Cisztás fibrózis módosító genetikai faktorok lehetséges szerepe kongenitális bilateralis vas
deferens hiányban Doktori tézisek Dr. Havasi Viktória
egyéni felkészülő Semmelweis Egyetem
Eméleti Orvostudományok Doktori Iskola
Konzulens:
Dr. Zsembery Ákos, egyetemi docens, Ph.D.
Hivatalos bírálók:
Dr. Garami Miklós, egyetemi docens, Ph.D.
Dr. Váradi András, egyetemi tanár, Ph.D., D.Sc.
Szigorlati bizottság elnöke:
Dr. Sasvári Mária, egyetemi tanár, Ph.D., D.Sc.
Szigorlati bizottság tagjai:
Dr. Csanády László, egyetemi docens, Ph.D.
Dr. Fodor László, kutató-fejlesztő, Ph.D.
Budapest
Bevezetés
A cisztás fibrózis (CF) a kaukázusi rassz leggyakoribb autoszomális recesszív öröklődésű kórképe, amely a tüdő, a pankreász, a máj, és az exokrin mirigyek érintettségével jár. A cisztás fibrózis tüneteinek kialakulásáért a cisztás fibrózis transzmembrán konduktancia regulátor (CFTR) gén mutációi felelősek. A génnek ezidáig több mint 1900 variánsát azonosították. Számos CFTR mutáció szerepe feltételezett nemcsak CF-ben, hanem egyéb CFTR génhez kapcsolt kórállapotok úgymint disszeminált bronchiectasia, allergiás bronchopulmonáris aszpergillózis, akut rekurrens vagy krónikus idiopathiás pankreatitisz, alkoholos pankreatitisz, és kongenitális bilaterális ductus deferens hiány (CBAVD) kialakulásában is. A CBAVD kétoldali ductus deferens hiány következtében jön létre, normális spermatogenezissel és obstructív azoospermiával jár, és a férfi sterilitásos esetek mintegy 2-3%-ának áll a hátterében. A CBAVD pontos pathogenezise nem tisztázott, de a kórkép bizonyítottan CFTR mutációkkal társul. A CFTR szekvencialtérések nagy száma miatt azonban, számos génhiba kóroki szerepe a CBAVD-ben egyelőre nem tisztázott. A CFTR gén F508del mutációja - 508-as pozícióban lévő fenil-alanin aminosav deléció - a leginkább vizsgált és leggyakoribb CF mutáció. Az F508del-n kívül egy másik mutációt - F508C (fenil-alanin cisztein csere)
- is leírtak ugyanebben az aminosav-pozícióban, ezt azonban olyan tünetekkel nem járó polimorfizmusnak véltek, amely sem a CF, sem a CBAVD kialakulásához nem járul hozzá.
Kísérletes adatok szerint az F508del csökkenti a protein folding hatékonyságát és korai CFTR fehérje degradációt eredményez, míg az F508C variáns lehetővé teszi a CFTR fehérje teljes érését. Du és munkatársai azt tapasztalták, hogy in vitro körülmények között az F508C-CFTR protein eléri a sejtfelszínt és mérhető Cl- ioncsatorna funkciót mutat. Cui és munkatársai pedig azt találták, hogy az F508C aminosavcsere megváltoztatja a CFTR Cl- ioncsatorna kapu-aktivitását.
Mivel még az egypetéjű iker CF-es betegek tünetei is jelentős variabilitást mutatnak, a CF létrejöttében szerepet játszó genetikai módosító faktorok vizsgálata intenzív kutatás tárgyát képezi. Drumm és munkatársai összefüggést mutattak ki a transzformáló növekedési faktor (TGFB-1) gén 10. kodon CC genotípusának (rs1800470 korábban rs1982073) gyakorisága és a tüdőtünetek súlyossága között homozigóta CF betegek esetében. Egy, a TGFB-1 gén 25. kodonjára lokalizálódó polimorfizmus (rs1800471) szerepe szintén felmerült a CF tüdő manifesztációjának befolyásolásában.
