FODORPATAKI LÁSZLÓ SZIGYÁRTÓ LÍDIA BARTHA CSABA
NÖVÉNYTANI ISMERETEK
KÖRNYEZETTUDOMÁNYI TANSZÉK
FODORPATAKI LÁSZLÓ SZIGYÁRTÓ LÍDIA BARTHA CSABA
NÖVÉNYTANI ISMERETEK
Scientia Kiadó Kolozsvár · 2022
Első magyar nyelvű kiadás: 2009 Második, változatlan kiadás: 2022
© Scientia 2022
Minden jog fenntartva, beleértve a sokszorosítás, a nyilvános előadás, a rádió- és televízióadás, valamint a fordítás jogát, az egyes fejezeteket illetően is.
ISBN: 978-606-975-061-2 Lektor:
Dr. Nagy-Tóth Ferenc (Kolozsvár)
Felelős kiadó:
Sorbán Angella
Kiadói koordinátor:
Szabó Beáta
Sorozatborító:
Miklósi Dénes
TARTALOM
Elõszó 9
1. A növénytani tudományágak helye
a biológiai tudományok rendszerében (B. Cs.) 11 2. A növényi életforma jellemvonásai (F. L.) 13 3. A növények helye
az élõvilág törzsfejlõdéstani rendszerében
(Sz. L.) 17
4. Rendszertani kategóriák (B. Cs.) 23
5. A növényi test szervezõdési formái,
szimmetriaviszonyai és elágazási típusai (F. L.) 26 6. Analóg, homológ és metamorfozált növényi testrészek (F. L.) 30 7. A növényi sejt sajátos alkotói (F. L.) 32 8. Növényi életmódú egysejtûek és teleptestûek (F. L.) 38 8.1. Az algák és szerepük a vízi ökológiai rendszerek primer
produkciójában, valamint a vízminõség monitorizálásában 38 8.2. Zuzmók és mohák, ezek szerepe a szárazföldi környezet
minõségének bioindikációjában 44
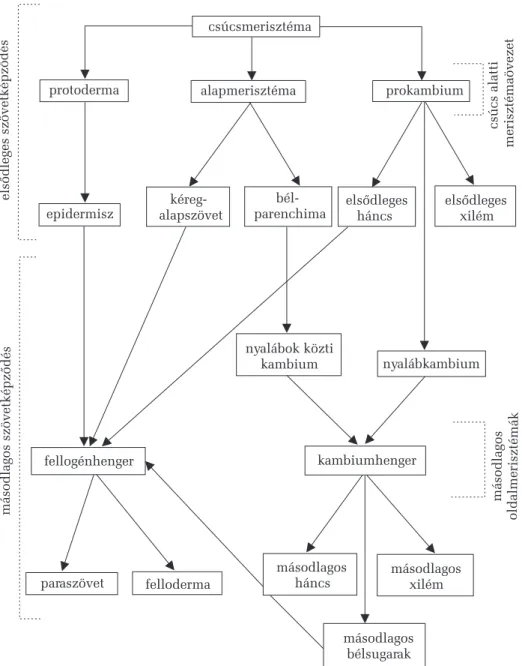
9. A hajtásos növények (Cormophyta) testszervezõdése (F. L.) 48 9.1. A szövetképzõ differenciálódás
és megnyilvánulási módozatai 48
9.2. A növényi szövetek osztályozása 52
9.2.1. A merisztémák (osztódó szövetek) 53
9.2.2. Védõszövetek 59
9.2.3. Felületi felszívó szöveti képzõdmények 65
9.2.4. Alapszövetek vagy parenchimák 67
9.2.5. Szilárdító- vagy támasztószövetek 72
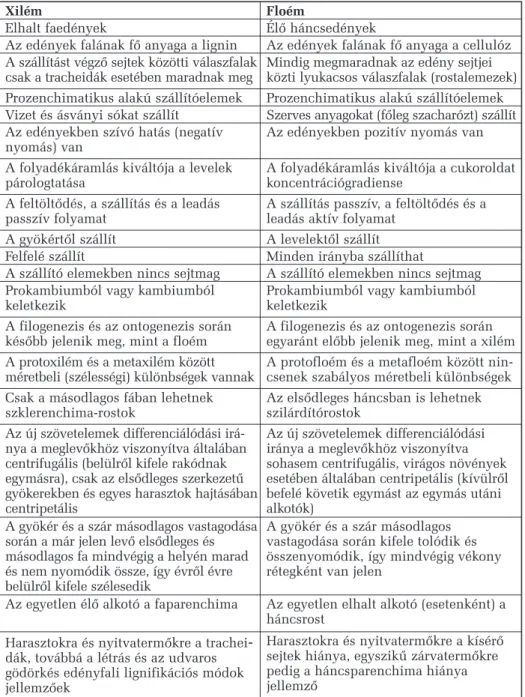
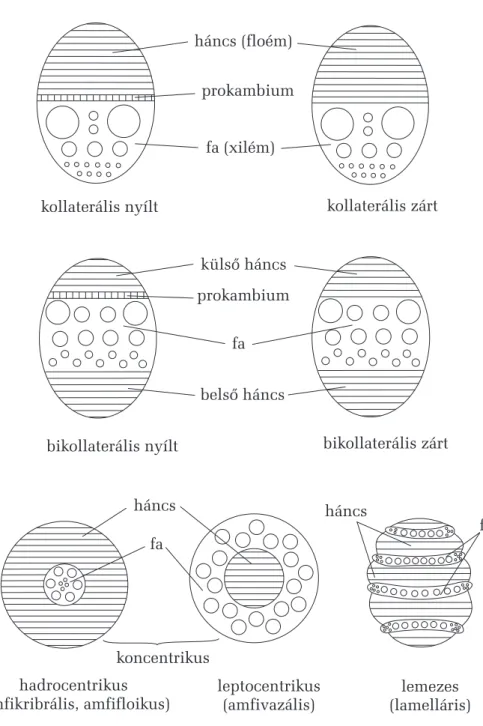
9.2.6. A szállítószövetek rendszere 74
9.2.7. A szállítónyalábok típusai 79
9.2.8. Szekréciós szöveti képzõdmények 83 9.2.9. Érzõ vagy ingerfelfogó szöveti képzõdmények 88 9.3. A hajtásos növények szervei (organográfia) 90
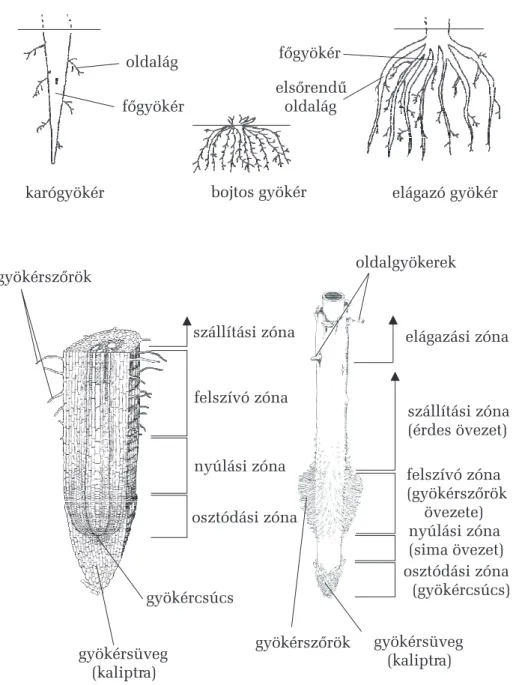
9.3.1. A gyökér (radix, rhyza) 92
9.3.1.1. Módosult gyökértípusok 93
9.3.1.2. A gyökércsúcs szervezõdése 97 9.3.1.3. A gyökér elsõdleges szerkezetének
általános jellemzõi 100
9.3.1.4. Az oldalgyökerek endogén eredete 102 9.3.1.5. A gyökér másodlagos szerkezete 103
9.3.2. A szár (caulis) 105
9.3.2.1. Módosult szártípusok 108
9.3.2.2. A hajtáscsúcs szervezõdése 110 9.3.2.3. A szár elsõdleges szerkezetének
általános jellemzõi 112
9.3.2.4. A gyökér és a szár szállítónyalábjainak
csatlakozása 114
9.3.2.5. A sztéle evolúciója
az elsõdleges szárszerkezetben 116 9.3.2.6. A szárszerkezet másodlagos
módosulásainak típusai 120
9.3.2.7. Évgyûrûs fatestszerkezet, szíjács és geszt,
a héjkéreg 123
9.3.3. A levél (folium, phyllum) 130
9.3.3.1. A levél törzsfejlõdési (filogenetikai)
és egyedfejlõdési (ontogenetikai) eredete 131 9.3.3.2. Levélmódosulások, levélállás (fillotaxia)
és a lombhullás szerkezeti alapja 138 9.3.3.3. A levél szerkezete és adaptív jellemvonásai
különbözõ környezeti körülmények között 142
TARTALOM 7 10. A hajtásos növények fõ rendszertani csoportjai,
virágtalan és virágos (magvas) növények (B. Cs. és Sz. L.) 156
10.1. A harasztok (Pteridophyta) 156
10.2. A nyitvatermõk (Gimnospermatophyta) 159 10.3. A zárvatermõk (Angiospermatophyta) 162 10.3.1. Kétszikûek osztálya –Dicotyledoneae 168 10.3.2. Egyszikûek osztálya –Monocotyledoneae 195 11. A növények testszervezõdésének
alkalmazkodása a környezet vízviszonyaihoz (F. L.) 206 12. A növények testének alkalmazkodása
a környezet fényviszonyaihoz (F. L.) 209
13. A növények szerkezeti és mûködési alkalmazkodása
a környezet hõmérsékleti körülményeihez (B. Cs. és F. L.) 210
14. Növényi életformák 212
Tárgymutató 215
Szakirodalom 237
A kötet szerzői 247
A könyv bevezetõ, alapozó növénybiológiai ismereteket foglal össze azon egyetemi hallgatók számára, akik szakképzésük során felhasználhat- ják a klasszikus növénytan aktualizált, a jelenlegi tudomány szintjének megfelelõ egységesítõ információit a növények testszervezõdésérõl, általá- nos tulajdonságairól, fontosabb rendszertani kategóriáiról, törzsfejlõdésta- ni és alkalmazkodási típusairól. A jegyzet anyaga nagymértékben támasz- kodik a Mikroszkópos növényszervezettan címû egyetemi tankönyv részletes szövettani, szervtani és alaktani adataira, valamint gazdag szem- léltetõ ábraanyagára, vázlatrajzaira és színes fényképeire, melyeket a jelen jegyzetben nem ismételtünk meg. A könyvbe foglalt tananyagot úgy állítot- tuk össze, hogy közös kiindulási alapot képezzen azon alkalmazott, gya- korlati növénytani ismeretek elsajátításához, amelyek felhasználhatók a környezet minõségének vizsgálatában, a környezetvédelemben, az erdõ- gazdálkodásban, a növénytársulástani és növényökológiai vizsgálatokban, a növénytermesztésben, illetve a korszerû kertészeti gyakorlatban. Továb- bá, a tananyag kiindulópont olyan modern növénytani csúcstudomány- ágakban való szakosodáshoz, mint amilyen a molekuláris növényélettan, a növényi sejtbiológia, az ökofiziológia és a növényi biotechnológia.
