MTA Doktori Értekezés Tézisei
Háziállatok szöveti lipidjeinek
zsírsavösszetételi elemzése eltér® élettani
állapotokban
Dr. Szabó András (Ph.D.)
Kaposvári Egyetem
Állattudományi Kar
Kaposvár, 2013
1. Bevezetés
Azállattenyésztésitudományokterületénegyrehangsúlyosabbszerepet
kapnakbiokémiai,illetveélettaniszempontok,kétf®területenis. Atáplál-
kozástudományésazállatitermékel®állításkapsolatamáraigenszorossá
vált,hiszenajómin®ség¶termékalapjatagadhatatlanulazegészséges és
hatástanilagisismertkörülményekközötttermel®állat. Amodernállat-
tartásaszelekiómellettszámosolyankörnyezeti(pl.tehnológiai)faktort
alkalmaz,melyekgyakarankevésbéismertmódonhatnakazállatokjólé-
tére(welfare)vagyegyszer¶enazéletmin®ségére. Atáplálkozástudomány
rohamosfejl®désévelösszhangban azállati eredet¶termékekösszetétele,
különösenafunkionáliskomponensekfokozottjelenléte prioritástnyert,
melynekmár a termékel®állítás során is egyre nagyobb jelent®sége van.
Miutánahumánzsírsavellátásegyharmadát-felétazállatitermékekbiz-
tosítják,fokozottgyelemilletimegazállatitermékekzsírsavösszetételét,
különösenpedig azesszeniáliszsírsavakat. A dolgozatbafoglalt, f®képp
azállatiszövetekzsírsavösszetételére vonatkozóközelítésekközelsemte-
kinthet®kátfogónakvagyteljesnek,mégisélzottanarrairányultak,hogy
megmutassákazsírsavösszetételbizonyoskörnyezetiésgenetikaitényez®k
hatásárabekövetkez®meglep®ennagyváltozatosságát. A lehet®ségekhez
mérten nem kizárólag a zsírsavakra, hanem a zsírsav-, és esetenként a
szervezetszint¶ lipidmetabolizmus vizsgálatára is vállalkoztam,illetve a
legtöbbesetbenlipidfrakióhozismegpróbáltamkötniatapasztaltválto-
zásokat.
AKaposváriEgyetemÁllattudományiKaránakÁllatitermékMin®sít®
Laboratóriumában az állati termékekmin®ségét a lehet® legszélesebben
értelmezi a munkasoportunk. Kapsolódva a Karon folyó, els®sorban
állattartásiéstakarmányozásikezeléseketalkalmazókísérletesmunkához
a dolgozatban bemutatandó kísérletes aktivitás arra irányult, hogy f®-
képpháziállatok(eml®sök,baromfajokéshalak)szöveteibenelemezzeaz
anyagsereintenzitás,a rendkívül intenzívnövekedés, valamint a takar-
mányeredet¶zsírsavakszövetilipidekre,lipidfrakiókzsírsavairaésennek
folyományaként azoxidatív stabilitásra gyakorolt hatásait. A dolgozat-
banennekmegfelel®ena 2004-2012 id®szaktámogatottkutatásaitfogla-
lom össze, melyekgazdasági és vadon él® állatokon, kontrollált kísérle-
tek,illetveszisztematikusmintagy¶jtésekkeretébenkészültekaKEÁTK
ÁllatitermékMin®sít®Laboratóriumában.
1.1. El®zmények
Állatieredet¶lipidekreirányulókísérletesmunkámataPh.D. ösztön-
díjas id®szakban kezdtem el (kizárólag nyúlon), majd ezt követ®en in-
tézményi szint¶ támogatás mellett aKaposváriEgyetemÁllatudományi
Karán2004-benlétrejöttazakkoriÁllatiTermékfeldolgozásésMin®sítés
Tanszék. ATanszékenlehet®ségetkaptamegylipidekkelfoglalkozólabo-
ratóriumirészkialakítására,RomváriRóbertProfesszorvezetésemellett.
Ezzelegyid®benháromévesOTKAPosztdoktoripályázatitámogatásban
részesültünk háromévre (2004-2007). A laboratóriumban annak f®, el-
s®sorbanazélelmiszer alapanyagokmin®sítésére irányulóprolja mellett
azótahazai -éskisebbrészben nemzetközi-támogatással folynakrend-
szeres, az állati (eml®s, madár éshal) lipidekre irányuló kutatási prog-
ramok. Szerensésmódon,alapozvaaKEÁTKspeiális állattenyésztési
adottságaira,sikerültezenvizsgálatoklegtöbbjébeazállatkísérleteslehe-
t®ségekkelrendelkez®társ-tanszékeketéskollégákatisbevonni.
1.2. A kutatások élja
Azértekezéstárgyáhozkapsolódóvizsgálatoknégyf®soportbasorol-
hatók,ésnégyalapvet®kérdésreirányultakaélkit¶zésekis.
1. Ajelent®seneltér®energetikaiállapotok(intenzívnövekedés,ener-
giamegvonás)metabolikushatásaihogyanírhatókleszelektáltbaromfa-
jok(pulyka,brojlersirke,tojótyúk)klinikai-kémiaivizsgálatával?
2. Az energetikai status milyen hatást gyakorol a szöveti lipidfrak-
iókzsírsavösszetételére madárfajokban? (Pulykamellizomzat ontogene-
zisalattimembránlipidösszetételi elemzése,ésbaromfajokkomparatív,
allometrikusvizsgálata (szív,máj,vese,tüd®(parenhymaéssurfatant)
ésagy).
