DOKTORI ÉRTEKEZÉS
A VÁNDORKAGYLÓ (DREISSENA POLYMORPHA) MEGTELEPEDÉSE, MENNYISÉGI VISZONYAI, ANYAGFORGALOMBAN BETÖLTÖTT SZEREPE, FILTRÁCIÓJA, VALAMINT BAKTÉRIUMKÖZÖSSÉGRE
GYAKOROLT HATÁSA
Készítette:
Balogh Csilla
okl. környezetmérnök
Pannon Egyetem, Környezettudományi Doktori Iskola, Veszprém
Belső konzulens: Dr. Padisák Judit tanszékvezető egyetemi tanár, az MTA Doktora
Pannon Egyetem, Mérnöki Kar, Limnológia Tanszék, Veszprém
Témavezető: Dr. B. Muskó Ilona
tudományos főmunkatárs, a biológiai tudomány kandidátusa
Készült: MTA Balatoni Limnológiai Kutatóintézet, Tihany
2008
A VÁNDORKAGYLÓ (DREISSENA POLYMORPHA) MEGTELEPEDÉSE, MENNYISÉGI VISZONYAI, ANYAGFORGALOMBAN BETÖLTÖTT SZEREPE, FILTRÁCIÓJA, VALAMINT BAKTÉRIUMKÖZÖSSÉGRE GYAKOROLT HATÁSA
Értekezés doktori (PhD) fokozat elnyerése érdekében Írta:
Balogh Csilla
Készült a Pannon Egyetem Környezettudományi doktori iskolája keretében Témavezetők: …Dr. Padisák Judit és Dr. B. Muskó Ilona…
Elfogadásra javaslom (igen / nem)
(aláírás) A jelölt a doktori szigorlaton …...%-ot ért el,
Az értekezést bírálóként elfogadásra javaslom:
Bíráló neve: …Dr. Korponai János… igen /nem
……….
(aláírás) Bíráló neve: …Dr. Nagy Sándor Alex… igen /nem
……….
(aláírás)
A jelölt az értekezés nyilvános vitáján …...% - ot ért el
Veszprém, ……….
a Bíráló Bizottság elnöke A doktori (PhD) oklevél minősítése…...
………
Az EDT elnöke
Tartalomjegyzék
Kivonat 1
Abstract 2
Auszug 3
Bevezetés 5
1. Irodalmi összefoglaló 7
1.1. A vándorkagylóelterjedése, mennyisége 7
1.1.1. Rendszertani besorolása, inváziója 7
1.1.2. Megjelenése a Balatonban, korábbi kutatási eredmények 11 1.1.3. A Balaton, mint a vándorkagyló élőhelye 14
1.2. A vándorkagyló megtelepedése, fejlődése, életciklusa 16 1.3. A vándorkagyló ökoszisztémára gyakorolt hatása 18
1.3.1. Kompetíciója más bevonatlakókkal 18
1.3.2. Filtrációja, fécesz és pszeudofécesz kibocsátása 21 1.3.3. Mikrobiális közösségre gyakorolt hatás 28
1.4. Kutatásaink előzményei 29
2. Célkitűzés 31
3. Anyag és módszer 32
3.1. A vándorkagyló megtelepedésének tanulmányozása 32 3.2. A vándorkagyló mennyiségi viszonyai 33
3.2.1. Hínáron 33
3.2.2. Kövön 34
3.2.2. Nádon 36
3.3. A terepi mintavételek részletei és az adatok feldolgozása 37
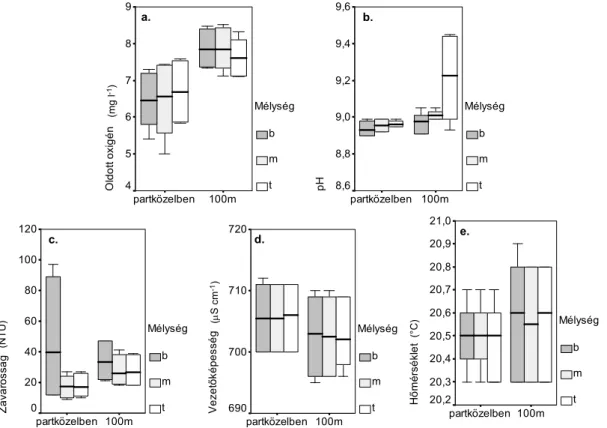
3.3.1. A mért környezeti paraméterek 37
3.3.2. A kagylóra vonatkozó paraméterek kalkulálása,
populációösszetétel vizsgálata 37
3.3.3. A használt statisztikai módszerek bemutatása 38
3.4. A vándorkagyló filtrációja a Balatonban 38 3.5. A vándorkagyló szerepe az anyagforgalomban 42 3.6. A vándorkagyló bakteriális közösségre gyakorolt hatása 44
3.6.1. Heterotróf baktériumtörzsek tenyésztése agaron 45
3.6.2. DNS izolálása és PCR 46
3.6.3. A közösség szerkezetének vizsgálata denaturális
gélelektroforézissel 47
3.6.4. A közösség anyagcseréjének nyomonkövetése
közösségi szintű fiziológiai profillal 47
3.6.5. Az üledék CHNS tartalma 49
4. Eredmények 50
4.1. A vándorkagyló megtelepedése térben és időben
és az ezt befolyásoló tényezők és környezeti paraméterek 50 4.2. A vándorkagyló mennyiségi viszonyai 57
4.2.1. 2001 és 2002-ben a balatoni hínárosban 57 4.2.2. 2003-2005-ben a Balaton köves parti zónájában 62 4.2.3. 2006-ban a Balatont szegélyező nádasban 68
4.3. A vándorkagyló szűrése a Balatonban 72 4.4. A vándorkagyló szerepe az anyagforgalomban 75 4.5. A vándorkagyló bakteriális közösségre gyakorolt hatása 82
4.5.1. A heterotróf kolóniaképző egységek (CFU)
számának elemzése lemezen 82
4.5.2. Mikrobiális aktivitás 85
4.5.3. A tápanyag együttes hasznosítása 87
4.5.4. A üledék CHNS tartalma 88
5. Eredmények megbeszélése, következtetések 90
5.1. A vándorkagyló megtelepedése térben és időben
és az ezt befolyásoló tényezők és környezeti paraméterek 90 5.2. A vándorkagyló mennyiségi viszonyai 95
5.2.1. 2001 és 2002-ben a balatoni hínárosban 95 5.2.2. 2003-2005-ben a Balaton köves parti zónájában 96 5.2.3. 2006-ban a Balatont szegélyező nádasban 99
5.3. A vándorkagyló szűrése a Balatonban 99 5.4. A vándorkagyló szerepe az anyagforgalomban 103 5.5. A vándorkagyló bakteriális közösségre gyakorolt hatása 106
6. Összefoglalás 110
7. Irodalomjegyzék 112
8. A doktori tanulmányok során elfogadott,
megjelent közlemények 129
8.1. Egyéb kiadványfüzetben megjelent,
elektronikusan is hozzáférhető közlemények 130
8.2. Benyújtott közlemények 130
9. Tézisek 131
10. Theses 134
Köszönetnyilvánítás 137
Kivonat
A vándorkagyló (Dreissena polymorpha) több kontinens édesvizeiben is gyorsan kolonizáló, széleskörű alkalmazkodóképességgel bíró faj, mely az adott ökoszisztéma összetételét, működését jelentősen befolyásolja.
Munkám során vizsgáltam azon élettelen és élő környezeti tényezőket (víz átlátszóság, mélység, vezetőképesség, pH, hőmérséklet, társulás összetétel) melyek hatással vannak a balatoni vándorkagyló térbeli és időbeli megtelepedésére, mennyiségi gyarapodására, olyan élőhelyeken, mint a hínáros, a nádas és a part menti köves zóna. Laboratóriumi körülmények között, inverz mikroszkóp, illetve CHNS-O elemanalizáló készülék segítségével tanulmányoztam a vándorkagyló táplálkozási (filtációs) dinamikáját, az élőhely (üledék) anyagforgalmában és a táplálékforrás (algafajok) lokális összetételének módosításában betöltött szerepét.
Továbbá lehetőségem adódott az észak-amerikai Nagy Tavakban élő vándorkagyló populációk az élőhely baktérium közösségének összetételére (PCR és DGGE elektroforézis), mennyiségére (CFU szám meghatározás), aktivitására és anyagcsere mintázatára (BIOLOG identifikációs teszt) gyakorolt hatásának felmérésére.
A Balatonban a vándorkagyló megtelepedésének dinamikája függött az aljzat minőségétől, a víz mélységtől és a parttól mért távolságtól. A vándorkagyló és a tegzes bolharák (Chelicorophium curvispinum) között a kolonizálható felületekért és a táplálékért folyó versengést állapítottunk meg, mely kapcsolat időben a vándorkagyló kezdeti inváziójának fokozatos csökkenésével, és ezzel párhuzamosan a tegzes bolharák dominanciájának növekedésével volt jellemezhető. A vándorkagyló tisztítási rátája 6-72 ml kagyló-1 óra-1 között változott. A Balatonban megtelepedett kagylóállomány - melynek össztömege elérheti a 88 tonnát – filtrációs kapacitását 1,7 millió m3 víz nap-1-ra becsültük, mely a teljes víztömeg megközelítőleg 0,1 %-át teszi ki. A vándorkagyló kibocsátott anyagcseretermékei szénnel és nitrogénnel terhelték a víztestet és elősegítették r-stratégista algafajok elszaporodását. A kagyló méretével együtt nőtt a kagyló filtrációs rátája és egyúttal a kibocsátott anyagcseretermék mennyisége, valamint a kagyló lágyszövetét alkotó szén, nitrogén és kén tartalom is. A vándorkagyló kolónia befolyásolta az élőhely üledék összetételét, ezzel kedvező körülményeket biztosított egyes baktériumfajoknak, valamint módosította a fenéküledék élőközösségének szerkezetét, aktivitását és anyagcsere diverzitását.