A tankönyv elõbb ismerteti a növényi életforma általános jellemvoná- sait, a növények helyét és szerepét az élõvilágban, valamint a növényvilág rendszerezésének alapjait. Ezek után röviden összefoglalja az egysejtû és telepes növények fõ csoportjainak ismertetését és ökológiai szerepét, majd bemutatja a hajtásos növényektest szervezõdésének alapvetõ összetevõit és változatait, áttekinti a hajtásos növények fontosabb törzsfejlõdéstani csoportjait, szintetizálva elsõsorban a nálunk gyakoribb és gyakorlati je- lentõségû zárvatermõk rokonsági köreinek rövid és szelektív bemutatását.
Végül a különbözõ környezeti feltételekhez és változatos életmódokhoz való alkalmazkodás legfontosabb megnyilvánulási formáit és szabálysze- rûségeit tárgyalja, mintegy bevezetõként a késõbbi ökofiziológiai ismere- tek elmélyítéséhez. Mindezek által olyan megbízható kiindulási alapot szeretne nyújtani, amely sikeres szakosodást tesz lehetõvé bármely mo- dern növénytani tudományágban és az ezeket felhasználó, szélesebb körû alkalmazott természettudományi tárgykörökben.
Kolozsvár, 2004. január 1. A szerzõk
1. FEJEZET
A NÖVÉNYTANI TUDOMÁNYÁGAK HELYE A BIOLÓGIAI TUDOMÁNYOK RENDSZERÉBEN
A biológia az élõvilág tanulmányozásával foglalkozó tudomány, mely jelenleg erõteljesen fejlõdik a kísérleti mûszerek és módszerek tökéletese- désének köszönhetõen. Elõreláthatóan a XXI. század tudományos fejlõdé- sének alapvetõ területe, alkalmazott ismeretei (például a géngyógyászat, a biotechnológiai termelés, a biológiai energiatermelés, a környezetminõsé- gi biomonitorizálás stb. által) döntõ módon befolyásolják életünk és az egész földi élet minõségét. Tudományágai két nagy területre oszthatók: az egyeden belüli (molekuláris, sejti, szervi) szervezõdési szintek vizsgálatá- ra és az egyed feletti szervezõdési szintek (populációk, fajok, életközössé- gek, ökológiai rendszerek) tanulmányozására.
1. Az idiobiológia az egyedi szervezeten belüli szintekkel foglalko- zik. Csupán a legalapvetõbb ágazatait említjük meg.
1a) A sejttan, a genetika és a biokémia együttesen a molekuláris bio- lógia fõ összetevõit képezi, ez jelenleg az élettudományok legdinamiku- sabban fejlõdõ része.
1b) A növénytan magába foglalja a növényanatómiát (szervezettant, ennek részei a szövettan vagy hisztológia, a szervtan vagy organográfia, valamint az alaktan vagy morfológia) és a növényélettant (fiziológiát), ez utóbbinak két fõ ágazata az anyagcsere-élettan (fotoszintézis, légzés, táp- anyagszállítás, ásványi táplálkozás, vízgazdálkodás stb.) és a fejlõdésélet- tan (növekedés, bioritmusok, hormonális szabályozás, virágzás, szaporo- dás, öregedés, ingerválaszok stb.). Speciális területei közül jelenleg a legerõteljesebb fejlõdést az ökofiziológia és a stresszélettan mutatja.
1c) Az állattan keretébe tartozik az anatómia (szövettan, szervtan, alaktan), az élettan (anyagcsere-élettan: táplálkozás, légzés, keringés, ki- választás, köztes anyagcsere és ezek hormonális szabályozása, az ideg- élettan, mozgás- és érzékelésélettan, szaporodásélettan), valamint az eto- lógia (viselkedéstan).
1d) A mikrobiológia két fõ részét a bakteriológia és a víruszológia ké- pezi.
1e) Az immunológia, az embriológia és a biotechnológia (genetikailag át- alakított élõlények felhasználása hasznos termékek ipari méretû elõállításá- ra) az elõbbi tudományágakra támaszkodó modern tárgyköröket képviseli.
2. A szinbiológia az élõ anyag egyed feletti szervezõdési szintjeinek tanulmányozását foglalja magába. Területei közül szintén csak néhány jellemzõbbet említünk meg.
2a) A rendszertan fõ ágazatai a növényrendszertan, a gerinctelen és gerinces állatok rendszertana, valamint a mikológia (a gombák tanulmá- nyozása).
2b) Az ökológia az élõlényeknek külsõ környezetükkel való kölcsön- hatásait vizsgálja, fontosabb területei közül megemlítjük a populációtant, a társulástant (cönológiát) és a hidrobiológiát, ez utóbbinak pedig a két fõ része a limnológia (szárazföldi folyó- és állóvizek tanulmányozása) és az oceanológia.
2c) Az evolúciótan az élõvilág fejlõdésének, elterjedésének és átala- kulásának törvényszerûségeit tanulmányozza a földtörténeti korok folya- mán, általános nézetet nyújtva az élõ anyag önszervezõdésének, önsza- bályozásának és környezetébe való beilleszkedésének tekintetében.
A számos biológiai tudományág között természetesen sokrétû átfedé- sek vannak, egymás ismereteire támaszkodnak és kölcsönösen kiegészítik egymást az élettudományok egységes rendszerén belül, ugyanakkor szoros kapcsolatban vannak a kémiai, fizikai, földrajzi, matematikai stb. tudomá- nyokkal. Továbbá, a biológiai tudományokra támaszkodik számos más, al- kalmazott tudományterület, mint például az orvostudomány, a mezõgaz- dasági tudományok, az erdõgazdálkodás, a környezettudományok stb.
Nem titkolható jelenség, hogy a XX. század második felében a moleku- láris biológia rendkívül látványos felemelkedéséhez a biológiai háborútól való félelem adta meg a lendületet. Ekkor a totális háborút „ABC-háború- nak” nevezték (A –atomic; B –biological; C –chemical), biológiai vetülete pedig abból indul ki, hogy genetikailag manipulált fertõzõ mikroorganiz- musok általi járványos megbetegítés útján nagy embertömegek küzdõképes- sége semmisíthetõ meg. Ezzel szemben jelenleg a lõfegyverek nélküli, új vi- lágméretû háborús veszélyt „DEF-háborúnak” nevezik (D –demographic; E –environmental; F –financial) és immár nemcsak az ökológusok számára vált nyilvánvalóvá, hogy egy országot környezetpolitikailag is tönkre lehet tenni, mindenféle hadüzenet nélkül. A kérdés az, hogy ha a biológiai hatás nem ismer országhatárokat, hogyan lehet meggátolni korlátlan terjedését.
Jelenleg ez az aggodalom készteti a társadalmi erõket az ökológiai és a bio- technológiai kutatások pénzügyi támogatására és lendületes fejlesztésére.
2. FEJEZET
A NÖVÉNYI ÉLETFORMA JELLEMVONÁSAI
A növények, nagy változatosságuk ellenére, számos közös általános tulajdonságot mutatnak, melyek által beilleszkednek az élõvilág egészé- be, másrészt pedig elkülönülnek egyéb élõlénytípusoktól.
A növények mutatják az élõ anyag általános tulajdonságait, mint amilyen az anyagcsere, az önszabályozás, az önálló növekedés és fejlõ- dés, az öröklékenység, a változékonyság, az alkalmazkodóképesség, az ingerelhetõség, a szaporodás.
Az anyagcsere vagy metabolizmus azt jelenti, hogy a növények a kül- sõ környezetbõl felvett anyagokat és energiát átalakítják testük saját élõ anyagává és sajátos energiaformákká, testanyagaikat állandóan megújítják és szabályozottan módosítják életmûködéseik során. Az anyagcsere felépí- tõ, energiát elhasználó folyamatai az anabolizmust, lebontó, energiafelsza- badító folyamatai pedig a katabolizmust képviselik. Az anyagcsere bioké- miai reakciók által valósul meg, melyek katalizátorai az enzimfehérjék.
Anyagcseréje során a növény környezetét is fokozatosan módosítja, hiszen bizonyos szervetlen anyagokat vesz fel (szén-dioxidot, ásványi só- kat), ezeket átalakítja és másfajta, általában szerves anyagokat ad le kör- nyezetébe (például gyökérsavakat, terpéneket, lehulló leveleinek, virága- inak és terméseinek fehérjéit, nukleinsavjait és összetett cukrait stb.). Így a növény nyílt rendszerként mûködik, mely állandó jellegû kétirányú kapcsolatban van környezetével, és anyagcseréjének önszabályozása ál- tal õrzi meg dinamikus (állandóan ingadozó) egyensúlyát változó külsõ körülmények között is. Mindezek által a növény élete alatt mennyiségi (növekedési) és minõségi (fejlõdési) visszafordíthatatlan változások soro- zata megy végbe. Ezek teremtik meg a feltételeket a szaporodásra, vagyis a szülõk örökletes tulajdonságainak különbözõ kombinációkban történõ átadására az utódokba.
Az alapvetõ tulajdonságok megõrzése és megnyilvánulása az egyedi élet és az egymás utáni nemzedékek során az örökletes program (a min- den sejtben meglevõ genetikai anyag) által történik. Ebben a programban mutációk vagy az ivaros szaporodás révén bekövetkezõ rekombinációk során jöhetnek létre véletlenszerû változások, amelyek az egyedeket egy-
mástól részben különbözõvé teszik és hozzájárulnak új tulajdonságok megjelenéséhez. Ezek az új tulajdonságok adott körülmények között le- hetnek többé vagy kevésbé elõnyösek vagy hátrányosak, így az élõhely környezeti körülményeihez való alkalmazkodás során szelektálódnak és különbözõ mértékben terjednek el az egyes változatok. A környezetbe va- ló sikeres beilleszkedés egyik feltétele az ingerelhetõség, ami által a nö- vény tudomást szerez a hozzá érkezõ külsõ jelzésekrõl és a számára leg- megfelelõbb módon irányított és ellenõrzött belsõ változásokkal próbál válaszolni ezekre.