3. Anatívszövetitriglieridekösszetételénekhelyzetspeikuszsírsav-
összetételielemzésealkalmas-eafaj-ésfajtaazonosításrahételtér®fajés
kétsertésfajta esetében?
4. Milyen kinetikávaljellemezhet®a takarmányeredet¶zsírsavak szö-
vetilipidekbeéslipidfrakiókbatörtén®inkorporáiójaédesvizihalakban,
afrikaiharsaéstilápiaesetében?
2. Anyagok és módszerek
2.1. Vizsgálatba vont állatok, tartás, takarmányo-
zás
Azalábbbemutatottvizsgálatoksoránmindenesetbenmeghatároztuk
atakarmányoktáplálóanyag-,ésszükségeseténzsírsavösszetételét is,az
állatvizsgálatokpedig aMunkahelyiÁllatkísérleti Bizottságengedélyezé-
sétkövet®entörténtek.
2.1.1. Eltér® energetikai állapotok metabolikus hatásainak jel-
lemzésebaromfajok klinikai-kémiai vizsgálatával
2.1.1.1. Tojótyúk mesterséges vedletése,a koplalás vérparamé-
terekkeltörtén® nyomonkövetése
Tojótyúkokon12napos,teljestakarmánymegvonásosmesterségesved-
letéstalkalmaztunk,melyperiódusbanháromnapontavérvételeketvégez-
tünk,klinikai-kémiaiszérumanalíziséljából. Ugyanezenmadarakszöve-
teitelemeztüka2.1.2.1. fejezetben.
2.1.1.2. Hústermelésre szelektált pulykavérbiokémiai paramé-
tereineknyomonkövetése,akelést®la vágásig
BUT Big 8 hímivarú, nagy növekedési erély¶pulykákat neveltünk az
intenzívnövekedés nyomon követése éljából, majd 3naposan, 4,8, 12,
16,illetve20heteskorbanvégeztünkvérvételeket,klinikai-kémiaiszérum
analízis éljából. Ugyanezen állomány izommintáit elemeztük a 2.1.2.2.
(illetve3.1.2.2.) fejezetben.
2.1.1.3. Abrojlersirkekliniko-kémiaivizsgálata(follow-up)
ateljesfelnevelés alatt
Avizsgálatba Ross 308hímivarúbrojlereketvontunk be, avérvétele-
ket0,1, 2, 3,4 és5hetesen végeztük,az intenzívnövekedés jellemzése
érdekében.
2.1.2. Az energetikai status hatása a szöveti lipidfrakiók zsír-
savösszetételére madárfajokban
2.1.2.1. Tojótyúkokmesterségesvedletése,adrasztikusenergia-
megvonáshatásaamájésszíveltér®lipidfrakióinak(triglierid
ésfoszfolipid)összetételére
Avizsgálatbana2.1.1.1. fejezetbenismertetettmadarakvettekrészt.
Akoplaltatásid®szakátmegel®z®en,ésannakvégén(0. és12. napok)10-
10madárbólaszívetésamájatzsírsavösszetételi(foszfolipidéstriglierid
frakiók)analízisreeltettük.
2.1.2.2. Pulykamellizommembránlipidekontogenezisalattivál-
tozásainakleírása
A2.1.1.2. fejezetbenismertetettállománybólpróbavágástkövet®enfel-
színesésmélymellizommintákatelemeztünkmindenmintavételiid®pont-
ban,a membránlipidekzsírsavproljára vonatkozóan, allometrikusadat-
értékeléssel.
2.1.2.3. Eltér® kifejlett kori testsúlyú madárfajok szívizmának
membránlipidjei,allometrikusanalízis
Azelemzésbe pulyka,lúd,tyúk,fáán, húsgalamb,balkánigerleésja-
pánfürjfajok 5-5 kifejlett egyedét vontukbe,ésaszívizom teljesfoszfo-
lipidjeinekzsírsavproljátelemeztük,allometrikusmódszertannal,atest-
súlyfügg®szabályosságokfeltárásaérdekében.
2.1.2.4. Eltér® kifejlett kori testsúlyú madárfajok tüd®, vese,
májésagyfoszfolipid zsírsavproljának allometrikus elemzése
A 2.1.2.3. fejezetben megadott fajokon túl foglyot is vizsgáltunk. A
tüd®, vese, máj ésagy mintákteljes foszfolipid zsírsavösszetételét hatá-
roztukmeg,azadatértékelés szinténallometrikusvolt.
2.1.2.5. A tüd® parenhymalis és surfatant foszfolipidjeinek
allometrikus elemzésebaromfajokban
A2.1.2.4. fejezetbenemlítettfajokbantüd®b®lmosottsurfatant(vala-
mintateljestüd®ésamosottszövet,kváziparenhyma)foszfolipidjeinek
zsírsavprolját határoztuk meg, allometrikus adatelemzésre alapozva, a
strukturális és a szekretált foszfolipid zsírsavprol testsúlyfügg® szabá-
lyosságainakvizsgálataéljából.
2.1.3. A szöveti triglieridek (TG) összetételének natív alaku-
lása;afaj-ésfajtaazonosításlehet®ségei aTG molekulaszerke-
zetrészletesanalízisére alapozva (regiospeikus analízis)
2.1.3.1. Afajspeikustulajdonságokelemzésetriglieridekmo-
lekulaszerkezetivizsgálatára alapozva
Gímszarvas,jávorszarvas,vaddisznó,extenzívennevelthússertés,borz,
húsnyúléslúdsubutanzsírmintáikerültekelemzésre. Atriglieridekb®l
hasnyállipázos hidrolízistkövet®enaz1,3 ésa2helyzetizsírsavproltis
meghatároztuk,azalábbiakszerint: 1,3helyzetzsírsavprolja,zsírsavan-
ként: 1,3%=(3xTG%-2MG%)/2,majddeniáltukazún. helyzetimeg-
oszlásifaktort: Helyzetimegoszlásifaktor(HMF):HMF=log(1,3%/2%).
