SZEGEDI TUDOMÁNYEGYETEM
TERMÉSZETTUDOMÁNYI ÉS INFORMATIKAI KAR MIKROBIOLÓGIAI TANSZÉK
BIOLÓGIA DOKTORI ISKOLA
TRICHODERMA TÖRZSEKEN ALAPULÓ NÖVÉNYVÉDŐ ÉS NÖVÉNYNÖVEKEDÉST SERKENTŐ BIOEFFEKTOROK FEJLESZTÉSE
Ph.D. értekezés
Körmöczi Péter
Témavezető:
Dr. Kredics László egyetemi docens
Szeged 2016
2 TARTALOMJEGYZÉK
Tartalomjegyzék ... 2
1. Rövidítések ... 5
2. Bevezetés ... 6
3. Irodalmi áttekintés ... 8
3.1. A Trichoderma fajok általános előfordulása ... 8
3.2. A Trichoderma nemzetség taxonómiai áttekintése ... 8
3.2.1. A Trichoderma nemzetség molekuláris taxonómiája ... 9
3.2.2. A Trichoderma nemzetség szekcióinak jellemzése ... 10
3.3. A növénypatogén gombák jelentősége, kártétele, védekezési stratégiák ... 11
3.3.1. A vizsgálatainkban szereplő növénypatogén gombák és mezőgazdasági kártételük ... 11
3.3.2. A növénypatogén gombákkal szembeni lehetséges védekezési stratégiák ... 13
3.4. A Trichoderma fajok jelentősége a mezőgazdaságban ... 14
3.5. A Trichoderma fajok biokontroll mechanizmusai ... 15
3.5.1. Fungisztázis a talajban ... 16
3.5.2. Tápanyagokért folytatott verseny ... 17
3.5.3. Mikoparazitizmus ... 17
3.5.4. Sejtfalbontó enzimek és szabályozásuk ... 20
3.5.5. Antibiózis, a másodlagos anyagcseretermékek szerepe a biológiai védekezésben ... 23
3.6. A lakkáz enzim előfordulása, hasznosítási lehetőségei ... 23
3.6.1. A Trichoderma fajok lakkáztermelése ... 23
3.7. A különféle fungicidek és azok csoportosítása hatásmechanizmusuk alapján ... 24
3.8. A növények növekedésének és fejlődésének serkentése és ennek mechanizmusai ... 25
3.8.1. Rizoszféra-kompetencia ... 25
3.8.2. A gyökér kolonizációja ... 27
3.8.3. Az ásványi anyagok szolubilizálása, tápanyagfelvétel növelése ... 27
3.8.4. A klorofilltartalom és a fotoszintetikus aktivitás fokozása ... 28
3.8.5. Az abiotikus stresszhatások mértékének csökkentése ... 28
3.8.6. Másodlagos metabolitok termelése ... 28
3.8.7. Növényi hormonok bioszintézisének fokozása ... 29
4. Célkitűzések ... 30
5. anyagok és módszerek ... 31
5.1. Mintavételezés, mintavételi helyek ... 31
5.2. Trichoderma törzsek izolálása és azonosítása ... 31
3
5.2.1. Trichoderma törzsek izolálása a zöldségrizoszféra-mintákból ... 31
5.2.2. Genomi DNS kivonása az izolált Trichoderma törzsekből ... 32
5.2.3. Az izolált Trichoderma törzsek azonosítása ... 32
5.3. Agar-konfrontációs teszt ... 33
5.4. Különböző környezeti paraméterek hatása a micéliumnövekedésre ... 34
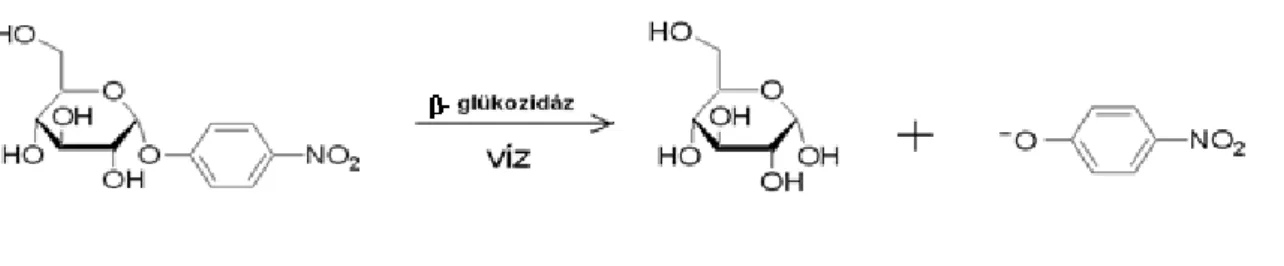
5.5. A Trichoderma törzsek cellulózbontó és foszfátmobilizáló képességének vizsgálata ... 35
5.6. A Trichoderma törzsek által termelt lakkáz enzimek vizsgálata ... 35
5.6.1. A Trichoderma törzsek lakkáz enzimeinek kvalitatív vizsgálata ... 35
5.6.2. A Trichoderma törzsek lakkáz-termelésének kvantitatív vizsgálata ... 36
5.6.3. Trichoderma törzsek által termelt lakkázok pH-függésének vizsgálata ... 36
5.7. Különböző fungicidek minimális gátló koncentráció (MIC) értékeinek meghatározása ... 36
5.8. Paradicsomnövényeken végzett kísérletek és mérések ... 37
5.8.1. Növénynevelés és a kísérleti elrendezés ... 37
5.8.2. A paradicsomnövények biomassza-produkciójának a vizsgálata ... 38
5.8.3. A sztómakonduktancia és a CO2-asszimiláció meghatározása ... 38
5.8.4. A klorofill a fluoreszcencia indukciós paramétereinek meghatározása ... 38
5.8.5. A fotoszintetikus pigmenttartalmak meghatározása ... 39
5.8.6. Az összcukortartalom meghatározása ... 39
5.9. Az adatok feldolgozása és értékelése ... 40
6. Eredmények és megvitatásuk ... 41
6.1. Törzsek izolálása, az izolált törzsek változatossága ... 41
6.1.1. A részletes vizsgálatainkhoz felhasznált Trichoderma törzsek kiválasztásának szempontjai ... 43
6.2. A Trichoderma törzsek in vitro antagonizmusa ... 43
6.3. A vizsgált Trichoderma törzsek ökofiziológiai jellemzése ... 46
6.3.1. A hőmérséklet hatása a micéliumnövekedésre ... 47
6.3.2. A pH hatása a micéliumnövekedésre ... 49
6.3.3. A vízaktivitás hatása a micéliumnövekedésre ... 50
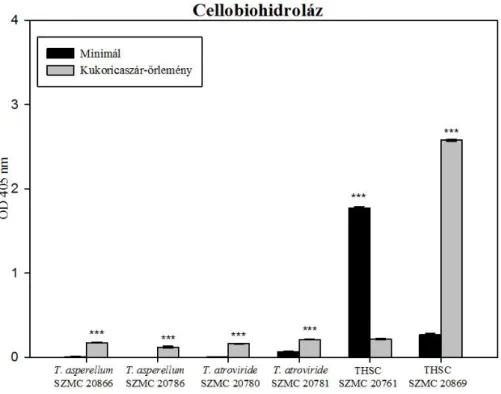
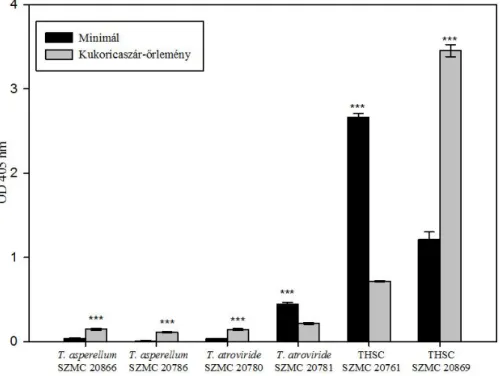
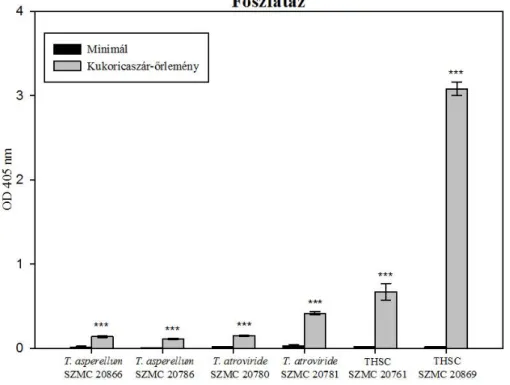
6.4. Cellulózbontó és foszfátmobilizáló képesség vizsgálata ... 54
6.4.1. A Trichoderma törzsek cellulózbontó képességének vizsgálata ... 54
6.4.2. A Trichoderma törzsek foszfatáz aktivitásának vizsgálata ... 57
6.5. A Trichoderma törzsek lakkáztermelésének vizsgálata ... 58
6.5.1. Lakkáztermelő Trichoderma törzsek azonosítása ... 58
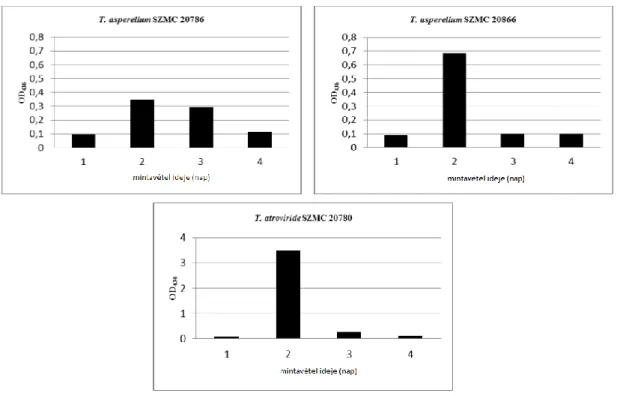
6.5.2. Az igazolt lakkáztermelő törzsek lakkáz-aktivitásának vizsgálata... 58
6.5.3. A lakkáz-aktivitás pH-függésének kimutatása ... 60
6.6. A törzsek fungicidekkel szembeni érzékenységének vizsgálata ... 61
4 6.7. A T. asperellum SZMC 20786-os törzs paradicsomnövények növekedésére és
fotoszintetikus aktivitására gyakorolt hatásának vizsgálata ... 63
6.7.1. A T. asperellum SZMC 20786 törzzsel történt kezelés paradicsomnövények biomassza- produkciójára gyakorolt hatásának vizsgálata ... 63
6.7.2. A T. asperellum SZMC 20786 törzzsel történő kezelés hatása a paradicsomnövények fotoszintetikus aktivitására ... 65
6.8. Növényvédő és növénynövekedést serkentő bioeffektor Trichoderma törzsek kiválasztása, termékfejlesztés ... 68
7. Összefoglalás ... 72
8. Summary ... 76
9. Irodalomjegyzék ... 80
10. Köszönetnyílvánítás ... 101
5 1.RÖVIDÍTÉSEK
6-PP 6-pentil-alfa-piron
ABTS [2,2′-azinobisz-(3-etilbenztiazolin-6-szulfonát)]
act aktint kódoló gén ATP adenozin-trifoszfát aw vízaktivitás
BCA Biological Control Agent – biokontroll ágens BCI Biokontroll Index
bp bázispár
cal kalmodulint kódoló gén DMSO dimetil-szulfoxid ech42 endokitinázt kódoló gén
FHB kalászfuzáriózis („Fusarium head blight”) FOSC F. oxysporum fajkomplexum
FSSC F. solani fajkomplexum
FT frisstömeg
Fv/Fm A második fotokémiai rendszer maximális kvantumhasznosítása
G guanozin
GPCR G protein-kapcsolt receptorok („G protein-coupled receptors”) HPLC Nagyteljesítményű folyadékkromatográfia
IES Indolecetsav
IGS Intergenic Spacer – gének közti szakasz