A növényeket egyéb élõlényektõl számos sajátos tulajdonság külön- bözteti meg, ezek közül legfontosabbak az autotróf (önellátó) táplálkozási mód, a korlátlan növekedés, a színtestek (plasztiszok) jelenléte, a kétszaka- szos (spórás és gamétás) szaporodási ciklus, a sejtek omnipotenciája, a nagy testfelület és szárazföldi növények esetében a helyhez kötött életmód.
A növények alapvetõ jellemvonása azautotrófia,az önellátó táplálko- zásmód. Ez azt jelenti, hogy környezetükbõl egyszerû, energiaszegény szervetlen anyagokat vesznek fel és a napfény energiájának felhasználásá- val ezekbõl önmaguk építik fel testük összetett, vegyi energiában (kovalens kötésekben) gazdag szerves anyagait: a fehérjéket, a cukrokat (szachari- dokat), a lipideket, a nukleinsavakat (DNS, RNS), a szekunder meta- bolitokat (például terpéneket, fenolszármazékokat, alkaloidokat) stb. A nö- vények két fõ tápláléka a szén-dioxid és a víz. A szén-dioxid-molekulák szénatomjaiból a fényenergia segítségével összeszerelik összes szerves anyaguk szénláncát, és mivel minden szerves anyagban hidrogénatomok is vannak, a szén-dioxid mellett szükségük van hidrogénforrásra is. Ezt szá- mukra a vízmolekulák képviselik, melyekbõl a hidrogénatomok felhaszná- lása után visszamarad az oxigén, ettõl pedig úgy szabadulnak meg, hogy leadják környezetükbe, vagy egy részét légzésük során oxidációs lebontó folyamatokban utólag hasznosítják. A felépítõ, szén-dioxidot asszimiláló folyamatok csak fényenergia segítségével zajlanak, sötétben az elõbbi tar- talékok egy része légzés útján lebomlik, hogy a következõ reggelig az élet- folyamatok a már beépített energia egy részének felszabadításával fenntart- hatók legyenek.
A szén-dioxid és a víz mellett a növények egyéb egyszerû anyagokat (ásványi sókat) is felvesznek: nitrogén-, foszfor-, kénvegyületek vizes olda- tát, fémionokat stb., és ezeket is beépítik testük élõ anyagába. Mivel szelek- tív anyagfelvételük során a rendelkezésre álló különbözõ ásványi sókból más-más mennyiségeket vesznek fel, környezetük vegyi összetételét nem-
2. A NÖVÉNYI ÉLETFORMA JELLEMVONÁSAI 15 csak minõségi, hanem mennyiségi szempontból is módosítják az idõk so- rán, aktív szerepet játszva az egyes elemek természetbeni körforgásában.
Autotrófiájukból származik a növények nélkülözhetetlen szerepe minden ökológiai rendszerben, hiszen a növények az új szerves anyagok elsõdleges termelõi és az élelmi láncokba új energiamennyiségeket beho- zó (napfényt vegyi energiává alakító) szervezetek, ezért csakis a növé- nyek által biztosított tápanyagokkal és energiával létezhetnek a fogyasz- tók és a lebontók, vagyis az állatok és a mikroorganizmusok.
Mivel külsõ energiaforrást használnak, a növényekre általában a nagy testfelület (elágazó és lapos testrészek kifejlesztése) jellemzõ, és mi- vel nem kell táplálék után kutatniuk, a nagyobb stabilitást biztosító hely- hez kötött életmód terjedt el a szárazföldi és vízi, nagyobb testû növé- nyek körében.
A növények másik jellemzõje a korlátlan növekedés, ami által életük végéig képesek újabb és újabb, fiatal testrészek létrehozására. Ez annak tulajdonítható, hogy a szárak rügyeinek hajtáscsúcsában és a gyökerek végein soha nem fogynak ki az osztódó szövetek. Ugyanakkor a növényi sejtek megõrzik omnipotenciájukat, vagyis azon képességüket, hogy elõbbi szakosodásukat elveszítve „megfiatalodjanak”, visszanyerjék osz- tódó képességüket és új testrészeket vagy teljesen ép új növényegyedet alakítsanak ki. Tehát növények esetében egyetlen sejtbõl egy egész szer- vezet regenerálódhat. E tulajdonságukat használják fel a növényszaporí- tási biotechnológiák a klónozás során.
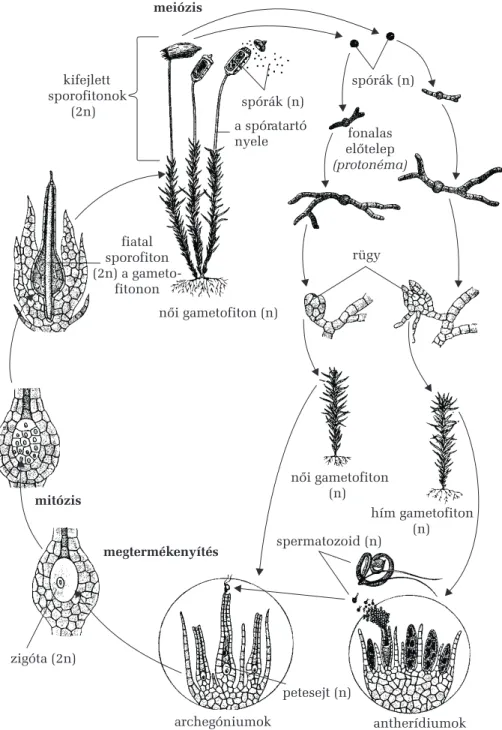
A növények sajátossága akétszakaszos szaporodási ciklusis. Ez azt jelenti, hogy kétféle szaporodási mód követi egymást (egy ivartalan és egy ivaros), és mindkétféle szaporítósejtbõl (a spórákból és a gamétákból) származik külön-külön egy-egy testforma az egyedfejlõdés során. Tehát minden növénynek van ivartalan és ivaros testformája, ez utóbbi pedig önmaga is kétféle lehet: nõi és hím ivaros életszakasz. Mindez annak tu- lajdonítható, hogy a szaporodási folyamat során a számfelezõ (redukciós) sejtosztódás vagy meiózis alkalmával nem keletkeznek közvetlenül gamé- ták, amelyek megtermékenyítéskor létrehozzák az új egyedet, hanem elõbb ivartalan szaporítósejtek (spórák) jönnek létre, ezek osztódásokkal létrehoznak egy új testet, melyen ivarszervek fejlõdnek, és csak ezekben alakulnak ki, nem számfelezõ, hanem számtartó sejtosztódással (mitózis útján) a gaméták. Így minden növény életében van egy diploid (kétszeres kromoszómakészletes) életszakasz, mely szaporodáskor spórákat képez, ezért sporofitonnak nevezzük, és egy haploid (egyszeres kromoszóma- készletes) életszakasz, mely szaporodáskor gamétákat hoz létre, ezért ga-
metofitonnak nevezzük. A két életszakasz szabályosan követi egymást minden életciklus során, a sporofiton a megtermékenyített petesejttõl (a zigótától) a spóraképzésig, vagyis a meiózis bekövetkeztéig tart, a game- tofiton pedig a spóráktól a gaméták létrejöttéig és egyesüléséig, vagyis a megtermékenyítésig terjed. Ezek alapján minden ivarosan szaporodó nö- vény szaporodási ciklusának fõ állomásai a következõk:
Zigóta (megtermékenyített petesejt, 2n) – számtartó (mitotikus) sejt- osztódások – diploid test (sporofiton, 2n) – ivartalan szaporító- (spóra- képzõ) szervek (2n) – a diploid spóraanyasejtek számfelezõ, redukciós osztódása (meiózis) – ivartalan szaporítósejtek (spórák, n) – haploid test (gametofiton, n) – ivaros szaporító- (gamétaképzõ) szervek (n) – a haploid gamétaanyasejtek számtartó osztódása (mitózis) – ivaros szaporítósejtek (nõi és hím gaméták, n) – megtermékenyítés (a kétszeres kromoszóma- készlet helyreállítása a két szülõtõl az utódnemzedékben) – zigóta (2n).
A növényvilág evolúciója (törzsfejlõdése) során a két életszakasz kö- zül a sporofiton válik egyre fejlettebbé, hiszen a kétszeres örökítõ anyag- készlet nagyobb változatosságot és jobb alkalmazkodóképességet biztosít számára, a gametofiton pedig fokozatosan redukálódik, elveszíti önálló- ságát és beépül a sporofitonba, mely védelmezi és táplálja. Ezért a virá- gos növényeknél a gametofiton életszakaszt nem látjuk szabad szemmel, hiszen a virágba beépülten fejlõdik.
3. FEJEZET
A NÖVÉNYEK HELYE AZ ÉLÕVILÁG TÖRZSFEJLÕDÉSTANI RENDSZERÉBEN
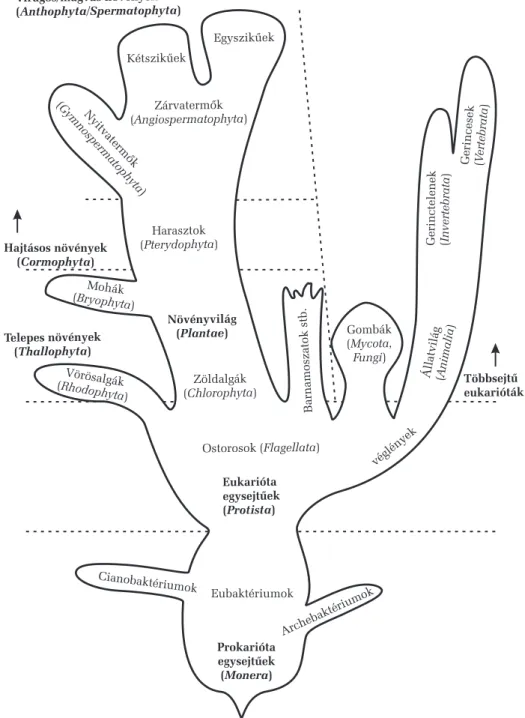
Az élõvilág öt nagy testszervezõdési kategóriára osztható, ezek a pro- karióták (a baktériumok), a protiszták (az egysejtû eukarióták), a többsejtû növények, a többsejtû gombák és a többsejtû állatok (1. ábra). Az életfor- mák tekintetében megkülönböztetünk baktériumokat, növényeket, gombá- kat és állatokat, a protiszták közül egyes egysejtûek növényi módon, má- sok gomba módon, ismét mások állati módon élnek, ezért életmûködési szempontból az eukarióta egysejtûek különbözõ képviselõi hozzátartoznak a növényvilághoz, az állatvilághoz vagy a gombák világához, habár test- szervezõdésük alapján a protiszták külön csoportjába sorolhatók. Ily mó- don, például, az egysejtû algák testfelépítésük tekintetében a protiszták- hoz, életmûködési szempontból pedig a növényekhez tartoznak.
