I N T R O D U C T O R Y L E C T U R E (Translated)
Ma r c e l Be s s i s
School of Medicine, University of Paris, and Centre National de Transfusion Sanguine
Micro-irradiation of living cells is employed for two main purposes : (i) to analyse the functions of cellular constituents by selectively altering certain of them ; (2) to study the effects of radiation on different parts of the living cell, effects which are very little understood, because they are masked by more general reactions when the radiation is spread over the whole of the cell.
The choice of technique of irradiation depends on the kind of radiation one wishes to use and the nature of the problem being studied. In reviews by Zi r k l e (1957) and Sm i t h (1964) are to be found theoretical treatments from the point of view of physics and of radiobiology as well as descriptions of the effects on cells of the different types of micro-beam, of u.v. and ionizing radiation. When lasers were invented it was natural to use them for cell micro-surgery,
( Be s s i s et al 1962). In this paper we intend to show briefly the tech- nical principles and the principal results obtainable (where one can hope to obtain them) in this form of experiment on the living cell.
ι. i r r a d i a t i o n t e c h n i q u e s
A. U.v. micro-puncture
The earliest experiments on localized u.v. irradiation date from 1912.
Ch a h o t i n used a magnesium spark, a monochromator and a reverse
quartz microscope, which formed a reduced image of rays at 280 nm on the specimen. The cells were observed with an ordinary microscope.
A fluorescent solution allowed one to locate the u.v. spot so that on replacement by the specimen its position was accurately known.
In 1954 and 1957, Ur e t z, Bl o o m and Zi r k l e (1954) and Ur e t z
and Pe r r y (1957) worked with an apparatus in which the u.v. beam
was directed through the objective which also served for observation.
The objective was a mirror system having a focus independent of wavelength. A spot of minimum diameter of 1-2 μ was obtained,
291
292 M I C R O - I R R A D I A T I O N O F C E L L S
adjustable in position beforehand using visible light. Ur e t z and
Pe r r y (1957) coupled this mirror-type objective with an optical system allowing of phase-contrast. A photomultiplier could be substituted for the microscope condenser to measure the intensity of radiation directed on to the cell.
Mo n t g o m e r y and his collaborators (1958) described a micro-
irradiation technique using a 'flying spot'. In this apparatus the utilizable u.v. energy is very feeble.
More recently a number of other arrangements have been put forward ( Be s s i s and No m a r s k i , 1959, i960; Cz i h a k , 1961; Du b r o v ,
1962). The first of these will be described in detail because with it the finest spots are obtainable and excellent phase-contrast images can be observed.
Principle of the apparatus
A special objective, replacing the microscope condenser, projects upon the cell to be irradiated an image of a small aperture illuminated by a u.v. beam. Circular apertures giving images of o-2-ιο μ diameter, or slits (image 0-5 χ ίο μ) are used. This objective needs to be well corrected and to have a very high N. A. if one is to obtain well-localized spots of sufficient intensity. We have replaced the Bausch and Lomb mirror objective used in our first apparatus by a Zeiss Ultrafluar lens objective; aperture 1-25, glycerine immersion, corrected as accurately for the u.v. as for the visible region. The object is observed and cine- photographed by an ordinary phase-contrast microscope. The phase- contrast system consists essentially of a ring source near the lamp and means of forming an image, with the aid of a semi-transparent mirror, on the focal plane before the Ultrafluar condenser-objective. One thus obtains the necessary phase-contrast illumination without restricting the u.v. beam. T o be able to localize the u.v. spot one illuminates the aperture with a minute source coloured green, for example, using a semi-transparent mirror and collector. The semi-transparent mirrors are of thin silica with dielectric layers, completely transparent to the u.v. and reflecting 30 per cent of visible light (see Fig. 1).
The u.v. source is either a hydrogen or deuterium lamp giving a u.v.
continuum, or a rotating electrode cadmium spark, followed by a Cooke monochromator, or sometimes a low-pressure mercury resonance lamp. The best optical conditions are sought to deliver to the object the maximum quantity of energy coupled with the most precise localization and the smallest dimensions of the spot. Thanks
I N T R O D U C T O R Y L E C T U R E 2 9 3
TABLE I
Hole source Spot image Luminance factor
o o 9 ο·2θ
80 o-io 062
120 0-13 ο·8ο
160 ο·ι8 ο·84
In the original article by Bessis and No m a r s k i (i960) there will be found theoretical considerations of the relationship of irradiation energy to spot dimension.
Method of use
Object preparation. The object is placed between a cover slip of silica, (of high-quality u.v. transparency), facing the condenser- objective, and a small glass slip, both about 0-15 mm thick (for the Bausch and Lomb or the Zeiss objectives described above). After sealing with paraffin the preparation is placed (in a metal support) between the objective and condenser-objective with immersion fluid on both sides, oil on the phase-contrast objective side and glycerine on the other.
Adjustment of spot and phase-contrast. The phase contrast is adjusted in the usual way; the hole source being placed at the same distance from the condenser as the field diaphragm, both are centred together.