2.1.3.2. Afajtaszint¶eltérésekvizsgálatasertésgenotípusoktri-
glieridjeinekmolekulaszerkezetivizsgálatával
A 2.1.3.1. fejezet módszertanára építve a fajtaszint¶ eltérések felde-
rítése volt a él, magyar nagyfehér és sz®ke mangalia egyedek szöveti
triglieridjeiben(szív-ésvázizom,hátszalonna,tüd®,vese,májéslép).
2.1.4. A takarmányeredet¶ zsírsavak szövetilipidekbe és lipid-
frakiókba történ® beépülésének (inkorporáió) vizsgálata, ha-
lakban
2.1.4.1. Afrikaiharsa(Clariasgariepinus)lézsírsavösszetétele
éshúsmin®sége,növényiolajok etetésétkövet®en
Avizsgálatéljaazvolt,hogymegállapítsuk,ahalolajrészlegeshelyette-
sítése(növényiolajokkal)aharsatakarmánybanmilyenváltozásokatidéz
el®ahúsmin®ségben,ahúskémiaiösszetételében,illetvehogyjellemezzük
azsírsavakizomszövetbe (annakteljeslipidtartalmába) történ®inkorpo-
ráióskinetikáját, azún. zsírsavkihígulási modellsegítségével, hathetes
kezelésiintervallumban,hárommintavételrealapozva.
2.1.4.2. Növényiolajokszövetilipidfrakiókbavalóbeépülésének
elemzéseNílusi tilápia (Oreohromis nilotius)szerveiben
A vizsgálat élja az volt, hogy jellemezzük eltér® növényi olajok és
halolaj(mintkontroll)zsírsavainakinkorporáiósüteméttilápiaszövetek
(lé,has¶rizsír,májésgonádok)lipidfrakióiba(triglieridekésfoszfolipi-
dek),hatheteskezelési periódusban. Azadatelemzéstazegyszer¶zsírsav
kihígulásimodellsegítségévelvégeztük.
2.2. Mintael®készítés és laboratóriumi vizsgálatok
A vérmintákból a klinikai-kémiai paramétereket automata készüléke-
kenhatározták meg. A szöveti lipidtartalom kivonása (tejesszövetb®l,
tüd®mosófolyadékból) mindenesetben Folh ésmtsai (J. Biol. Chem.,
1957, 226:497-509) módszeréveltörtént, antioxidánsjelenlétében,míg a
komplexlipidek frakionálása szilikagéloszlopon(emelked®polaritású e-
luenssorozatokkal),illetve szilikagélG60 lemezekentörtént. Azsírsavak
gázkromatográás elválasztását és elemzését megel®z® származékképzés
minden pontban NaOCH
3
alapú, báziskatalizált módszer volt. A lipi-
dekkelkapsolatospreparatívlépések aKE ÁTK ÁllatitermékMin®sít®
Laboratóriumában történtek,míg gázkromatográás analízis azÁllatte-
nyésztésiésTakarmányozásiKutatóintézetbenkészült(egytanulmány,a
harsalé(2.1.4.1.) kivételével),Shimadzu2010 készüléken. A malondi-
aldehid(MDA)konentráiótnatív, -70 o
C-ontároltszövetekb®laSzIE
MKK Takarmányozástani Tanszékén határoztákmeg, fotometriás mód-
szerrel, míg a hagyományos húsmin®ségre irányuló vizsgálatokat a KE
ÁllatitermékMin®sít® Laboratóriumában végeztük. A Weendeianalízis
soránszabvány módszereketkövettünkafelsoroltakszerint: szárazanyag
(MSZ-08-1783-1: 1983); nyersfehérje, Nalapon, Kjeldahl módszer (MSZ
6367-11:1984);nyerszsír(6830-6:1984);nyershamu(MSZISO5984).
2.3. Statisztikai értékelés
Azsírsavprolalapadatok értékelésesoránsoportonként,illetvekeze-
lésenként a kétszeres szórástávolságon kívül es® adatokat (els®dleges és
származtatott esetekben is) kihagytam az elemzésb®l. A vérparaméte-
rekid®pontonkéntiösszehasonlítása sorántöbbtényez®svarianiaanalízist
(kezelésid®tartama,testsúly)alkalmaztam,Tukey-féleposthoteszttel.
Azenergiamegvonáshatásánakelemzésekorakezd®észáróid®pontokban
felvettadatok(zsírsav,malondialdehid)összevetésérekétmintásfüggetlen
t-próbátalkalmaztam. Azallometrikuselemzéseksorán(3.1.2.2.-3.1.2.5.)
azsírsavprolprimerésszármaztatottadataitmintatestsúlyx-edikhat-
ványaithatároztammeg(zsírsavmol%=konstans+testsúly x
).
A2.1.4. fejezetbenalkalmazottpredikiós modellsorán Jobling (Aqu-
ault. Res., 2004, 35:706-709) ún. zsírsav kihígulási modelljét vettem
alapul. Amodell szerintazsírsavak inkorporáiója legtöbbszöraszerve-
zetbenfellelhet®készletkihígulásávaljellemezhet®folyamat,ennélfogva
azviszonylagegyszer¶modellelleírható,azalábbiakszerint:
Pt= Pr+ (Pi Pr) /(Qt -Qi),ahol
Pt: tid®pontbanazadottzsírsav%-a
Pi: akiindulásiid®pontbanmértzsírsav arány
Pr: arefereniasoportzsírsav-részaránya
Qi: kvantitatívmutató(pl. szervtömeg)akiinduláskor
Qt: éstid®pontban
AstatisztikaielemzésekbenmindenesetbenSPSS10forWindows(1999)
programothasználtam,azábrázolást pedigazallometrikusmodellekese-
tébenQtiPlot0.9.7.6. (2009)szoftverrelkészítettem,egyLinuxmunkaál-
lomáson.