ITS riboszomális RNS génklaszter köztes átíródó elválasztó régiója („Internal Transcribed Spacer”)
MAMP mikróba-asszociált molekuláris mintázat („microbe-associated molecular pattern”)
MAPK mitogén-aktiválta protein kináz
MEA malátakivonatos táptalaj („Malt extract agar”)
MIC minimális gátló koncentráció („Minimal Inhibitory Concentration”) NCBI National Center for Biotechnology Information
NPQ nem fotokémiai kioltás
NRPS nem riboszómális peptid-szintetáz („nonribosomal peptide synthetase”) OD optikai denzitás
PCR polimeráz láncreakció („polymerase chain reaction”)
Phi PSII a második fotokémiai rendszer effektív kvantumhasznosítása
PK poliketid
PKS poliketid-szintáz
PSII a második fotokémiai rendszer qP fotokémiai kioltás
ROS reaktív oxigénfajták („reactive oxygen species”) rpb2 RNS polimeráz II
SM másodlagos anyagcseretermékek („secondary metabolites”) SSCP szubtilizin-szerű S8 proteáz
T timin
tef1 transzlációs elongációs faktor 1-alfa alegysége THSC T. harzianum fajkomplexum
VOC illékony szerves vegyület („volatile organic compound”) YEG élesztő-glükózos táptalaj („yeast extract - glucose agar”)
6 2.BEVEZETÉS
A Trichoderma nemzetség számos képviselője az emberi jólétre jelentős hatást gyakorló mikroorganizmusok közé sorolható. A Trichoderma fonalasgombák alkalmazására több lehetőség is nyílik. A Trichoderma törzsek közül kerülnek ki a leggyakrabban használt biofungicidek és növénynövekedés-serkentők, ezen túl számos iparág által hasznosított extracelluláris enzimek forrásai is lehetnek. Változatos másodlagos metabolitok termelésére képesek, néhány fajuk klinikai jelentőséggel is rendelkezik, míg más törzseket úgy módosítottak, hogy általuk lehetőség nyíljon fehérjék heterológ megtermeltetésére. A talaj bioremediációjában is szerepet játszanak a szerves és szervetlen vegyületek, valamint nehézfémek felvételének szabályozása révén (Schuster és Schmoll, 2010; Harman 2011a,b; Mukherjee és mtsai., 2013).
Az utóbbi évtizedekben kétségkívül megnövekedett a Trichoderma fajokról szóló tanulmányok, valamint szabadalmak száma. A „Google Scholar Database” adatbázis a
„Trichoderma” keresőszóra a 2013-as évben ~138 ezer találatot adott ki, míg ez a szám a 2016-os évre közel duplájára emelkedett, elérve a ~269 ezer találatot. A nemzetséget még az 1794-es évben Persoon (Persoon, 1794; Mukherjee és mtsai., 2013) írta le, az egyes fajok hasznosításában rejlő lehetőségeket azonban csak az 1930-as években ismerték fel.
Mindez Weindling (1932, 1934) munkásságának köszönhető, aki bizonyította a Trichoderma fajok mikoparazitizmusát, antibiotikum-termelését és a biológiai védekezés céljaira történő alkalmazásuk lehetőségét. Az elkövetkező 30 évben viszonylagos csend állt be a Trichoderma törzseken alapuló biológiai védekezéssel kapcsolatos vizsgálatok területén, míg az 1970-es években szabadföldi kísérletek során be nem bizonyították hatékonyságukat. Napjainkban már a legtöbbet, és legszélesebb körben tanulmányozott mikroorganizmusok közé tartoznak, ami a rendelkezésre álló szakirodalom mennyiségéből is jól látható.
Kezdetben a Trichoderma törzsek azonosítása során főleg a morfológiai jegyekre támaszkodtak, ami a nemzetség pontatlan taxonómiájához vezetett. Áttörést a molekuláris biológiai módszereken, köztük a vonalkód-szekvenciákon alapuló módszerek („barcoding”) hoztak (http://www.isth.info/). Ma már számos, pontosan beazonosított törzs érhető el különböző törzsgyűjteményekben, és nagy mennyiségben állnak rendelkezésre DNS-szekvenciaadatok is. A Trichoderma törzsek esetében sajnos a pontos, fajszintű azonosítást sokszor elhanyagolják, pedig esetükben ez két okból is igen fontos lenne: 1.
egyes előnyös tulajdonságok faj- illetve törzsspecifikusak lehetnek, ennél fogva fontos a
7 pontos azonosítás az egyes törzsekben rejlő összes lehetőség maximális kihasználása érdekében; 2. a pontos azonosítás még fontosabb a biztonsági szempontok szem előtt tartása miatt, hogy az alkalmazni kívánt törzs a gyártóra, a felhasználóra és a környezetre se jelentsen kockázatot. Itt meg kell említeni, hogy a rendelkezésre álló technikák, valamint internetes adatbázisok ellenére sajnálatos módon a kereskedelemben elérhető Trichoderma-alapú készítményekben jelenlevő törzsek jelentős része tévesen, vagy semmilyen formában sem lett azonosítva.
8 3.IRODALMI ÁTTEKINTÉS
3.1. A Trichoderma fajok általános előfordulása
A Trichoderma nemzetség képviselői kozmopoliták, széles körben elterjedtek és világszerte számos ökoszisztéma állandó tagjai (Kubicek és mtsai., 2008; Kredics és mtsai., 2014). A Trichoderma fajok előfordulásában különböző faktorok játszanak szerepet, melyek között megtaláljuk az adott élőhely mikroklímáját, a rendelkezésre álló szubsztrátumokat, valamint komplex ökológiai kölcsönhatásokat (Hoyos-Carvajal és Bissett, 2011). A Trichoderma gombák túlélése az eltérő földrajzi adottságokkal rendelkező élőhelyeken a rendelkezésre álló tápanyagok széles körben történő hasznosításával, a magas reproduktív kapacitással, és a hatékony kompetitív tulajdonságokkal hozható összefüggésbe (Cardoso Lopes és mtsai., 2012). Változatos élőhelyekhez történő alkalmazkodásra való képességüket szemlélteti, hogy izoláltak már Trichoderma törzseket tengeri szivacsokból (Paz és mtsai., 2010; Gal-Hemed és mtsai., 2011), fűfélékből és lágyszárú növényekből, valamint endofita gombaként is számon tartják őket (Samuels és mtsai., 2006a; Zhang és mtsai., 2007; Mulaw és mtsai., 2010). A nemzetség képviselői a különféle gombafajokról, korhadt fákról, fakéregről, talajból és a növények rizoszférájából leggyakrabban izolált gombafajok között is jelen vannak.
Gombaellenes hatásuk a nagy mennyiségben termelt extracelluláris enzimeknek és antibiotikus hatású vegyületeknek köszönhetően érvényesül, így képesek váltani a szaprotróf és biotróf életmód között. A jelenleg rendelkezésre álló szakirodalmi adatok alapján az összes Trichoderma faj képes biotróf kapcsolat kialakítására (Jaklitsch, 2009;
Druzhinina és mtsai., 2011; Druzhinina és Kubicek, 2013).