Emellett Darrah és munkatársai szoros korrelációt mutattak ki az A-típusú endothelin receptor (EDNRA) gén két polimorfizmusa (rs1801708, rs5335) és a cisztás fibrózis tüdő
manifesztációjának súlyossága között. A CBAVD-s betegek egy kis hányadában nem lehet CFTR génhibát azonosítani, azonban az érintettek mintegy 80-97%-a egy, 50-93%-a pedig két CFTR mutációt hordoz. Egy CFTR mutáció jelenléte azonban nem minden esetben vezet CBAVD-hez, ugyanis férfiak milliói bírnak egy hibás CFTR génnel, mégsem alakul ki náluk ezen kórkép. Ezek alapján feltételezhetjük, hogy a CFTR mutációkon kívül egyéb tényezők is szükségesek a kórkép kialakulásához, azonban ezek ma még nem ismertek.
Mindezek alapján, kísérleteink fő célja az volt, hogy cisztás fibrózis genetikai módosító faktorok CBAVD-ben betöltött lehetséges szerepét megvizsgáljuk.
Célkitűzések
Vizsgálataink központi hipotézise az volt, hogy a) egy
„enyhe” CFTR mutáció (F508C) hozzájárulhat a CBAVD fenotípushoz és b) cisztás fibrózis módosító gének nemcsak a CF manifesztációját befolyásolják, hanem hozzájárulnak a CBAVD kialakulásához is. Céljaink közt szerepelt, hogy:
1. A CFTR gén F508C variánsának gyakoriságát és feltételezett kórokozó szerepét CF hordozó, cisztás fibrózis és CBAVD-s betegekben meghatározzuk.
2. Ismert cisztás fibrózis módosító polimorfizmusok - rs18004700, rs1800471, rs1801708, rs5335 - szerepét CBAVD-s beteganyagon meghatározzuk.
3. Bebizonyítsuk azt a feltételezést, miszerint CF módosító gének a CBAVD kórkép penetranciáját is befolyásolják.
Beteganyag és módszerek
F508C polimorfizmus vizsgálatunkban azon férfiak mintáit használtuk fel analízisra, akiknél Ambry TestTM:CF genetikai teszt felhasználásával teljes CFTR génanalízis történt 2002.
január és 2007. június között az Ambry Genetics laboratóriumában, Kaliforniában. A vizsgálati populációt 6970 férfi alkotta, akik közül 850-et CF hordozó szűrésre, 5938-at cisztás fibrózis megalapozott gyanújával és 182-t CBAVD klinikai diagnózisával küldtek genetikai analízisre. A CBAVD genetikai vizsgálatát minden esetben fizikális vizsgálat előzte meg. Kutatásunkat az érintett betegek beleegyezésével végeztük. A genomiális DNS izolálása leukocytákból történt, majd az izolálást követően a DNS minőségi és mennyiségi vizsgálatát agaróz gélelektroforézissel végeztük. A minták analízise Ambry TestTM:CF használatával történt, amelynek során a minta analízise először módosított hőmérséklet grádiens elektroforézissel (mTTGE), majd ezt követően DNS szekvenálással történik. A teszt a CFTR gén összes exonjának
vizsgálatára, valamint az exon-intron határokon lévő 20 bázis és néhány fontosabb intronban elhelyezkedő mutáció azonosítására alkalmas. A vizsgálni kívánt exonok és intronikus régiók felerősítése polimeráz láncreakcióval (PCR) történt. A PCR termékeket először denaturáltuk majd - a maximális heteroduplex kialakulásához szükséges optimális körülményeket biztosítva - lassan lehűtöttük. A felerősített szakaszokat ismert, normális CFTR szekvenciájú DNS mintákkal kevertük össze, hogy az eltéréseket analizálni tudjuk. A PCR termékeket ezután DCode rendszeren módosított hőmérséklet grádiens elektroforézisnek vetettük alá. A minták elektroforézise után a poliakrilamid géleket ethídium-bromiddal festettük meg, majd UV fénnyel megvilágítva GelDoc rendszerben értékeltük ki. Amennyiben az ismert szekvenciájú kontrollokhoz képest eltérés volt a viszgált PCR produktum mintázatában, a kérdéses minta DNS szekvenciája meghatározásra került. A mintákat egy újabb PCR reakcióban szekvenáló primerekkel felerősítettük, majd ExoSAP-It kittel megtisztítottuk. A szekvenálás standard láncterminációs módszerrel történt Beckman Coulter CEQ8000 kapilláris elektroforézis készüléken.
A CBAVD módosító gén-vizsgálatunk során 80 CBAVD-s beteg valamint 51 kontroll férfi DNS mintáját analizáltuk.