Eredményeink alapján a vándorkagyló a Balaton parti övének domináns tagja. Dinamikus filtrációja következtében befolyásolja az üledék szén és nitrogéntartalmát, az élőhely közösség alga és baktérium összetételét, ezáltal döntően képes meghatározni a tavak ökológiai állapotát.
Abstract
SETTLING, QUANTITATIVE TRENDS, FILTRATION OF ZEBRA MUSSEL (DREISSENA POLYMORPHA) AND THE EFFECT OF THE MUSSEL ON THE MATTER-ENERGY FLUX
AND ON THE BACTERIAL COMMUNITY
The present study dealt with studying the conditions of settling and the impact on the surrounding of the rapidly colonizing invasive species, the zebra mussel (Dreissena polymorpha).
The environmental parameters (water transparency, depth, conductivity, pH and temperature) influencing the spatio-temporal pattern of colonization on submerged macrophytes, reed, and stones of the littoral zone of Lake Balaton were studied in situ. In laboratory conditions, the filtration rate and clearance potential, the contribution in matter-energy flux and on the composition of the phytoplankton community were investigated by microscopic follow up and a CHNS-O analyzer. In North American lakes the impact of the mussel on the bacterial community structure, metabolic activity and metabolic diversity was studied by determination the colony forming units (CFU), applying BIOLOG assay, PCR technique, and denaturing gradient gel electrophoresis (DGEE).
The settling dynamic of zebra mussel depended on the surface of the substrate, the water depth and the distance from the shoreline. The density on exposed subtrata increased by 18,643 ind m-2 d-1.A competition for the settling place and food was detected between Dreissena and the amphipod Chelicorophium curvispinum. After the initial invasion the relative abundance of zebra mussels decreased and finally Chelicorophium curvispinum became dominant species on the exposed substrata.
However, in most of the sampling zone the permanent dominance of zebra mussel was observed especially in its spawing period. They were also found to be the first and most successful re-colonizing species when dry period was over. Filtration rate of zebra mussel ranged between 6-72 ml ind-1 h-1. It could be estimated that the assumed 88 tons Dreissena of the lake are able to filter out algae from 1,7 million m3 water day-1 which volume means the 0,1% of Lake Balaton. Excretes of the mussel increased carbon and nitrogen content of the sediment and therefore provide an excellent nutrient for the r-strategist algae species. The carbon, nitrogen and sulfur concentrations measured from mussel tissue as well the filtration rate increased together with the body size. Dreissena clusters provided better condition for certain bacteria, hence could modify the structure, activity and metabolic diversity of the benthic bacterial community.
Zusammenfassung
DIE ANSIEDLUNG, DIE QUANTITATIVE VERHÄLTNISSE, DIE FILTRIERUNG UND DIE ROLLE IN DEM STOFFWECHSEL BZW. AUF DIE BAKTERIELLE GEMEINSCHAFT DER
WANDERMUSCHEL (DREISSENA POLYMORPHA)
Der Schwerpunkt unser Studien war die Untersuchung der Ansiedlung, der quantitative Festlegung, der Rolle in dem Stoffwechsel und des Einflusses auf die bakterielle Gemeinschaft der Wandermuschel (Dreissena polymorpha).
Parallel zu den Muschel-Probeentnahmen wurden auch wasserchemische Messungen durchgefürt (Bestimmung der Sichttiefe mit der Secchi-Scheibe, Wassertiefe, Leitfähigkeit, pH, Temperatur). Wir haben die Frage gestellt, welche Faktoren die Ansiedlung der Wandermuschel und die räumliche und zeitliche Wandlung der Ansiedlung beeinflussen und bestimmen. Zur Bestimmung der Muscheldichte wurden Proben von der Wasserpest, vom Steinufer und vom Schilfdickicht in dem Plattensee genommen. Die Individuendichte der Algen wurden mit der Hilfe von Stereomikroskop und der Inhalt des Kohlenstoffs, des Stickstoffs und des Schwefels von Proben wurden mit dem CHNS-O Apparat bestimmt. Ihre Filtrierung wurde untersucht, ihre Filtrationsrate und ihre Wasserreinigung-Potenzial kalkuliert und ihre Rolle im Stoffwechsel des Sediment bzw. in der Modifikation der Artenzusammensetzung von Algen bestimmt. Zu Untersuchungen von Bakterien wurden molekularbiologische und mikrobiologische Methoden verwendet.
In dem Kolonisation-Experiment haben wir die Kompetition zwischen Dreissena polymorpha und Chelicorophium curvispinum untersucht. Am Ende des Experiments haben wir bemerkt, dass Chelicorophium curvispinum die dominante Art war. Im Plattensee ist deshalb dem Ergebnis nach, vom Anfang der Invasion auch Chelicorophium curvispinum an verschiedenen Oberflächen (Schilf, Stein, Wasserpest) dominant. Im Jahre 2004, in den Dürreperioden, wurde die Muschel ganz zurückgedrängt. Später wurde es bemerkt, dass die Muschel an der wasserbedeckten neuen Oberflächen parallel mit der Steigung der Wasserhöhe weit verbreitet war. Die Filtrationsrate der Wandermuschel schwankte zwischen 6-72 ml Muschel-1 Stunde-1. Die in dem Plattensee sich verbreitende Muschelpopulation, -Gesamtgewicht 88 Tonnen- kann täglich 1,7 Millionen m3 Wasser von den Algen reinigen. Diese Wassermenge ist 0,1 Prozent der Gesamtwassermenge des Plattensees.
In unseren Experimenten haben die r-Strategen der Algenarten an den durch die Muschel entleerenden Substanzen regelmäßig sich vermehrt. Zusammen mit der Gröβe der Wandermuschel ist ihre
Filtrationsrate und zugleich die Menge der entleerenden Stoffwechselprodukten, bzw. der Kohlenstoff, der Stickstoff und der Schwefel in dem Muschelgewebe gewachsen. Die Dreissena Kolonie wirkt auf die in der Nähe vorgehende Vorgängen ein, unter anderem auf die Umgebung des Sediment. So macht es möglich für bestimmte Bakterienpopulationen dominant zu sein, und gleichzeitig ändert es die Struktur, die Aktivität, den Stoffwechsel der Gemeinschaften von Bodensediment.
Bevezetés
A Káspi-tó és a Fekete-tenger vidékéről származó vándorkagyló (Dreissena polymorpha) az egyik legsikeresebb r-stratégista faj (Vanderploeg és mts., 2002), melynek inváziója az 1920- as években kezdődött meg, majd a hajóközlekedés fejlődésével párhuzamosan teljesedett ki.
Egész Európában, majd Amerikában is gyorsan elterjedt. A Balatonban tömeges inváziójára 1933-ban figyeltek fel (Sebestyén, 1934; Entz és Sebestyén, 1942). A kagyló ma, az invázió után is nagy sűrűségben található elsősorban a part megerősítését szolgáló köves zónában, a nádon (Lakatos és mts., 2001), a hínáron (Muskó és Bakó, 2005) és egyéb, megtelepedésre alkalmas, alámerült szubsztrátumon, a parti öv gerinctelen élővilágának jelentős hányadát képezve.
A vándorkagyló megjelenésével előre megjósolhatatlan változásokat indukál. Nagy peteprodukciója a megtelepedésre alkalmas felületeken a kagyló tömeges megjelenését eredményezi. A vándorkagyló a víztestből rendkívül hatékonyan képes kiszűrni a részecskéket, eltávolítva ezáltal az algákat, a lebegőanyagokat, a baktériumokat és a zooplanktont, melynek révén nő a víz átlátszósága és módosulhat az alga összetétele (MacIsaac és mts., 1991, 1995;
Cotner és mts., 1995; Horgan és Mills, 1997; Pillsbury és mts, 2002). Az átszűrt részecskék egy részét szervesanyagban gazdag (Ricciardi és mts., 1997) fécesz és pszeudofécesz formában tömöríti és bocsátja vissza a víztestbe, ezáltal fontos szerepet tölt be az itt végbemenő anyag- és energiaforgalomban (Stanczykowska és mts., 1976). A vándorkagyló táplálékforrása a bukó vízimadarak és a halaknak (Ponyi, 1994; Speciár és mts., 1997; Bozzay és Hegedűs, 1978).
A kagylók fontos láncszemet jelentenek a litorális régió anyag és energiaforgalomában, többek közt az elsődleges megtermelt szerves anyag átalakítása, a lebegőanyagok és egyéb szerves anyagok lebontása, valamint a bentikus makroszkópikus gerinctelenek és a vízi csúcsragadozók szempontjából (Entz és Sebestyén, 1942; Specziár és mts., 1997).