3.2. A Trichoderma nemzetség taxonómiai áttekintése
A Trichoderma fajok taxonómiája (Hypocraceae, Hypocreales, Ascomycota) rövidebb múltra tekint vissza, mint a korábbi taxonómiai terminológiában használt teleomorf Hypocrea nemzetségé. Kezdetben a Trichoderma fajok azonosítása különböző morfológiai bélyegek alapján történt. Ez alapján az egyes fajok elkülönítése igen nehéznek bizonyult a viszonylag kevés eltérő morfológiai karakternek köszönhetően, ezáltal átfedések jöttek létre a fajok között, ami pontatlan fajazonosításokhoz vezetett (Samuels, 2006). Mivel a morfológiai alapon történő azonosítás még napjainkban is komoly kihívás, a nemzetséggel kapcsolatos áttörést a molekuláris alapon történő azonosítás hozta meg.
Samuels (2006), valamint Druzhinina és Kopchinski (2006) a vonalkód-szekvenciákon
9 alapuló meghatározás segítségével 100 Trichoderma faj jellemzését végezték el. Ezt követően a Trichoderma törzsek diverzitásának vizsgálatát célzó tanulmányok sokkal nagyobb jelentőségre tettek szert, mellyel párhuzamosan kiterjedt mintavételezés folyt az egész világon (Chaverri és mtsai., 2003a; Hoyos-Carvajal és mtsai., 2009; Jaklitsch, 2009, 2011; Migheli és mtsai., 2009; Gal-Hemed és mtsai., 2011), így a nemzetség taxonómiája rövid idő alatt nagy fejlődésen ment keresztül.
3.2.1. A Trichoderma nemzetség molekuláris taxonómiája
A nukleinsav-polimorfizmus molekuláris módszereken alapuló jellemzése nagyon nagy számban biztosít lehetőséget a potenciális markerek taxonómiai vizsgálatok céljaira történő azonosítására (Lieckfeldt és mtsai., 1998). Az 1990-es évek végére a szekvenciaelemzés elfogadott és standard módszerré vált a gomba-rendszerbiológiában (Fujimori és Okuda, 1994; Kuhls és mtsai., 1996; Kindermann és mtsai., 1998; Dodd és mtsai., 2000), melynek következtében a Trichoderma fajok azonosítása is egyre megbízhatóbbá vált (Druzhinina és Kubicek, 2005; Druzhinina és mtsai., 2006; Samuels, 2006; Jaklitsch, 2009) és folyamatosan növekedett az azonosított fajok száma. Az első 100 molekulárisan jellemzett Trichoderma fajról 2006-ban nyújtottak áttekintést Druzhinina és munkatársai (2006).
A nemzetséget érintő első molekuláris taxonómiai vizsgálatok során a riboszomális RNS génklaszter köztes átíródó elválasztó (ITS) régiójának 1-es, majd később a 2-es szakaszát, valamint az 5,8S rRNS-t kódoló gént használták fel (Kindermann és mtsai., 1998; Lieckfeldt és mtsai., 1998; Dodd és mtsai., 2000). Az ITS-szekvenciák ugyan nem biztosítanak megfelelő mértékű filogenetikai felbontást, de alkalmasnak bizonyultak a vonalkód-alapú meghatározáshoz (Druzhinina és mtsai., 2005; Kredics és mtsai., 2014).
Az ITS1 és ITS2 szekvenciákban megtalálható „horgony”-szakaszok azonosítása az interneten elérhető TrichOKey program (www.isth.info) segítségével lehetővé teszi a leggyakrabban előforduló fajok azonosítását. Mindezek mellett újabb és újabb markerek jelentek meg, amelyek segítségével növelni lehetett a filogenetikai fák felbontását. Ezen markerek között megtalálhatóak a 18S, valamint a 28S RNS-t kódoló szakaszok, a transzlációs elongációs faktor 1-alfa (tef1) különböző szakaszai, továbbá az endokitinázt (ech42=chi18-5) (Lieckfeldt és mtsai., 2000; Kullnig-Gradinger és mtsai., 2002), az RNS polimeráz II-t (rpb2) (Chaverri és mtsai., 2003a), az aktint (act) és a kalmodulint (cal1) (Samuels és mtsai., 2006a,b; Jaklitsch és mtsai., 2008a) kódoló gének egyes szakaszai. A nemzetségen belül ezen markerek közül a filogenetikai vizsgálatok céljaira a tef1 gén 4. és
10 5. intronjai bizonyultak a legalkalmasabbnak (Jaklitsch és mtsai., 2006a,b; Samuels és mtsai., 2006a; Jaklitsch, 2009, 2011).
3.2.2. A Trichoderma nemzetség szekcióinak jellemzése
A T. longibrachiatum és a T. pseudokoningii külön-külön a Rifai által 1969-ben létrehozott 9 fajaggregátum részét képezték (Samuels és mtsai., 2012), később pedig a Longibrachiatum szekció alapjául szolgáltak. Kuhls és munkatársai 1996-ban írták le a T.
reesei-t, mint a szekció új képviselőjét, melynek az ivaros (teleomorf) alakja korábban Hypocrea jecorina-ként volt ismert. Druzhinina és munkatársai 2012-ben elvégezték a Longibrachiatum szekció felülvizsgálatát, és az újonnan leírt fajokkal együtt 21 önálló fajt soroltak a szekcióba.
A legtöbb Trichoderma faj a Trichoderma szekcióba tartozik (Kulling-Gradinger és mtsai., 2002, Druzhinina és Kubicek, 2005). Ez a szekció alapvetően megegyezik a Bissett (1991b) által bevezetett szekcióval, később azonban számos faj került át ide, főleg a Pachybasium szekcióból, többek között a T. hamatum, a T. pubescens és a T. strigosum (Kindermann és mtsai., 1998; Dodd és mtsai., 2000; Lieckfeldt és mtsai., 2001; Kulling- Gradinger és mtsai., 2002). A közelmúltban a T. koningii szubkládot, valamint a Viride és a Viridescens kládot írták le a szekció új tagjaiként (Jaklitsch és mtsai., 2006a; Samuels és mtsai., 2010). Később Jaklitsch (Jaklitsch, 2009, Jaklitsch és mtsai., 2012) munkásságának köszönhetően a Trichoderma szekció újabb 27, majd 43 fajjal bővült.
A Pachybasium szekciót Bissett (1991a) a T. hamatum-ra (korábban Pachybasium hamatum) alapozta, melyet később további 20 fajjal, többek között a T. harzianum-mal, T.
piluliferum-mal, T. polysporum-mal és a teleomorf H. gelatinosa–val bővített (Bissett, 1991b). A szekcióról azonban kiderült, hogy parafiletikus (Kindermann és mtsai., 1998;
Kulling-Gradinger és mtsai., 2002), ezért további 5 kládra osztották fel. A T. hamatum-ot áthelyezték a Trichoderma szekcióba, az összes többi hozzá hasonló morfológiájú fajt pedig egymástól független kládokba (Ceramica, Chlorospora, Harzianum, Semiorbis, Strictipilosa és Stromaticum) sorolták (Chaverri és Samules, 2003; Jaklitsch, 2009;
Jaklitsch, 2011). A T. hamatum áthelyezésével a Bissett (1991a) által létrehozott szekciónevet már nem használták tovább. A szekciót később Lu és munkatársai (2004) szervezték újjá a T. polysporum/H. pachybasioides köré, a T. minutisporum-mal és egyéb újonnan leírt fajokkal együtt. A szekció a Pachybasioides nevet kapta, ám később Jaklitsch-nak (2011) köszönhetően Pachybasium „core” központi csoportnak nevezték el,
11 mely tovább bővült a Podostroma és Podocrea nemzetségekkel (Chamberlain és mtsai., 2004; Jaklitsch és mtsai., 2008b).
A Hypocreanum szekciót Bissett (1991a) a T. lacteum-ból eredeztette. A későbbiek folyamán Overton és munkatársai (2006a,b) vizsgálták felül a szekciót, majd a legújabb multilókusz-vizsgálaton alapuló filogenetikai elemzés során 13 fajt soroltak a Hypocreanum szekcióba, többek között a T. austriacum-ot, a T. citrinum-ot, a T. decipiens- t, a T. phellinicola-t, a T. pulvinatum-ot és a T. sulphureum-ot.
A Bissett által felállított szekciókon túl számos további kládot hoztak létre. A legnagyobb ezek közül a Harzianum klád, melybe zöld aszkospórákkal rendelkező fajok tartoznak (Jaklitsch, 2009; Druzhinina és mtsai., 2010). A klád fő képviselője a kozmopolita T. harzianum sensu lato fajkomplex, amely nagyfokú filogenetikai változatosságot mutat. További filogenetikai vizsgálatokat Chaverri és Samuels (2002, 2003), valamint Chaverri és munkatársai (2003b) végeztek, majd Druzhinina és munkatársai (2010) tovább folytatták a klád vizsgálatát. Druzhinina és munkatársai (2010) vizsgálataik alapján két, a H. lixii-től különböző, genetikailag eltérő anamorf fajt igazoltak, a T. harzianum sensu stricto-t és a korábban még le nem írt T. „afroharzianum”-ot. Mivel egyértelműen kiderült, hogy a H. lixii és a T. harzianum sensu stricto nem egy faj, a H. lixii – T. harzianum teleomorf – anamorf kapcsolatot megszüntették (Druzhinina és mtsai., 2010). A H. lixii és a T. harzianum sensu stricto mellett a klád 18 fajt számlál, melyek között megtalálható a T. aggressivum (Samuels és mtsai., 2002), a T. pleuroti és a T.
pleuroticola, melyek termesztett gombák zöldpenészes fertőzéséért tehetők felelőssé (Park és mtsai., 2006; Komoń-Zelazowska és mtsai., 2007).