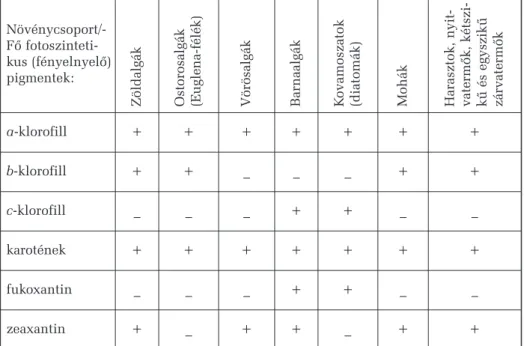
Aprokariótákat a baktériumok képviselik. Ezek sejtmag nélküli egy- sejtûek, minden élõhelyet benépesítenek, nagyon változatos táplálkozási módokra képesek: vannak közöttük lebontók, élõsködõk, fotoszintetizáló és kemoszintetizáló autotrófok stb. Nagyon gyorsan szaporodnak (ketté- osztódással), rendkívül alkalmazkodóképesek. Legõsibb képviselõik az archebaktériumok, amelyek ma csak néhány szélsõséges életkörülmény között, oxigénmentes helyeken (például vízfenéki iszapban) élnek és na- gyon különböznek az összes többi élõlénytõl. A többi baktérium az eubaktériumok csoportját képviseli, és ide tartoznak a cianobaktériumok is, melyeket sötétzöld színük és oxigéntermeléssel társuló fotoszintézi- sük alapján régebb tévesen a növényvilágba, az algák közé soroltak és a ma már helytelen kékalga vagy kékeszöld moszat névvel illettek. Ma már tudjuk, hogy ezek az élõlények baktériumok, nincs sejtmagjuk, egyetlen kromoszómájuk van sejtenként és egyszerû kettéosztódással (amitoti- kusan) szaporodnak. A növényvilággal való hasonlóságuk azzal magya- rázható, hogy valószínûleg egyes õsi formáikból alakultak ki a fotoszin- tézist végzõ növényi kloroplasztiszok, tehát mûködési szempontból a zöld színtestekkel egyenértékûeknek tekinthetõk.
A cianobaktériumok aerob légzésû, fény jelenlétében fejlõdõ, fotoszin- tézisük során oxigént felszabadító prokarióták. Fõ fényelnyelõ pigment-
ként a klorofillok típusai közül mindig csakis a-klorofillt tartalmaznak.
A sejthártya betüremkedéseibõl származó fotoszintetikus membránjaik fe- lületén félgömb alakú testecskékbe (fikobiliszómákba) csoportosuló kék és vörös pigmentmolekulák (fikocianin, allofikocianin, fikoeritrin) egészítik ki az a-klorofillnak és a karotenoidoknak köszönhetõ fényelnyelési színké- pet, akárcsak a vörösalgák esetében. Egyesek (például azAnabaenafajok) a molekuláris nitrogén megkötésére és ammóniummá való átalakítására képesek, így például az elárasztott rizsültetvények vizében élõ cianobakté- riumoknak köszönhetõen nem szükséges az ilyen rizsföldek nitrogéntrá- gyázása. A cianobaktériumok fõ káros hatása a vízi életterekben a vízvirág- zás (habár ezt rajtuk kívül egyes algák is okozhatják). Ezt nagymértékû túlszaporodásuk okozza meleg nyári napokon, fõleg olyan vizekben, ame- lyek szervetlen tápanyagokban (fõleg a nitrogén és a foszfor vegyületeiben) feldúsulnak (eutrofizálódnak), mert a környezõ megmûvelt területek túl- zott trágyázása során sok nitrogén- és foszforsó mosódik be az illetõ vízbe.
Ilyen körülmények között a hirtelen felszaporodó cianobaktériumok sötét- zöld bevonatot alkotnak a víz felszíni rétegében, egymást is beárnyékolják, a fényhez nem jutó sejtek elpusztulnak, lesüllyednek és közben lebomla- nak, ez a folyamat pedig elhasználja az oxigént a vízbõl, így számos egyéb vízi élõlény is megfulladhat. Például a halak ilyenkor tömegesen elpusztul- nak. Emellett számos cianobaktérium toxinokat (cianotoxinokat) is termel, melyek károsíthatják az állatok idegrendszerét, továbbá az örökítõ anyag, bizonyos enzimek, sejtmembránok szintjén levõ szállítómolekulák mûkö- dését zavarják meg. Ilyen toxintermelõ és vízvirágzást okozó egysejtû cianobaktériumok például aMicrocystisfajok. Mások viszont fehérjékben, vitaminokban és ásványi sókban gazdag takarmányként, élelmiszeri ada- lékként vagy kozmetikumként hasznosíthatók és e célból üzemi mértékben termeszthetõk. Ilyen például aSpirulina platensis(Arthrospira fusiformis), melynek sejtjei spirálisan csavarodott fonalas telepet alkotnak az erõsen lúgos és meleg vizekben.
Léteznek a baktériumoknál sokkal egyszerûbb szervezõdési formák is, amelyek az élõ anyag egyes tulajdonságait mutatják, ezek viszont nem tekinthetõk teljes értékû élõlényeknek, mert nem léteznek önállóan, ha- nem sejteken belül élõsködnek, nincs saját anyagcseréjük. Ilyenek a ví- rusok, valamint a náluk is egyszerûbb viroidok (fertõzõ RNS-molekulák) és a prionok (fertõzõ fehérjerészecskék). Mindezek sejtszerkezet nélküli biológiai entitások, az élettelen és az élõ anyag határán állnak, valószínû- leg õsi prokariótákból fejlõdtek vissza, vagy pedig sejtes szervezetekbõl kivált, részlegesen önállósult örökítõ (információhordozó) anyagrészek.
3. A NÖVÉNYEK HELYE AZ ÉLÕVILÁG TÖRZSFEJLÕDÉSI RENDSZERÉBEN 19 A prokariótáknál fejlettebb testszervezõdési szintet a protiszták, va- gyis az eukarióta (valódi sejtmaggal rendelkezõ) egysejtûek képviselik.
Ezek változatos táplálkozási formákra képesek, egyesek fény jelenlétében autotrófok, sötétben pedig heterotrófok (például számos ostoros egysejtû).
Rájuk már általánosan jellemzõ az aerob (légköri oxigént felhasználó) lég- zés és ezen a szinten megjelenik az ivaros szaporodás, aminek alapján a számfelezõ sejtosztódás (a meiózis) áll. Ezáltal az utódokban két szülõ örökletes tulajdonságai keverednek különbözõ kombinációkban, így na- gyobb változatosságot mutatnak és többféle vízi élõhelyet népesítenek be.
Egyes õsi formáik képviselik a közös õseit az állatvilágnak, a gombáknak és a növényvilágnak, amelyek többsejtû szervezetekként fejlõdtek tovább.
Az állatok teste diploid sejtekbõl áll (diplonták), nincs sejtfaluk és szilárd szerves táplálékot is képesek megemészteni. A gombák szintén kész szerves anyagokkal táplálkoznak, de ezeket vizes oldat formájában szívják fel. Testükre a haploid sejtek jellemzõek (haplonták).
A gombák (MycobiontavagyFungi) lehetnek szaprofiták (televényla- kók) vagy paraziták (élõsködõk). Az elõbbiek a baktériumokkal együtt az elpusztult szervezetek szerves testanyagainak a lebontása által fontos szerepet töltenek be az anyagok természetbeni körforgásában (lebontók), általuk jutnak vissza a talajba a különbözõ ásványi sók, amelyeket a nö- vények újra felvehetnek. Az élõsködõ gombák számos veszélyes betegsé- get okoznak növények és állatok testében egyaránt. Nincsenek plaszti- szaik, csakis heterotróf táplálkozási módra képesek. Sejtfaluk fõ alkotója általában az ízeltlábú állatok külsõ testvázát is felépítõ kitin. Fõ tartalék tápanyaguk a glikogén (ugyanaz az összetett cukor, amit az állati sejtek is raktároznak). Testük megnyúlt, fonalas egységekbõl: hifákból áll, ez lehet egysejtû vagy többsejtû. A hifák lazább vagy tömörebb szövedéke a mi- célium. Spórákkal és gamétákkal egyaránt szaporodhatnak, gyakori a hifasejtek két külön sejtmagot viselõ állapota, a dikarion (n + n). A nyál- kagombák (Myxomycetes) sejtfal nélküli, állábas sejttömegeket alkotnak.
A moszatgombák (Zygomycetes) legismertebb képviselõi a fejespenész és a szõlõragya. A tömlõsgombák (Ascomycetes) spóraképzõ testrésze zárt tömlõ, ismertebb képviselõik az ecsetpenész (Penicillium fajok), a sör- élesztõ, a talajban élõ szarvasgomba. A pálcikaspórás vagy bazídiumos gombák (Basidiomycetes) spórái szabadon állnak ujjszerû kis nyúlványok végén. Egyik legveszélyesebb parazita képviselõjük a gabonarozsda, melynek életciklusa kétféle növényben (egy köztes gazdán és egy gaz- dán) zajlik. A pálcikaspórás gombák közé tartoznak a tipikus kalapos gombák is, ezeknél a talajból kiemelkedõ, sûrûn összefonódó hifarészek
tönköt és kalapot alkotnak. A tönk körül fent gallér, lent pedig bocskor le- het jelen. A kalap alsó részén van a lemezes vagy csöves termõréteg a spórákkal. Egyes fajoknál a kalap szélén maradhat egy fátyol, a kalap fel- sõ felületén pedig lehetnek kalappikkelyek. E testrészek jellemvonásai nagyon fontosak az egymáshoz hasonló ehetõ és mérgezõ gombafajok megkülönböztetésében. Például a csiperkét a gyilkos galócától (mely erõs idegmérget termel) az különbözteti meg, hogy a csiperke kalapjának alsó oldalán a termõréteg rózsaszínû vagy barna, tönkje körül nincs bocskor, míg a gyilkos galóca kalapjának termõrétege fehér, a tönk alsó része kö- rül pedig hártyás bocskor található. Általános javaslat, hogy ha egy gom- bafajt nem ismerünk bármely fejlõdési stádiumában teljes bizonyosság- gal, akkor ezt ne szedjük le fogyasztás céljából.