Ebből 19 beteg és 20 kontroll minta Spanyolországból, 31
CBAVD-s és 31 kontroll minta Törökországból, és további 30 CBAVD-s minta Portugáliából származott. A vizsgálatok során portugál kontroll minták nem álltak rendelkezésünkre.
Kutatásunkhoz olyan CBAVD-s betegek DNS mintáit használtunk, akiknél legalább egy CFTR gén mutáció volt kimutatható. A kontroll csoportba azonos régióból származó, egészséges férfiak mintái kerültek. A vizsgálati protokollt mind a University of Alabama at Birmingham, mind pedig a helyi portugál, spanyol és török egyetemek etikai bizottságai jóváhagyták, és a résztvevők hozzájárultak ahhoz, hogy mintájukat kutatásunkhoz felhasználhassuk. A TGFB-1 gén (NT_011109) 5’ végén elhelyezkedő 453 bázis hosszúságú szakaszát az alábbi saját tervezésű primerek felhasználásával erősítettük fel: 5’-GAGGACCTCAGCTTTCCCTC-3’
(forward) és 5’-CTCCTTGGCGTAGTAGTCGG-3’
(reverse). A felerősített szakasz mind az rs1800470, mind pedig az rs1800471 TGFB-1 génpolimorfizmusokat magában foglalta. Az EDNRA gén (ENSG00000151617) promoterének 480 bázist felölelő szakaszát, amely az rs1801708 polimorfizmust foglalja magába, a következő saját tervezésű
primerekkel erősítettük fel: 5’-
GTGGAAGGTCTGGAGCTTTG-3’ és 5’-
TTCCCAGCTCTCGTCTTCTC-3’. Az EDNRA gén 8.
exonjában lévő SNP (rs5335) detektálásához az alábbi
(szintén saját) primereket alkalmaztuk: 5’-
CTGCTGCTGTTACCAGTCCA-3’ és 5’-
TGACCAGTTCCCATTGAACA-3’. A PCR reakció szekvenálás előtti tisztítását QIAquick PCR Purification Kittel végeztük, majd a tisztított PCR termékeket a szekvenálás előtt BigDye Terminator v. 3.1 Cycle Sequencing Ready Reaction kit-tel erősítettük fel. A szekvenálási reakció a gyártó előírásai szerint történt Applied Biosystems 3730 Genetic Analyzer készülékén, POP-7 polimer alkalmazásával.
A szekvenciák analízisét és referenciával történő összeillesztését a Chromas Lite és Clustal W Multiple Sequence Alignment szoftverekkel végeztem.
Az F508C allél-frekvencia különbségek jelentőségét hármas kontingencia teszttel határoztuk meg az alábbi csoportokban: I. cisztás fibrózis gyanújával genetikai vizsgálatra küldött minták; II. CFTR mutáció hordozó-szűrés csoport; és III. klinikailag igazolt CBAVD csoport. A statisztikai analízisek a VassarStats internet alapú programjával történtek. Mivel három vizsgálati csoportunk közül kettőben az F508C variáns kevesebb mint 5 mintában volt jelen, hármas Monte Carlo szimulációs tesztet futtattunk 10000 mintára. Emellett páronkénti összehasonlításokat és esélyhányadosok meghatározását végeztük el.
Az EDNRA és TGFB-1 polimorfizmusok genotípus eloszlásának összehasonlítása χ2 próba, az allél-frekvencia különbségek meghatározása pedig t-próba alkalmazásával történt. Az összehasonlításokat az összes beteg és összes kontroll csoportok, valamint az egyes alcsoportok (spanyol, portugál, török) között is elvégeztük. A statisztikai analíziseket SPSS statisztikai szoftvercsomag felhasználásával végeztük.
Eredmények
A CF hordozó-szűrés csoportban (n=850) 3 esetben detektáltuk egyedüli eltérésként az F508C variánst. Ebben a csoportban nem találtunk olyan mintát, amelyikben az F508C eltérés egy másik CFTR mutációval társult volna. A klinikailag bizonyított CBAVD csoport (n=182) analízise során 3 betegben az F508C mutáció egy második pathológiás CFTR eltéréssel társult (1. Táblázat).
1. Táblázat. F508C hordozó CBAVD-s betegek
Beteg Mutáció 1 Mutáció 2 Klinikum Életkor
1 F508del F508C CBAVD 39
2 F508del F508C CBAVD, sinusitis, asztma
34
3* L206W F508C CBAVD 40
A CF klinikai gyanúval genetikai vizsgálatra küldött (n=5938) csoportban 18 esetben az F508C variáns volt az
további mintájában az F508C eltéréshez egy másik kórokozó CFTR mutáció is társult (2. Táblázat).