Irradiation. With the aperture illuminated with visible light and a spot size chosen, the desired point on the specimen is brought over the spot and the u.v. admitted. Irradiation times usually vary between some tenths of seconds and 30 sec. After control of the illumination of the phase-contrast system by photo-cell the effects are cine-photo- graphed.
to the use of a very wide beam it is possible to localize the place where the radiation is absorbed not only horizontally but also in depth ; the beam is convergent with wide angle, and the intensity is relatively small within it except just at the focus. Table 1 gives relative luminance factors for different hole sources, at a wavelength 250 nm and with an objective of 100 χ , Ν.A. 1.
2 9 4 MICRO-IRRADIATION OF CELLS
FIG. Ι. Diagram of the path of rays in the micropuncture apparatus
(BESSIS and NOMARSKI, i 9 6 0 ) .
in the u.v. they may reach ο·2 μ. The potentialities of localization for u.v. irradiation (in contrast to ionizing radiations) depend entirely on the geometrical relationships of the different parts of the cell. For example, one cannot irradiate the cell nucleus without also affecting the cytoplasm lying immediately underneath it; nevertheless one can Dimension and shape of the spots. The practical limits of spot diameter are given by the resolving power of the condenser-objective, and also depend on the wavelength used. For visible light they are about 0-4 μ;
I N T R O D U C T O R Y L E C T U R E 295 concentrate the radiation on a very tiny volume situated within the nucleus.
Circular spots are most frequently used, of diameter 0-2-20 /x.
Other shapes are easily formed, since the hole source size (pierced through black plastic sheet) is 2-20 mm. It is equally possible to protect as well as to irradiate any precise areas by replacing the hole source by a thin silica plate. The phase-contrast microscope is simultaneously in focus on this and on the object. With a pen dipped in Indian ink one can easily outline on the plate the shapes of nucleoli, for example, and then by filling in with ink one can irradiate all the cell except the nucleoli. Alternatively one can irradiate the nucleoli only.
B. Micropuncture by laser
Lasers are of very recent invention and will certainly be subject to great development in the near future. The commercially available laser which has proved to be the most useful for micropuncture is the ruby laser (wavelength 694 nm). Our apparatus is based on that described above for u.v. irradiation (1959-60). Nevertheless, certain novel matters have to be taken into account. In particular the duration of emission from the laser is very short (about 1/1000 sec), and it is difficult to centre the microspot. For this reason we propose the following arrangement (Bessis, Gires, Ma y e r and No m a r s k i , 1962).
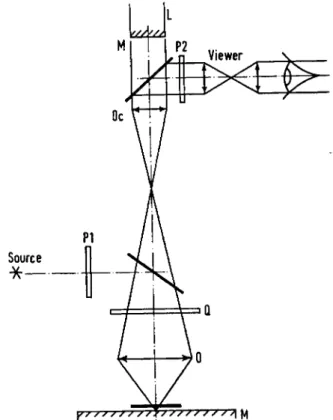
A microscope illuminated by reflected light is used, the object being placed on a dielectric mirror. Centering is effected by observing a second image of the object by autocollimation upon the front face of the laser, the luminosity being regulated by two polarizers and a quarter-wave plate, removable after use. One can then localize the position where the spot will be, thanks to the weak image which can be reflected off the laser front face. Before the irradiation experiment the image of a small test source is made to coincide with the place of the true spot.
For work on a living cell a preparation is made in the usual way and is illuminated by transmitted light with the aid of a condenser and phase-contrast. The useful diameter of lasers used is 3 mm; their length 50 mm. The rear face of the laser cylinder is heavily silvered, but allowing a small fraction of light to pass which can be monitored photo-electrically; the front face is semi-transparent, transmitting 6 per cent. The excitation is given by a Xenon flash lamp of spiral silica tube, and fed by a capacity of 160 μ¥ charged to 2500 V.
2q6 m i c r o - i r r a d i a t i o n o f c e l l s
The laser produces a light spot 5 mm in diameter at a 1 m distance.
The beam is closely parallel, with a divergence of about 5 χ ί ο- 3. With the diameter of the laser used smaller than the pupil of a microscope objective, it is of interest to set up the laser before the eyepiece of a
FIG. 2. Diagram of the path of rays in the laser illuminated microscope
(BESSIS et al, 1962). L, Laser; M , half-silvered surface; Oc, ocular;
Ο, objective ; M , object carrier ; Q, J wave plate ; Pj and P2, polarizers.
microscope. Here the beam is limited by the eyepiece ring. We have used an eyepiece of 6 χ and an objective of 100 χ , which diaphragms the laser one-third; the calculated spot diameter is then 2-5 μ.
T o avoid dangers at the eyepiece exit due to the violence of the light emission two precautions are taken ; special light filters absorbing in the red are used in the eyepiece, and the effect of micropuncture is observed on a television screen. (Vidicon, again protected by a selec- tive light-absorbing screen.)
INTRODUCTORY LECTURE 297 The use of a laser appears to have very great advantages over other
methods of micropuncture. These include (1) the energy is delivered in an exceedingly short space of time (1/1000 sec for ordinary lasers, perhaps dropping to one ten-millionth of a second for new types).
Under these conditions the irradiated points reach extremely high temperatures. All substances absorbing at the laser wavelength are destroyed, and those alone. Recent technical developments give promise of lasers of wavelengths reaching from the infra-red to the u.v.
Use of the apparatus
The required part of the cell is brought into position over the test spot by methods similar to those described for u.v. irradiation above, and then the laser is discharged. The destruction of the irradiated spot (if it absorbs the laser ray) is observed on the television screen. The diameter of the spot is adjusted by lenses of suitable power, placed just before the eyepiece. The intensity of irradiation can be regulated partly by changing the power of the Xenon flash and partly by a small absorber situated by the front face of the laser.