A Harzianum klád mellett további 8 kisebb klád (Chlorospora, Spinulosa, Strictipilosa, Ceramica, Semiorbis, Brevicompactum, Psychrophyla, valamint Lutea) került még meghatározásra (Chaverri és Samuels, 2003; Degenkolb és mtsai., 2008; Jaklitsch 2009, 2011).
3.3. A növénypatogén gombák jelentősége, kártétele, védekezési stratégiák
3.3.1. A vizsgálatainkban szereplő növénypatogén gombák és mezőgazdasági kártételük
A takarmánynövényeket érintő betegségek óriási jelentőséggel rendelkeznek a mezőgazdaságban, és évről-évre súlyos terméskieséseket és gazdasági károkat képesek okozni (Viterbo és Horwitz, 2010; Mesterházy és mtsai., 1999). A talajban előforduló és
12 azon keresztül fertőzni képes mikroorganizmusok közül is a talajgombák a legjelentősebbek (Viterbo és Horwitz, 2010).
A Fusarium nemzetség a növénypatogén gombák egy nagyon változatos csoportját alkotja, képviselői széles gazdaspektrummal és földrajzi elterjedtséggel rendelkeznek (Doohan és mtsai., 2003). A növénypatogén Fusarium-ok a gabonák számos különböző megbetegedését képesek előidézni, többek között a palánták levélfoltosodását („seedling blight”), fuzáriumos tőrothadását („foot rot”), valamint a kalászfuzáriózist (FHB –
„Fusarium head blight”). A kukoricanövények legtöbb részét is képesek megfertőzni a csírázás kezdetétől egészen az érés folyamatának legvégéig (Sutton, 1982; Parry és mtsai., 1995).
A Fusarium fajokhoz hasonlóan az egész világon nagy számban okoznak fertőzéseket az Alternaria nemzetség képviselői. Gazdanövényeik közé sorolhatunk számos gazdaságilag jelentős zöldséget és gyümölcsöt (pl. alma, brokkoli, karfiol, répa, burgonya, paradicsom) (Johnson és mtsai., 2001; Mora és Earle, 2001; Weir és mtsai., 1998). Számos dísznövényt és fűfélét is képesek megfertőzni (Laemmlen, 2001a). Az Alternaria fajok által okozott fertőzések leggyakrabban a növények föld feletti részein következnek be. A fertőzés legelső, kezdeti tünete egy apró fekete pont megjelenése a levél felszínén, melynek mérete a fertőzés előrehaladtával folyamatosan növekszik.
Különféle környezeti körülmények között képes a növények körében globális problémát okozni a gombák által kiváltott palántadőlés is. A palántadőlés elnevezése a hajtás és a gyökér szöveteinek szétesésére, vizenyőssé és puhává válására utal (Laemmlen, 2001b). A legtöbb palántadőlést kiváltani képes növénypatogén gomba a kifejlett növényen is képes különféle fertőzések kialakítására. A palántadőlésért felelős két leggyakoribb gomba a Rizoctonia solani, valamint a Pythium-ok, de a betegséget okozhatják Botrytis, Sclerotinia, Alternaria, Phytophthora és Fusarium fajok is (Laemmlen, 2001b).
Több mint 500 gazdanövénnyel (mezőgazdasági-, és dísznövények, fák) és nagyfokú negatív gazdasági hatással rendelkezik a Rhizoctonia solani (Anderson, 1982;
Sneh és mtsai., 2013; Kai és mtsai., 2007). A 2000-es években Európában drámai növekedés következett be a betegség előfordulásainak számában, ezáltal meghatározó szereplőjévé vált a növénypatogén gombák által kiváltott fertőzéseknek (Grosch és mtsai., 2005). A R. solani szaprotróf és nekrotróf törzsei széles körű földrajzi elterjedtséggel rendelkeznek, szkleróciumaik igen ellenállóvá teszik a szélsőséges környezeti behatásokkal szemben (Kai és mtsai., 2007).
13 A tökfélék ismert kártevője a Phoma cucurbitacearum, amely a növények növekedése során a ragacsos szárfoltosságot idézi elő, a termés megjelenését követően pedig fekete rothadást képes kiváltani (Keinath és mtsai., 1995). A Cucurbita nemzetségen belül a Phoma törzsek által okozott fekete rothadásnak a sütőtök esetében is jelentős gazdasági szerepe van, ugyanis betakarítás előtt és a raktározás során is súlyos károkat képes okozni (Zitter és Kyle, 1992).
A levegőben a szél és az eső által terjesztett egyik legjelentősebb nekrotróf növénypatogén gomba a Botrytis cinerea, amely több mint 200 gazdanövénnyel rendelkezik világszerte (Williamson és mtsai., 2007a), köztük facsemetékkel, zöldségekkel, dísz-, és szántóföldi növényekkel, de a gyümölcsösök is ki vannak téve a Botrytis törzsek által okozott lágyrothadásnak és szürkepenészes megbetegedésnek. A Botrytis törzsek betakarítás előtt és után is egyaránt képesek a fertőzés kiváltására, mely a legtöbb esetben a növény föld feletti részein következik be (Williamson és mtsai., 2007b).
A kifejlett, vagy már öregedő fázisban lévő kétszikű növények vannak leginkább kitéve a fertőzéseknek. A Botrytis elleni védekezés igen nehézkes, mivel a talajban micélium és konídium formájában is huzamosabb ideig túlél, valamint rövid idő alatt kialakulhat rezisztencia a védekezéshez használt fungicidekkel szemben, ezért a legtöbb esetben kombinált kezeléseket alkalmaznak (Williamson és mtsai., 2007a).
3.3.2. A növénypatogén gombákkal szembeni lehetséges védekezési stratégiák
A növénypatogén gombák ellen történő védekezéshez elengedhetetlenül szükséges, hogy alapos információval rendelkezzünk a növénypatogén kórokozóról, a gazdanövényről és a fertőzés kialakulásához szükséges környezeti feltételekről (Koike és mtsai., 2003).
Egy hatásos védekezési stratégiának mindenféleképpen gazdaságosnak is kell lenni, a védekezésre fordított összeg nem haladhatja meg a betakarítást követően a terményből származó hasznot (Koike és mtsai., 2003). Ezért a védekezés során nagyon nagy a jelentősége az elővigyázatosságnak és a növények folyamatos megfigyelésének, különös tekintettel azokra a növényekre, amelyeken már jelentkeznek valamely fertőzés kezdeti tünetei. A fertőzéssel szembeni beavatkozás időzítése is kulcsszereppel bír. Mindezek mellett ahhoz, hogy a védekezés gazdaságos legyen, az alábbi feltételeknek is meg kell felelni: legyen egyszerű, biztonságos és költséghatékony, továbbá kellően hatásos ahhoz, hogy a fertőzést, elfogadható mértékben tudja csökkenteni vagy megszüntetni. A leghatásosabb azonban mégis az, ha több különböző stratégiát alkalmazunk egyszerre (Koike és mtsai., 2003).
14 A növénytermesztés során alapvető lehet a különféle növénypatogén ágensekkel szemben ellenálló, nemesített növényfajták használata, így elkerülve a fertőzések okozta terméskieséseket (Wink, 1988; Hammond-Kosack és Parker, 2003).
A növény optimális fejlődéséhez szükséges körülmények kialakítása is fontos szereppel bír. Az ideális körülmények megteremtése során minimalizálható az esélye annak, hogy a kórokozó találkozzon a gazdanövénnyel. A vetés és ültetés előtt a megfelelő talajelőkészítés ebben az esetben elengedhetetlen. Továbbá segít, ha a vetés olyan földbe történik, ahol még nem fordult elő korábban az adott kórokozó által kiváltott megbetegedés. Emellett a megfelelő minőségű vetőmag használata is nélkülözhetetlen (Koike és mtsai., 2003).
A növénypatogénekkel szembeni védekezés eszköze a kémiai védekezés is, a vegyszerek használata azonban nagyon költséges lehet és szigorúan szabályozott, mindezen túl pedig a környezetre is kockázatot jelenthet. A kémiai védekezés eszközei közé tartozik például a vetést megelőző talajfertőtlenítés, amely hatékonyan képes a növénypatogén gombák számának csökkentésére. A csávázott vetőmagok használata is segítséget nyújt a magokat illetve a palántákat megtámadó kórokozók ellen. A kémia védekezés hátránya, hogy az adott növénypatogén gombáknál a rezisztencia hamar megjelenhet (Tjamos és mtsai., 2013).
Erre nyújthat megoldást az alább felsorolt védekezési stratégiákon túl a biológiai védekezés is, amely egyre nagyobb teret nyer a hatékony antagonista biokontroll ágenseknek köszönhetően, így megvalósítva az eredményes integrált növényvédelmet (Monte, 2001).