2. Táblázat. CF gyanús F508C hordozó betegek Beteg Mutáció
1
Mutáció 2
Klinikum Életkor 1 G551D F508C Pozitív újszülöttkori
szűrés
>1 hónap 2 V574M F508C CF alapos klinikai
gyanúja
1 hónap 3 F508del F508C CF alapos klinikai
gyanúja 24 év
4 F508del F508C CF alapos klinikai
gyanúja 11 év
5 F508del F508C CF alapos klinikai
gyanúja 32 év
Az F508C allél-frekvencia a klinikailag igazolt CBAVD csoportban mintegy 4,68-szor volt magasabb, mint a CF hordozó-szűrés populációban (3. Táblázat). A kétirányú χ2 próba azt mutatta, hogy az F508C variáns frekvenciája szignifikánsan magasabb a CBAVD-s csoportban, mint bármelyik más vizsgálati populációban. (3. Táblázat) Ezen hármas összehasonlítás során, a CBAVD csoport F508C allél- frekvenciája (3/363) szignifikánsan magasabbnak bizonyult mind a CF hordozó-szűrés csoport (3/1700), mind pedig a CF klinikai gyanúval vizsgálatra küldött csoport (23/11876) allél- frekvenciájánál (χ2=6,95, p=0,031). A 10000 mintás Monte Carlo szimuláció 0,0486 kummulatív valószínűséget eredményezett.
3. Táblázat. Az F508C allélek megoszlása vizsgálati csoportjainkban
CBAVD (F508C egy
másik mutációval)
CF hordozó
szűrés (csak F508C )
CF gyanús minták
Összes F508C egy
másik CFTR mutációval
Csak F508C mutáció F508C
mutációt hordozó betegek
3 3 5* 18 29
Összes beteg Allélek
182 364
850 1700
5938 11876
5938 11876
6970 13940 F508C
allél- frekvencia
0,824% 0,176% 0,042% 0,151% 0,208%
0,193%
χ2, (p) 2,38
(0,073)
21,94 (0,002)
5,87 (0,023) 4,01 (0,04) Esélyhánya
dos (OR) 4,701 4,283
A χ2 és az esélyhányados értékek a klinikailag igazolt CBAVD csoport és a CF hordozó-szűrés csoport, valamint a klinikailag igazolt CBAVD csoport és az összes többi betegcsoport közötti összehasonlításokra vonatkoznak. A p értékek a 10000 mintás Monte Carlo szimulációsösszehasonlításra lettek korrigálva.
*Az F508Cmutáció mellett egy további pathológiás CFTR mutációt hordoznak.
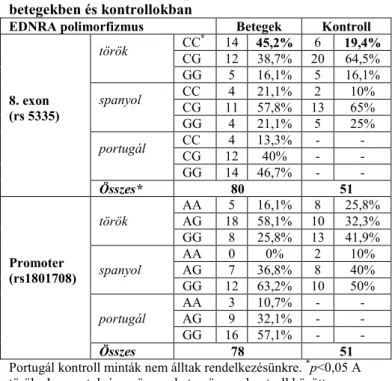
Módosító gén tanulmányunkban az EDNRA gén rs5335 SNP CC genotípus emelkedett gyakoriságát találtuk a CBAVD-s betegcsoportban (4. Táblázat). A CC genotípus szignifikánsan gyakrabban fordult elő a legnagyobb létszámú alcsoportunkban (török betegek 45,2% vs. török kontrollok 19,4%, p<0,05 χ2-próba), és az összes beteg és összes kontroll közötti különbség megközelítette a statisztikai szignifikanciát
(27,5% versus 15,7%, p=0,06). Az EDNRA promoter SNP (rs1801708) nem befolyásolta a CBAVD penetranciát (p=0,39) (4. Táblázat).