Preparation of objects
The preparations are made as described above for u.v. work. In cells apparently colourless, as white corpuscles, the laser produces no effect. By contrast, if the corpuscles are stained by certain dyes absorbing at the laser wavelength, such as Janus Green or Neutral Red, one observes the cell to burst on laser discharge. By using weak discharges and selective stains which fix selectively on cell structures one can destroy these alone. Similarly one can use ingestible pigments like lamp black or Trypan Blue, or natural colourants such as chloro- phyll in chloroplasts. It is interesting to note the effects observed on cells where the absorption of the laser wave length appears negligible, as for red blood corpuscles. Where touched by laser action coagulation occurs. With the lower intensities one can see the round trace left by the spot, while, after some minutes, one observes the haemolysis of the corpuscle, which bleaches progressively except at the spot, where the haemoglobin coagulates.
II. TECHNIQUES FOR EVALUATION
The results of micropuncture can be evaluated in three principal ways : one can study the successive changes by time lapse cinemicrography ; one can mark the cell very carefully, embed it, cut it, examine it
298 M I C R O - I R R A D I A T I O N O F C E L L S
with the electron microscope ( Bl o o m , i960; Si l v e s t r e et al, 1961);
and, lastly, above all one can evaluate the metabolic disturbances in the cell by study of changes in the incorporation of labelled material, revealed by autoradiography ( Pe r r y et al, 1961).
The advantages of micro-irradiation by laser compared with u.v.
are the following: the irradiation is of extremely short period and allows rapidly moving cells to be treated ; the intensity is very great, so that all absorbing material is selectively destroyed; the lowest concentration of absorbing substance, invisible as colour to the eye, is enough to produce a considerable effect and to lead to selective destruction of some of the cell contents. U.v. irradiation allows the more or less selective destruction of nucleic acids (using the wave- length 265 nm), or to coagulate the proteins (using 243 nm, Be s s i s and
No m a r s k i , i960). With the laser one can use a very large spot covering the whole of the cell surface after having first coloured the cell con- stituents with specific stains. The first results obtained by us relate to the staining of mitochondria by Janus Green ( Bessis and St o r b, in press) and the vacuoles of Golgi structures by Nile Blue. Here is a general technique capable of very great developments, and it is prob- ably the principal advantage of the microbeam using laser emission.
A. Technique of flux measurement at the irradiated surface The quantitative measurement of the absolute flux passing through a tiny surface, as in micro-irradiation, is fraught with considerable difficulty. The calculation of the dosage of radiation absorbed by the part of the cell irradiated is still more elusive. Comparison by two photocells suitably placed to compare the flux before and after passage through the cell has been suggested. It has also been proposed to use a biological unit, as, for example, the time for haemolysis of a human red blood corpuscle under the particular conditions. A better biological unit would be the inactivation of phages, because they are very small compared with the spot size, and also because this method is already used in macroscopic work (v o n Bo r s t e l , 1962). The inactivation of phages should be measured using the technique of micro-drops, which is a difficult manipulation, particularly when one uses very small spots and immersion objectives.
B. Dynamic studies of successive changes after micropuncture This work is carried out with the phase-contrast microscope, with the aid of speeded-up cinematographic registration. It is difficult to observe
I N T R O D U C T O R Y L E C T U R E 299
a cell radiated with a very small spot for longer than 3-6 h; after this period the cell dies spontaneously, since the narrow confinement required by the short focal lengths of the objectives prevents proper cell development. On the contrary, with much larger spots it is relatively easy to observe the effects of irradiation of tissue cultures for several consecutive days. This technique allows one to examine changes of cell motions, intra-cell movements, alterations of mitosis (BLOOM et al> 1955), pathological morphology of cell structures and the different forms of agony and of cell death.
C. Electron-microscope studies
T o observe an irradiated cell under an electron-microscope is very difficult. The collection of the cell, its fixation, its embedding in various plastics, its sectioning and identification under the electron- microscope, all present problems, and although a matter of great interest little work has been done and no sure and easy solution reached.
We describe here the main lines of the technique used by SILVESTRE
et al (1961), inspired by that of BLOOM (1960a).
Mounting of the specimen
It is necessary that the cells adhere to the cover-glass, covered or not with formvar. Centering: For experiment the object must be centered in the microscope field; this is facilitated by a grating eye-piece.