3.4. A Trichoderma fajok jelentősége a mezőgazdaságban
A modern mezőgazdaság sikereinek ellenére még mindig vannak olyan gyakorlatok, melyek növelik a növényeket érintő betegségek kialakulásának kockázatát és a rovarok által okozott kártétel mértékét. Ezek közé tartozik a klonális növények folyamatos monokultúrákban történő termesztése, valamint a kórokozókkal szemben érzékeny fajtanövények használata is. További problémát vet fel a nitrogénalapú műtrágyák túlzott használata, mely szintén növelheti a növényeket érintő fertőzések kockázatát (Harman és mtsai., 1996). A talajban előforduló kórokozók közül a súlyos veszteségeket okozó gombák jelentősége a legnagyobb. Számos növénypatogén gomba, pl.
a Pythium, Phytophthora, Botrytis, Rhizoctonia és Fusarium fajok kártételének mértéke
15 világszerte egyre jelentősebb. Negatív hatásuk nem kizárólag a fejlődő növényen jelentkezik, hanem az elraktározott terményeken is (Chet és mtsai., 1997). A mezőgazdaságban a vegyszerek alkalmazásával (kémiai védekezés, „chemical control”) számos gazdaságilag jelentős haszonnövényt érintő növényi kórokozót sikerült visszaszorítani. A vegyszerek használatával szemben azonban aggályok merültek fel, mivel a rovarkártevők és kórokozók fogékonysága ezekre a vegyszerekre nézve folyamatosan csökkent, és a rezisztencia is megjelent velük szemben. Mindezek mellett a kémiai növényvédőszerek a környezetre és az emberek egészségére is egyaránt káros hatást fejthetnek ki. Éppen ezért napjainkban egyre nagyobb érdeklődés övezi a különböző alternatív mezőgazdasági technikákat, melyekkel lehetőség nyílik a biológiai alapú növényvédelemre. A biológiai védekezés, vagy biokontroll („biological control”,
„biocontrol”) során egy vagy több mikroorganizmusnak (természetes ellenség, kompetitor, antagonista), vagy azok valamilyen termékének előre eltervezett és jól átgondolt alkalmazása valósul meg a különböző kártevő és kórokozó mikroorganizmusok populációinak visszaszorítása céljából. A biológiai védekezés ezen felül irányulhat a haszonnövényt érintő negatív hatások csökkentésére is. Manapság számos baktérium-, és gombaalapú biokontroll készítmény vált elérhetővé, melyek között a Trichoderma nemzetség képviselői is megtalálhatóak. A nemzetségen belül a T. virens, T. viride fajok és a T. harzianum fajkomplexum (THSC) képviselői a leggyakrabban alkalmazott biokontroll ágensek (Grondona és mtsai., 1997). A Trichoderma törzsek, mint biokontroll ágensek („biocontrol agent”, BCA) sikere számos kedvező tulajdonságukon alapul. Ilyen a magas reproduktív kapacitás, a kedvezőtlen körülmények nagyfokú tolerálása, és az a tulajdonságuk, hogy kiváló hatékonysággal képesek a különféle tápanyagok hasznosítására, továbbá a rizoszféra kolonizációjára. Ezek a többségében talajban előforduló mikroorganizmusok sikerrel kerültek a kereskedelmi forgalomba annak érdekében, hogy növeljék a terméshozamot a növény növekedésének serkentése, és a széles körben elterjedt növénykórokozókkal (baktériumok, gombák, oomycoták) szembeni biológiai védekezés révén (Monte, 2001; Harman és mtsai., 2004a).
3.5. A Trichoderma fajok biokontroll mechanizmusai
A biokontroll ágensek élő szervezetek, melyek aktivitása jelentős mértékben függ a környezet fizikokémiai paramétereitől. Ebből az okból kifolyólag a Trichoderma törzsek által kifejtett biokontroll tevékenység sokszor megjósolhatatlan. Az egyes biokontroll
16 Trichoderma fajok törzsei közötti genetikai sokszínűség megismerése, valamint biokontroll mechanizmusaik alaposabb tanulmányozása révén a törzsek sokkal hatékonyabb alkalmazása valósulhat meg. A biokontroll mechanizmusok összetettek, a kórokozókkal szembeni védekezés a különböző mechanizmusok együttes hatása által valósulhat meg (Howell, 2003). Mindezek hátterében állhat a tápanyagért és élőhelyért folytatott kompetíció, és a Trichoderma által termelt másodlagos anyagcseretermékek, melyek gátolhatják a növénypatogén gombák spóráinak csírázását (fungisztázis). A biokontroll Trichoderma törzsek el is pusztíthatják a célszervezetet (antibiózis), vagy módosíthatják a rizoszférát azáltal, hogy savanyítják környezetük pH-ját, így megakadályozva a kórokozók növekedését. A biokontroll megvalósulhat a patogén és a BCA közötti direkt kapcsolat, azaz a mikoparazitizmus által, ami a fizikai kapcsolaton túl magába foglalja különféle hidrolítikus enzimek, továbbá toxikus vegyületek és antibiotikumok termelését, melyek együttesen is hathatnak a kórokozóra. A Trichoderma BCA-k pozitív hatást gyakorolnak a növényekre a növény növekedésének serkentése által (biofertilizáció), valamint a növények védekező rendszerének serkentése révén is (Harman és mtsai., 2004b).
3.5.1. Fungisztázis a talajban
A talaj fungisztatikus hatása (Lockwood, 1997), valamint az a képességünk, hogy erősítsük, vagy éppen gyengítsük a talajnak ezt a tulajdonságát (Papavizas és Lumsden, 1980), jelentős hatással lehet a természetes úton jelenlevő vagy a mesterségesen kijutatott Trichoderma fajok túlélésére és populációméretére (Papavizas, 1985). A Trichoderma konídiumok ellenállóak lehetnek ezzel a fungisztatikus hatással szemben (Mitchell és Dix, 1975), de vannak érzékenyek fajok is (Lockwood, 1997). Az érzékenység semleges vagy lúgos kémhatású talajokban jobban érvényesül (Danielson és Davey, 1973a).
A jó antagonista képességgel rendelkező Trichoderma törzsek képesek a talaj más mikroorganizmusok és növények metabolitjai által kialakított fungisztatikus hatásának kiküszöbölésére, így képesek ebben a kompetitív környezetben túlélni (Benítez és mtsai., 2004). A Trichoderma törzsek a talajba juttatást követően gyors növekedésre képesek, melyben jelentős szerepet játszik, hogy nagyfokú természetes ellenálló-képességgel rendelkeznek számos toxikus vegyülettel szemben. Ez a rezisztencia az ABC transzport- rendszerek működésének köszönhetően képes létrejönni (Vyas és Vyas, 1995). A toxikus vegyületek között különféle herbicidek, fungicidek és peszticidek egyaránt megtalálhatóak (Chet és mtsai., 1997).
17 3.5.2. Tápanyagokért folytatott verseny
A Trichoderma fajok közös jellemzője, hogy képesek más gombafajokat kiszorítani a rizoszférából, így mind az élőhelyért, mind pedig a tápanyagokért zajló verseny kitűnő kompetitorai (Harman és mtsai., 2004b). Azok a Trichoderma törzsek, amelyek hatékonyabb tápanyagfelvételt és gyorsabb növekedést mutatnak, jelentős előnyhöz juthatnak.
A talajban normális körülmények között a vas oldhatatlan állapotban van jelen, azonban a baktériumok és a gombák egy speciális rendszer segítségével kelátolják és felvehető formájúvá alakítják. Ez a rendszer magába foglalja a sziderofórok, valamint olyan peptidek termelését, amelyek ligandként és vastranszporterként működnek. Jalal és munkatársai (1986) T. virens esetében 3 hidroxamát sziderofórt azonosítottak, míg Wilhite és munkatársai (2001) elvégezték a T. virens peptid-szintetázt kódoló psy1 génjének delécióját, amelynek valószínűsíthetően a sziderofór-szintézisben van szerepe.
Eredményeik alapján azonban nem mutatkozott különbség a deléció hatására a P. ultimum- mal és R. solani-val szembeni biokontroll képességben.
Bár még nem jelenthető ki teljes bizonyossággal, hogy a vas felvételéért sziderofórok révén megvalósuló verseny a biokontroll mechanizmusok része, a rendelkezésre álló eredmények azonban erre utalnak. Három Trichoderma faj, a T. reesei, THSC és T. atroviride összehasonlító genomelemzésének eredményei alapján a Trichoderma nemzetség biokontroll képességekkel rendelkező képviselői kiterjedt, a kelátolt vas felvételében közreműködő sziderofór-transzporter rendszerekkel rendelkeznek (Kubicek és mtsai., 2011; Kredics és mtsai., személyes közlés).
3.5.3. Mikoparazitizmus
A talajra összetett biológiai élőhelyként tekinthetünk. Különösképpen igaz ez a rizoszférára, ahol jelentős számú biológiai kölcsönhatást figyelhetünk meg a baktériumok, gombák, nematódák és a növények között (Singh és mtsai., 2004). Minden egyes organizmus a túlélésért küzd ebben az erősen kompetitív környezetben. A Trichoderma fajok és más gombák között létrejövő direkt kapcsolatokat hagyományosan nekrotróf hiperparazitizmusnak vagy mikoparazitizmusnak nevezzük (Harman, 2011a), melynek során olyan antagonista kapcsolat jön létre két gombafaj között, ahol az egyik faj képviselője közvetlenül támadja a másik faj képviselőjét. A gombák közötti parazita kapcsolatokat két fő csoportba sorolhatjuk az alapján, hogy a parazita mekkora agresszivitást mutat a gazda felé. Az első csoportba a biotróf kapcsolatokat soroljuk,
18 melynek során a gazda tápanyaggal látja el a parazitát, mely nem okozza a gazdaszervezet pusztulását. A második nagy csoportot a nekrotróf kapcsolatok alkotják. Az ebbe a csoportba sorolható mikoparazita gombák szélesebb gazdakörrel és nagyobb fokú agresszivitással rendelkeznek, és végső soron a gazdaszervezet pusztulását okozzák. Mivel a mikoparazita Trichoderma fajok az elpusztult gombákból álló biomasszát is képesek tápanyagforrásként hasznosítani, ezért életmódjukat mikoparazita helyett mikotrófnak is nevezhetjük, mely elnevezés magába foglalja mind a biotróf, mind pedig a szaprotróf táplálkozási stratégiákat (Druzhinina és mtsai., 2011). A Trichoderma nemzetség azon képviselőit, melyekre nekrotróf mikoparazitizmus jellemző, igazoltan is hatékony biokontroll ágenseknek tekinthetjük (Herrera-Estrella és Chet, 2003).