3. Eredmények és értékelésük
3.1.1. Eltér® energetikai állapotok metabolikus hatásainak jel-
lemzésebaromfajok klinikai-kémiai vizsgálatával
3.1.1.1. Tojótyúk mesterséges vedletése,a koplalás vérparamé-
terekkeltörtén® nyomonkövetése
Teljestakarmánymegvonásosmesterségesvedletésnekkitetttojótyúkok
klinikai-kémiai analízisére alapozva megállapítottuk, hogy az általános
energiahiányeseténatojástermelésiiklusgyorsanmegszakad,amitavér-
benlegérzékenyebben a szérum metabolitok (sökken® triglierid, emel-
ked®HDL ésösszkoleszterin) jeleznek. A máj és szívizom esetében ta-
pasztalt, kés®bbiekben(3.1.2.1.) bemutatásrakerül®jelent®skárosodás,
illetvekatabolizmusazintraellulárisenzimekszérumbanmértaktivitásá-
naknövekedésébensaka
γ
-GTésaLDHesetébent¶ntreálisnak. Fontosmegjegyezni,hogyafentiekbenbemutatottvizsgálatindirektéljaazvolt,
hogyateljestakarmánymegvonásosvedletésimódszerekkelkapsolatosan
olyan, refereniaként is értelmezhet®eredményekettegyenközzé, melyek
alapjánannakháttérbeszorulása voltvárható. Id®közbenezenvedletési
módszermárbetiltásrakerültHazánkbanis.
3.1.1.2. Hústermelésre szelektált pulykavérbiokémiai paramé-
tereineknyomonkövetése,akelést®la vágásig
Azegyirányúan hústermelésre szelektált madaraknál nagyon gyakran
fordulnak el® miopátiás esetek; elemzésünk kimondottan ezen esetleges
kórkép,vagya nagyonintenzívizom hipertróaokozta jelenségekre irá-
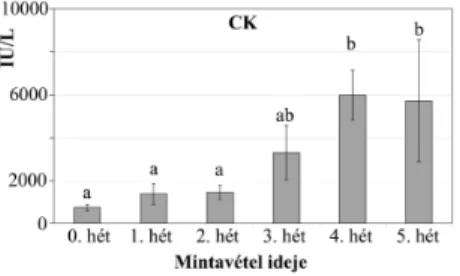
nyult. Anagynövekedésierély¶hústípusúpulykákszérumelemzéseolyan,
koránér® madársajátosságaira utalt, melynéljellemz®a korai, szikfel-
használásból fakadó triglierid sús,majd azigen intenzívizomfejl®dés
(magas laktát dehidrogenáz; extrém magas és tovább emelked®szérum
kreatinkinázaktivitás,1. ábra),ésvégüligen nagykifejlettkoritestmé-
retetérel. Érdekestapasztalat,hogyapetoralis izomzatlineárisnöveke-
dését(mellizomsúly=0.2787testsúly123.67;R 2
=0.9935,P<0.001)
er®sALP aktivitás-sökkenés kísérte(2. ábra),mely asontrendszer és
azizomzataszinkronfejl®déséreutaltebbenaszéls®ségesenszelektáltál-
lományban.
1. és2. ábraAkreatinkináz(CK)ésazalkalikusfoszfatáz(ALP)
aktivitásváltozásaahústípusúpulykafelnevelésesorán
Bárazalkalmazottmetodikaésközelítésrutinanalízisenalapul,asze-
lekióel®rehaladtávalahústermel®állományok folyamatosantávolodnak
akiindulásiponttól(bronzpulyka),ígyeredményeink,különösenaszelek-
ióhátrányoskövetkezményeikapsánújszer¶nektekinthet®k.
3.1.1.3. Abrojlersirkekliniko-kémiaivizsgálata(follow-up)
ateljesfelnevelés alatt
Amindösszesenöthétigtartónövekedésifolyamatsorán(2800gél®tömeg
eléréséig)egyjellegzetesenkoraiéréstmutató(koraitriglieridésösszko-
leszterinkonentráiósúsokésgyorsanemelked®konentráiójúplazma
fehérjék)madáradatait rögzítettük. A hústermelésreirányulószelekió
olyan mérték¶ izom hipertróát eredményezett a vizsgált Ross 308 ge-
notípusban, mely enyhe hiperkalémiához, magas AST és hasonlóan a
hústermel®pulykához-extrémmagasCKaktivitáshozvezetett(3. ábra).
3. ábraAkreatinkinázaktivitás-emelkedésea
brojlersirkékfelnevelésesorán
Érdekesmódon a nagyonrövid nevelési id®szak nemtette lehet®vé a
korhatásánakhagyományosértékelését(szervetlenfoszfátésNaminimális
uktuáiója),ámmindajellegzeteskoraérés,mindpedigazextrémmagas
hústermelésikapaitásjólkimutathatóvoltezenszelektáltállománybanis.