Számos gomba társul egyéb élõlényekkel, így egyesek fák vagy lágy szárú növények gyökereivel alkotnak mikorizákat, elõsegítve a víz és a benne oldott ásványi sók felszívását, mások pedig a zuzmók felépítésé- ben vesznek részt.
A protiszták és a növények közötti kapcsolatot az algák vagy mosza- tok teremtik meg. Egysejtû, sejtcsaládos és többsejtû képviselõik egy- aránt ismertek és növényi típusú életmódot folytató, fõleg vízi autotróf szervezetek. A vörösalgák (Rhodophyta) önálló fejlõdési vonalat képvi- selnek, a többi növénynél mélyebb tengervizekben is megélnek, ahol a fény már nagyon gyenge és csupán kék és zöld sugarakat tartalmaz. Egy másik fejlõdési vonalon alakultak ki a barnás-sárgás színû kromofiták (Chromophyta), amelyek közé a barnaalgák, a kovaalgák, a sárgaalgák és rokonaik tartoznak, ezek a közepesen mély vízrétegeket népesítik be, ahova a kék és zöld mellett a sárga fénysugarak is bejutnak (de a vörös fény nem). A vizek felszínhez közeli, jobban megvilágított és vörös suga- rakban gazdag rétegében élnek a nagyon változatos zöldalgák (Chlorophyta), egyesek közülük a nedves szárazföldre is kijutottak és bi- zonyos õsi formáikból származtathatók egyrészt a mohák, másrészt a ha- rasztok.
A mohák (Bryophyta) számos képviselõje már szárazföldi növény, a májmohák telepteste még homogén, a lombos moháké szárszerû és levél- szerû részekre tagolódik. A többsejtû algákkal együtt a mohák ateleptes- tû növényeket (Thallophyta) képviselik, amelyeknek még nincsenek va- lódi szöveteik és szerveik.
A harasztok (Pterydophyta) fõ képviselõi a korpafüvek, a csipkeha- rasztok, a zsurlók és a páfrányok (csak a nagylevelû harasztokat nevezzük páfrányoknak). Ezek teste már valódi szervekre tagolódik, melyek szako-
3. A NÖVÉNYEK HELYE AZ ÉLÕVILÁG TÖRZSFEJLÕDÉSI RENDSZERÉBEN 21 sodott szövetekbõl épülnek fel. A vegetatív vagy önfenntartó szervek a gyökér, a szár és a levél, a generatív vagy szaporító testrészek pedig a spó- raképzõ szervek és a gamétaképzõ szervek. Viráguk és magjuk nincs, ezért a virágtalanok (Cryptogamae,rejtett szaporodásúak) szintjét képviselik.
A nyitvatermõk (Gymnospermae,csupasz magvúak), mint amilyenek a fenyõfélék, már virágok formájában elkülönült szaporítószervekkel (por- zókkal, termõkkel) rendelkeznek, megporzás és megtermékenyítés után a virágok magkezdeményeibõl magok fejlõdnek, amelyek magukba zárják, védik és táplálják az új csíranövényt. A nyitvatermõk magjai még szaba- don állnak, nincsenek termésbe zárva.
A zárvatermõk (Angiospermae, bezárt magvúak) a legfejlettebb nö- vénycsoportot képviselik, magvaik védelmét és sikeresebb elterjesztését (ezáltal pedig a fajok elterjedési területének bõvítését) a termések biztosít- ják, amelyek a virág termõjének zárt magházából fejlõdnek ki. A nyitvater- mõkkel közös õsbõl elõbb a kétszikû zárvatermõk alakultak ki (Dicotyledo- natae), melyeknél a magban levõ embriónak két sziklevele van. Belõlük származtathatók az egyszikû zárvatermõk (Monocotyledonatae), melyeknél a csíranövénynek már csak egyetlen sziklevele fejlõdik ki.
A nyitvatermõk és a zárvatermõk (kétszikûek és egyszikûek) a virá- gos vagy magvas növényeket képviselik (Anthophyta vagy Spermato- phyta), a harasztokkal együtt pedig a szervekre tagolt testû,hajtásos nö- vények(Cormophyta) fejlõdési szintjét alkotják.
A fentiekbõl kitûnik, hogy a növények lehetnek egysejtûek, teleptes- tûek és hajtásos növények. Közös tulajdonságuk, hogy kétszakaszos sza- porodási ciklusuk van (haplodiplonták) és fotoszintézisük által autotróf életmódot folytatnak, aminek sejtszintû szerkezeti alapja a színtestek (plasztiszok) jelenléte.
Teljes mértékben helytelen és pontatlan az „alacsonyabb rendû” és a
„magasabb rendû növény” megnevezés, hiszen ebbõl nem lehet tudni, hogy melyik fejlõdési szintrõl van szó. Például, a moha az algákhoz képest magasabb rendû, a harasztokhoz viszonyítva pedig alacsonyabb rendû.
Prokarióta egysejtűek (Monera)
Archebaktériumok Eubaktériumok
Eukarióta egysejtűek (Protista)
Cianobaktériumok
Barnamoszatokstb.
véglények Gombák (Mycota,) Fungi
Állatvilág (
) Animalia Gerinctelenek ()Invertebrata
Gerincesek
()
Vertebrata
Zöldalgák (Chlorophyta) Vörösalgák
(Rhodophyta)
Növényvilág (Plantae) Mohák
(Bryophyta)
Harasztok (Pterydophyta) Nyitva
termők (
) Gymnospermatop
hyta Kétszikűek
Egyszikűek
Zárvatermők (Angiospermatophyta) Virágos/magvas növények
(Anthophyt /Spermatophytaa )
Hajtásos növények (Cormophyta)
Telepes növények (Thallophyta)
Többsejtű eukarióták
Ostorosok (Flagellata)
1. ábra.Az élõvilág törzsfájának egyszerûsített vázlata (Margulis 1970 nyomán, módosítva)
4. FEJEZET
RENDSZERTANI KATEGÓRIÁK
Az élõvilág nagy változatossága az áttekinthetõség céljából szükséges- sé teszi a rendszerezést, a csoportosítást közös tulajdonságok, törzsfejlõ- déstani rokonsági viszonyok alapján. Az élõvilág rendszerezésének alap- egysége a faj (species). Minden élõlény valamely fajhoz tartozik, a fajon belül az egyedek keresztezõdhetnek egymással és termékeny utódokat hozhatnak létre, így a fajnak egy közös génállománya van, ami egyben megkülönbözteti egyéb fajoktól. A fajt különbözõ élõhelyeket azonos vagy más-más idõben benépesítõ egyedcsoportok, populációk alkotják. Ha va- lamilyen elszigetelõdés a különbözõ helyi körülményekhez sajátosan al- kalmazkodó populációkategóriák elkülönülését eredményezi, akkor fajon belüli alegységek: alfajok vagy fajták, változatok és formák jöhetnek létre.
Ha ezek egyedei egymással találkoznak, szabadon keresztezõdhetnek, ami azt mutatja, hogy bizonyos tulajdonságok elkülönülése ellenére az alegy- ségek ugyanazon fajon belüliek. Az alfaj kifejezést vadon élõ, a természet- ben önmagától létrejött fajon belüli változatokra használjuk, míg fajtákról akkor beszélünk, ha ezeket az ember hozta létre mesterséges kiválogatás és nemesítés során.
A Botanikai Nevezéktan Nemzetközi Kódexe jelenleg 12 hierarchikus (egymásba épülõ) rendszertani alapkategóriát állapít meg, de indokolt eset- ben megengedi köztes egységek beiktatását is (alcsalád, fõrend, alosztály, altörzs stb.). A 12 alapkategória, a nagyobbtól az egyre kisebb rokonsági kö- rök felé haladva, a következõ: világ, törzs, osztály, rend, család, tribusz, nemzetség (génusz), szekció, sorozat (szériesz), faj, változat, forma. Ha a rendszertani kategória nevét vagy rangját nem kívánjuk megadni, akkor ál- talánosan a taxon kifejezést használjuk. A tudományos nevezéktanban a nemzetség (nagyon közeli rokon fajok csoportjának) neve mindig egyes számú latin fõnév, a fajnév pedig ehhez nyelvtanilag illeszkedõ melléknév.
A fenti rendszertani kategóriák közül az alapvetõek a faj, a nemzetség, a család, a rend, az osztály és a törzs. A közeli rokon, sok közös tulajdon- ságot mutató fajok egyazon nemzetséghez (génuszhoz) tartoznak, a rokon nemzetségek egy családot alkotnak, a közeli családok egyazon rend részei,
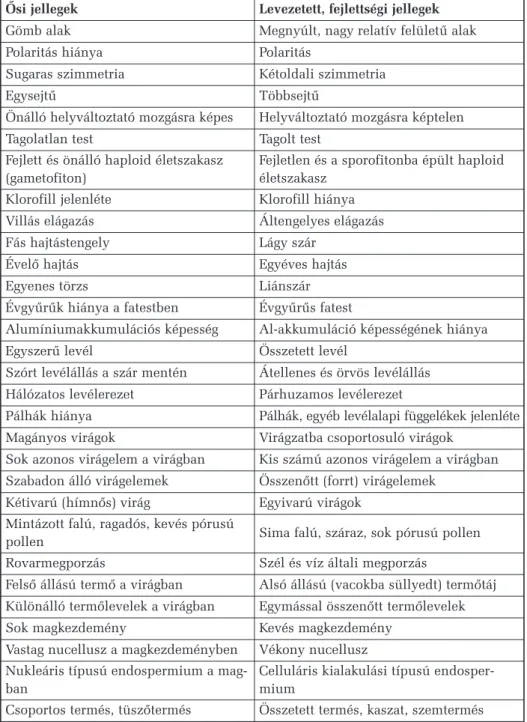
1. táblázat.Õsi és levezetett növényi tulajdonságpárok
Õsi jellegek Levezetett, fejlettségi jellegek Gömb alak Megnyúlt, nagy relatív felületû alak
Polaritás hiánya Polaritás
Sugaras szimmetria Kétoldali szimmetria
Egysejtû Többsejtû
Önálló helyváltoztató mozgásra képes Helyváltoztató mozgásra képtelen
Tagolatlan test Tagolt test
Fejlett és önálló haploid életszakasz
(gametofiton) Fejletlen és a sporofitonba épült haploid életszakasz
Klorofill jelenléte Klorofill hiánya Villás elágazás Áltengelyes elágazás
Fás hajtástengely Lágy szár
Évelõ hajtás Egyéves hajtás
Egyenes törzs Liánszár
Évgyûrûk hiánya a fatestben Évgyûrûs fatest
Alumíniumakkumulációs képesség Al-akkumuláció képességének hiánya
Egyszerû levél Összetett levél
Szórt levélállás a szár mentén Átellenes és örvös levélállás Hálózatos levélerezet Párhuzamos levélerezet
Pálhák hiánya Pálhák, egyéb levélalapi függelékek jelenléte Magányos virágok Virágzatba csoportosuló virágok
Sok azonos virágelem a virágban Kis számú azonos virágelem a virágban Szabadon álló virágelemek Összenõtt (forrt) virágelemek
Kétivarú (hímnõs) virág Egyivarú virágok Mintázott falú, ragadós, kevés pórusú
pollen Sima falú, száraz, sok pórusú pollen Rovarmegporzás Szél és víz általi megporzás
Felsõ állású termõ a virágban Alsó állású (vacokba süllyedt) termõtáj Különálló termõlevelek a virágban Egymással összenõtt termõlevelek Sok magkezdemény Kevés magkezdemény
Vastag nucellusz a magkezdeményben Vékony nucellusz Nukleáris típusú endospermium a mag-
ban Celluláris kialakulási típusú endosper-
mium
Csoportos termés, tüszõtermés Összetett termés, kaszat, szemtermés
4. RENDSZERTANI KATEGÓRIÁK 25 a rokon rendek ugyanazt az osztályt építik fel, a közvetlen közös õsbõl származtatható osztályok pedig ugyanahhoz a törzshöz tartoznak.