Marking: This is necessary to find the object again, and is carried out by making a circular trace with a diamond (mounted on the revolving piece) on the upper surface of the cover-glass. Fixation and pre- embedding: For fixing, the cover-glass, separated from its support, is placed, specimen downwards, in a 1 per cent solution of osmic acid for 30-45 min. A pre-embedding treatment is then required; the glass is placed successively in baths of alcohol, methacrylate and catalyst identical with that used for the final embedding. The glass is then placed on the special jig for embedding and the object is centered on this support. Embedding: This uses a jig consisting simply of a metal piece, screwed into the tube and carrying a truncated cone of rhodoid (plexiglass or metal cannot be used as methacrylate adheres in a way impossible to remove). One places on a plate a little cylinder of gelatin shaped as a capsule. All that is necessary is to place the tube fitment over the cylinder to obtain automatic centering. It is lifted off gently and the cylinder filled with semi-polymerized methacrylate of a pasty consistency. The jig is lowered again for final centering so that it is
300 M I C R O - I R R A D I A T I O N O F C E L L S
immersed in the plastic without allowing it to overflow. The container is put in an oven for 12-15 h at 6o° C . When polymerization is complete the cover-glass is solidly attached to the plastic block. T o detach it without disturbing the embedded material a small piece of solid carbon dioxide is placed on it for about 1 min. Shaping to a pyramid: T o give the plastic block the pyramidal form necessary for the microtome a special precision shaping machine is used which produces a pyramid with axis coincident with that of the support. The object will then be found within the small square forming the pyramid tip. It remains only to place it on the microtome and take sections precisely parallel to the truncated pyramid tip. This operation is delicate, for the object is attacked immediately by the knife.
D. Evaluation of metabolic disturbances in the cell
These are evaluated above all by autoradiography. Labelled pre- cursors (thymidine, uridine, amino-acids, etc.) are introduced into the culture ; neighbouring non-irradiated cells serve as standards, and one notes for the radiated cell the absence of fixation or an abnormal localization of a labelled substance. One can thus study the effects of localized irradiations on the synthesis of D N A , R N A or of proteins.
When the irradiated cell is large enough micro-chemical methods may be used, as, for example, a study of oxygen consumption ( Sk r e b and
Sk r e b , i960). It has also been proposed ( Cr u i c k s h a n k , 1962) to use the technique of fluorescent anti-bodies. For example, after irradiation of the nucleus of a cell infected by a virus one can evaluate the effect upon the quantity of virus formed by using a fluorescent anti-viral anti-body.
E. Effect of cell changes upon the environment
We have used speeded-up microcinematography to study the effects of an irradiated cell upon its neighbours. These effects have been described as necrotaxis and necrophobia. Micro-irradiation can also be used to cause a cell to burst and distribute its contents, whose effects on the environment can then be examined. If, for example, a cell contains an anti-body, one can, by making it burst artificially, agglutinate or lyse neighbouring cells containing the corresponding antigen. One can thus demonstrate the presence of a specific anti-body in a given cell. The same trial can be made for any other product with the condition of having round the cell a suitable substrate (histamine, héparine, enzymes, etc.).
I N T R O D U C T O R Y L E C T U R E 3OI
I I I . P R I N C I P A L O B S E R V E D R E S U L T S
A. Irradiation of the nucleus at rest
Ur e t z et al (1954) and Zi r k l e et al were the first to observe that irradiation of a small part of a cell nucleus by u.v. caused in a few minutes loss of contrast in the zone, which became pale when examined under phase-contrast. The zone absorbed less u.v. at 265 nm. Pe r r y
(1957) studied this effect, after irradiating the nucleus with hetero- chromatic u.v. at an intensity of 2 ergs/μΑ Irradiation of a surface 50 μ2 in area showed a very clear diminution of absorption. With a larger surface the effect is less, even though the energy per unit of surface remains the same. The diminution of absorption is constant within 10 per cent for 240, 260 and 280 nm. The phenomenon of paling is also shown when chromosomes are irradiated. Zi r k l e and
Ur e t z (i960) used a microbeam 8 μ in diameter to study the wave-
length differences for the intensity of paling of chromosomes. They obtained the following results :
Wave length, nm Relative energy 225 220 240 30 250 10 260 16 270 19 280 10 3ΟΟ 2*2 Their results suggest that paling is due to the destruction of nucleo- tides and of proteins containing tyrosine, tryptophan or both.
Bl o o m (i960) and Bl o o m and Le i d e r (1962) carefully examined the modifications associated with paling, using optical and electron microscopy. They confirmed that the refractive index and the absorp- tion at 260 nm diminished in the irradiated zone; the zone became negative to the Feulgen reaction and to staining by Methyl Green.
It also became negative to the colour reaction of Alfert Geschwind for basic proteins. These results, associated with those of Pe r r y, show that 50-60 per cent of the D N A disappears after irradiation under the conditions of experiment. For this reason the authors propose to replace the term paling by that of D N A steris.
Electron-microscopical examination of irradiated cells has shown that the A constituent of chromosomes (corresponding probably to
302 m i c r o - i r r a d i a t i o n o f c e l l s
fibres of D N A ) had strongly diminished or disappeared, while the Β constituent (network) remained practically unchanged. The authors insist that fixation must be made in neutral formol, followed by staining with phospho-tungstic acid. Permanganate or osmic acid fixation does not give such good results. Am e n t a (1961) stained chromosomes with D N A steris by the Gomori technique for alkaline phosphatase. He observed no difference between the pale zone and the remainder of the chromosome. He concluded that the amount of alka- line phosphatase seemed independent of the presence or absence of D N A . However, he did not use the photometric technique.
Synthesis of DNA and RNA
Comparative studies between cells with nuclei and those which have been deprived (not needing a true microbeam) have been made on certain material, of which the shape (Amoeba) or the anatomy (Acetabolaria) readily lend themselves for examination. One half of the cell is protected by an aluminium screen. These results have been confirmed by the use of much smaller beams. Er r e r a and Va n d e r - h a e g h e (1957) noted that irradiation greatly affects the survival and regeneration of fragments containing no nucleus. Sk r e b and Er r e r a
(1957) and Sk r e b and Sk r e b (i960) studied the action of u.v. on nucleolated and denucleolated fragments of Amoeba. Irradiation caused a fall in basophilia, both in the nucleus and in the cytoplasm, and a fall in respiration of 50 per cent for denucleolated fragments compared to the normal. They found photo-recovery by visible light equal in the two cases.