Mindebből adódóan méltán illetik e világviszonylatban jól kutatott fajt az „ecosystem engineer” (Jones és mts., 1994; Karatajev és mts., 1997, 2002;Vanderploeg és mts., 2002), azaz az „ökoszisztéma arculatának jelentős formálója” megjelöléssel.
A Balatonban a vándorkagyló funkcionális ökológiai jellemzőiről kevés adat áll rendelkezésre, annak ellenére, hogy a vándorkagyló domináns a Balaton megtelepedésre alkalmas felületein. Jelentőségét a fentebb leírtak egyértelműen alátámasztják, így vizsgálata
fontos és meghatározó a tó ökoszisztémájának ismerete szempontjából. A részleteket tekintve kevéssé ismeretes a Balatonban a vándorkagyló mennyisége, a kagyló megtelepedése, táplálkozása és a táplálkozással kapcsolatos ülepítési tevékenysége (fécesz és pszeudofécesz képzése), a remineralizáció és a tisztítási ráta. Valamint annak ellenére, hogy jelentős interakció áll fenn a baktériumközösség és a vándorkagyló között, igen keveset tudunk a vándorkagyló bakteriális közösségre gyakorolt hatásáról.
1. Irodalmi összefoglaló
1.1. A vándorkagyló elterjedése, mennyisége
1.1.1. Rendszertani besorolása, inváziója
A Dreissena polymorpha Pallas magyarul vándorkagyló, vagy oroszkagyló, a Mollusca (puhatestűek) törzs, Lamellibranchiata (Kagylók) osztály, Heteroconchae (felsőrendű kagylók) alosztály, Eullamellibranchiata (lemezkopoltyúsok) rend, Dreissenidae családba tartozó, szilárd felületen megtelepedő, szűrő szervezet, mely intenzív inváziója révén hívta fel magára a figyelmet világszerte (1. kép).
1. kép. Vándorkagyló turzás az IJsermeer tó (Hollandia) partján
A „dreisenid” kagylók, mint a Dreissena polymorpha és az azonos családba tartozó Dreissena bugensis általában nem őshonosak jelenlegi élőhelyeiken. Eredeti hazájuk a Káspi-tó és a Fekete-tenger édes és brakkvízelvezető medencéi (Starobogatov és Andreeva, 1994).
Vándorlásuk véletlenszerű, jellemzően hajókon és csónakokon történő transzport, melyekre bisszuszfonal segítségével erősítik magukat, lárvájuk pedig a hajók ballasztvizében jut el távoli vízi ökoszisztémákba. A gazdaság globalizációja és a kereskedelem fejlődése jelentősen hozzájárult az inváziós édesvízi kagylófajok (legjelentősebbek: Dreissena polymorpha, Dreissena bugensis, Corbicula fluminea, Corbicula fluminalis és Limnoperna fortunei) tömeges előretöréséhez. Az inváziós fajok elterjedése általában nem egy lassú folyamat, hanem nagy távolság megtétele igen rövid idő alatt történik, mely gyors előretörésükhöz, térhódításukhoz vezet. Ezt számos emberi tevékenység felgyorsíthatja, pl. hajózható csatornák kiépítése, víztározók létesítése, politikai országhatárok módosulása, a politikai rendszer átalakulása, mely maga után vonja a határok elhelyezkedésének, a határon történő átlépés feltételeinek, a kereskedelemnek és a környezetjognak a változását, beleértve az esetleges migrációt is (Karatayev és mts., 2007). A származás vizsgálatok azt mutatják, hogy a vándorkagyló valamivel több, mint két évszázad alatt igen gyors ütemben terjedt el mesterséges és természetes vízi utakon, a kereskedelem fejlődésével kiépített csatornákon és az ezzel összeköttetésben lévő
folyókon. Dél Oroszországból a keleti-tengeri tartományokba az 1700-as évek végén, az 1800-as évek elején (Karatayev és mts., 2003), s innen szintén közvetítő csatornákon át egész Európába (Nalepa és Schloesser, 1993), majd Amerikába is (Herbert és mts., 1989) eljutott.
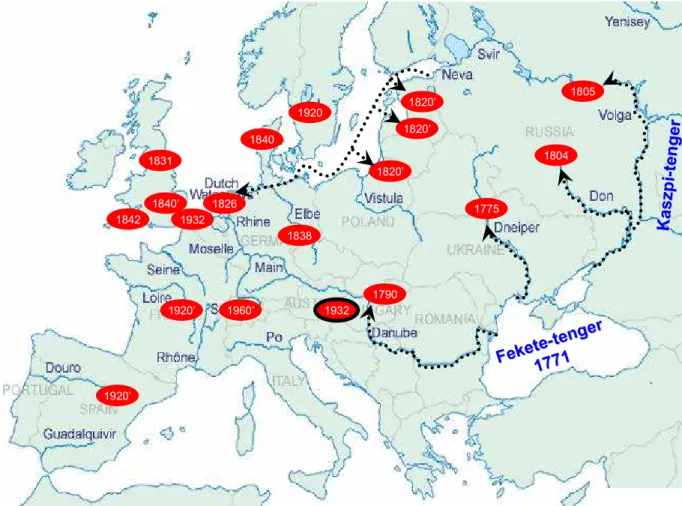
Inváziója és mennyisége elsősorban a geomorfológiai adottságok, a fitoplankton és a vízminőség függvényében alakul (1. táblázat).
1. táblázat. A vándorkagyló maximális denzitása különböző tavakban és folyókban.
Terület Denzitás [ind m-2] Referenciák
Lake Mikolajskie 2.200 Stanczykowska, 1975
River Rhine The Netherlands (1989) 21.000 Bij de Vaate és mts., 1992
Polish Lakes 6.720 Stanczykowska és Lewandowski, 1993a
Lake Garda 20.000 Franchini, 1978 *
Lake Constance 21.000 Walz, 1975
Lake Constance ∼ 330.000 Werner és mts., 2005
Lake Zurich 30.000 Burla és Lubini-Ferlin, 1976 *
Lake Dojran 4.000-5.000 Sapkarev, 1975 *
Dneprodzerzhinsk Reservoir 25.000-36.000 Gaidash és Lubanov, 1978 *
Szczecin lagoon 114.000 Wiktor, 1963 *
Lake Erie (1989-1990) 342.000 Leach, 1993
Lake Huron 300.000 Nalepa és mts., 1995
Lake Ontario (2000) 38.865 Dermott és mts., 2003
Illinois River (1993) 100.000 Schloesser és mts., 1996
*(Bij de Vaate, 1991)
Számos tanulmány kulcsfontosságú tényezőként említi még a kalcium koncentrációt a kagylók elterjedését illetően (Mellina és Rasmussen, 1994; O'Neill, 1996), és egyben rámutattak arra, hogy a legkiválóbb módszer a vándorkagyló élőhelyének meghatározására a kalcium koncentráció mérése. A kagylóhéj fő komponense a kalcium, melyből a kagyló héjképzése során jelentős mennyiséget igényel és egyben esszenciális a kagyló fejlődéséhez és reprodukciójához.
Sprung (1987) kimutatta, hogy 12 mg l-1 kalcium koncentráció alatt a kagyló nem termel lárvát.
A belorusz tavakban a vándorkagyló csak 25,4 mg l-1 kalciumtartalom mellett volt megtalálható (Karatayev, 1995), Észak-Amerikában azonban az európai adatoktól eltérően már 13 és 25 mg l-1 értékek mellett, sőt 8,5 mg l-1 kalciumtartalomnál is előfordultak kagylók (Strayer és mts., 1996;
Mellina és Rasmussen, 1994; Hincks és Mackie, 1997).
A vándorkagyló a Duna vízrendszerében 1790-ben jelent meg, Magyarországon az első példányt ekkor írta le Grossinger János a Holt-Zsitva és a Büdös-ér vizsgálata során (Bozzay és Hegedűs, 1978). Hazánkban a Balatonon (1932) kívül a Dunában és mellékvizeiben fordul elő (Oertel és Nosek, 2003; Bódis, 2007). A Szigetközben az ugyancsak inváziós Corbicula fluminea a leggyakoribb faj, a vándorkagyló pedig második legnagyobb gyakorisággal fordul elő. Míg az 1990-es évekig az erős áramlásnak kitett területen, addig 2001-től különösen a főági szakaszon található meg (Bódis, 2006). A Tiszában elvándorolt egészen Szolnokig és itt benyomult a Zagyvába is. A Tisza alsó folyásában a múlt század elején már nagy valószínűséggel élt, ahogy azt a Nemzeti Múzeum gyűjteményében lévő, Törökbecse mellett gyűjtött példányok bizonyítják (Bozzay és Hegedűs, 1978).