3.5.3.1. A gazdagomba jelenlétének érzékelése
A (nekrotróf) mikoparazitizmushoz vezető út komplex folyamat, melynek első lépése a gazdagomba jelenlétének érzékelése. Ennek megismeréséhez 3 különböző Trichoderma faj genomszekvenálásából (Kubicek és mtsai., 2011), valamint transzkriptomikai analíziséből (Lorito és mtsai., 2010; Seidl és mtsai., 2009) származó adatok nyújtottak átfogó ismereteket. A különböző Trichoderma törzseknél a gazdagombával létesített tényleges kapcsolat kialakítása előtt megfigyelték számos proteázt, és oligopeptid transzporter fehérjét kódoló gén kifejeződését (Seidl és mtsai., 2009; Suárez és mtsai., 2007). Ezen proteázok többsége a T. cf. harzianum CECT 2413 törzs transzkriptómájáról biokontroll körülmények között szerzett adatok alapján a szubtilizin-szerű szerin proteázok csoportjába tartozott (Suárez és mtsai., 2007; Seidl és mtsai., 2009). Azok a T. atroviride törzsek, amelyek túltermelik a prb1 gén által kódolt proteázt, emelkedett mikoparazitikus aktivitással rendelkeznek (Flores és mtsai., 1997). Ezek a proteázok működésük során oligopeptideket szabadítanak fel a gazdagomba sejtfalából, melyek ezután a T. atroviride megfelelő receptoraihoz kötődve segítik a nitrogén hiányának érzékelését (Seidl és mtsai., 2009) (1. ábra). A G protein-kapcsolt receptorokat („G protein-coupled receptors”, GPCRs) tekintjük ezen oligopeptidek szenzorának. További G protein-kapcsolt receptorok is szerepet játszanak a gazdagomba érzékelésében. Ilyen például a Gpr1, amely a cAMP receptor-szerű GPCRs család tagja, mely szintén a T. atroviride mikoparazitizmusában játszik szerepet (Omann és mtsai., 2009).
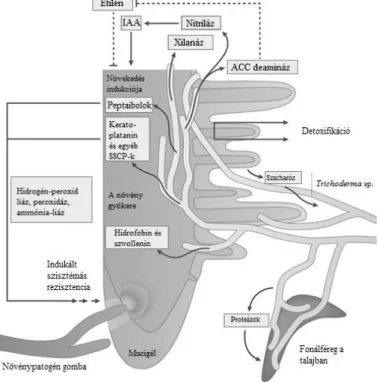
19 1. ábra. A Trichoderma fajok mikoparazitizmusa a talajban.
A Trichoderma fajok által termelt és szekretált proteázok hatására a gazdagombából különféle peptidek szabadulnak fel. Ezek a peptidek vagy a G protein-kapcsolt receptorokhoz (Gpr1) vagy a nitrogén-érzékelő receptorokhoz kötődnek, így elindítva egy kaszkádmechanizmust, mely a MAPK-okon keresztül egy eddig
ismeretlen transzkripciós faktort aktivál, ami aztán a sejtfal lebontásában szerepet játszó enzimek és másodlagos metabolitok termelődését váltja ki. A gazdagomba által termelt metabolitok és a reaktív oxigén
gyökök (ROS) kiváltják a Trichoderma törzsek stresszválaszát, valamint detoxifikációját. Druzhinina és mtsai. (2011) nyomán.
A gombákban megtalálható jelátviteli utak közül a legjelentősebb a mitogén- aktiválta protein kináz (MAPK) útvonal (Schmoll, 2008) (1. ábra). A Trichoderma fajok genomjában három MAPK-t kódoló gént találhatunk. Az egyik az úgynevezett patogenitásért felelős MAPK (TmkA; ismert még Tvk1 és Tmk1 néven). A következő a sejtintegritásért felelős kináz (TmkB), a harmadik pedig az ozmoregulációban játszik szerepet (Hog1) (Schmoll, 2008). A tmkA deléciója a T. virens ʽPʼ törzséből (gliovirin- termelő, Pythium fajokkal szemben hatásos biokontroll törzs) a Sclerotium rolfsii-val szemben mutatott antagonizmusának elvesztését eredményezte, ellenben a R. solani elleni antagonizmusra való képessége megmaradt (Mukherjee és mtsai., 2003; Viterbo és mtsai., 2005). Mindezekkel ellentétben a tmkA deléciója a ʽQʼ törzsben (nagy mennyiségben termel gliotoxint, R. solani-val szemben hatásos) tovább javította a törzs R. solani-val valamint, P. ultimum-mal szemben mutatott biokontroll képességét. Minderre a másodlagos metabolitprofilban jelen lévő különbség szolgáltathat magyarázatot. A másik két MAPK-ról (TmkB és a Hog1) sokkal kevesebb információ áll rendelkezésünkre, mivel a vizsgált mutánsok gyenge növekedést mutatnak, ami kizárja őket a sikeres
20 antagonizmusból. A TmkB mutáns T. virens mikoparazitizmusa sérült S. rolfsii-val szemben (Kumar és mtsai., 2010), míg a Hog1 mutáns T. atroviride nem mutatott mikoparazita aktivitást (Delgado-Jarana és mtsai., 2006).
3.5.3.2. A tényleges kapcsolat kialakulása a mikoparazita és a gazdagomba között A mikoparazitizmus megvalósulásához a gazdagomba hifájának felismerésén túl szükség van a direkt kapcsolat kialakulására is, amely az úgynevezett ʽcoilingʼ mechanizmus révén valósul meg. Ennek során a Trichoderma hifa jellegzetes hurokszerű növekedést mutat a gazdagomba micéliuma körül (1. ábra). Ez a jelenség a gazdagomba hifájának felszínén jelen lévő lektinek felismerésén alapul (Inbar és Chet, 1996). Azonban a növényi lektinek is ugyanolyan mértékben képesek a ʽcoilingʼ mechanizmust indukálni, ami arra utal, hogy nem a lektinek határozzák meg a folyamat gazdaspecificitását (Rocha- Ramirez és mtsai., 2002). Egyes Trichoderma törzsek gazdaszervezet jelenlétének hiányában képesek a saját hifáik körül is körbetekeredni, ami viszont arra utalhat, hogy nincs szoros összefüggés a ʽcoilingʼ és a mikoparazitizmus között.
A Trichoderma fajok mikoparazita támadása során a gazdagomba hifájának mentén történő növekedéssel párhuzamosan papillaszerű képletek kialakulását is megfigyelhetjük (1. ábra). A folyamat későbbi lépései során a gazdagomba sejtfalának lebontása és a penetráció is ezen papilla-szerű képlet kialakulásának helyén fog bekövetkezni (Harman, 2011b; Harman és mtsai., 2004a; Chacón és mtsai., 2007). Ezek a képletek analógok a növénypatogén gombák appresszóriumaival.
3.5.3.3. A mikoparazitizmus végső lépése
A mikoparazitizmus legvégső lépésében a Trichoderma törzsek gombaellenes másodlagos anyagcseretermékeik (antibiózis) és sejtfalbontó enzimek együttes hatása révén a gazda gombák pusztulását okozzák (1. ábra). A penetráció után a Trichoderma hifa intracellulárisan növekedik a gazdagomba hifájának belsejében, és tápanyagként hasznosítja annak beltartalmát.
3.5.4. Sejtfalbontó enzimek és szabályozásuk
A mikoparazitizmus során a felismerés, majd a Trichoderma hifa azt követő morfológiai változásai után a hidrolítikus enzimek termelése kulcsfontosságú szerepet töltenek be a sikeres mikoparazita kölcsönhatás létesítésében, a gombasejtfal jelenti ugyanis az első akadályt a gazdagomba és a mikoparazita között. A sejtfal lebontásában számos enzim (kitinázok, glukanázok, N-acetilglükózaminidázok, proteázok) együttesen
21 játszik szerepet (Di Pietro és mtsai., 1993; Schirmböck és mtsai., 1994; Mishra, 2010).
Mindezeket alátámasztandó, különféle kísérleti körülmények között vizsgálták Trichoderma törzsek rázatott tenyészeteit. Amikor a törzsek növekedéséhez szükséges szén-, illetve nitrogénforrások jelen voltak a tápközegben, akkor a litíkus enzimek kifejeződéséért felelős gének represszált, nyugalmi állapotban voltak (Mach és mtsai., 1999; Olmedo-Monfil és mtsai., 2002). Mivel a hidrolítikus enzimek specifikus szerepet töltenek be a mikoparazitizmus során csakúgy, mint a lehetséges gazdagomba felismerésében, így két különböző Trichoderma törzs együttes leoltásával vizsgálták az enzimeket kódoló gének kifejeződését. A tápanyag mennyiségének csökkenése által bekövetkezett korlátozó körülmények sem eredményezték azonban az enzimszint növekedését (Reithner és mtsai., 2011). Másrészről tápanyaglimitáló körülmények között a gazdagomba sejtfalkomponenseinek tápközeghez történő hozzáadása a sejtfalbontó enzimeket kódoló gének kifejeződésének fokozását idézte elő (Olmedo-Monfil és mtsai., 2002; Suárez és mtsai., 2007).