3.1.2. Az energetikai status hatása a szöveti lipidfrakiók zsír-
savösszetételére madárfajokban
3.1.2.1. Tojótyúkokmesterségesvedletése,adrasztikusenergia-
megvonáshatásaamájésszíveltér®lipidfrakióinak(triglierid
ésfoszfolipid)összetételére
Atojótyúkok ipariszint¶ tojástermeléserendszerint nemsakegy, ha-
nemtöbb iklusban valósul meg, melyekethagyományosan mesterséges
vedletésiprogramokszakítanakmeg. Tekintettelarra, hogyavedlésifo-
lyamat megindítása priméren az energiaraktárak kimerülésének köszön-
het®,lipidanyagsereszempontjábólkiemelked®en érdekesezen élettani
helyzet,melyetjelenfejezetbenamájésaszívtriglierid(TG)ésfoszfo-
lipid(PL)zsírsavanyagseréjénekelemzésénkeresztüljellemeztünk.
Amájtriglieridekbenakoplaláshatásáraapentadekánsav(C15:0),
agondoénsav(C20:1n9)ésanervonsav(C24:1n9)részarányaemelkedett,
míg a palmitinsav (C16:0) aránya és az n6/n3 hányados er®sen lesök-
kent. Fontos megjegyezni,hogy aC20< PUFA el®állításának legjellem-
z®bbhelyeamáj,melyakoplalásidejénazextrahepatikusszervekPUFA
ellátásábanszintekizárólagosszerep¶,miutánaraktározott,adiposaere-
det¶triglieridezenzsírsavakbanmeglehet®senszegény.Érdekes,bizonyos
mértékigellentmondvaafentieknekmindazarahidonsav,mindpedig a
dokozahexaénsav határozott retenióját (paradoxial onservation) ta-
pasztaltukamájtriglieridekben.
Amáj foszfolipidjeinekenergiamegvonásraadott reakióiközül ki-
emelend® apalmitinsav, a palmitoleinsav (C16:1n7), az olajsav (C18:1
n9),a dokozahexaénsav (C22:6 n3)ésaz összes n3 zsírsav aránysökke-
nése,azt tükrözve, hogyahosszan tartóenergiahiány meghatározómér-
tékbenbefolyásolja amáj membránlipidjeinekösszetételét. Afoszfolipid
zsírsavprolt tekintveúgyt¶nt, hogyamája normál, élettani memb-
ránlipidintegritástnem képes12 napos koplalás alatt fenntartani, és a
jelent®sszervsúlyveszteség(24%)mellettkomolyabbmembránkárosodás-
salisszámolnikell(melyavérparaméterekbenisigazolhatóvolt,3.1.1.1.
fejezet).
A miokardiális triglieridekben, melyeksak igen sekély válasz-
reakiót mutattak, az
α
-linolénsav aránya sökkent, míg aγ
-linolénsav részaránya szignikánsan emelkedett a 12 napos kezelés következtében,azt jelezve, hogy az éhezési állapot bár megemeli a szívizom triglierid
konentráióját(+12%),annakmin®ségitulajdonságaitsakminimálisan
befolyásolja.
Aszívmembránlipidjeibenalegfontosabbváltozáskéntértékelhet®,
hogyazeikozadiénsav,azeikozatriénsav(C20:3n6)ésadokozahexaénsav
részarányokszignikánssökkenéstmutattak,mígaC20:4n6/C20:3n6
hányadoskéntmegadott
Δ
5deszaturázindexértékejelent®senemelkedett, arrautalva,hogyadrasztikusenergiamegvonástojótyúkbanamiokardi-álismembránlipidekdegradáiójávaljár,melyafoszfolipidekzsírsavainak
homeosztázisát negatívanbefolyásolja.
A lipidperoxidáió mértékének beslésére alkalmazott malondialde-
hidkonentráiója amájban igener®senmegemelkedetta12. napra(4.
ábra),melynekháttereamagasPUFArészaránnyalrendelkez®strukturá-
lislipidekdegradáiójalehetett,miutánaMDAf®legaháromvagytöbb
telítetlenkötésttartalmazózsírsavakbólkeletkezik.
4. ábraAmájésaszívizomlipideperoxidáiójánakváltozása
mesterségesvedletéssorán
Azeredményeketösszefoglalvaazttapasztaltuk,hogyavedletésiprog-
ramamájban mindatároló, mindastrukturálislipidekbenmennyiségi
ésmin®ségiváltozásokatisel®idéz. Úgyt¶nik,12naposteljestakarmány-
megvonáskomolyabbmájsejtmembránkárosodástokozott,melyetafoko-
zottlipidperoxidáióis jelzett. Mígamáj triglieridekkiemeltszerepet
játszottak a polién zsírsavak homeosztázisának fenntartásában, addig a
szívtriglieridekszintesemmilyenmin®ségi változástnemmutattak. A
szívtriglieridekmennyiségi növekedése, ésváltozatlanzsírsavösszetétele
kapsán amáj speiális kardioprotektívszerepe merültfel a koplaltatás
idején.
3.1.2.2. Pulykamellizommembránlipidekontogenezisalattivál-
tozásainakleírása
A hústermelésre szelektált madarakközül napjainkra talána pulyká-
ban történt a legnagyobb változás atestösszetétel ésa növekedési ütem
vonatkozásában. A pulykamellizommára korai,kelést követ® id®szak-
ban igen fontos szerepet kap, különösen a termogenezisben, míg annak
ontogenikus fejl®désével kapsolatosan meghatározó informáió, hogy a
tyúkalakúak(Galliformes) jellemz®enkoraérés¶,fészekhagyó fejl®désme-
netetmutatnak,melysorán azembrionális ésakifejlettkori energetikai
tulajdonságok jelent®sen eltérnek. A mellizom membránlipidek ontoge-
nezis alatti összetételi változásait allometrikus módszertannal elemezve
számosesetben szignikánsallometrikus felj®désmenetetírtunk le,külö-
nösena prekurzor-termékzsírsavak (C18:3n3 -C22:6 n3) reláiójában.