Br ä c h e t and Ol s z e w s k a (i960) showed for Acetabularia that the nucleus (or the part of the cell containing it) controlled the synthesis of R N A and of sulphur proteins. Bo r s t e l and Wo l f f (1955) studied the effect of u.v. on the haploid egg of Habrobracon. This measures about 600 μ by 150 μ. The easily seen nucleus is in the front part.
Irradiation of the part containing the nucleus is statistically much more effective. Exposure to visible light induces a photorecovery after nuclear irradiation, but not after irradiation of the cytoplasm alone.
Ja g g e r and Pr e s c o t t (1962) irradiated Amoeba Proteus by poly- chromatic u.v. and observed that it needed sixty times more energy upon the cytoplasm as upon the nucleus to obtain the same effect;
either to retard mitosis, or to kill the cell. If the cell was wholly irradiated much feebler dosages were sufficient to produce the same effects. Photorecovery by light of 330-440 nm could be observed,
I N T R O D U C T O R Y L E C T U R E 303
whether the nucleus or cytoplasm had been irradiated. Synthesis of R N A (measured by tritium containing uridine incorporation) was stopped for periods of 1-22 h by irradiation of the nucleus, but not by irradiation of the cytoplasm. No photorecovery of R N A synthesis was observed. De n d y (1962) irradiated tissue cultures of embryonic or adult fibroblasts with heterochromatic microbeams of u.v. of 3-5 μ diameter in order to see the effect on the yield of R N A synthesis, determined by autoradiography, by counting the number of grains situated above the nucleus of irradiated cells and of fifty neighbouring ones. The irradiated sites were (1) the nucleolus, (2) the nucleus, (3) the cytoplasm, and (4) the liquid around the cell. Estimation of D N A synthesis was made at times 0-2 to 6 h after irradiation. For case (4) above, there was little significant inhibition, with a return to normal after 6 h. For cases (1), (2) and (3) a maximum inhibition at 6 h was found, with no notable difference between them. It was surprising that irradiation of the cytoplasm equally inhibited D N A synthesis, and the authors speculated on the possibility of the production of a nuclear toxin by radiation of the nutritive liquid.
B. Irradiation of the nucleolus
Ga u l d e n and Pe r r y (1958) showed that irradiation for 3 sec of a single nucleolus in a neuroblast of the grasshopper with a u.v. microspot of high intensity, from the telophase to the middle of the prophase, stops mitosis. After the middle of the prophase, the cell becomes insensitive to irradiation. Irradiation of a non-nucleolus region slows up but does not stop mitosis. Er r e r a, Ha l l and Pe r r y (1958) showed by auto- radiography that the uptake of amino acids begins at the same time and in the same manner in the nucleolus, the nucleus and the cyto- plasm. If the nucleolus is irradiated so that cytidine is no longer taken up, the take-up of amino acids by the nucleus remains unchanged, but it decreases 30 per cent both for the nucleolus and for the cytoplasm ; from which they conclude that the nucleolus controls the metabolism of certain proteins of the cytoplasm. Pe r r y, Me l and Er r e r a (1961) returned to this work. They observed that irradiation of the nucleolus of Hela cells in tissue culture affected the quantity of R N A in the cell.
Their conclusion is that the nucleolus controls the uptake of two-thirds of the R N A of the cytoplasm and of one-third of the R N A of the nucleus. The greater part of the R N A of the cytoplasm seemed to be synthesized in the nucleolus. Mo n t g o m e r y and Hu n d l e y (1961) used a flying spot to irradiate nucleoli of cells in tissue cultures for very long
304 M I C R O - I R R A D I A T I O N O F C E L L S
periods, up to 12 h. They noticed that after 4-6 h the u.v. absorption of the nucleoli diminished, after which it remained constant. The cell neither died nor showed any change in its movements or behaviour.
They gave no explanation of this kind of paling of nucleoli. Ca r l s o n
et al (1961) and Ca r l s o n (1961) used u.v. at 254 and 280 nm, and
Ga u l d e n (i960) using heterochromatic u.v. also observed a retarda- tion of mitosis when irradiation was made between the telophase and the prophase.