4. Táblázat. EDNRA genotípusok eloszlása CBAVD -s betegekben és kontrollokban
EDNRA polimorfizmus Betegek Kontroll
8. exon (rs 5335)
török CC* 14 45,2% 6 19,4%
CG 12 38,7% 20 64,5%
GG 5 16,1% 5 16,1%
spanyol CC 4 21,1% 2 10%
CG 11 57,8% 13 65%
GG 4 21,1% 5 25%
portugál CC 4 13,3% - -
CG 12 40% - -
GG 14 46,7% - -
Összes* 80 51
Promoter (rs1801708)
török
AA 5 16,1% 8 25,8%
AG 18 58,1% 10 32,3%
GG 8 25,8% 13 41,9%
spanyol
AA 0 0% 2 10%
AG 7 36,8% 8 40%
GG 12 63,2% 10 50%
portugál
AA 3 10,7% - -
AG 9 32,1% - -
GG 16 57,1% - -
Összes 78 51
Portugál kontroll minták nem álltak rendelkezésünkre. *p<0,05 A török alcsoportok és az összes beteg-összes kontroll között
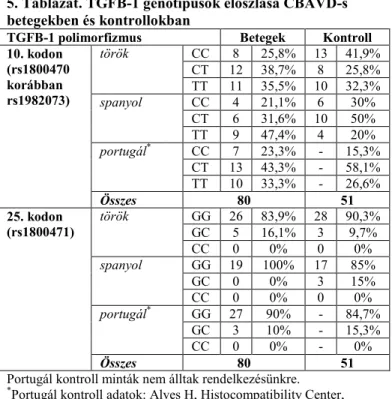
Az rs1800470 (TGFB-1 10. kodon) SNP T allélja nagyobb gyakorisággal fordult elő a CBAVD betegek között, mint a kontroll csoportban (összes beteg: 57%, összes kontroll:
45%), de szignifikáns különbséget nem sikerült kimutatni sem az alcsoportok és megfelelő kontrolljaik, sem az összes beteg
és összes kontroll összehasonlítása során. A TGFB-1 25.
kodonra lokalizálódó SNP (rs1800471) és a kongenitális ductus deferens hiány között semmilyen asszociációt nem tudtunk azonosítani (5. Táblázat).
5. Táblázat. TGFB-1 genotípusok eloszlása CBAVD-s betegekben és kontrollokban
TGFB-1 polimorfizmus Betegek Kontroll 10. kodon
(rs1800470 korábban rs1982073)
török CC 8 25,8% 13 41,9%
CT 12 38,7% 8 25,8%
TT 11 35,5% 10 32,3%
spanyol CC 4 21,1% 6 30%
CT 6 31,6% 10 50%
TT 9 47,4% 4 20%
portugál* CC 7 23,3% - 15,3%
CT 13 43,3% - 58,1%
TT 10 33,3% - 26,6%
Összes 80 51
25. kodon (rs1800471)
török GG 26 83,9% 28 90,3%
GC 5 16,1% 3 9,7%
CC 0 0% 0 0%
spanyol GG 19 100% 17 85%
GC 0 0% 3 15%
CC 0 0% 0 0%
portugál* GG 27 90% - 84,7%
GC 3 10% - 15,3%
CC 0 0% - 0%
Összes 80 51
Portugál kontroll minták nem álltak rendelkezésünkre.
*Portugál kontroll adatok: Alves H, Histocompatibility Center, University of Porto, Portugália
Következtetések
1. Az F508C allél-frekvencia szignifikánsan gyakoribb volt a CBAVD-s betecsoportban mind a CF betegek mind pedig a CF hordozókhoz viszonyítva. Az F508C csere CBAVD-t vagy CF-et is eredményezhet, mivel azonban szignifikánsan gyakoribb volt a CBAVD-s betegekben, ezért az eltérés nagyobb valószínűséggel fog CBAVD-t eredményezni, mint CF-et, amennyiben egy második pathológiás CFTR mutációval társul.
2. A vizsgált TGFB-1 poliformizmusok - rs1800470 és rs1800471 – nem befolyásolták a CBAVD penetranciát. Ezek alapján feltételezhetjük, hogy a TGFB-1 nem CBAVD módosító gén.
3. Az EDNRA exon 8 CC genotípus (rs5335) emelkedett gyakoriságát találtuk mind a török, mind pedig a teljes betegcsoportban. Az EDNRA promoter SNP (rs1801708) nem volt hatással a CBAVD fenotípusra a betegeinkben.
Eredményeink alapján feltételezhetjük, hogy az EDNRA szerepet játszik a CBAVD létrejöttében.