Az Európát behálózó csatornák megépítését követően a nemzetközi kereskedelem jelentős fellendülését követően a londoni Docks-ban 1824-ben látták először a vándorkagylót s ma már Anglia és Skócia minden folyójában jelen van (Kerney és Morton, 1970). Poroszországi kikötőkben 1825 óta gyakori. 1826-ban a Rajna torkolatáig jutott, később pedig bevándorolt a Loire folyóba is (Richnovszky és Pintér 1979). A vándorkagylót később behurcolták Németországba (1830) (2.
kép), Belgiumba (1833), Franciaorszába (1838), Dániába (1840), majd az 1920-as évektől Európa délnyugati és középső részére (Starobogatov és Andreeva, 1994). Annak ellenére, hogy Angliában az 1800-as években megjelent, Írországban csak 1997-ben (Minchin, 2000), Spanyolországban pedig csak 2001-ben (Bij de Vaate és mts., 2002) írták le először. Az Európában való elterjedést mutatja be a 1. térkép.
2. kép. Kőre telepedett vándorkagyló Mondsee (Ausztria)
1. térkép. A vándorkagyló inváziószerű terjedése Európában
A vándorkagylót 1986-ban Észak-Amerikába is behurcolták, a Kelet-Európából Amerikába közlekedő hajók ballasztvize által. Első példányát a St. Claire tóban találták 1986-ban (Herbert és mts., 1989). A megjelenést követően, 2 év alatt rendkívüli gyorsasággal elszaporodott a Nagy Tavak mindegyikében és a környező, ezekkel összeköttetésben lévő vizekben is elszaporodott, számos más édesvízi szervezettel együtt. A Mississipi folyó keleti részében nagy mennyiségben előfordul, többek közt annak csatornahálózatában (Bobeldyk és mts., 2005). A vándorkagyló hatása az észak-amerikai vizekben igen jelentős: a nagy vándorkagyló denzitás (1.
táblázat) és a geomorfológiai adottságok következtében megjelenő tömeges inváziója révén. Ezen tavakra jellemző köves fenéküledék és a vándorkagyló számára ugyancsak kiváló, megtelepedésre alkalmas felületet jelentő, azonos családba tartozó Dreissena bugensis (a D.
polymorpha-val ellentétben ez a faj a puha üledékben is képes élni) későbbi megjelenése (May és Fekete
Fekete--tenger tenger 1771 1771
Kaszpi Kaszpi--tenger
tenger
1804
1775
1805
1826 1932
1932 1790 1838
1840 1920
1960’
1820’
1842
1920’
1920’
1831
1840’
1820’
1820’
bugensist a Nagy Tavakban a vándorkagyló megjelenésével közel egy időben, az 1980 évek közepén hurcolták be (Mills és mts., 1993), ezzel szemben Európában nem terjedt el, annak ellenére, hogy a 20. század folyamán ezekkel a területekkel összeköttetésben lévő vizeken intenzív hajóközlekedés volt jellemző.
A vándorkagyló évente több millió dolláros kárt okoz az ipari és háztartási létesítmények vízvezetékének eltömítésével, korróziós folyamatok indukálásával (3. kép).
3. kép. Vándorkagyló kísérletben használt laboratóriumi eszköz csavarjainak korróziója
1.1.2. Megjelenése a Balatonban, korábbi kutatási eredmények
Sebestyén Olga adatai szerint (Sebestyén, 1934) a Dreissena polymorpha első két példányát 1932. szeptember 12.-én találták a tihanyi Kis-öbölben egy-egy tavi kagylóra (Anodonta cygnea) telepedve. A Balatonba való bekerülését tekintve, nagy valószínűséggel halászati eszközökkel és vízi járművekkel (4. kép) hurcolták be a Dunából a Sió-csatornán keresztül a tegzes bolharákkal (Chelicorophium curvispinum; Sars, 1895) együtt (Grossinger, 1791; Sebestyén, 1934, 1938).
4. kép. A Dunán és a Sió-csatornán át közlekedő hajó;
feltehetően ennek fenékdeszkájára erősödve hurcolták be a vándorkagylót a Balatonba
A Dreissenával közel rokon a Congeria Partsch nemzetség, melynek fajai valamikor miocén- és pliocénkori, kiédesülő-félben lévő tengereket népesítettek be. Így a Balaton helyén hullámzó Pannon tengerben éltek nagy számban. Congeria maradványok (fosszíliák) a Balaton környékén gyakoriak és búbrészük "kecskekörmök" néven általánosan ismeretes.
1933 nyaráig a Balatonban mindössze tíz magányos vándorkagylót gyűjtöttek össze (mindegyiket Tihanyban), a nyár második felében azonban a tó több területén egyre nagyobb
számban volt megtalálható. 1933 júliusától 1934 áprilisáig az összegyűjtött kagylók 67,2%-a vándorkagyló volt. 1934-ben már a strandolók figyelmét is felkeltették a lépcsőkön, parti tárgyakon megtelepedő vándorkagyló csoportosulások (Sebestyén, 1937).
1934 októberére a felnövekedett egyedek hossza elérte a 2-16 mm-t, melyek 75 %-a 4-11 mm közötti volt. 1935 tavaszára a kagylóturzások jelentős mennyiségét a vándorkagyló alkotta a Tihanyi-rév környékén. 1936 októberben 2-10 mm méretű egyedek voltak jellemzőek, melyek 77,6 %-a 2-5 mm-es volt. Mindezek a növekedés lassulását mutatják (Sebestyén, 1937).
1936-ban a vándorkagyló lárvák mennyisége a vízmélységtől függően 4-27 ind l-1 között változott Tihanynál (Entz és Sebestyén, 1942).
1963-ban a Keszthelyi-öbölben csupán üres vándorkagyló héjakat (51 ind m-2) találtak (Entz és mts., 1963), 1982-ben szintén csak héjakat találtak, mennyiségük azonban sokkal jelentősebbnek (8.000 ind m-2) mutatkozott (Entz, 1982).
1979-ben Lakatos 2-8 egyedet talált 100 cm2 nádfelületen, a parti köves zónában pedig tömeges előfordulását regisztrálta. A tihanyi szennyvíztisztító telep befolyójánál és a Tihany- Gödrös nádas területénél (kontroll) végzett biotekton vizsgálatok (élőbevonat analízis) során megállapították, hogy a szennyvíztisztító telep befolyójánál kevesebb a vándorkagyló, mint a kontroll területen, mely egyértelműen a szennyező hatásnak tudható be (Lakatos, 1979).
A Balatonban a vándorkagyló a bukó vízimadarak (Fuligula ferina, Bucephala clangula, Fulica atra, Aythya fuligula és Anas platyrchynchos) és a halak (főként ponty és keszegfélék:
Cyprinus carpio, Blicca bjoerkna) - különösen a veresszárnyú koncér (Rutilus rutilus) - számára szolgál jelentős táplálékforrásul (Entz és Sebestyén, 1946; Bozzay és Hegedűs, 1978; Bíró, 1974;
Ponyi, 1994; Specziár és mts., 1997). Entz 1981-es munkája szerint a vándorkagyló mennyiségének drasztikus csökkenése a szárcsaállomány jelentős őszi-téli növekedésével (kifalással) magyarázható (Entz, 1981). A Bódeni-tó sekély vízterületein ősztől tavaszig vándorkagyló állomány megcsappanása figyelhető meg, mely ugyancsak a szárcsák tömeges kifalásának a következménye (Werner és mts., 2005). A veligera lárva a halivadékok értékes táplálékforrása lehet (Stanczykowska, 1977).
Richnovszky (1981) a vándorkagyló állomány csökkenése szempontjából nem tartja elégséges magyarázatnak a halak és a szárcsák tömeges kifalását, inkább Ponyi és más szerzők megállapításait támogatja, miszerint a tó eutrofizálódásának előrehaladását jelzi a kagylópusztulás. A Mazuri-tavak esetében ugyanis a fokozódó eutrofizációval magyarázták az
egyre jobban visszaszoruló vándorkagyló állományt, egyes tavakban a drasztikusabb eutrofizációból adódóan mára már teljesen eltűntek a vándorkagylók (Stanczykowska, 1997;
Stanczykowska és Lewandowsky, 1993a, 1993b).
Ponyi 1982-ben összegezte a vándorkagyló egyedszámra vonatkozó adatokat, s megállapította, hogy míg 1966-ban 25-3.436 ind m-2, addig 1968-ban 0-3.720 ind m-2 között változott a Balaton egyes medencéiben. Eredményei alapján egy kifejlett balatoni vándorkagyló megközelítőleg 1 cm nagyságú (Ponyi, 1985).
1998-ban a Balatonban Tihanynál a mederfenék közelében 2.000-6.000 ind m-2 vándorkagylót találtak (Kozák és mts., 1998). Az első vándorkagyló biomasszára vonatkozó adatokat Muskó és Bakó (2002, 2005) közölte.
A Balatonba helyezett mesterséges felületeken (kémcsőkefe és csempe) megtelepedő gerinctelen állatok vizsgálata során azt tapasztalták, hogy a vándorkagyló a kolonizáló állatok jelentős hányadát alkotta (Lakatos és mts., 1997; Kozák és mts., 1998; Muskó és Russo, 1999). A mesterséges aljzatok benépesülési dinamikájának tanulmányozása során a kihelyezett kémcsőkefékre eleinte a vándorkagyló telepedett meg, később azonban a tegzes bolharák (Chelicorophium curvispinum) kiszorította a vándorkagylót (Muskó és Russo, 1998, 1999).