A törzsfejlõdéstani rendszerezés szemléltetésére konkrét példaként tekintsük át a tulipán rendszertani besorolását egyre nagyobb egységekbe:
– faj (species): pompás tulipán (Tulipa gesneriana) – nemzetség (genus): tulipánok (Tulipa)
– család (familia): liliomfélék (Liliaceae) – rend (ordo):Liliales
– osztály (classis): egyszikûek (LiliopsidavagyMonocotyledonopsida) – törzs (phylum): zárvatermõk (MagnoliophytavagyAngiospermato-
phyta)
– subregnum: hajtásos növények (Cormobionta) – világ (regnum): növényvilág (Plantae).
Látható, hogy minden rendszertani kategória tudományos nevének jellegzetes végzõdése van („eae” a család, „ales” a rend, „opsida” az osz- tály, „phyta” a növénytörzs számára stb.).
A helyes rendszertani besorolásban gyakran segít a megfigyelhetõ tu- lajdonságok õsi, primitív vagy levezetett, fejlett jellege. A növények kö- rében érvényes ilyen õsi és levezetett tulajdonságpárok összesítését tar- talmazza az 1. táblázat.
A NÖVÉNYI TEST SZERVEZÕDÉSI FORMÁI,
SZIMMETRIAVISZONYAI ÉS ELÁGAZÁSI TÍPUSAI
Az egysejtûek, a telepesek és a hajtásos növények teste egyaránt sok- féle alakot ölthet a változatos környezeti feltételekhez való alkalmazko- dással és az örökletes változatossággal összefüggésben.
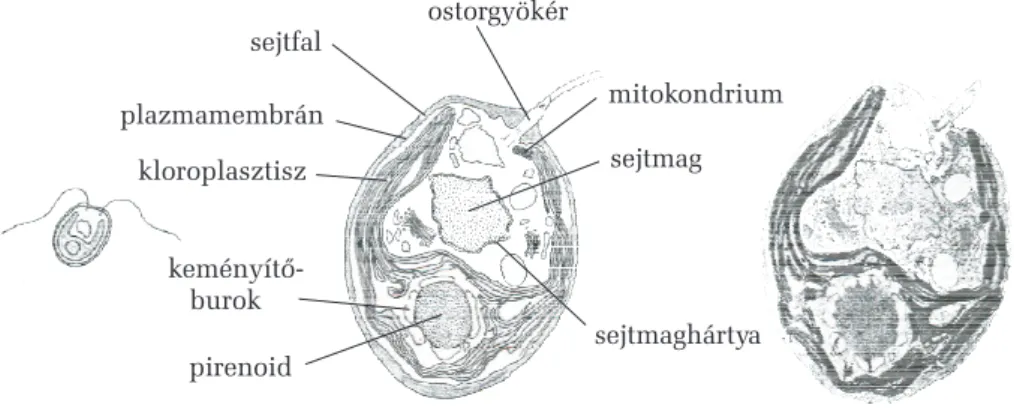
Az egysejtûek õsi formája sejtfal nélküli és önálló helyváltoztató mozgásra képes vízi élettérben, ostorok vagy állábak segítségével. Az os- tor és a fény irányának érzékelésében szerepet játszó szemfolt a sejt egyik, elülsõnek megfelelõ oldalán foglal helyet, így az ostoros sejtek már polaritást mutatnak (két szemben álló végük nem egyforma). Ennek kö- zönséges példája a zöldostoros. A polaritás a differenciálódás elõfeltéte- le, és alapja valamely anyagnak a más-más mennyisége (koncentráció- különbsége) az illetõ testrész két szemközti oldala között. Ha a sejtfal nélküli, alakját változtatni képes sejt több sejtmagot tartalmaz, melyek egyenértékûek, mert ugyanazon kezdeti sejtmag osztódásai során kelet- keztek, akkor plazmódiumról beszélünk. Más esetekben az egysejtû nö- vény merev sejtfallal rendelkezik, így állandó testalakot tart fenn, de önálló mozgásra képtelen. Az ilyen egysejtû lehet polaritás nélküli (pél- dául gömb alakú, ez az õsi forma), vagy a sejt elülsõ és hátsó része egy- mástól különbözõ lehet. Amikor több sejt összeolvadásából egy nagyobb sejt jön létre, melynek több, különbözõ eredetû sejtmagja van, tömlõtest- rõl beszélünk. Az egysejtûek és a többsejtûek közötti átmenetet azok a testfelépítési változatok képviselik, amelyek esetében önálló életû rokon egysejtûek valamilyen módon egymás mellett maradnak. Ennek egyik változata az, amelyben a sejtek az általuk kiválasztott nyálkaburokban csoportosulnak, más esetben a közös anyasejtbõl egyszerre megszületõ ikersejtek oldalfalaik által összetapadtan maradnak, továbbá vannak olyan egysejtûek, amelyek különbözõ korú rokon egyedekbõl álló sejtcsa- ládjában állandósuló sejtközi kapcsolatok vannak és a csoport különbö- zõ tagjai szakosodnak a táplálkozásra, a helyváltoztatásra és a szaporo- dásra. Ennek tipikus példája a gömbmoszat.
A valódi többsejtû telepes növények teste állhat egyszerû vagy elága- zó sejtfonalból (erre példa a békanyál, illetve az érdes békanyál), lapos le-
5. A NÖVÉNYI TEST SZERVEZÕDÉSI FORMÁI… 27 mez alakú sejttömegbõl (például a tengeri saláta) vagy a tér minden irá- nyába növekvõ masszív teleptestbõl (például a csillárkamoszat teste).
Néha a sejtfonalak annyira szorosan egymás mellé rendezõdnek (de nem kommunikálnak egymással), hogy álszövetes tömör test alakul ki. A fej- lettebb teleptest, habár belsõ felépítése minden részében egyforma, elkü- lönül gyökérszerû, szárszerû és levélszerû részekre, de ezek nem állnak valódi szövetekbõl. Ilyen számos nagytestû alga és sok moha telepteste.
A hajtásos növények teste valódi szakosodott szövetekbõl álló szer- vekre tagolódik, melyek közt kölcsönös függõségi viszony van. A vegeta- tív, önfenntartó szerepû szerveket a gyökér, a szár és a levél képviseli, a generatív vagy szaporítószervek lehetnek spóraképzõ és gamétaképzõ szervek, virágos növények esetében ezeket a virág részei képviselik, va- lamint a virág magkezdeményeibõl kialakuló magvak. Zárvatermõknél a virág magházából kifejlõdõ termés is a szaporodást szolgáló szerv.
Szimmetriájukat tekintve a növényi testrészek négyfélék lehetnek.
1. Asugarasvagy radiális szimmetriájú testrészek nagyon gyakoriak, ez a legõsibb szimmetria típus, amely esetében a testrész több szimmet- riasíkkal osztható két szemben álló, egyenértékû (egymás tükörképének megfelelõ, egymással fedõhelyzetbe hozható) félre. Ilyen szimmetriájuk van a hengeres száraknak, gyökereknek, levélnyeleknek, virágkocsá- nyoknak, a gömbölyû vagy hengeres magvaknak és terméseknek. A suga- ras szimmetriájú virágokat aktinomorf („csillag alakú”) virágoknak ne- vezzük. Ilyen például a magnólia, a tavirózsa, a boglárka, a rózsa, a káposztafélék (keresztesvirágúak), a liliomfélék virága.
2. Akétoldali vagy bilaterális szimmetriájú testrészek csak egyetlen szimmetriasíkkal rendelkeznek, mely általában a testrész hosszában ha- lad. Leggyakoribb esetben a jobb és a bal fél egyforma, de a felsõ és az al- só fél alakja egymástól eltérõ. Ilyen például a legtöbb levéllemez. A kétol- dali szimmetriájú virágokat zigomorf virágoknak nevezzük. Jellemzõek, többek között, a hüvelyesekre (pillangós virágok), az árvacsalánfélékre (ajakos virágok), az orchideákra (mézajakos virágok). A fészekvirágzatúak- nál (a napraforgó rokonainál) a csöves virágok sugaras, a steril nyelves vi- rágok pedig kétoldali szimmetriájúak.
3. A biszimmetrikus testrészeknek két, egymásra merõleges szim- metriasíkjuk van, így egymással egyforma egyrészt a bal és a jobb fél, másrészt pedig a felsõ és az alsó fél. Ilyenek például a fügekaktusz ella- posodott, nyelv alakú szárrészei vagy az elliptikus levéllemezek. Ez egy ritkább szimmetriatípus.
4. Azaszimmetrikustestrészek egyetlen szimmetriasíkkal sem oszt- hatók két egyforma félre, mert alakjuk szabálytalan. Ez is egy ritka válto- zat, ilyenek a begóniák és a szilfák levéllemezei, aCanna indicavirágai.
A növényi tengelyszervek általában elágaznak, hogy a tér minél több irányába terjesszék a testrészeket. Ezek az elágazási módok szabályos mintázatok formájában ismétlõdnek a növény élete során. Három fõ elága- zási típus ismeretes, és ezek mindegyikének két vagy három altípusa van.