C. Irradiation of the mitotic apparatus
The same paling observed for the nucleus in interphase, as described above, is also found for chromosomes. The time to produce the same effect, however, seems much shorter for the latter ( Zi r k l e, Bl o o m
and Ur e t z , 1958). When one segment of a chromosome is irradiated the paling progresses from the irradiated part in both directions for 20-30 min until the length affected is two or three times as long as the irradiated spot. This paling is observed at all mitotic stages from prophase to the anaphase, with almost equal exposure times. Ur e t z , Bl o o m and Zi r k l e (1954) observed that the irradiation of centro- meres prevented the coherent movements of the chromosomes ; they no longer moved towards the equatorial plane. Nevertheless, if a chromosome was near a daughter nucleus it was re-incorporated in the re-constructed nucleus ; if it was far off it formed an accessory nucleus on its own, a micronucleus, which could, however, become a lobe of a principal nucleus. Bl o o m, Zi r k l e and Ur e t z (1955) observed that when several chromosomes in metaphase were exposed to microbeams bridges were formed between two irradiated parts and the two chromosomes became adhesed. The nuclear division then took place in any asymmetric manner. Ur e t z and Zi r k l e (1958) studied the relative energy at different wavelengths necessary to obtain both paling and adhesion of chromosomes from heart cultures of Triton, with a 8 μ spot. The two effects were found to be linked. Zi r k l e , Bl o o m and Ur e t z (1958) noted that the adhesion was localized at the irradiated points and that the effect was a direct action. Ur e t z and Zi r k l e, in 1955, observed the spindle body in polarized light, because it is strongly biréfringent in the blastomeres of sea-urchin eggs. After irradiation of the cytoplasm of one of the two blastomeres (the other serving as a control) the spindle immediately disappeared. When the blastomer was irradiated before the appearance of the spindle the latter never developed. The spindle of Triton heart cells in culture
I N T R O D U C T O R Y L E C T U R E 305
could be made to disappear after irradiation with heterochromatic u.v., even if the irradiated place was 30 μ away. These authors think that some poison is formed photochemically from the spindle. Spindle destruction is followed by very anomalous movements of the chromo- somes. They lose their usual configuration in metaphase and form a rosette whose centromeres are directed towards an attractive region different from the normal one. There follows a false anaphase, in which the undivided chromosomes separate into two rosettes; then the cytoplasm divides normally. The daughter cells contain, then, two groups of complete chromosomes instead of two groups of half chromosomes. Fragments of chromosomes can also be seen. Zi r k l e , Ur e t z and Ha y n e s ( 1960) also studied the disappearance of the spindle and of phragmoblastes after irradiation of cytoplasm of plant cells.
D. Irradiation of cytoplasm
As early as 1921, Ch a h o t i n had shown the susceptibility of the cell surface to irradiation. On sea-urchin eggs an oedema is obtained, localized at the spot level, which are often resorbed by the cell;
if the radiation is stronger the cell surface bursts. Ur e t z and Zi r k l e
(1955) observed the same localized oedema on sea-urchin eggs.
Ch a h o t i n (1935,1936) showed that many protozoa, after irradiation, threw out a part of their cytoplasmic contents. Sometimes they formed external protuberances, which separated after a time from the body of the cell. One could have up to seven expulsions of cytoplasm reducing the volume of the cell before final cytolysis. Ch a h o t i n (1935) also observed modifications of movement within and external to the cell. With Amoeba a cessation of protrusion of pseudopods and of cytoplasmic cyclosis was observed. The changed portion of the cytoplasm, coagulated or liquified, is consumed by the remainder of the cell. Te r n i (1933), using Ch a h o t i n ' s microbeams on urodele spermatozoa, found that the tail movements were not affected by irradiation of the head or neck, but that irradiation of the flagella caused immediate cessation of movement of the fragment treated.
This spermatozoan has an undulant membrane which permits easy observation. Ch a h o t i n (1936) observed that the peduncle of Vorticella contracted violently on irradiation and was able to find the most sensitive region. He also observed that radiating the contractile vacuole caused it to cease pulsating. With Euglena his experiments led him to believe that the small red body called stigma seemed to play a part in phototaxis. When the stigma was irradiated phototaxis had
20
3θ6 M I C R O - I R R A D I A T I O N O F C E L L S
disappeared. Ch a h o t i n (1938) has even described the production of conditioned reflexes in irradiated infusoria. Effects on the nuclei after irradiation of the cytoplasm have been described by several workers
( Ch a h o t i n , 1920; Er r e r a and Va n d e r h a e g h e , 1957; Bl o o m et al,
1955). Often the nucleus shows marked changes, vacuolization and increase of basophilia. Am e n t a (1962) irradiated polynuclear eosino- phils. Radiation of the granules caused their disappearance followed after a short delay by pycnosis of the nucleus. He postulated the liberation of a substance toxic to the nucleus. Ya s h i m a and Iz u t s u
(i960) studied the effects of u.v. microbeams on Triton eosinophil centrosomes. When the centrosome was irradiated its movements stopped and the cell became spherical. When they irradiated a small part of the cytoplasm they observed a change of direction of the cell towards the injured part. Be s s i s and No m a r s k i (i960) showed that, at a wavelength of 243 nm (Cd-arc) one could obtain in 5 sec haemolysis of a red blood corpuscle. With powerful doses a localized coagulation of haemoglobin preceding haemolysis occurred. The same effect can be seen when a ruby laser is used, which seems surprising in view of the long wavelength of the radiation ( Bl o o m et al, 1962). Be s s i s et al (1964) studied the conditions of irradiation of different parts of the cytoplasm by a laser, as well as the theoretical and practical effects of organic stains. Weak staining by Janus Green, which colours the mitochondria only, enables selective destruction of these to be achieved. Other stains allow of action on the nucleus and on different parts of the Golgi apparatus.
E. Different types of cell death
Be s s i s (1964a) studied aspects of the agony and death of cells after irradiation by different u.v. wavelengths. Varied aspects were found, depending on the point of application and the wavelength. Coagulation of the protoplasm, for example, occurs at 254 nm, while at 258 and 270 nm the nucleic acids are affected. Depending on the place of application death can take place by rigidification, oedema, nucleolysis, etc.