1999 nyarán a vándorkagyló lárvák tekintélyes mennyiségben voltak jelen a Balaton nyíltvizében kelet-nyugat irányú (B.akarattya-Tihany-B.akali-Szigliget) emelkedő gradienst (9,4- 75,5 ind l-1) mutatva. Májusban, amint a víz hőmérséklete a 17°C-ot elérte megjelentek az első lárvák, melyek nyár közepére már jelentős mennyiségben voltak jelen, egyes nyugati területen akár a 250 ind l-1 értéket is meghaladták (Muskó, 2001; G.-Tóth, 1999). 2001-ben a vándorkagyló lárva denzitása a Balatonban átlagosan 0,86-74,59 ind l-1 volt 2002-2004 között lecsökkent 0,28- 16,25 ind l-1 értékre, majd a vízszintemelkedéssel párhuzamosan növekedett a veligera lárva mennyisége és 2007-ben már 2,57-33,34 ind l-1 között változott (G.-Tóth és mts., benyújtott dolgozat). A hínáros élőbevonatában a vándorkagyló 2-64 %-ban fordult elő (G.-Tóth és mts., 2002).
Az Alsóörs-Siófok-Szigliget-Balatonfenyves tengely mentén vett mintavételek alapján a parttól távolodva a zooplanktonban a vándorkagyló lárva százalékos arányának növekedése figyelhető meg. Míg partközelben a zooplankton 3%-át teszi ki, a nyíltvízben elérheti akár a 10
%-ot is (Németh, 2003).
Összességében azt lehet mondani, hogy a kezdeti tömeges, inváziószerű vándorkagyló megtelepedés (1930-as évek) oka a megfelelő életkörülmények és a természetes ellenség hiánya volt. Egy bizonyos maximális kifejlődés után a kagyló denzitása csökkent, jelentős pusztulást követően pedig (Sebestyén, 1937) egy új biocönotikus egyensúly állt be (Entz, 1941), melyben a kezdeti értékekhez képest mind a kagylók mérete, mind a száma jelentősen csökkent. A Balatonban tapasztaltak (Richnovszky, 1981), néhány Balatonban végzett terepi felmérés (Lakatos, 1979) és nemzetközi eredmény (Stanczykowska, 1997; Stanczykowska és Lewandowsky, 1993b) alapján is arra a következtetésre jutottak, hogy az eutrofizálódás fokozódásával visszaszorult a vándorkagyló. A kagyló mennyisége időnként csökkent, időnként pedig nőtt, s több mint 70 év elteltével a lárva adatok alapján napjainkban újra a kagyló előretörése tapasztalható (G.-Tóth és mts., 2002; G.-Tóth és mts., benyújtott dolgozat).
1.1.3. A Balaton, mint a vándorkagyló élőhelye
A Balaton a Föld egyik legjobban kutatott, Közép-Európa legnagyobb édesvizű sekély tava (átlagos mélység: 3,2 m; területe: 596 km2; partvonala: 240 km; 78 km hosszú; 4,5-12 km széles;
térfogata: 2 km3). A tó legmélyebb pontja a Tihanyi-félsziget déli csúcsa előtt végignyúló árokszerű mélyedés, a „Kút”, melynek mélysége megközelítőleg 11,5 méter. A tó medencéjének elhelyezkedése, alakja, valamint a domináns, erős észak-nyugati szél miatt a hullámverés a déli partot építi, az északi part vonalát pedig pusztítja. A tó teljes víztömegének mozgása szempontjából a legjelentősebbek a tihanyi-szorosban és a délnyugati tórészben keletkező áramlások.
A Balaton nagyobb mint 5000 km2-es vízgyűjtő területe hazánk határain belül található. A tó vizének évi természetes ingadozása közel 1 m-t tesz ki, s e periodikus ingadozásokat 1863 óta próbálják szabályozni a Sió-csatornával, valamint a Sió-zsilippel. Csapadékos esztendőben a zsilip kinyitásával elárasztják a Sió-csatornát, azonban főként a nyári időszakban a nagy párolgásból adódó vízszintcsökkenés miatt a Sió-zsilipet időnként zárva tartják. A tó vízszintjét 1997-től 70-110 cm (minimum és maximum szint) közt szabályozták (Varga, 2005). 2000 nyarától a vízszint folyamatosan csökkent, az extrém meleg, csapadéktalan nyár és a nagy párolgás következtében. 2003 októberére a vízszint jelentősen csökkent (23,7 cm), a tó
Átlagos vízszint: 75 cm Átlagos vízszint: 88 cm
történetének egyik legaszályosabb periódusát (2000 - 2003) követően (Varga, 2005). Hasonlóan alacsony vízszinteket korábban 1921-ben, 1949-ben és 1961-ben is feljegyeztek (1. ábra).
1. ábra. A vízszint változása 1901-2003 között (Varga, 2005).
A Balaton partvonalának hossza jelentős, ebből adódóan a parti öv - keskenysége ellenére - igen fontos élettér. A mederfenék iszapos (Lóczy, 1894; Miller és Wagner, 1978), homokos üledék, így a partot megerősítő, 105 km hosszan elnyúló mesterséges kőszórások, a nagy kiterjedésű nádasok és a szigetszerűen elterülő hínárosok felülete (a tó 5%-át borítja vízi növényzet, mely túlnyomó részt hínár és nád [Ponyi, 1981]), mint szilárd, megtelepedésre alkalmas szubsztrátum jelent elsősorban élőhelyet a vándorkagylónak.
A Balaton nagy kiterjedésű állóvíz, azonban sekélységéből adódóan a vízszínt változása jelentősen befolyásolja a megtelepedésre alkalmas felületeken a vándorkagyló mennyiségének alakulását. A vízszintcsökkenés hatására a tó litorális zónája eltolódik, változik a fény lehatolása, a fény intenzitása a mederfenéken, valamint a bentosz vertikális zonációja figyelhető meg (Palomäki, 1994). Az ebből adódó fényviszonybeli változások a litorális zónában a legszembetűnőbbek, ahol számos bentikus organizmus él együtt.
A tó 1960-1980 közti eutrofizációs periódusát (Herodek, 1988) oligotrofizálódás követte.
A fitoplankton biomasszája azonban nemcsak időben, de térben, a tó hossztengelye mentén is változik. A hossz-szelvény vizsgálatok alapján a Szigligeti és a Keszthelyi-medence a tó
legeutrófabb területe. A Keszthelyi-medencében kis mértékben, esetenként jelentősen, nagyobb biomassza jellemző, mint a Szigligeti-medencében és a keleti medence felé haladva csökken a biomassza értéke (Padisák, 1994, 2002; Padisák és Reynolds, 1998; Tátrai és mts., 2000; Vörös és mts., 2000; Jolánkai, 2001).
A Balaton vízminőségének javulása elsősorban a kis-balatoni „szűrőrendszer”-nek köszönhető, mely természetes úton tisztítja a Zala folyó vizét. Így a foszforterhelés kb. felére csökkent, a tározó az algák számára legkönnyebben felvehető foszfátnak 90%-át tartja vissza, s emellett a nitrogén visszatartás sem elhanyagolható. A tó közvetlen parti sávjának és a vízgyűjtő terület településeinek csatornázottsága jelentős mértékben kiépült, ezzel megakadályozva a talajvíz fokozódó elszennyeződését, valamint a szennyezőanyagok ilyen módon történő tómederbe jutását.
1.2. A vándorkagyló megtelepedése, fejlődése, életciklusa
Radiológiai vizsgálatok a vándorkagyló élettartamát 3-4 évben állapították meg, az állat csak speciális esetben él 4 évnél tovább (Morton, 1969). Más szerzők szerint a vándorkagyló általában 2-9 évig él, a maximális méretét - mely 40 mm körüli - 1-2 év alatt éri el (Mackie és mts. 1989;
Smirnova és Vinogradov 1990; Mackie 1993; Mackie és Schloesser 1996). A Balatonban egy kifejlett kagyló megközelítőleg 1 cm nagyságú (Ponyi, 1985).
A gametogenezis ősszel, vagy télen kezdődik, a peték általában nem egyszerre, hanem az egész vegetációs időszak alatt (késő nyárig, vagy kora őszig nyúlik el) termelődnek. A peték 70 µm nagyságúak, igen jelentős mennyiségben keletkeznek (több tízezer, millió darab) (Mackie és mts. 1989; Sprung 1993; Mackie és Schloesser 1996; Nichols 1996; McMahon 1996) és azonnal osztódni kezdenek. A nőstény által fehér felhő formájában kibocsátott peték gyorsan leülepednek a mederfenékre. A spermiumok a hímekből szintén fehér felhőként távoznak. Az állatok fő tenyészideje májustól október közepéig tart, melyhez a szükséges minimális hőmérséklet 12-15
°C (Lvova és mts., 1994).
A kagylólárvák 2-9 nappal a megtermékenyítést követően már 70-100 µm hosszú héjjal rendelkeznek (Sprung 1993; Ackerman és mts. 1994). A planktonikus lárvaállapot stádium (2.
ábra/a, b kép) 6-8 hétig tart, s mivel a lárvák nagyon aprók, úszóképességük pedig nagyon csekély, korán letelepszenek (2. ábra/c-e kép). A planktonikus lárva mozgása rendszerint egy passzívan mozgó részecskéhez hasonlítható, azaz véletlenszerű (Zimmer és Butman, 2000).
