1. táblázat Az egyes kísérletek helyszíneinek megjelölése
Kisvízfolyás neve Kód 1. kísérlet 2.
kísérlet
3.
kísérlet
4.
kísérlet
5.
kísérlet Víztest jellege
Cuha-patak C1 x természetes
Cuha-patak C2 x természetes
Torna-patak T1 x természetes
Torna-patak T2 x természetes
Torna-patak T3 x mesterséges
Torna-patak T4 x x mesterséges
Csigere-patak Cs1 x természetes
Csigere-patak Cs2 x x x x módosított
Veszprémi-séd VpS x természetes
Vázsonyi-séd VzS x x x természetes
A Cuha-, a Csigere és a Torna-patak (1. ábra) a Bakonyban (Bakony északi és délnyugati részén) fut, hosszuk 12-63 km között változik. A kísérleti helyszínek általános jellemzői reprezentálják a patakszakaszok jelentős részét.
A Cuha-patakon két kísérleti helyszínt jelöltem ki. Az első ponton (C1, Függelék 1. kép), a folyómeder anyaga szemcseméretét tekintve megalital (Ø > 40 cm, nagyobb kövek, tömbök), makrolital (Ø: 20 - 40 cm, durva kövek) és mezolital (Ø: 6 - 20 cm, ökölnyi kövek) keveréke. A patak az erdőben fut, melynek uralkodó fafajai a Quercus robur, a Quercus petraea és a Carpinus sp.
A másik kijelölt Cuha-patak ponton (C2, 1. ábra, Függelék 2. kép), a fent említett három fafajon kívül a Populus tremula is jelentős egyedszámmal képviselteti magát. Az patak aljzata ezen a szakaszon akal (Ø: 2 mm - 2 cm, finom és közepes méretű kavics), psammal (Ø: 6 µm - 2 mm, homok), és néhány ponton argyllal (Ø < 6 µm agyag), a patak meanderezik, sodrása lassabb, mint a C1-es helyszínen.
A Csigere-patak első pontján (Cs1, 1. ábra, Függelék 3. kép) az uralkodó fafaj a P. tremula.
A meder kanyarog, az aljzatot akal (Ø: 2 mm - 2 cm, kavics), psammal (Ø: 6 µm - 2 mm, homok, iszap), és néhány ponton argyllal (Ø < 6 µm iszap) alkotja. A Csigere-patak második pontján (Cs2, 1. ábra, Függelék 4. kép) nincs természetes fás vegetáció, csak szórványosan található néhány Salix caprea és Populus tremula egyed. Az aljzat anyaga microlital (Ø: 2 cm - 6 cm, durva kavics) és akal (Ø: 2 mm - 2 cm, finom és közepes méretű kavics). A meder enyhén kanyarog, a part menti vegetáció igen szegényes. A Cs2-es pont az 1.
táblázatban módosított helyszínként (jelen van több fontos természetességre és módosításra utaló elem is) jelöltem, ennek oka az, hogy a meder természetes jellegű mind lefutását, mind szemcseméretét tekintve, míg a partja módosított, egyrészt a szántóföldek másrészt a kaszálások és a fás vegetáció hiánya miatt.
21 A Torna-patakon összesen négy vizsgálati helyszín volt, melyből két természetes jellegű, hasonlóan a Cuha-patak C1 és C2, valamint a Csigere-patak Cs1 természetes és átmeneti jellegű Cs2 pontjaihoz. A két természetes helyszínen (T1 és T2, 1. ábra Függelék 5. és 6.
kép) többek között Betulaceae fajok alkotják a Bakonyra jellemző, patakot kísérő erdőt. A meder mikrolital (Ø: 2 - 6 cm, durva kavics) és akal (Ø: 2 mm - 2 cm, finom és közepes méretű kavics) mindkét helyen. A másik két vizsgálati helyszín a Torna-patak (T3 és T4, 1.
ábra, Függelék 7. és 8. kép) erősen módosított szakaszán lett kijelölve, ahol a meder kiegyenesített, trapéz alakú. A T4-es helyszínen a trapézmeder alja és oldala betonlapokkal kirakott. A mederanyag főként mikrolital (Ø: 2 cm - 6 cm) és akal (Ø: 2 mm - 2 cm). A parton semmilyen természetes fás vegetáció nem található, mert a T3-as pont mezőgazdasági területen fut, a T4-es pont pedig Devecser urbanizált területén.
Veszprémi-séd (VpS) hossza mintegy 70 km; a kísérleti helyszín a séd forráshoz közelebbi szakaszán volt kijelölve (forrástól mintegy 20 km-re), mielőtt a patak beérne Veszprém városának körzetébe (VpS, 1. ábra, Függelék 9. kép). A Veszprémi-séd enyhén kanyargó a kísérleti területen. Az aljzat mezolitalból (20%) és mikrolitalból (60%) áll. Jellemzőek a kisebb felhalmozási xylal (kisebb fatuskók, gallyak) zónák, melyek mögött feltorlódik a CPOM. A meder mentén kétoldalt 5-7 méter szélességben található fás vegetáció Salix, Sambucus, Robinia fajokkal. A Veszprémi-séd a vizsgálat helyszínén természetes jellegű, a partot szegélyező fás szárú sáv mellett kaszáló terül el, melyet néhol kiskertek szakítanak meg.
Vázsonyi-séden (VzS, 1. ábra, Függelék 10. kép) a mintavételi helyszín Nagyvázsonyon és Pula falu között található. Természetesebb környezetű a Csigere-patakhoz (Cs2) képest, mivel a patak jobb partján természetes fás vegetáció található kb. 200 méter szélességben, itt meghatározó mennyiségben vannak jelen a különböző fenyőfélék, a mezei (Acer campestre) és a hegyi juhar (Acer pseudoplatanus). Ezen kívül még elszórtan találhatunk közönséges gyertyánt (Carpinus betulus). A patak partján közvetlenül viszont a kosárfonó fűz (Salix viminalis) és a rezgő nyár (Populus tremula) a leggyakoribb fafaj, de 14 lombhullató fafaj és különböző lágyszárúak és fenyőfélék is képviseltetik magukat (Vass 2010).
1. ábra A kísérlet helyszínei