Összefoglalva azt mondhatjuk, hogy a CBAVD egy multifaktoriális betegség, amelynek létrejöttében számos módosító tényező szerepet játszik, amelyek vizsgálatához további nagyobb populációkon végzett kutatások és genom- szintű asszociációs vizsgálatok (GWAS) szükségesek.
Publikációs lista
A disszertáció az alábbi közleményeken alapul
Havasi V, Rowe SM, Kolettis PN, Dayangac D, Ahmet Şahin, Grangeia A, Carvalho F, Barros A, Sousa M, Bassas L, Casals T, Sorscher EJ (2010). Association of cystic fibrosis genetic modifiers with congenital bilateral absence of the vas deferens. Fertil Steril 94(6):2122-7.
Havasi V, Keiles S, Hambuch T, Sorscher EJ, Kammesheidt A (2008). The role of the F508C mutation in congenital bilateral absence of the vas deferens. Genet Med 10(12):910- 4.
További saját közlemények jegyzéke
Bányai K, Gentsch JR, Martella V, Bogdán A, Havasi V, Kisfali P, Szabó A, Mihály I, Molnár P, Melegh B, Szücs G (2009). Trends in epidemiology of human G1[8] rotaviruses:
a Hungarian study. J Infect Dis 200 (Suppl 1):S222-7.
Komlósi K, Talián GC, Faragó B, Magyari L, Cserép V, Kovács B, Bene J, Havasi V, Kiss CG, Czirják L, Melegh B (2008). No influence of SLC22A4 C6607T and RUNX1 G24658C genotypic variants on the circulating carnitine ester profile in patients with rheumatoid arthritis. Clin Exp Rheumatol 26(1):61-6.
Havasi V, Hurst CO, Briles TC, Yang F, Bains DG, Hassett DJ, Sorscher E (2008). Inhibitory effects of hypertonic saline on P. aeruginosa motility. J Cyst Fibros (4):267-9.
Papp E, Havasi V, Bene J, Komlósi K, Talián G, Fehér G, Horváth B, Szapáry L, Tóth K, Melegh B (2007). Does glycoprotein IIIa gene (Pl(A)) polymorphism influence clopidogrel resistance? : A study in older patients. Drugs Aging 24(4):345-50.
Komlósi K, Havasi V, Bene J, Süle N, Pajor L, Nicolai R, Benatti P, Calvani M, Melegh B (2007). Histopathologic abnormalities of the lymphoreticular tissues in organic cation
transporter 2 deficiency: evidence for impaired B cell maturation. J Pediatr 150(1):109-111.e2.
Havasi V, Szolnoki Z, Talián G, Bene J, Komlósi K, Maász A, Somogyvári F, Kondacs A, Szabó M, Fodor L, Bodor A, Melegh B (2006). Apolipoprotein A5 gene promoter region T-1131C polymorphism associates with elevated circulating triglyceride levels and confers susceptibility for development of ischemic stroke. J Mol Neurosci 29(2):177-83.
Havasi V, Komlósi K, Bene J, Melegh B (2006). Increased prevalence of glycoprotein IIb/IIIa Leu33Pro polymorphism in term infants with grade I intracranial haemorrhage.
Neuropediatrics 37(2):67-71.
Szolnoki Z, Havasi V, Talián G, Bene J, Komlósi K, Somogyvári F, Kondacs A, Szabó M, Fodor L, Bodor A, Melegh B (2006). Angiotensin II type-1 receptor A1166C polymorphism is associated with increased risk of ischemic stroke in hypertensive smokers. J Mol Neurosci 28(3):285-90.
Metzger S, Bauer P, Tomiuk J, Laccone F, Didonato S, Gellera C, Mariotti C, Lange HW, Weirich-Schwaiger H, Wenning GK, Seppi K, Melegh B, Havasi V, Balikó L, Wieczorek S, Zaremba J, Hoffman-Zacharska D, Sulek A, Basak AN, Soydan E, Zidovska J, Kebrdlova V, Pandolfo M, Ribai P, Kadasi L, Kvasnicova M, Weber BH, Kreuz F, Dose M, Stuhrmann M, Riess O (2006). Genetic analysis of candidate genes modifying the age-at-onset in Huntington's disease. Hum Genet 120(2):285-92.
Bene J, Komlósi K, Havasi V, Talián G, Gasztonyi B, Horváth K, Mózsik G, Hunyady B, Melegh B, Figler M (2006). Changes of plasma fasting carnitine ester profile in patients with ulcerative colitis. World J Gastroenterol 12(1):110-3.