1. Avillásvagy dichotomikus elágazás esetén közvetlenül a csúcs vá- lik ketté. Ez a legõsibb elágazási típus, ma algák teleptestén, kislevelû ha- rasztok (csipkeharasztok, korpafüvek) gyökerén és szárán, valamint egy õsi eredetû nyitvatermõ: a páfrányfenyõ (Ginkgo biloba) levélerezetében lelhetõ fel. Két altípusa van, aszerint, hogy a két ág egyforma vagy nem.
1a) Izotom villás elágazásról akkor beszélünk, amikor a csúcsból szétváló két ág egyforma és mindkettõ ugyanúgy ágazhat tovább.
1b) Anizotom villás elágazás esetében a csúcsból kialakuló egyik ág hosszabb, mint a másik, és csak ez a hosszabb ágazik el ismét egyenlõt- lenül.
2. A közalapos vagy monopodiális elágazás esetében a korlátlanul növekedõ, egységes közös tengely végighalad az egész elágazási rendsze- ren és csúcsa alatt bizonyos távolságokra jelennek meg az oldalágak.
Ezek általában az alaptól a csúcs felé haladva, csúcskeresõ (akropetális) irányban alakulnak ki egymás után, így a legalsó oldalágak a legidõseb- bek és a leghosszabbak. Ilyenek például a fürtös virágzatok és az egysé- ges törzsû fák szára. Aszerint, hogy a tengely hossza mentén ugyanazon a szinten hány oldalág képzõdik, három altípusa lehet.
2a) Váltakozó (altern) vagy szórt elágazás esetén egy szinten egyetlen oldalág ered.
2b) Átellenes (opponált) elágazáskor egymással szemben két oldalág található.
2c) Örvös (verticillát) elágazásról akkor beszélünk, amikor a csúcs alatti egyazon szinten több mint két oldalág fejlõdik a tér különbözõ irá- nyaiban.
3. Áltengelyes vagy szinpodiális elágazás esetében az eredeti ten- gelyrész korlátolt növekedésû, korán leáll és csúcsa alatt indul ki egy vagy több olyan ág, ami túlnövi az eredeti szakaszt. A jelenség ugyanígy többször megismétlõdik, így az elágazási rendszer tengelyét több egymás utáni, különbözõ szakasz építi fel. Aszerint, hogy a növekedésében meg-
5. A NÖVÉNYI TEST SZERVEZÕDÉSI FORMÁI… 29 álló csúcs alatt hány irányba folytatódik a hosszanti növekedés, ennek is három változata van.
3a) Monokázium esetében az eredeti csúcsot egyetlen oldalág növi túl.
3b) Dikázium vagy álvilla akkor alakul ki, amikor a fejlõdésében le- álló csúcs alatt két, szemben álló oldalág által folytatódik a növekedés.
3c) Pleiokáziumról akkor beszélünk, amikor a növekedését nem foly- tató csúcsot a közvetlenül alatta megjelenõ oldalágak több mint két irányba növik túl.
A korlátlan növekedés és a sorozatos elágazás által egyes növények tes- te valóságos térbeli és idõbeni rekordokat ér el az élõvilág egészére vonat- koztatva. Ezt az alábbiakban néhány példával szemléltetjük. A leghosszabb testû ma élõ szervezetek aMacrocystisnemzetségbe tartozó tengeri barna- algák, melyek telepteste eléri a 300 m hosszúságot. A leghosszabb hajtásos növény aCalamus rotangnevû trópusi lián, melynek tekeredõ szára eléri a 170 métert. A legmagasabb ma élõ fák az eukaliptuszok, melyeknek törzs- magassága 125 és 150 méter között van. A mammutfenyõk (Sequoia gigantea) a legsúlyosabb élõlények, testük több száz tonna lehet. A leghos- szabb életû lények szintén növények, észak-amerikai zord éghajlatú magas- hegyi fennsíkokon élõ, nagyon lassú anyagcseréjû és nagy ellenálló képes- ségû fenyõk (Pinus longeviva), egyes példányok 4600 évnél idõsebbek.
A leghosszabb életû levél a Namíb-sivatagban élõ õsi típusú nyitvatermõé (Welwitschia mirabilis), mely ugyanazzal a két, folyamatosan növekvõ levél- lel él több száz évig. A legszélesebb virág aRafflesia arnoldiinevû trópusi parazita növényé, a kinyílt virág átmérõje 1 méter. A legmagasabb: 1,5 mé- teres virága az elõbbi növény egyik közeli rokonának van (egy Amor- phophallus faj). A legnagyobb mag 6 kg, tulajdonosa a maldív (Lodoicea seychelliana) pálmafa, a leghosszabb terméssel pedig a tengeri bab (Entada scandens) rendelkezik (1,5 métert elérõ hüvelytermés).
ANALÓG, HOMOLÓG ÉS METAMORFOZÁLT NÖVÉNYI TESTRÉSZEK
Mind a törzsfejlõdéstanilag rokon fajok közös örökletes programja, mind a konkrét környezeti feltételekhez való alkalmazkodás rányomja bé- lyegét a növényi testrészek szervezõdésére. A fenotipikus tulajdonságok szintjén a genotípus és a környezet kölcsönhatásában gyakran az egyik- nek erõteljesebb nyomai látszanak a szervek alakjában és belsõ szerkeze- tében, mint a másiknak. A környezet módosító hatása leginkább olyan helyzetekben nyilvánul meg, amikor bizonyos növények evolúciós alkal- mazkodásuk során szélsõségesen specializálódnak sajátos élõhelyeken va- ló sikeres fennmaradás céljából, és ekkor a jellegzetes életmód szerkezeti alapja egy tipikusan alakult testforma lesz, aminek ismerete fontos az al- kalmazkodási stratégiák megismerésében. Ilyen adaptív bélyegek szemlél- tetõi az analóg, a homológ és a metamorfozált növényi testrészek.
Azanalóg testrészekeredete különbözõ, de azonos környezeti feltéte- lekhez való egyforma alkalmazkodás során egymáshoz nagyon hasonlóvá váltak. Ebben az esetben tehát nem a rokonság, hanem a környezet a meg- határozó. Az analóg testrészek az összetartó vagy konvergens evolúció so- rán jöttek létre. Például, egymással analóg az afrikai sivatagok kutyatej fa- jainak és az amerikai sivatagok kaktuszainak oszlopos, pozsgás, aszszimiláló és vízraktározó szára. Emiatt morfológiai szempontból nem lehet felismerni, hogy az adott növény kutyatej-e vagy kaktusz. Másik pél- da a szõlõkacs és a borsókacs: mindkettõ kapaszkodásra szolgál, és az azo- nos szerep hatékony betöltéséért hasonlóvá váltak, de a szõlõkacs a szár, a borsókacs pedig a levél egyik részébõl alakult ki, tehát eredetük eltérõ.
Ugyanígy egymással analóg a kaktusz, az akác és a kökény tövise: mindhá- rom egyforma felépítésû, mert ugyanazt a mechanikai védõ szerepet szol- gálja, de a kaktuszé levélbõl, az akácé levélalapi pálhából, a kökényé pedig szárból származik. A lombos mohák levélkéi a hajtásos növények levelei- vel analóg teleprészek, levélszerûen lapos asszimiláló testrészek, de a te- leptestnek, nem a hajtásnak a részei. A rucaöröm (Salvinia natans) nevû, vízen úszó páfrány minden szárcsomóján a három levél közül az egyik a gyökerekkel analóg, mert csak elágazó levélerezete nyúlik alá és szívja fel
6. ANALÓG, HOMOLÓG ÉS METAMORFOZÁLT NÖVÉNYI TESTRÉSZEK 31 a vizet az ásványi sókkal, éppúgy, mint a többi növény gyökere. A szúrós csodabogyó korlátolt növekedésû oldalágai (szárrészei) a levelekkel analó- gok, melyekhez a megtévesztésig hasonlítanak, csakhogy szárazság idején jobban gazdálkodnak a vízzel, mint a levelek, mert kevesebbet párologtat- nak, de ugyanakkora fényfelfogó és asszimiláló felületet biztosítanak.
A homológ testrészek közös eredetûek, ezért felépítési alaptervük azonos, de a hosszas széttartó vagy divergens evolúció során, különbözõ környezeti körülményekhez és eltérõ életmódokhoz, illetve más-más mûködési szerep hatékonyabb betöltésére alkalmazkodva, végsõ megjele- nési formájuk nagyon különbözõvé vált. Például, egymással homológ szervek a kaktusz tövise, a lednek kacsa és a kancsóka rovarfogó csapdá- ja, hiszen mindhárom levéleredetû, de más-más irányba (védelemre, ka- paszkodásra, illetve rovaremésztésre) szakosodtak alkalmazkodásuk so- rán, így már nem hasonlítanak egymásra, habár kialakulásukkor a fiatal levél jellegeit mutatják. Szintén egymással homológ a virág csészelevele, sziromlevele, termõlevele és porzója: mindegyik levélbõl származik, de különbözõvé váltak, mert más-más szerep betöltésére szakosodtak, így eltérõ irányokba fejlõdtek tovább.
A metamorfozált szervek alakja és szerkezete erõteljesen átalakult az illetõ szervtípus általános megjelenési formájához viszonyítva, mert szélsõségesen alkalmazkodtak egy-egy speciális mûködési szerep betölté- sére, ami eltér a szerv közönséges funkciójától. Figyelmes ontogenetikai és anatómiai vizsgálattal a metamorfozált testrész szervi hovatartozása felismerhetõ, hiszen az illetõ szerv alapvetõ általános szervezõdési bé- lyegeit, ha módosulások ráépülésével is, de megõrzi, csakhogy a sajátos szerep szempontjából fontos szövetelemek kerülnek túlsúlyba. Például, a retek és a sárgarépa fogyasztható része módosult, raktározó gyökér, a bur- gonya gumója metamorfozált földbeni szárrész, a kaktuszok tövise módo- sult levél, a karfiol „feje” metamorfozált virágzat.
A NÖVÉNYI SEJT SAJÁTOS ALKOTÓI
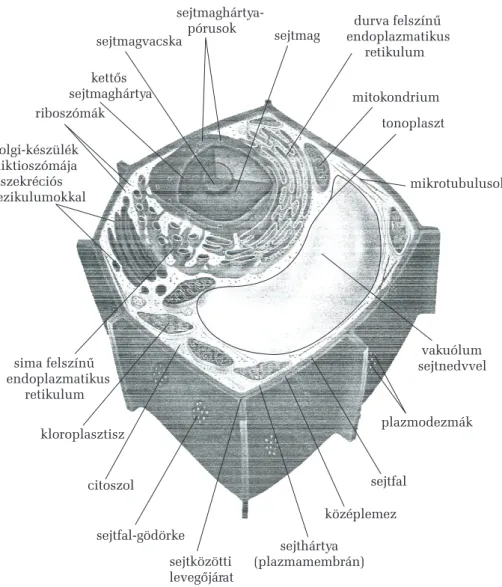
Az eukarióta sejtek általános alkotói mellett (sejthártya vagy plazma- membrán, sejtmag sejtmagvacskával és kromatinállománnyal, riboszó- mák, sima és riboszómákat viselõ, érdes endoplazmatikus retikulum, diktioszómák által képviselt Golgi-készülék, mitokondriumok, alapcito- plazma vagy citoszol és a benne levõ sejtvázelemek: mikrotubulusok, mikrofilamentumok és intermedier filamentumok, esetleg ostor vagy csillók) a növényi sejtekben egyéb, sajátos összetevõk is jelen lehetnek.