Azontogenezis alatt rögzített eredményekeltértek akora- éskés®nér®,
kis- és nagy test¶ madarak összevetéséb®l nyert logenetikus összefüg-
gésekt®l,amennyibenemelked®testsúlymellettallometrikusanemelked®
telítetlenségi indexet (UI) éssökken® telített zsírsav aránytigazoltunk
(5. ábra).
5. ábraAtelítettzsírsavakésatelítetlenségiindex(UI)változásaia
pulykamellizomfoszfolipidjeiben,atestsúlytólfügg®en
Afentieredményekalapjánindokoltnakt¶nikakérdés,hogyvajonsak
ahústípusúpulykamembránlipidösszetételénekalakulásailyenspeiális,
vagymaga azontogenezis alattifejl®dési menettérelnagymértékbena
logenetikusösszefüggésekt®l.
3.1.2.3. Eltér® kifejlett kori testsúlyú madárfajok szívizmának
membránlipidjei,allometrikus analízis
Azalapvet®enkomparatívvizsgálatkeretébeneltér®kifejlettkoritest-
súlyúmadarakszívizmánakmembránlipidjeitelemeztük,mígamunkahi-
potézisalapja az volt, hogy amembránok, mintmetabolikusütemadók
elméletetmadarak esetében sak mellizomban és szub-elluláris szinten
(máj mitokondriális membránokban és vese mikroszómában) igazolták.
A madárszívizom membránlipidzsírsavösszetételi eredmények szerint a
vizsgálatbavont fajok közöttjelent®sebb interspeikus eltérésekállnak
fenn. A származtatott zsírsav adatok közül a telítetlenségi index (UI)
ésaz összesPUFAarányaallometrikussökkenéstmutatott,míg utóbbi
zsírsav soportban anagyobb test¶madarak alasonyabb n3részarányt
mutattak,az összes monoén ésösszes n9monoénzsírsav pedig allomet-
rikusemelkedést. Azegyedizsírsavak közülapalmitoleinsav, azolajsav,
avakénsav, agondoénsav ésaz eikozatriénsav (C20:3 n3) allometrikus
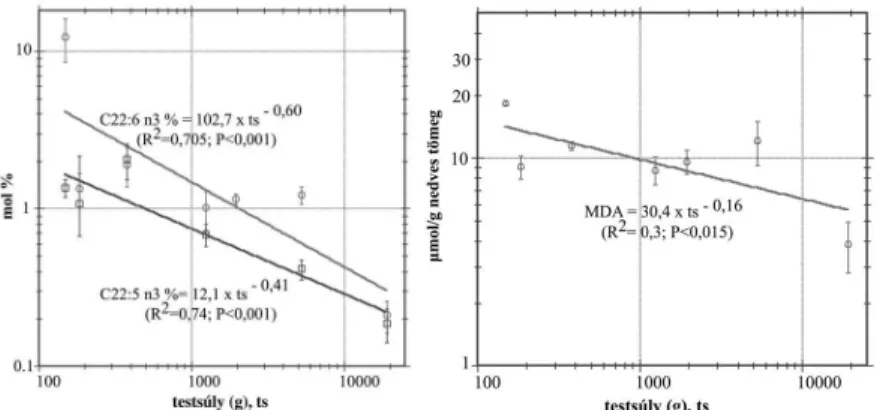
arányemelkedéstmutatott,mígapalmitinsav,asztearinsav,adokozapen-
taénsav(C22:5 n3)ésa dokozahexaénsav aránya (6. ábra) allometrikus
aránysökkenésselvoltjellemezhet®. Aszívizommalondialdehidkonent-
ráiójanegatívallometrikusösszefüggéstmutatott(7. ábra).
6. és7. ábraAdokozapentaén-ésdokozahexaénsavrészarányának,
valamintamalondialdehidkonentráiójánakallometrikusváltozásai
madárszívizomban
Amadárszívmembránlipidekzsírsavproljábantapasztalttestsúlyfügg®
szabályosságok alapvet® egyezést mutattak a madármellizom és eml®s
szívizomesetében eddig korábban publikált adatokkal. Ismert, hogy az
alapanyagsere intenzitása allometrikus változást mutat, melyetf®leg a
sejtmembránok,azokonbelül pedig leginkább alipidek szabályoznak. A
korrelatív,testsúlyfügg®membránlipidösszetételiszabályosságok,különö-
senatöbbszöröstelítetlenségmértékeésDHArészaránya,azazamemb-
ránlipid mili® er®sen befolyásolja a membránhoz kötött enzimek, f®leg
a Na +
/K +
ATPáz molekuláris aktivitását. Bemutatott eredményeink
arrautalnak,hogyaszívizombanisigazoltzsírsavösszetételi szabályossá-
gokegyszélesebbösszefüggésrészeikéntértelmezhet®k,melyekmadárban
nemsak avázizomzatban, de a szívben, valamint a vese mikroszómális
membránesetébenisfennállnak. Aszívizombantapasztalteredményekre
alapozvaatovábbiakbanamadaraktöbb,metabolikusanaktívszervéreis
kiterjesztettükavizsgálatot, arrakeresveaválaszt,mennyireáltalános a
madárszervekbenamembránlipidekzsírsavainakallometrikuselrendez®-
dése.