F. Effect of cell death on the environment
Micropuncture allows the killing of one cell among its fellows; by using speeded-up cinematography one can follow the effect of this death upon the environment. Thus one can study a phenomenon named necrotaxis ( Bessis, 1964). Certain cells are attracted towards
FIG. 3 . U . v . i r r a d i a t i o n of the n u c l e o l u s of a l y m p h o c y t e i n c u l t u r e . (a) U n d e r p h a s e - c o n t r a s t ; note the large d a r k n u c l e o l u s , (b) T h r e e spots of different d i a m e t e r s of v i s i b l e l i g h t are f o r m e d o n the left of the c e l l , (c) O n e of these s p o t s is c e n t e r e d o n the n u c l e o l u s a n d the r a d i a t i o n c h a n g e d to u . v . for a b o u t 15-sec e x p o s u r e , (d) A f t e r 15 m i n ; the n u c l e o l u s h a s d i s a p p e a r e d .
FACING P. JO 6
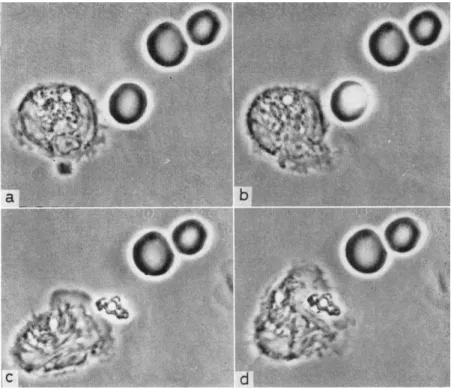
FIG. 4 . E f f e c t of laser i r r a d i a t i o n o n a red b l o o d c o r p u s c l e a n d p h a g o - cytose b y a l e u c o c y t e , (a) L e u c o c y t e at centre, three red c o r p u s c l e s o n the r i g h t , (b) T h e spot is c e n t e r e d o n one of the c o r p u s c l e s a n d the laser is o p e r a t e d , (c) T h e red c o r p u s c l e is d e s t r o y e d a n d a part of the h a e m o g l o b i n is left t o w a r d s w h i c h the l e u c o c y t e m o v e s , (d) T h e l e u c o c y t e ingests the r e m a i n s of the r e d c o r p u s c l e .
I N T R O D U C T O R Y L E C T U R E
the dying ones. In human blood, for example, when a white or red corpuscle is irradiated, as soon as the cell shows signs of change neighbouring cells move towards it, and, in a few minutes, one obtains what the anatomo-pathologists call a 'rosette'. These cells consume the injured cell, usually by emptying it, each cell carrying away a part of its prey; in a few minutes only the cell skeleton is left. By means of speeded-up films it becomes evident that it is only after the cell has undergone plasmolysis, allowing its cell contents to escape, that the neighbouring cells move in, often from some distance (500 μ). One can see polynuclear new-comers violently throw aside those rosette forming cells consuming the corpse in order to obtain their share of the feast. Sometimes it can be seen that the polynuclears, after having consumed some of the dead cell, leave the part, which then no longer attracts further cells, even those which by chance pass close by; it seems that here the necrotactic substances are exhausted. It would be interesting to identify these substances and to discover if they are specific for particular cells. Many other effects on the environment such as liberation of specific anti-bodies may be studied by this technique (Bessis, 1964).
In the course of these experiments we have also observed a pheno- menon of reverse movement, which is a form of negative Chemotaxis.
When certain individuals of a culture of Euglena are irradiated, within about 1 min all the others spread away from the killed cells ( B e s s i s and B u r t e , 1964b). This technique allows one to study certain aspects of cell ecology.
R E V I E W S
Ra y n a u d A. (1963) In Techniques de laboratoire (Loiseleur J., ed), Vol. 2, p. 571, Masson, Paris.
Sm i t h C.L. (1964) Inter. Rev. Cytol. 1 6 , i33~53, Acad. Press, New York.
Zi r k l e R . E . (1957) Adv. Biol. Med. Physics 5, 103-46.
R E F E R E N C E S Am e n t a F . S . (1961) Stain Tech. 3 6 , 15-19.
Am e n t a F . S . (1962) Anat. Ree. 1 4 2 , 81-8.
Bessis M. (1964a) In Ciba Foundation Symposium on Cellular Injuries, I vol., Churchill, London.
Bessis M . , Gi r e s F . , Ma y e r G . and No m a r s k i G . (1962) C.R. Acad. Sei., Paris 2 5 5 , 1010-12.
Bessis M. and Bu r t e Β. (1964b) Necrotaxis Texas Report, Biol, and Med.
Bessis M. and No m a r s k i G. (1959) C.R. Acad. Set., Paris 2 4 9 , 766-8.
Bessis M . and No m a r s k i G. (i960) J. Biophys. Biochem. Cytol. 8, 777-91.
3°8 MICRO-IRRADIATION OF CELLS
Bessis M. and Te r p o g o s s i a n M . (1964c) Micropuncture of cells by LASER microbeam, Ν. Y. Acad, of Science (in press).
Bl o o m W . (1960a) J. Biophys. Biochem. Cytol. 7 , 191.
Bl o o m W . (1960b) Science, 1 3 1 , 1316.