Metzger S, Bauer P, Tomiuk J, Laccone F, Didonato S, Gellera C, Soliveri P, Lange HW, Weirich-Schwaiger H, Wenning GK, Melegh B, Havasi V, Balikó L, Wieczorek S, Arning L, Zaremba J, Sulek A, Hoffman-Zacharska D, Basak AN, Ersoy N, Zidovska J, Kebrdlova V, Pandolfo M, Ribai P,
Kadasi L, Kvasnicova M, Weber BH, Kreuz F, Dose M, Stuhrmann M, Riess O (2006). The S18Y polymorphism in the UCHL1 gene is a genetic modifier in Huntington's disease. Neurogenetics 7(1):27-30.
Nádasi E, Bene J, Havasi V, Komlósi K, Talián G, Melegh G, Papp E, Gasztonyi B, Szolnoki Z, Sándor J, Mózsik G, Tóth K, Melegh B, Wittmann I (2005). Detection of the Leu40Arg variant of the platelet glycoprotein IIb/IIIa receptor in subjects with thrombotic diseases. Thromb Res 116(6):479- 82.
Szolnoki Z, Havasi V, Talián G, Bene J, Komlósi K, Somogyvári F, Kondacs A, Szabó M, Fodor L, Bodor A, Melegh B (2005). Lymphotoxin-alpha gene 252G allelic variant is a risk factor for large-vessel-associated ischemic stroke. J Mol Neurosci 27(2):205-11.
Bányai K, Pálya V, Benkő M, Bene J, Havasi V, Melegh B, Szücs G (2005). The goose reovirus genome segment encoding the minor outer capsid protein, sigma1/sigmaC, is bicistronic and shares structural similarities with its counterpart in Muscovy duck reovirus. Virus Genes 31(3):285-91.
Komlósi K, Kellermayer R, Maász A, Havasi V, Hollódy K, Vincze O, Merkli H, Pál E, Melegh B (2005). Maternally inherited deafness and unusual phenotypic manifestations associated with A3243G mitochondrial DNA mutation.
Pathol Oncol Res 11(2):82-6.
Bányai K, Forgách P, Erdélyi K, Martella V, Bogdán A, Hocsák E, Havasi V, Melegh B, Szücs G (2005).
Identification of the novel lapine rotavirus genotype P[22]
from an outbreak of enteritis in a Hungarian rabbitry. Virus Res 113(2):73-80.
Papp E, Havasi V, Bene J, Komlósi K, Czopf L, Magyar E, Fehér C, Fehér G, Horváth B, Márton Z, Alexy T, Habon T, Szabó L, Tóth K, Melegh B (2005). Glycoprotein IIIA gene (PLA) polymorphism and aspirin resistance: is there any correlation? Ann Pharmacother 39(6):1013-8.
Szolnoki Z, Havasi V, Bene J, Komlósi K, Szöke D, Somogyvári F, Kondacs A, Szabó M, Fodor L, Bodor A, Gáti I, Wittman I, Melegh B (2005). Endothelial nitric oxide synthase gene interactions and the risk of ischaemic stroke.
Acta Neurol Scand 111(1):29-33.
Komlósi K, Bene J, Havasi V, Tihanyi M, Herczegfalvi A, Móser J, Melegh B (2004). Phenotypic variants of A3243G mitochondrial DNA mutation in a Hungarian family. Orv Hetil 145(35):1805-9.
Melegh B, Bene J, Mogyorósy G, Havasi V, Komlósi K, Pajor L, Oláh E, Kispál G, Sümegi B, Méhes K (2004).
Phenotypic manifestations of the OCTN2 V295X mutation:
sudden infant death and carnitine-responsive cardiomyopathy in Roma families. Am J Med Genet A 131(2):121-6.
Komlósi K, Havasi V, Bene J, Storcz J, Stankovics J, Mohay G, Weisenbach J, Kosztolányi G, Melegh B (2005). Increased prevalence of factor V Leiden mutation in premature but not in full-term infants with grade I intracranial haemorrhage.
Biol Neonate 87(1):56-9.
Komlósi K, Havasi V, Bene J, Ghosh M, Szolnoki Z, Melegh G, Nagy A, Stankovics J, Császár A, Papp E, Gasztonyi B, Tóth K, Mózsik G, Romics L, ten Cate H, Smits P, Méhes K, Kosztolányi G, Melegh B (2003). Search for factor V Arg306 Cambridge and Hong Kong mutations in mixed Hungarian population samples. Acta Haematol 110(4):220-2.