Amikor a lárva érintkezik a felülettel a bisszusz mirigy által termelt váladék a vízben fonalszerűen megszilárdul, s ezzel a 100-200 merev szálból álló bisszusszfonallal lehorgonyozza magát különböző tárgyakra, kövekre, rákokra, kagylókra (általában Unio- vagy Anodonta fajok héjaira), hínárra, nádszálakra, esetenként az üledékbe (Ackerman és mts., 1994; Mellina és Rasmussen 1994; Nichols 1996; Berkman és mts. 1998). Előfordulhat az is, hogy a végleges megtelepedés előtt esetlegesen néhány órán, néhány napon keresztül egyéb élőhelyre elmászik (Kobak, 2001), jobb felületet keres a letelepedéshez: ez esetben a kagyló akár 7 cm-t is megtesz egy éjszaka alatt (Ackerman és mts., 1994). Gyakran a post-veligera és a fiatal kagyló a hullámokat kihasználva, a lesüllyedést elkerülve, a bisszusz fonalak segítségével úszó felületekre telepedve nagy távolságok megtétele után áttelepedik más felületre (Lewandowski, 1982; Martel, 1993, Baker és Mann, 1997, Horváth és Lamberti, 1997).
A letelepedő stádium elején, az aktív táplálkozás megkezdésekor igen jelentős mortalitás tapasztalható, mely európai vizek esetén akár 99%-ot is elérhet. A veligera (mérete 167-300 µm, 2. ábra) potenciális préda az igen széles mérettartományban (0,4 µm-től [Cotner és mts., 1995]
1,2 mm-ig [Horgan és Mills, 1997]), rendkívül hatékonyan és intenzíven szűrő kifejlett vándorkagyló számára. Feltehetően ez is jelentősen hozzájárulhat a magas lárvamortalitáshoz, amely a Bodeni tóban (Walz, 1978b), vagy a lengyel Taltowisko tóban (Lewandowski, 1982;
Stanczykowska, 1997) elérheti akár a 99-99,86%-ot is. Mindebből adódóan a vándorkagyló életciklusa során a veligera stádium a legkritikusabb.
A letelepedett állat (2. ábra/e kép) egy év múlva, azaz a következő évre válik ivaréretté, amikor a héj méret megközelítőleg 8 mm (Sprung, 1992). A kifejlett állat bármikor leválhat az aljzatról, akár vándorolni, vagy újból letelepedni is képes, friss bisszuszt képezve (Entz és Sebestyén, 1942; Ackerman és mts., 1994). Általában csomókban, fürtöket alkotva élnek.
2. ábra A vándorkagyló fejlődési alakjai (planktonikus lárvaállapot stádiuma (a, b) és a letelepedett állat (c-e) (Ackerman, 1994).
A téli időszakban összezárulnak, elszakítják bisszuszfonalukat és a parti fövénybe fúródnak, telepeket alkotva védik meg magukat a fagyástól (Entz és Sebestyén, 1942).
1.3. A vándorkagyló ökoszisztémára gyakorolt hatása
1.3.1. Kompetíciója más bevonatlakókkal
Az egzotikus fajok, mint a vándorkagyló is, lényegesen képesek átalakítani környezetüket kompetitív kihívást jelentve őshonos fajoknak (Griffits, 1993; Nalepa és mts., 1998). A vándorkagyló a vízbe kerülő tárgyakon és jóformán bármely vízi élőlényen képes megtelepedni, így nagyon gyakran egyéb kagylófajok (Unio, Anodonta) héján, nádszálakon, hínáron, vagy más vízi élőlényeken, valamint köveken, bójákon, oszlopokon és a hajók, csónakok oldalain és bármely más csupasz felületen (5. kép).
A vándorkagyló fejlődési alakjai
a.
b.
c.
d.
e.
Láb
Fejlődés Növekedés
Metamorfózis:
A vitorla eltűnik Dissoconch Pediveliger
(>167 −−−−<300 µm)
Plantigrade (>158 −−−−500 µm)
Juvenile (500 −−−−5000 µm)
Kifejlett kagyló
Spermium (4 −−−−9 µm)
Pete (40 −−−−96 µm) Megtermékenyítés
Embrió Trochophore (57 −−−−121 µm) D-alakú veligera
(70 −−−−160 µm) Veliconcha
(120 −−−−280 µm) Pediveliger
(>167 −−−−<300 µm)
Prodissoconh II Vitorla
Prodissoconh I
5. kép. A vándorkagyló megtelepedése különböző aljzatokon.
A Balatonban, a köves-kavicsos medrű tavakhoz képest viszonylag kevés megtelepedésre alkalmas felület található, mely megfelelő életteret jelentene a vándorkagylónak, vagy egyéb helyhez kötött szervezet számára, így ezek birtoklásáért nagy harc folyik az ökoszisztémán belül.
A Balatonban a gerinctelen állatok közül a vándorkagyló telepedik meg legsikeresebben a vízbe bekerült különböző tárgyakon (Muskó és Görög, 2000). A Balatonba helyezett mesterséges aljzatokon (kémcsőkefe és csempe) megtelepedő gerinctelen állatok vizsgálata rámutatott arra, hogy a vándorkagyló jelentős hányadát alkotja a bevonat faunájának (Lakatos és mts., 1997;
Kozák és mts. 1998; Muskó és Russo, 1998, 1999).
Az aljzatként szolgáló élőlények életkörülményire, életműködésére negatív hatással van a rátelepedő élőlény, számos esetben a pusztulásukhoz is vezethet. Mindez jól megfigyelhető az Unionidae és a vándorkagyló ilyen jellegű kapcsolata esetén (6. kép). Az unionidák szempontjából a héjra telepedő vándorkagyló mind a mozgás (ami amúgy is lassú), mind pedig a táplálkozás (hasonló) szempontjából negatív hatást jelent. Az Unionidae fajok pusztulása esetén azonban csökken a mederfenéken a rögzülésre alkalmas aljzat, mely egyúttal csökkentheti a vándorkagyló mennyiségét.
6. kép. A vándorkagyló megtelepedése nagyobb méretű Anodonta kagylón
7. kép. A szivacs megjelenése a vándorkagyó ágyakban.
A vándorkagyló egyik konkurrense a szivacs (Spongillidae). A szivacs intenzív növekedése során, mintegy befonja a kagylót (7. kép), miközben a sipho nyílásához bevezető rés fokozatosan szűkül, lassan pedig az egész elzárul, a kagyló nem jut tápanyaghoz, rövidesen éhen pusztul (Sebestyén, 1934).
A kecskerák (Astacus leptodactylus) páncélját gyakran vándorkagylók lepik el, mely a kecskerák vedlését nem akadályozza (Sebestyén, 1934).
A Balatonban a vándorkagylót és a tegzes bolharákot közel egy időben (1932) hurcolták be, itt a két faj koegzisztál, mennyiségük fluktuál különböző környezeti tényezők hatására (Muskó, 2001; G-Tóth és mts., 2002; Muskó és Bakó, 2005). E tekintetben eltérés tapasztalható a Balaton és a Rajna között, mivel az utóbbi esetében a kagylóinváziót (1826) követően jelent meg a tegzes bolharák (1987), mely a kolonizálásra alkalmas felületekért, valamint a táplálékért
komoly konkurrenciaharcot folytatott a kagylóval és annak visszaszorulásához vezetett (van der Velde és mts., 1994, 1998; Rajagopal és mts., 1998).
1.3.2. Filtrációja, fécesz és pszeudofécesz kibocsátása
A vándorkagyló a víztestből rendkívül hatékonyan és igen széles, 0,4 µm-től (Cotner és mts., 1995) 1,2 mm-ig (Horgan és Mills, 1997) terjedő mérettartományban képes kiszűrni a részecskéket (8. kép), eltávolítva ezáltal az algákat és a lebegőanyagot (Kryger és Riisgard, 1988;
Reeders és mts., 1989; Reeders és Bij de Vaate, 1990; Bunt és mts., 1993; Stanczykowska és Lewandowski, 1993a; Roditi és mts., 1996; Caraco és mts., 1997; Horgans és Mills, 1997; G.-
Tóth, 1999), a baktériumokat (Cotner és mts., 1995; Silverman és mts., 1996), a protozoákat (Lavrentyev és mts., 1995; Jack és Thorp, 2000) valamint a zooplanktont, beleértve a saját lárváját is (Shevtsova és mts., 1986; MacIsaac és mts., 1991, 1995; Lavrentyev és mts., 1995).
8. kép. Aktívan szűrő vándorkagylótelep.
A táplálék az inhalens (orális) szifón keresztül a nyálkás váladékot termelő csillós kopoltyúhámon át, a részecskéket tömörítő, szétválasztó és továbbító csillókkal jut a köpenyüregbe. A kagylónál a gyomor simaizmait egy forgó mozgást végző kristálynyél helyettesíti, melyből emésztést elősegítő mirigyváladék válik ki. A tápanyag a gyomrot körülvevő középbéli mirigyben diffúzióval szívódik fel. A mirigy végzi az emésztést, felszívja a megemésztett tápanyagot, raktározza azt, elvégzi a szükséges biokémiai átalakításokat, méregtelenít, majd diffúzióval juttatja a hemolimfába, onnan az egyes szervekbe az azok működéséhez szükséges anyagokat. Az emésztetlen anyagok, az anyagcsere végterméke (fécesz) a lábban futó középbéli szakaszon a szívburok által körbenőtt utóbélen keresztül az exhalens (anális) szifón át távoznak.