3.1.2.4. Eltér® kifejlett kori testsúlyú madárfajok tüd®, vese,
májésagyfoszfolipid zsírsavproljának allometrikus elemzése
Hasonlóan a 3.1.2.3. fejezetben bemutatott, fajok összehasonlítására
alapozottvizsgálathoz,jelenfejezetbennyolmadárfajeddignemelemzett
szerveinekmembránlipidzsírsavproljátelemeztük. Amunkaújszer¶sége
abbanállt,hogya mellizomésszívizommellett többtovábbi, metaboli-
kusanaktívszerv (teljestüd®, máj, veseésagy)teljes szövetifoszfolipid
tartalmánakanalízisérekerültsor. Eredményeinkszerintavese,amájésa
tüd®foszfolipidzsírsavproljánakallometrikusösszefüggéseinagybanha-
sonlítottakakorábbanváz-ésszívizomban,valamintszub-ellulárisfrak-
iókbanisigazoltszabályosságokhoz (testsúllyalemelked®egyszerestelí-
tetlenség,közelállandótöbbszöröstelítetlenség,kompetitívn3-n6zsírsav
arányokésnagyoner®snegatívallometrikusösszefüggéstmutatódokoza-
hexaénsavarány(8. ábra)). Érdekesésfontostény,hogyezenösszetételi
sajátosságokatmégaPLzsírsavprolraszignikánshatástgyakorlótakar-
mányeredet¶zsírsavaksemfedtékel. Ennekmegfelel®enmegállapítható,
hogyazagy kivételévelalegtöbb,metabolikusanaktívszervbenjólnyo-
mon követhet® a membránok, mint ütemadókelv biokémiai szabályozó
szerepe.
8. és9. ábraAdokozahexaénsav(DHA)ésamalondialdehid(MDA)
allometrikusváltozásaiavizsgáltnyolmadárfajvese,máj,tüd®ésagyi
foszfolipidjeiben
AMDAtestsúlyfügg®konentráióváltozása (9. ábra)avesében,máj-
banéstüd®benarrautal,hogyamembránlipidektelítetlenségeperoxidá-
ióvalszembenérzékennyétesziezen szerveklipidjeit, melybenkulssze-
repetkapadokozahexaénsav.
3.1.2.5. Eltér® kifejlett kori testsúllyal rendelkez® madárfajok
tüd®parenhymaéssurfatantelemzése,allometrikus módszer-
tannal
Amadárlégz®szervrendszerénvégzettvizsgálatkérdésfelvetésea3.1.2.4.
fejezetben egy részleteiben nemvizsgált ponthoz kapsolódott, melyben
ateljes madártüd®összes foszfolipidjének zsírsavproljában amembrá-
nok, mint az anyagsere ütemadói elmélet igazolhatónak bizonyult. A
madártüd®azonban foszfolipidekben igen gazdag (a. 80%)surfatan-
totis tartalmaz (illetve szekretál), melynek a teljes PL zsírsavprolhoz
való hozzájárulása afentivizsgálatban ismeretlenmaradt. Jelen munka
élkit¶zéseazvolt,hogyelemezzeamadársurfatantPLzsírsavproljá-
banesetlegesentestsúly-függést(áttételesenpedig alapanyasere intenzi-
tás)mutatózsírsavakat,valamintkülönfelmérjeasurfatantmentestüd®
parenhymaszerepétateljestüd®rekapottallometrikusösszefüggésekki-
alakításában. F®bb eredményeinkszerint a madár teljes tüd®szöveti és
tüd®parenhymálisfoszfolipidjeihatározottannegatívallometrikusössze-
függést mutatnakaDHAvonatkozásban, viszonylagkiegyenlítettpolién
zsírsavszintmellett,amembránok,mintazanyagsereütemadóielmélet-
nekmegfelel®en. Kisebbmeredekséggelugyan,deasurfatantPLfraki-
ójában(mintszekretált,ésnemstrukturálisfoszfolipidben)isigazoltuka
DHAnegatívallometrikusösszefüggését (10. ábra).
10. és11. ábraAdokozahexaénsav(DHA)arányánakésa
malondialdehid(MDA)konentráiójánakallometrikusváltozásai
madarakteljestüd®szövetében,tüd®parenhymájában,illetvea
surfatantfoszfolipidjeiben
A szöveti malondialdehid konentráiót a natív ésa mosott tüd®szö-
vetb®lhatároztukmeg,ésmindkétesetbener®sennegatívtestsúlyfüggést
igazoltunk(11. ábra). AzegyediMDAésDHAadatokközöttikorreláió
közepesnek,illetveszorosnakbizonyultakétszövettípusesetében(0,39és
0,679anatívésamosottmintákban,rendre).
Eredményeinkszerintamadárlégz®szervrendszerében(mindatüd®pa-
renhymában,mindpedigasurfatantban)adokozahexaénsavkiemelten
fontosszabályozószereppelrendelkezikmintmembránlipidkomponensés
mintasurfatant szekréiómértékétbefolyásolófaktor. ADHArészará-
nyaésanemenzimatikus lipidperoxidáió szoros korrelatívösszefüggése
arrautal,hogyamembránlipidektöbbszöröstelítetlensége,különösenpe-
digaDHAarányajelent®senbefolyásoljaaperoxidáiómértékétamadár
pulmonárisfoszfolipidjeiben.