3.5.4.1. Kitinázok
Bár a gazdagomba sejtfalának lebontása több litíkus enzim együttes hatása révén valósul meg, a mikoparazitizmusban a kitinázok játszák a kulcsszerepet, a legtöbb gomba sejtfalában ugyanis a legnagyobb arányban kitint találunk (Baek és mtsai., 1999; Peberdy, 1990). A kitinolítikus enzimeket három fő csoportba sorolhatjuk (Harman és mtsai., 1993;
Sahai és Manocha, 1993). Az első csoport képviselői közé tartoznak az endokitinázok (EC 3.2.1.14), amelyek véletlenszerűen hasítják a kitinláncokat kitotetraóz, kitotrióz és kitobióz egységekre. A második csoportba az exokitinázok – vagy kitobiozidázok (EC 3.2.1.29) – tartoznak, melyek kitobióz-egységeket hasítanak le a kitinláncról anélkül, hogy egyéb mono-, vagy oligoszacharid képződne. Az utolsó csoportot az N-acetil-β-D- glükózaminidázok – más néven kitobiázok (EC 3.2.1.30) – alkotják, melyek monomereket hasítanak le a kitin, illetve a kitin oligomerek nem redukáló végeiről.
3.5.4.2. Glukanázok
A kitinázok mellett igen fontos szerepet játszanak a mikoparazitizmusban a β-1,3- glükanázok (laminarinázok) (Harman és mtsai., 2004a) is, melyek két fő csoportra oszthatók (Manczinger és mtsai., 2002). Az endo-β-1,3-glükanázok (EC 3.2.1.29) a β- kötéseket random helyeken hasítják a poliszacharidlánc mentén, ezáltal rövid oligoszacharidokat szabadítanak fel. Ezzel szemben az exo-β-1,3-glükanázok (EC
22 3.2.1.58) glükóz egységeket hasítanak le a szubsztrát nem redukáló láncvégéről. Az első Trichoderma β-1,3-glukanáz enzim tisztítását Tangarone és munkatársai (1989) hajtották végre egy T. longibrachiatum törzsből. Később Vázquez-Garciduenas és munkatársai (1998) egy THSC törzs β-1,3-glukanáz rendszerének 7 tagját azonosították laminarin- indukció révén.
3.5.4.3. Cellulázok
Mivel a Trichoderma fajok többsége szaprotróf életmódot folytat, így részt vesznek a cellulóz, hemicellulóz és kitin biopolimerek lebontásában is (Schuster és Schmoll, 2010).
A Trichoderma fajok celluláz (EC 3.2.1.4.) termelését ezért elsősorban ipari jelentősége miatt tanulmányozzák nagyon széles körben (Kubicek és mtsai., 1990). Amíg a cellulóz β- 1,4 kötéssel kapcsolódó glükóz egységekből épül fel, addig a hemicellulóz egy heteropolimer, amely különböző pentózokból és hexózokból épül fel. Ezek gyakran acetilálva is vannak és elágazásokat is tartalmaznak (Do Vale és mtsai., 2014). A cellulóz és hemicellulóz lebontásához számos extracelluláris enzim együttes hatására van szükség.
A Trichoderma fajok rendelkeznek a lebontáshoz szükséges összes enzimmel. A legjelentősebb celluláztermelő fajuk a T. reesei, amely rendkívüli gazdasági jelentőséggel rendelkezik. Az általa termelt cellulázokat felhasználják az élelmiszeriparban (Galante és mtsai., 1998b), a textiliparban (Galante és mtsai., 1998a), papírgyártás során (Buchert és mtsai., 1998), valamint bioüzemanyag gyártásához is (Seiboth és mtsai., 2011). Mindezek mellett egyes jó celluláztermelő képességgel rendelkező Trichoderma törzsek az aratást követően a talajba történő kijuttatásuk esetén segíthetik a növényi szármaradványok lebomlását is Khan és Amin (2012).
A Trichoderma fajok cellulázai az Oomycota törzsbe tartozó Pythium fajokkal szembeni mikoparazitizmus során is fontos szerepet játszanak a gazdagomba hifájába történő penetrációban (Benhamou és Chet, 1997).
3.5.4.4. Proteázok
Egyéb sejtfalbontó enzimek kisebb polimereket és fehérjéket hidrolizálnak, melyek végső soron hozzájárulnak a gazdagomba sejtfalának teljes széteséséhez. Flores és munkatársai (1997) egy THSC törzs biokontroll aktivitásának emelkedését mutatták ki egy proteináz fehérje túltermeltetése következtében, így igazolva a proteázok szerepét a gazdagomba sejtfalában található fehérjekomponensek lebontásában Szekeres és mtsai., 2004).
23 3.5.5. Antibiózis, a másodlagos anyagcseretermékek szerepe a biológiai védekezésben
A másodlagos anyagcseretermékek („secondary metabolites”, SM) kisméretű szerves molekulák, melyek jelenléte nem létszükséglet az organizmus növekedéséhez, fejlődéséhez és szaporodásához. Specifikus körülmények között azonban fontos szerepet töltenek be különféle jelátviteli utakban és szükségesek az egyéb mikroorganizmusokkal való kapcsolatok kialakításához. Ennél fogva a biokontroll folyamatok során betöltött szerepük vizsgálata nagy jelentőséggel bír. Ismeretes, hogy a Trichoderma fajok számos kis molekulatömegű, apoláros másodlagos metabolitot (pl. pironokat, terpenoidokat, szteroidokat és poliketideket) képesek termelni. A Trichoderma fajok képesek sziderofórok termelésére is, és nagy számban különféle peptaibolt is előállítanak, amelyek rendhagyó aminosavakban gazdag, nem riboszómális fehérjeszintézis útján szintetizálódó metabolitok (Degenkolb és mtsai., 2006).
3.6. A lakkáz enzim előfordulása, hasznosítási lehetőségei
A lakkázok (EC 1.10.3.2, benzéndiol: oxigén oxido-reduktázok) polifenol- oxidázok, melyeket ’multicopper oxidase’-nak is neveznek, mivel 4 rézatomot tartalmaznak. A lakkázok széles körben elterjedtek a magasabbrendű növényekben, valamint a rovarok és baktériumok körében is, de különféle gombák által termelt lakkázokat is azonosítottak (Gianfreda és mtsai., 1999). Az eddig legtöbbet vizsgált, gombák által termelt lakkázokat a fehérkorhasztó gombákból tisztították (Kiiskinen és mtsai., 2004). A gomba-lakkázok szerepet játszhatnak a növényekkel szembeni patogenitásban, a pigmenttermelésben és a lignocellulóz lebontásában (Thurston, 1994;
Gianfreda és mtsai., 1999).
A lakkázok számos szubsztrátot képesek hasznosítani, ezért a különböző iparágak nagy érdeklődést mutatnak irántuk. Alkalmasak lehetnek a textilfestékek fehérítésére, a papírgyártás során pedig a lignocellulóz lebontására. Felhasználhatók továbbá bioszenzorokban, detoxifikációs és bioremediációs folyamatokban is (Gianfreda és mtsai., 1999; Xu, 1999)
3.6.1. A Trichoderma fajok lakkáztermelése
Kutatások bizonyították a Trichoderma fajok (T. atroviride, THSC) lakkáztermelését, melyet a zöld pigment termelésével hoztak összefüggésbe (Hölker és mtsai., 2002; Sadhasivam és mtsai., 2008; Chakroun és mtsai., 2010). Két talajmintákból származó törzs, egy T. viride és egy T. reesei esetében is kimutatták a lakkáztermelést, a
24 két törzs molekuláris azonosítása azonban még nem történt meg (Gochev és Krastanov, 2007; Krastanov és mtsai., 2007). Mindezen munkák ellenére is csak igen kevés tanulmány foglalkozik a Trichoderma törzsek által termelt lakkázokkal.
3.7. A különféle fungicidek és azok csoportosítása hatásmechanizmusuk alapján A növénypatogén gombákkal szembeni lehetséges védekezési stratégiák közé tartozik a kémiai védekezés, ahogy ez már a 3.3.2.-es fejezetben is említésre került. A kémiai védekezés különböző fungicidek révén valósítható meg. A fungicidekkel történő védekezés kulcsfontosságú szerepet játszik a különféle növénykórokozókkal szembeni integrált növényvédelemben. Használatuk során a rezisztencia megjelenése fontos korlátozó tényezővé vált a fungicidek hatékonyságát és használhatóságának időtartamát figyelembevéve, így az újabb és újabb fungicidek kifejlesztése egyre magasabb költségekkel jár (Ma és Michailides, 2005). Megoldást nyújthat azonban az egyes biokontroll ágensek és a csökkentett mennyiségű fungicidek együttes alkalmazása az integrált növényvédelem keretein belül (Monte, 2001).
A különböző fungicideket hatásmechanizmusuk alapján csoportosíthatjuk, így megkülönböztethetünk kontakthatású és szisztémás hatású fungicideket.
A kontakthatású szerek közé tartoznak a réz-, és kénvegyületek, valamint a ditiokarbamát származékok (mankozeb, maneb, tirám), továbbá különböző ftálimid- származékok is, mint pl. a folpet és a kaptán, melyek a sejtlégzés gátlására képesek (Siegel, 1971).