Nagy alga koncentráció esetén a kagyló több részecskét szűr ki, mint amennyit feldolgoz az emésztőrendszere, a felesleget pedig pszeudofécesz (általában nagyobb mennyiségben jelen
lévő és, vagy rosszabb minőségű, csillókkal tömörített részecskék, szeszton, mely a köpenyüregből golyók formájában, nyállal keveredve távozik a bemenő szifón át) formájában bocsátja ki, mielőtt az a bélbe juthat (Hawkins és Bayne, 1992). Mérsékelten nagy koncentrációjú részecskemennyiség esetén az aktuális szűrési ráta adaptálódhat az emésztőrendszer kapacitásához, a kopoltyú aktivitásának szabályozása révén (Sprung és Rose, 1988). Alacsony táplálék koncentráció esetén a táplálékfelvétel megegyezik a zavartalan körülmények mellett jellemző szűrési rátával, azaz a filtrációs vagy fogyasztási kapacitással.
A vándorkagyló filtrációs/tisztítási rátájának világirodalmi adatai a 2. táblázatban találhatók.
2. táblázat. A vándorkagyló filtrációs rátája világirodalmi adatok alapján.
Szerzők Táplálék Kagylóméret
(mm)
Hőmérséklet (C°)
Tisztítási/Filtrációs ráta (ml ind-1 óra-1)
Kondratev (1963) Agyag szuszpenzió 22 16-17 133*
Mikheev (1966) In situ 22 20-22 52*
Mikheev (1967) In situ 22 - 2-50*
Mikheev és Sorokin. (1966) 14C izotóppal jelölt táplálék 22 - 46*
Stanczykowska (1968) In situ 22 - 35*
Alimov (1969) Agyag szuszpenzió 22 18-20 159*
L'vova-Katchanova (1971) - 22 - 43-56*
Morton (1971) Grafit 22 5-30 6,9*
Különféle algák 29 5-30 5-180
Chlamydomonas globosa 29 30 180
Euglena spirogyra 29 30 47
Hinz és Scheil (1972) Szuszpenzió 22 5 13*
Stanczykowska és mts. (1975) In situ 22 22-25 8-44*
Stanczykowska és mts. (1976) In situ - - 10-100
Walz (1978a) Nitzschia 22 15 63*
Benedens és Hinz (1980) Grafit 22 17,7 4*
Kryger és Riisgard (1988) Szuszpenzió 22 20 287*
Reeders és mts. (1989) Chlorella 22 4,5-17,7 14-78
Alga, agyag 15-25 4,5-17,7 2-287
Reeders és Bij de Vaate (1990) In situ 22 10-21 40-75*
In situ 22 3-5 18-51*
Hudson folyóvíz
Bastviken és mts. (1998) 100 µm-es hálón átszűrt Hudson folyóvíz
Átlagosan 11,5
16-20 24-63
Lei és mts. (1996) Mikrogömbök, szeszton 12-23 15 74
Horgan és Mills (1997) Chlamidomonas sp. 9-22 17 48-179
Mallamonas sp. 9-15 17 51-125
Cryptomonas ovata 9-22 17 88-339
Baldwin és mts. (2002) Chlamidomonas 20
Ackerman (1999) 0,2 µm-es hálón átszűrt Erie tó víz és
Chlorella pyrenoidosa
32,5 20,6 658
MacIsaac és mts. (1995) Keregesféreg, veligera 6-22 16 0-100
*(Reeders és mts., 1993)
Morton (1971) munkájában összehasonlítva az addigi világirodalmi adatokat, azt tapasztalta, hogy a vándorkagyló szűrési rátája sokkal kisebb, mint általában a tengeri kagylóké (Mytilus edulis, Cardium edule, Pecten irradians, Venus mercenaria, Mya arenaria és Srobicularia plana). A vándorkagyló lárvájának tisztítási rátája Dionisio Pires és mts. (2004) szerint 2,58 és 16,66 µl h-1 között változik, MacIsaac és mts.-nak (1992) eredményei alapján pedig 245 és 418 µl ind-1 h-1 között.
Néhány tanulmány arról számol be, hogy a kagyló feltehetően a bemenő szifójánál kisebb részecskéket tudja kiszűrni, eltekintve néhány speciális tulajdonságokkal rendelkező algától (pl. a cianobaktériumok; Nicholls és Hopkins, 1993; Horgan és Mills, 1997). A kagyló a természetes szesztont gyorsabban szűri, mint az inert anyagokat (Morton, 1971; Lei és mts., 1996).
Egyes tanulmányok szerint a kagyló képes szelektív szűrésre, bizonyos algafajokat szelektíven kiválaszt és azonnal vissza is bocsát a víztestbe laza pszeudofécesz formában, mely életképes algákból áll (Reeders és Bij de Vaate, 1990; Lavrentyev és mts., 1995; Baker és mts., 1998; Bastviken és mts., 1998; Vanderploeg és mts., 2001; Naddafi és mts., 2007), mások azonban ennek ellenkezőjéről számolnak be (Nicholls és Hopkins, 1993; Horgan és Mills, 1997).
Ten Winkel és Davids (1982) eredményei úgyszintén mutatnak szelekciót. Ez alapján a köpenyüregben és a gyomorban jelen lévő 15-45 µm átmérőjű részecskék pozitív, ezzel szemben az ennél kisebb és nagyobb méretűek hiánya negatív szelekcióra utal. Naddafi és mts. (2007) szerint a vándorkagyló szűrésének szelektivitását az elérhető táplálék méretének és minőségének
függvényében szabályozza és filtrációs rátája jelentősebbnek mutatkozik heterogén algakultúra esetén, mint egy algafaj jelenlétében. A kibocsátott pszeudofécesz általában 50 µm-nél nagyobb és 7 µm, vagy annál apróbb fitoplanktont tartalmaz nagy mennyiségben, mely a fitoplankton minőségi és morfológiai szelekciójával magyarázható, mivel a méret és a sejtfal struktúrája, felülete is meghatározó a szelektiviás hatékonyságának szempontjából (Baker és mts., 1998;
Naddafi és mts., 2007). Számos esetben előfordulhat, hogy a fitoplankton a sejtfalában ill.
felületén található glikoproteidek révén tapad meg a kagyló tápcsatornájában (Waite és mts., 1995).
A 7 µm-nél kisebb méretű fitoplanktonnak alacsony a hosszú szénláncú telítetlen zsírsav koncentrációja, mely nem előnyös a kagylónövekedés szempontjából (Vanderploeg és mts., 1996). Ten Winkel és Davids (1982) és Bastviken és mts. (1998) szerint a vándorkagyló az algafajok közül elsősorban a Cryptomonas fajokat preferálja.
Jorgensen és mts. (1984) szerint a vándorkagyló az 1 µm-nél nagyobb részecskéket közel 100%-ban visszatartja. Mások szerint a tápanyag visszatartás hatásfoka csak 5 µm-nél nagyobb részecskeátmérő esetén maximális (Sprung és Rose, 1988). Lei és mts. (1996) különböző átmérőjű mikrogömböket használtak filtrációs kísérleteik során, melyben hasonlóan az előbbiekhez azt tapasztalták, hogy a 0,5 µm átmérőjű részecskékhez képest az 1 µm átmérőjű részecskéknél a filtráció 4,8-szorosára, az 1,5 µm-es gyöngyök esetén pedig 3,5-szörösére nőtt.
Azonban 1,5 µm-nél nagyobb átmérőjű részecskéknél nem tapasztaltak jelentős változást a szűrésben, a visszatartás hatékonysága pedig 87-100%-nak mutatkozott.
Morton (1969) munkájában azt írta, hogy a táplálék szelekciója inkább mennyiségi, mint minőségi folyamat. Kísérlete során azt tapasztalta, hogy a vándorkagyló jobban preferálja a kisebb méretű részecskéket, ez esetben azonban nagyobb koncentráció szükséges a maximális szűrési ráta eléréséhez (Morton, 1971).
Azt is megfigyelték, hogy míg a kiszűrt algákat 20%-ban, addig a baktériumokat 50%- ban asszimilálja a kagyló (Mikhaev, 1966), természetes tápláléka is főként egysejtű algákból és baktériumokból áll (Morton, 1971), Stanczykowska (1977) szerint tavasszal elsősorban diatómával, kék- és zöld algával táplálkozik.
Pillsbury és mts.-nak (2002) összefoglaló munkájában a Huron tó Saginaw öblének fitoplankton közösségében bekövetkező változások azt mutatják, hogy a vándorkagyló
zöldalgák biomasszájának feltehetően a jobb fényviszonyokból adódó növekedése. A kagyló megjelenését követően négy évvel Microcystis spp. algavirágzás volt jellemző a Saginaw öbölben, melynek hátterében a Microcystis spp. szelektív elutasítása (rossz íz, az alga által termelt microcystin nevű toxin kellemetlen szaga), esetlegesen a normálisan, korábban jelen lévő fitoplankton hiánya állhat (lásd Pillsbury és mts., 2002).