Bl o o m W . and Le i d e r R J . (1962) J. cell. Biol. 1 3 , 269-301.
Bl o o m W . , Zi r k l e R.E. and Ur e t z R.B. (1955) Ann. N.Y. Acad. Sei. 5 9 ,
503-13.
Bo r s t e l R . C .v o n (1962) Cited by Smith C.L. in Nature, Lond. 1 9 6 , 728-9.
Bo r s t e l R . C .v o n and Wo l f f S. (1955) Proc. nat. Acad. Sei., Wash. 4 1 , 1004-9.
Br ä c h e t J. and Ol s z e w s k a M.J. (i960) Nature, Lond. 1 8 7 , 954-5.
Ca r l s o n J.G. (1961) Ann. N.Y. Acad. Sei. 9 5 , 932-41.
Ca r l s o n J. G . , Ga u l d e n M.E. and Ja g g e r J. (1961) In Progress in Photo-
biology, Proc. of the 3rd International Congress on Photobiology. Elsevier, Amsterdam, London, New York, Princeton.
Ch a h o t i n S. (1912) Biol. Zbl. 3 2 , 623.
Ch a h o t i n S. (1920) C.R. Soc. Biol., Paris 8 3 , 1593-5.
Ch a h o t i n S. (1935) C.R. Acad. Set., Paris 2 0 0 , 26, 2217-19.
Ch a h o t i n S. (1936) C.R. Acad. Sei., Paris 2 0 2 , 12, 1114-16.
Ch a h o t i n S. (1936) Ann. Protist., Paris 5, 1-55.
Ch a h o t i n S. (1938) Arch. Inst, prophyl. 1 0 , 119-33.
Ch a h o t i n S. (1961) C.R. Soc. Biol., Paris 8 4 , 464-6.
Cr u i c k s h a n k J.G. Cited by Smith C.L. (1962) Nature, Lond. 1 9 6 , 728.
Cz i h a k G . (1961) Zeiss 6 Mitteil. 2 , 165-72.
De n d y P.P. (1962) Cited by Smith C.L. (1964) In Proc. of Int. Cong. Rad.
Res., Harrogate (in press).
Du b r o v A.P. (1962) Biophys. Moscou 2 4 , VIII, 634.
Er r e r a M., Ha l l A. and Pe r r y R.P. (1958) Biochem. Biophys. Acta 4 9 , 58-63.
Er r e r a M. and Va n d e r h a e g h e F . (1957) Exp. Cell Res. 1 3 , 1.
Ga u l d e n M.E. (i960) In The Cell Nucleus, vol. I, pp. 15-17, Mitchell,
Butterworth & Co. London.
Ga u l d e n M.E. and Pe r r y R.P. (1958) Proc. nat. Acad. Set., Wash. 4 4 , 553-9.
Ja g g e r J. and Pr e s c o t t D. M . (1962) Cited by Smith C.L. in Microbeam Conf. Aug. 2, Cambridge.
Mo n t g o m e r y P.O'B. and Bo n n e r W.A. (1958) A.M.A. Pathol. 6 6 , 418-21.
Mo n t g o m e r y P.O'B. and Hu n d l e y L.L. (1961) Exp. Cell Res. 2 4 , 1-5.
Pe r r y R.P. (1957) Exp. Cell. Res. 1 2 , 546.
Pe r r y R.P. (i960) Exp. Cell. Res. 2 0 , 216-20.
Pe r r y R.P., Me l A. and Er r e r a A. (1961) Biophys. Biochem. Acta 4 9 , 47-57.
Si l v e s t r e J., Bu r t e B. and Ro u s s e a u R. (1961) Bull. Mic. Appliquée 1 1 ,
109-15.
Sk r e b Y. and Er r e r a M. (1957) Exp. Cell Res. 1 2 , 649-56.
Sk r e b Y. and Sk r e b N. (i960) Biochim. Biophys. Acta 3 9 , 540-1.
Sm i t h C.L. (1962) Nature, Lond. 1 9 6 , 728-9.
Te r n i T . (1933) Bull, de Γ Ass. des Anatomistes, C R . de Γ Ass. des Anato- mistes, 28 ème réunion, pp. 651-4.
INTRODUCTORY LECTURE
Ur e t z R . , Bl o o m W . and Zi r k l e R . E . (1954) Science, 120, 197-9.
Ur e t z R . B . and Pe r r y R . P . (1957) Rev. sei. Instum. 28, 861-6.
Ur e t z R . B . and Zi r k l e R . E . (1955) Biol. Bull. 1 0 9 , 370.
Ur e t z R . B . and Zi r k l e R . E . (1958) Radiation Res. 9 , 296.
Ya s h i m a Y . and Iz u t s u K. (i960) Mie Med.J. 1 0 , 223-38.
Zi r k l e R . E . , Bl o o m W . and Ur e t z R . B . (1958) Radiation Res. 9 , 319.
Zi r k l e R . E . , Bl o o m W . and Ur e t z R . B . (1956) In Proc. Int. Conf. Peaceful uses of Atomic Energy, Vol. II, p. 273, Genève. Cited by Zirkle.
Zi r k l e R . E . and Ur e t z R . B . (i960) Science 1 3 1 , 1316.
Zi r k l e R . E . , Ur e t z R . B . and Ha y n e s R . H . (i960) Ann. N.Y. Acad. Sei.
9 0 , 435-9.