Szolnoki Z, Somogyvári F, Kondacs A, Szabó M, Bene J, Havasi V, Komlósi K, Melegh B (2003). Increased prevalence of platelet glycoprotein IIb/IIIa PLA2 allele in ischaemic stroke associated with large vessel pathology.
Thromb Res 109(5-6):265-9.
Sándor J, Havasi V, Kiss I, Szücs M, Brázay L, Sebestyén A, Ember I (2002). Small area inequalities in breast cancer mortality and screening. Magy Onkol 46(2):139-45.
Havasi V, Sándor J, Kiss I, Szűcs M, Brázay L, Ember I (2001). Mortality of breast cancer and frequency of mammography in Hungary. Orv Hetil 142(50):2773-8.
Könyvfejezetek
dr. Havasi Viktória, Dr. Melegh Béla: Familiáris hypercholesterinaemia. In: Kornya László (szerk.), Betegségenciklopédia I-II. Springer Kiadó, Budapest, 2002:
270-272.
dr. Havasi Viktória, Dr. Melegh Béla: “Fish-eye” betegség.
In: Kornya László (szerk.), Betegségenciklopédia I-II.
Springer Kiadó, Budapest, 2002: 272-273.
dr. Havasi Viktória, Dr. Melegh Béla: Fucosidosis. In:
Kornya László (szerk.), Betegségenciklopédia I-II. Springer Kiadó, Budapest, 2002: 276-277.
dr. Havasi Viktória, Dr. Melegh Béla: Haemochromatosis.
In: Kornya László (szerk.), Betegségenciklopédia I-II.
Springer Kiadó, Budapest, 2002: 297-299.
dr. Havasi Viktória, Dr. Melegh Béla: Histidinaemia. In:
Kornya László (szerk.), Betegségenciklopédia I-II. Springer Kiadó, Budapest 2002: 312-313.
dr. Havasi Viktória, Dr. Melegh Béla: I. A típusú hyperlipoproteinaemia. In: Kornya László (szerk.), Betegségenciklopédia I-II. Springer Kiadó, 2002., 326-327.
dr. Havasi Viktória, Dr. Melegh Béla: I. B típusú hyperlipoproteinaemia. in: Betegségenciklopédia I-II. Szerk.:
Kornya László, Springer Kiadó, Budapest, 2002: 328-329.
dr. Havasi Viktória, Dr. Melegh Béla: Lecitin-koleszterin- aciltranszferáz hiány. In: Kornya László (szerk.), Betegségenciklopédia I-II. Springer Kiadó, Budapest, 2002:
351-352.
dr. Havasi Viktória, Dr. Melegh Béla: Mannosidosis. In:
Kornya László (szerk.), Betegségenciklopédia I-II. Springer Kiadó, Budapest, 2002: 352-353.
dr. Havasi Viktória, Dr. Melegh Béla: Örökletes methaemoglobinaemia. In: Kornya László (szerk.), Betegségenciklopédia I-II. Springer Kiadó, Budapest, 2002:
392-393.
Köszönetnyilvánítás
Köszönettel tartozom a következő embereknek:
Konzulensemnek, Dr. Zsembery Ákosnak, aki lehetővé tette számomra, hogy az irányítása alatt egyéni felkészülő PhD hallgatóként dolgozhassak; amerikai mentoromnak, prof. Dr.
Eric Sorschernek; korábbi témavezetőimnek: prof. Dr. Miseta Attilának és id. prof. Dr Kellermayer Miklósnak; a PTE ÁOK Genetikai Intézet összes munkatársának, kiváltképpen Berenténé Bene Juditnak, aki megismertette velem a molekuláris biológia alapjait, és akitől számtalan esetben kaptam segítséget; a University of Alabama at Birmingham, Genomics Core Facility of the Howell and Elizabeth Heflin Center for Human Genetics és az Ambry Genetics összes dolgozójának a minták analízisében nyújtott színvonalas technikai segítségért; szüleimnek, testvéremnek és barátaimnak akiknek a támogatása nagyban hozzájárult munkám sikeréhez; végül de nem utolsósorban az összes vizsgálati személynek, akik hozzájárultak ahhoz, hogy mintáikat a kutatásainkhoz felhasználhassuk.