A Huron- és az Onedia tóban és a Saginaw öbölben megfigyelhető cianobaktérium virágzás okát más munkákban úgyszintén azzal magyarázták, hogy a kagyló elősegíti a virágzást, mivel a cianobaktériumot egyáltalán nem, vagy kevésbé aktívan szűri ki, mint a többi algát (Heath és mts., 1995; MacIsaac, 1996; Vanderploeg és mts., 1996; Budd és mts., 2001;
Vanderploeg és mts., 2001; Contoy és mts., 2005).
A vándorkagyló az Erie tóban is hozzájárulhatott a fitoplankton biomassza gyarapodásához, feltehetően azáltal, hogy fokozta a tápanyag „turnover” idejét, és egyúttal elősegítette a cianobaktérium virágzást is, melyhez Vanderploeg és mts. (2001) szerint az is hozzájárul, hogy a kagyló a Microcystis spp. algát szelektíven elutasítja.
Nagy kagyló denzitás és hosszú ideig tartó, előbbiekben említett szelektív filtráció, a leírt jelentős mértékű változásokhoz is vezethet (Baker és mts., 1998).
Mások ezzel szemben azt tapasztalták, hogy a kagyló eredményesen távolítja el a cianobaktériumokat (Bastviken és mts.,1998; Reed-Andersen és mts., 2000). Jack és Thorp (2000) Ohio folyóban végzett in situ kísérlete azt mutatta, hogy a nagy vándorkagyló denzitás visszaszorította a cianobaktérium virágzást és hozzájárult a diatomák denzitásának növekedéséhez. Az algavirágzást ugyanis sok egyéb tényező is kiválthatja úgy, mint a tápláléklánc manipuláció, számos inváziós faj megjelenése és egyéb tényezők (Vanderploeg és mts., 2001). Raikow és mts. (2004) 61 michigani tó tanulmányozása során azt találta, hogy azokban a tavakban, ahol jelen van a vándorkagyló, a klorofill-a és a számolt alga alapján alacsonyabb a teljes fitoplankton biomassza, mint azokban, ahol nincs jelen a kagyló.
Amennyiben a teljes foszfor koncentráció 25 µg l-1-nél kisebb volt, a vándorkagyló invázió hozzájárult a Microcystis dominanciájához. A hollandok biomanipulációs kísérletekben előszeretettel alkalmazzák a vándorkagylót. Kísérleteik azt mutatják, hogy mind a kolóniás, mind a szálas, akár toxikus, akár nem toxikus algát (Microcystis és Planktothrix) eltávolítja a sekély holland tavakban (Dionisio Pires és mts., 2005).
A vándorkagyló intenzív szűrése révén nő a víz átlátszósága, így a fény lehatolása (Stanczykowska és mts., 1976; Holland, 1993; Leach, 1993; Lowe és Pillsbury, 1995; MacIsaac, 1996; Mayer és mts., 2002; Pillsbury és mts., 2002, Barbiero és Tuchman, 2004), mely előnyös lehet a bentikus életközösségnek, növelheti a biodiverzitást (Strayer és mts., 1999), hozzájárulhat a hínár előretöréséhez (Skubinna és mts., 1995) valamint csökkenti a fitoplankton biomasszát (Reeders és mts., 1989; Fahnenstiel és mts., 1995; Fanslow és mts., 1995; Caraco és mts., 2006) és jelentősen módosíthatja az algaösszetételt is (Griffits, 1993; Dermott és Munawar, 1993; Heath és mts., 1995; Skubinna és mts., 1995; Horgan és Mills, 1997; Baker és mts., 1998; Bastviken és mts., 1998; Smith és mts., 1998; Strayer és mts., 1999; Naddafi és mts., 2007).
Hatékony filtráció révén feldúsul és helyben marad a kagyló esetlegesen élő algát is tartalmazó anyagcsereterméke, módosítva ezáltal az energiaáramlást a pelágikus zónából a bentikus régióba, dinamikus összeköttetést teremtve a bentikus és a pelágikus zóna között (Reeders és Bij de Vaate, 1990; Fahnenstiel és mts., 1995; Karatayev és mts., 1997; Strayer és mts., 1999). A fécesz és a pszeudofécesz képzésével elősegíti a lebegőanyagok kiülepedését (Ten Winkel és Davids, 1982; Stanczykowska és Planter 1985; Griffits, 1993; Karatayev és mts., 1997, Roditi és mts., 1997; Baker és mts., 2000). Daunys és mts. (2006) által egységnyi felületre kalkulált biodepozíciós ráta a kagylópopuláció által megtelepedett területen nagyobb volt, mint a fizikai ülepedési ráta a kagylókolóniáktól mentes mederfenéken.
A kagyló által kibocsátott fécesz és pszeudofécesz fontos táplálékforrása többek közt a helyhez kötött fenéklakó élőlényeknek (Wolnomiejski, 1969) és fontos szerepet tölt be az itt végbemenő anyagforgalomban (Stanczykowska és mts., 1976; Stanczykowska, 1997;
Stanczykowska és Lewandowski, 1993a). A szén a detritusz átcsoportosítása révén a nyíltvízből a bentikus zónába kerül, gyorsítva a különféle vízi életterek elemei közt lezajló folyamatokat (Carpenter és mts., 1985). Contoy és mts. (2005) szerint az Erie tóban a kagyló ammónium- és foszfátkibocsátása révén megrövidíti a nitrogén és a foszfor „turnover” idejét és ezzel módosítja a nitrogén valamint a foszfor forgalmat. Feltehetően a nitrogén forgalomban fontosabb szerepe van, mint a foszforéban.
A kagyló „top-down” hatását két tényező befolyásolhatja jelentősen, a felkeveredés (a bentoszban az alga eltávolításra lehet hatással) és a mélység, mely leginkább partközeli és sekély területeken lehet jelentős hatás (Edwards és mts., 2005). A kagylók, mint a Dreissena, vagy a Mytilus edulis az összes lebegőanyagból nem szelektálja ki a szerves, vagy ásványi anyagokat,
hanem a lebegőanyag koncentrációjának megemelkedését követően növekelik a filtrációs rátát (Reeders és mts. 1989). A kagyló a fitoplankton biomasszát leginkább a jól felkeveredő torkolati területeken szabályozza (Roditi és mts., 1996; Strayer és mts., 1996; Caraco és mts., 1997; Smith és mts., 1998).
Leach (1993) az Erie-tó nyugati felében vizsgálta a vándorkagyló elterjedése és a vízminőség közötti összefüggést (klorofill-a tartalom és átlátszóság). A kezdeti (1988) 4 lárva l-1 sűrűség 1990-re 96 lárva l-1 értékre nőtt, ezzel párhuzamosan az átlátszóság 85%-kal nőtt, az átlagos klorofill-a koncentráció pedig 43%-kal csökkent a nyugati medencében. Ugyanakkor a nyugati-központi medencében az átlátszóság 52%-kal nőtt, az átlagos klorofill-a koncentráció pedig 27%-kal csökkent. Leach mindezt a megtelepedett vándorkagylóknak tulajdonítja, melyek denzitása elérte a 342.000 ind m-2-t.
Az Erie tó bioenergetikai modellje alapján az ott élő vándorkagylópopuláció a planktonikus elsődleges termelés akár 16-36%-át is képes áthelyezni a bentikus zónába (Madenjian, 1995). A vándorkagyló a Huron tóban és a Saginaw öbölben a klorofill-a 59%-át (Fahnenstiel és mts., 1995), a Hudson folyóban 85% (Caraco és mts., 1997), az Erie tó északi részén pedig akár 90%-át is eltávolítja (Leach, 1993; Nicholls és Hopkins, 1993). A teljes diatóma mennyiség 80%-kal csökkent a halak számára fontos táplálékforrást jelentő vándorkagyló invázióját követően, mely végül egy magasabb trofitási szinthez vezetett (Holland, 1993).
Burnt és munkatársainak az Erie-tó nyugati részén végzett 1993-as kísérlete rámutatott arra, hogy a vándorkagyló naponta akár a víztest 39-96%-át is képes átszűrni (Bunt és mts., 1993), a lebegőanyag ülepítése révén pedig jelentősen hozzájárul a vizek minőségének javulásához. Stoeckmann és Garton (1997) szerint egy kagyló 3,16 kalóriát fogyaszt el naponta, erre alapozott számításaik alapján 10.000-50.000 ind m-2 denzitás mellett egy nyár folyamán a populáció az elsődleges termelés 10-50%-át fogyasztja el. Az Erie tó nyugati részén kolonizált vándorkagyló populáció a teljes szeszton több mint 40%-át képes eltávolítani a víztestből.
Mindebből kifolyólag Reeders és Bij de Vaate (1990, 1992) javasolják a vándorkagyló egyedszámának növelését az eutrofizáció csökkentése céljából.
Összegezve elmondható, hogy a vándorkagyló többféleképpen (direkt és indirekt módon) lehet hatással az alga összetételére. Közvetett módon a fény és a tápanyag módosulása révén - mely kedvez a fitoplanktonnak-, esetleg úgy, hogy olyan nagy sebességgel távolítja el az algát a