A szisztémás hatású fungicidek közé soroljuk a dikarboximid fungicideket, például az iprodiont, amely a gombák DNS-szintézisének gátlásán keresztül fejtik ki hatásukat (Pappas és Fisher, 1979).
Egy másik, nagy csoportot alkotnak a benzimidazolok, melyek közé sorolható a benomil, a karbendazim és a tiofanát-metil. Hatásmechanizmusukról megállapították, hogy a tubulin szintézisének gátlására képesek (Davidse, 1973).
Újabb nagy csoportot alkotnak a szterol-bioszintézis gátlói, ezeken belül megkülönböztetjük a szterol-C14-demetiláz inhibitorokat, melyeket az angol elnevezésük alapján DMI-fungicideknek neveznek. Legfőbb képviselőik az azolok, mint a triazol, imidazol és a pirimidin fungicidek (Russell, 1995). A különféle morfolinszármazékokkal (spiroxamin) a ∆8-∆7 izomeráz és a ∆14 reduktáz gátlása lehetséges (Russell, 1995).
25 3.8. A növények növekedésének és fejlődésének serkentése és ennek mechanizmusai
Évekkel ezelőtt még tartotta magát az a nézet, miszerint a Trichoderma fajok növénynövekedést serkentő hatásukat a csekély veszélyt jelentő növénypatogén gombák elpusztításán keresztül érik el. Ez a felvetés azt sugallta, hogy a nem steril környezetben lévő növények mindegyike szenved valamilyen mértékű fertőzéstől, így maximális növekedési potenciáljukat csak abban az esetben tudják elérni, ha a Trichoderma elnyomja a növénypatogén gomba által okozott negatív hatást, ezáltal növénynövekedést serkentő hatást kifejtve (Stewart és Hill, 2014). A kórokozók gátlása azonban nem lehet az egyedüli magyarázat, mivel a Trichoderma fajok hatása steril illetve félsteril körülmények között, hátrányos talajmikroflóra nélkül is érvényesül. Azóta számos egyéb mechanizmus szerepe került előtérbe a Trichoderma fajok növénynövekedést serkentő hatása kapcsán, többek között a növényi hormonok szintézise, vitaminok termelése, a talajban található tápanyagok szolubilizálása, a gyökérzet fejlődésének erősítése, valamint a fotoszintézis és a növények védekező rendszerének fokozása (Harman, 2000, 2006; Harman és mtsai., 2004a; Inbar és mtsai., 1994). Valószínű, hogy az egyes Trichoderma törzsek ezen tulajdonságok közül egy, vagy akár több mechanizmus együttműködése révén fejtik ki pozitív hatásukat. A THSC T-22 irányította növénynövekedés-serkentést megfigyelték pl.
üvegházi, valamint szabadföldi kísérletek során is, melynek hátterében a már említett tulajdonságok állhatnak. Fontos mechanizmus a gyökerek gyorsabb fejlődésének elősegítése is, ami növeli a növények szárazsággal szembeni ellenállóképességét (Altomare és mtsai., 2000; Harman, 1999; Harman és mtsai., 2004b, Mastouri és mtsai., 2010).
3.8.1. Rizoszféra-kompetencia
A növények növekedésének serkentése kapcsán meg kell említeni a Trichoderma törzsek rizoszféra-kompetenciáját. A rizoszféra, mint élőhely általánosnak tekinthető a Trichoderma törzsek esetében, ezen az élőhelyen lehetőségük van mind biotróf, mind szaprotróf életmód kialakítására. Mindezeket alátámasztja az a vizsgálat, melynek során nagyon nagy számban és változatosságban izoláltak Trichoderma törzseket egy etiópiai kávéültetvény kávénövényeinek (Coffea arabica) rizoszférájából (Mulaw és mtsai., 2010).
Ezzel ellentétben, egy olaszországi tanulmány során a nem rizoszféra-jellegű talajminták vizsgálatakor kevés Trichoderma fajt izoláltak (Migheli és mtsai., 2009).
A Trichoderma fajok rizoszférához mutatott affinitását két okkal is magyarázhatjuk. Az első lehetséges magyarázat, hogy a szárazföldi növények 92%-ának
26 gyökerén megtalálhatóak a mikorrhizaalkotó gombák, melyek potenciális gazdaszervezetei a mikoparazita Trichoderma törzseknek. A mikorrhizaalkotó gombák és a Trichoderma fajok között létrejövő kapcsolat azonban még mindig kevéssé tanulmányozott (Calvet és mtsai., 1993; Datnoff és mtsai., 1995; McAllister és mtsai., 1994; Nemec és mtsai., 1996;
Siddiqui és mtsai., 1996; Green és mtsai., 1999). Egyes tanulmányok szerint szinergista hatás figyelhető meg a két gomba között (Calvet és mtsai., 1993; Datnoff és mtsai., 1995;
Nemec és mtsai., 1996), míg mások megfigyelései alapján a Trichoderma fajok elnyomják az arbuszkuláris mikorrhizát képző gombákat (McAllister és mtsai., 1994; Siddiqui és mtsai., 1996; Green és mtsai., 1999).
2. ábra. A Trichoderma fajok és egyéb szervezetek között létrejövő kapcsolatok.
A Trichoderma fajokból számos vegyület szabadul fel, melyek képesek indukálni a növények szisztémás rezisztenciáját (peptaibolok és a kerato-platanin), így végső soron különféle enzimek (hidrogén-peroxid liáz, peroxidáz, ammónia-liáz) szintjének emelkedéséhez vezetnek. A xilanáz (Eix) feltehetően mikróba-asszociált
molekuláris mintázatként („microbe-associated molecular pattern”, MAMP) viselkedik, ezáltal erősíti a növények védekező rendszerét. Az ACC deamináz gátolja a növényben az etilén képződését, ami a gyökér növekedéséhez fog vezetni. Az indolecetsav (IES) képződéséhez konstitutív nitriláz-szekrécióra van szükség.
A Trichoderma törzsek gyökéren történő megtapadásában segít a szwollenin és a hidrofobin. A Trichoderma törzsek szénforrásként hasznosítják a növények által termelt szacharózt. Nematofág hatásukat a szekretált kitináznak és a szubtilizin-szerű S8 proteáznak köszönhetik (SSCP). Druzhinina és mtsai. (2011) nyomán.
A második magyarázat a növények gyökerén megtalálható gyökérsüveg legkülső sejtrétege által kiválasztott gélszerű kapszula, az ún. mucigél, melynek fő alkotója a pektin és hemicellulóz (ramnogalakturonán és arabinoxilán) (2. ábra). Ezek a komponensek könnyen lebonthatóak a Trichoderma hemicellulázok által, melyek azért fejlődhettek ki,
27 hogy lehetővé tegyék a gazdagombák által lebontott növényi részekből felszabaduló poliszacharidok lebontását (Druzhinina és mtsai., 2011).
A növények által a rizoszférába kiválasztott mono-, és diszacharidok fontos szénforrást jelentenek a mikorrhizaalkotó gombák számára (Nehls és mtsai., 2010), melyek közül a szacharóznak további szerepe van a rizoszféra T. virens általi kolonizációjában (Vargas és mtsai., 2009) (2. ábra). A T. virens ugyanis specifikus szacharóz- transzporterrel rendelkezik, mely a gyökér kolonizációjának kezdetén indukálódik, és nagyfokú hasonlóságot mutat a növények szacharóz-transzportereivel (Vargas és mtsai., 2011). Mindez arra utal, hogy a szacharóz aktívan juthat el a növénytől a gombáig.
3.8.2. A gyökér kolonizációja
Egyes Trichoderma törzsek a gyökér kolonizációjára is képesek, melynek során a gyökér epidermiszén átjutva intracelluláris növekedést mutatnak. A gyökér epidermisze, a kortex, illetve az edénynyalábok ugyanakkor változatlanok maradnak. A gyökér kolonizációja kallóz és cellulóz felhalmozódása révén a környező sejtekben a sejtfal megvastagodását váltja ki. Mindemellett fenolos vegyületek is termelődnek, amelyek peroxidáz-irányította keresztkötések révén még nagyobb stabilitást eredményeznek a sejtfalban. A sejtfal ily módon történő módosítása meggátolja, hogy a gomba a gyökér mélyebb sejtrétegeibe is eljusson (Yedidia és mtsai., 1999).
A gyökér kolonizációja során a Trichodermá-nak együtt kell működnie a növény védekező rendszerével, mely különféle antimikrobiális vegyületeket (fitoalexineket) termel. A sikeres Trichoderma-növény kapcsolat maga után vonja a gomba detoxifikáló és védekező rendszerének aktiválását (2. ábra). Ebből adódóan a gombának hatásos rendszerrel kell rendelkeznie, hogy az őt károsító vegyületeket el tudja távolítani. Ruocco és munkatársai (2009) T. atroviride-ben azonosították a TAABC2 ABC-transzportert, mely különböző környezeti feltételek mellett is segítette a sikeres kolonizációt.
3.8.3. Az ásványi anyagok szolubilizálása, tápanyagfelvétel növelése
A talaj tápanyagtartalma fontos tényező, mely jelentős hatással van a növénynövekedést serkentő gombák növekedésére és aktivitására. Ennek a serkentő hatásnak a mértéke a tápanyagban szegény talajok esetében rendelkezik a legnagyobb jelentőséggel. A nitrogénről tudjuk, hogy nélkülözhetetlen a növények fejlődéséhez és a maximális terméshozam eléréséhez. Harman és Björkman (1998) THSC T22 törzsével történő kukoricakezelést követően megállapították, hogy a kezelt növények nagyobbak és