Ilyenek a színtestek vagy plasztiszok, melyek kizárólagosan bizonyos nö- vényi sejtekben fordulnak elõ, a jellegzetes felépítésû sejtfal (mely más- fajta, mint a baktériumok és a gombák sejtfala), a vakuólumok (melyek sajátos lizoszómáknak felelnek meg), a mikrotestek sajátos növényi for- mái (levélperoxiszómák, szferoszómák, glioxiszómák stb.) és a növé- nyekre jellemzõ anyagcsere-zárványok (keményítõszemcsék, olajcsep- pek, aleuronszemcsék, kalcium-oxalát kristályok stb.). A növényi sejt általános felépítését a 2. ábra szemlélteti.
A színtestekvagy plasztiszok fõ szerepe a szerves tápanyagok foto- szintetikus elõállítása és raktározása. Két burkolómembránnal rendelkez- nek, melyek közül a belsõnek az áteresztõ képessége sokkal szelektívebb, mint a külsõé. Folyékony belsõ alapállományuk a sztróma, ez a plasztisz típusától függõen többé vagy kevésbé fejlett belsõ membránrendszert vesz körül. Prokarióta eredetûek, ezt bakteriális típusú örökítõ anyaguk, fehérjeszintetizáló rendszerük és amitotikus kettéosztódási képességük tükrözi (legalábbis bizonyos fejlõdési stádiumokban). Több típusuk is- mert, ezek meghatározott irányban egymásba átalakulhatnak a fejlõdés során. Legfontosabb formáik a proplasztiszok, az etioplasztiszok, a kloro- plasztiszok, a kromoplasztiszok és a leukoplasztiszok.
1. A proplasztiszok az osztódó, merisztematikus sejtekben vannak jelen. Színtelenek, kettéosztódásra képesek, még nincs kifejlett belsõ membránrendszerük, de ez fokozatosan kialakulhat a belsõ burkoló- membrán egyes szakaszainak a színtest hossztengelyével párhuzamos, megnyúlt kesztyûujjszerû betüremkedésével. Proplasztiszokból alakul ki a differenciálódás során az összes többi színtest.
7. A NÖVÉNYI SEJT SAJÁTOS ALKOTÓI 33
2. Az etioplasztiszok fény hiányában kifejlõdõ (fehéressárga színû, etiolált) hajtásokban jelennek meg a proplasztiszokból. Belsejükben sza- bályosan egymás mellé tömörülõ csövecskékbõl álló, kristályos elrende- zõdésû membránrendszer van a klorofill színtelen elõanyagával. Ez a prolamelláris test, mely a belsõ burkolómembrán betüremkedéseibõl szár-
kettõs sejtmaghártya riboszómák
sejtmagvacska
sejtmaghártya-
pórusok sejtmag
mitokondrium tonoplaszt
mikrotubulusok Golgi-készülék
diktioszómája szekréciós vezikulumokkal
endoplazmatikusínû retikulum durva felsz
sima felszínû endoplazmatikus
retikulum
citoszol
kloroplasztisz plazmodezmák
sejthártya (plazmamembrán) sejtközötti
levegõjárat
középlemez sejtfal-gödörke
sejtfal
vakuólum sejtnedvvel
2. ábra.A növényi sejt térmodellje (Mauseth 1995 nyomán)
mazik. Fény jelenlétében az etioplasztiszok kloroplasztiszokká alakulnak, miközben a hajtásrészek kizöldülnek.
3. A kloroplasztiszok vagy zöld színtestek proplasztiszokból vagy etioplasztiszokból képzõdnek. Bennük történik a fotoszintézis: a fényener- gia elnyelése a klorofillok és karotenoid pigmentek által, a fényenergia át- alakítása kémiai energiává, a vízbontás és az ezzel járó oxigéntermelés, a szén-dioxid asszimilációja az átalakított fényenergia segítségével, ami új, energiadús szerves anyagok képzõdését eredményezi. A kloroplasztiszok belsejében zárt, lapos hólyagokat vagy korongokat alkotó sajátos membrán- rendszer található, ez a tilakoidrendszer. Egy tilakoidkorong falát a tila- koidmembrán képezi, ez elhatárol a színtest sztrómájától egy folyadékkal telt belsõ teret, melyet tilakoidlumennek nevezünk. Egyes tilakoidkorong szakaszok egyenként végignyúlnak a sztróma bizonyos övezetében, ezek a sztrómatilakoidok (külsõ felületük érintkezik a plasztisz alapállományával).
Helyenként a tilakoidmembránok többszörös kitüremkedéssel egymást fedõ rövidebb korongok (gránumkorongok) tekercseként rendezõdnek. Itt az egy- másra tapadó membránszakaszok nem érintkeznek a sztrómával, ezért eze- ket gránumtilakoidoknak nevezzük. A gránum- és sztrómatilakoid szaka- szok egymással folytonossági (kontinuitási) viszonyban vannak. Az egymásra tevõdõ tilakoidkorongok kötege a gránum. Az algák kloroplasz- tiszaiban nincsenek gránumok, itt csupán egyenként haladó, hosszú tilakoidlemezek vannak jelen, ezért az algák színtestjeit lemezes típusúak- nak nevezzük. Másik sajátossága az algasejtek plasztiszainak, hogy bennük jellegzetes fehérjetest: a pirenoid lehet jelen. A gránumos plasztiszok a mo- hákra és a hajtásos növényekre jellemzõek.
A tilakoidmembránokba ágyazottan, fehérjékhez kötött elrendezõ- désben vannak jelen a fényelnyelõ klorofillmolekulák (zöld pigmentek) és a sárga karotenoid pigmentek (karotének és xantofillok). Ebbõl követ- kezik, hogy egy zöld növényben csak a kloroplasztiszok belsejében levõ membránok: a tilakoidmembránok zöldek. Ezekben történik a fényener- gia megkötése és átalakítása, amihez a vízbontás (oxigénfelszabadítás), redukáló erõ (a szerves anyagokhoz szükséges hidrogén) képzõdése és ATP-szintézise társul. Mindez a fotoszintézis fényszakaszát képviseli.
A sztrómában találhatók az alapanyagok és az enzimek az új szerves anyagok felépítéséhez, itt zajlik a szénasszimilációs anyagcsere, mely ál- talában szintén nappal megy végbe, akár a fényszakasz is (ezért helyte- len sötétszakasznak nevezni).
A kloroplasztisznak bakteriális típusú, körkörös kromoszómája van, ez számos egyforma példányban (amplifikált génekkel) van jelen a sztró-
7. A NÖVÉNYI SEJT SAJÁTOS ALKOTÓI 35 mában. Génjei anyai ágon öröklõdnek, mert a zigóta kialakulásakor csak a petesejtbõl jutnak át színtestek az utódba. A színtestnek saját, prokari- ótákra jellemzõ típusú riboszómái is vannak, így fehérjeszintézisre ké- pes. Az eukarióta sejttel való hosszas együttélés (endoszimbiózis) során a kloroplasztisz számos génje átköltözött a sejtmagba, így a színtest szá- mos fehérjéjét importálja a citoszolban levõ riboszómák felõl.
A nappal folyamán képzõdõ szerves anyagokból felgyûlõ ideiglenes fölösleget az elszállításig a kloroplasztisz a sztrómában megjelenõ kemé- nyítõszemcsék formájában tárolja. A sztrómában karotenoid tartalékot magába foglaló olajos cseppek: plasztoglobulusok is lehetnek.
4. Akromoplasztiszoka fotoszintetikus pigmentek közül csak karote- noidokat tartalmaznak, ezért sárga, narancssárga vagy téglavörös színûek.
Klorofillok hiányában fotoszintézisre nem képesek, belsõ membránrend- szerük kevésbé fejlett, mint a kloroplasztiszoké. Fõleg virágszirmokban, érett húsos termések falában gyakoriak.
5. A leukoplasztiszok színtelenek, nem tartalmaznak fényelnyelõ pigmenteket. Belsõ membránrendszerük fejletlen. Szerepük a tartalék tápanyagok raktározása, ezért gyökerekben, földbeni szárakban, magvak- ban és termésekben gyakoriak. Keményítõt, olajat vagy fehérjét raktároz- hatnak. Leggyakoribb formájuk az amiloplasztisz, mely megszilárduló keményítõszemcsét képez.
A sejtfal a plazmamembránt kívülrõl körülvevõ vastag, élettelen, rostos szerkezetû növényi sejtalkotó. Alapszerkezetét amorf alapanyagba ágyazott cellulózrostok rétegezett hálózata képezi, melybe utólag a szö- vetek differenciálódása során jellegzetes szilárdító- vagy szigetelõanya- gok rakódhatnak. A sejtfal elõanyagait a Golgi-készülék választja ki szek- réciós vezikulumokba csomagoltan a sejt felszínére. A sejtosztódás során keletkezõ két leánysejt között kialakuló közös válaszfal a középlemez, mely összecementezi a szomszédos sejteket, fõ összetevõje a kalcium- pektát. Erre rétegezi mindegyik sejt a saját belsõ tere felõl az elsõdleges sejtfalát, mely fõleg pektinbõl és hemicellulózból álló amorf alapállo- mányba (mátrixba) ágyazódó cellulózrostok kötegeibõl: mikrofibrillu- mokból áll. Ez a sejtfal még viszonylag vékony és áteresztõ. Az elsõdle- ges sejtfal után, ennek belsõ felülete mentén, kialakulhat egy kívülrõl befele (centripetális irányban) vastagodó másodlagos sejtfal. Ez cellulóz- rostokban még gazdagabb, mint az elsõdleges, és legalább három, egy- mást borító rétegbõl áll, melyekben a cellulózrostok kötegeinek elrende- zõdési iránya eltérõ. A sejtfalnak nemcsak a vastagsága, hanem vegyi összetétele is változhat. Ilyenkor a cellulóz mikrofibrillumok közé mere-