3.1.3. A szöveti triglieridek összetételének natív alakulása; a
faj-ésfajtaazonosításlehet®ségeiaTGmolekulaszerkezetrész-
letesanalízisére alapozva (regiospeikus analízis)
3.1.3.1. A fajspeikus tulajdonságok elemzése, triglieridek
molekulaszerkezeti vizsgálatáraalapozva
Napjainkban egyre nagyobb és fontosabb feladat az állati termékek
eredetének igazolása. A szöveti lipidek élelmiszertermékek szintjén már
kevéssé alkalmasak az eredet igazolására, miután az adiposasejtek DNS
tartalmaalasony,másrészt azok leginkább olvasztott állapotbantartal-
mazzákaz állati zsírokat. Tudvalev® az is, hogy a takarmány hatása a
neutrálislipidekösszetételérenagyonmarkáns(3.1.4. fejezet), ígyameg-
bízhatófaj-vagyfajta azonosításlegtöbbszörnemlehetséges. Jelenvizs-
gálatmunkahipotéziseazvolt,hogyafajiszint¶különbségeketaTGmo-
lekulaszerkezetbenrészleteibenfeltárjuk,majdazeredményekalapjánfaj
elkülönítési lehet®ségeket vizsgáljuk, hét állatfaj (házisertés, vaddisznó,
borz,nyúl,lúd,gím-ésjávorszarvas)mintáiraalapozva. Amódszertana
triglieridekrészleges hidrolízisén ésazsírsavak helyzetspeikus (1,3és
2)összetételielemzésénalapult.
Megállapítottuk, hogyfajtól függetlenül, deasertéskivételévela 2-es
helyzetbenazsírsavaklánhosszameghaladjaaTG-ek(ésaz1,3helyzet)
savainaklánhosszát. Asertések,atöbbifajtóleltér®ena2-eshelyzetben
magasabbtelítetlenségetmutattak,minta triglierid molekula1,3 pozí-
iójában. Érdekesmódon avadésháziasítottsertés genotípusokTGés
2MGzsírsavproljanemtérteljelent®sebben. ATGmolekulaszerkezeti
vizsgálatávaligazoltukaztatapasztalatimeggyelést,hogyazonosszén-
lánhosszúságú éstelítetlenség¶,de szimmetrikusésaszimmetrikuszsír-
savak közül el®bbiekpreferáltanatriglierid 2-espozíiójában aileznek
(12. ábra).
12. ábraAzonoslánhosszúságúéstelítetlenség¶,szimmetrikus(C18:1
n9)ésaszimmetrikus(C18:1n11)zsírsavakglierinailezési
prefereniája,fajonként
A kidolgozott módszertan kis mintaigény¶ (a. 50 mg TG, melyb®l
anatív ésahidrolizált forma iselemezhet®), viszonylag jól ismételhet®,
ésjól kivitelezhet® mindenmagas zsírtartalmúszövetmintán,vagy akár
olvasztott zsírokonis. A módszer limitáiójaként említhet® annaknagy
manuálismunkaigényeésrelatívmagaslaborköltsége(extrakió,hidrolízis,
vékonyrétegkromatográaésmintánkéntkétgázkromatográásanalízis).
Amódszermegbízhatóságát ésfelbontását atovábbiakbanmárnem
fajok,hanemeltér®fajtákelkülönítésénteszteltük(3.1.3.2.). Agyakorlat-
bannagyonsokszormerülfelugyanisazakérdés,hogyahungarikumként
számontartott mangalia mintákvagytermékek(különösenzsíréshús)
elkülöníthet®k-eahússertésekmintáitól.
3.1.3.2. Afajtaszint¶eltérésekvizsgálatasertésgenotípusoktri-
glieridjeinekmolekulaszerkezetivizsgálatával
Irodalmiéssaját(3.1.3.1.) adatokalapjánisismert,hogyazsírsavakel-
rendez®déseaTGmolekulánbelülnemrandom. AsertésTG-ekspeiális
zsírsav-megoszlására, nevezetesen a entrálisan igen gyakori palmitinsav
el®fordulásra Hildith és Stainsby (Biohem. J., 1935, 29: 90-99) már
igenkoránutalásttettek. Vizsgálatunkéljanemsakazvolt,hogyazed-
digiekbenkevésséismertszervek(szív-ésvázizom,hátszalonna,tüd®,vese,
májéslép)esetébenjellemezzükaTGés2-monoglierid(2MG)zsírsav-
prolt,hanemhogyesetlegesfajta-elkülönítést(mangaliaéshússertés)is
lehet®vétegyünk. Azeredményekhatszervbenmindazsírsavprol,mind
pedigazegyeszsírsavakhelyzetielrendez®désetekintetébenakétfajtában
meglehet®senhasonlókvoltak,amájkivételével,aholapalmitinsavnagy
entrálisel®fordulásigyakoriságanemvoltigazolható.
A sertés fajták TG-jeinek és 2MG-jeinek direkt analízisére alapozva
igazoltukamájbanéstovábbihatszervben,hogyajellegzetessertésTG
mintázatamájatkövet®enalakulki. Afajtákközöttieltérésekminimáli-
sakvoltak,els®sorbanazsírszövetbentaláltunkmegbízhatóelkülönítésre
alapotadókülönbséget,amennyibenapalmitinsavteljesmennyiségeman-
galiábana2-eshelyzetbenfordult el®,míghússertésbensakvalamivel
alasonyabb2-eshelyzetigyakoriságotigazoltunk(a. 90%). AHMFésa
regiospeikus analízis, úgyt¶nik,fajok szintjéig nyújtlehet®séget meg-
bízhatókülönbségtételre, amódszer felbontása afajtákközötti eltérések
megállapítására már kevésbé volt alkalmas (amennyiben léteznekfajták
közöttieltérések),mégkéttávoli fajta esetébenis.
3.1.4. A takarmányeredet¶ zsírsavak szövetilipidekbe és lipid-
frakiókba történ® beépülésének (inkorporáió) vizsgálata, ha-
lakban
3.1.4.1. Afrikaiharsa(Clariasgariepinus)lézsírsavösszetétele
éshúsmin®sége,növényiolajok etetésétkövet®en
Atakarmány-,illetvetáplálékeredet¶zsírsavakközülkülönösenatöbb-
szörösentelítetlenn3soportnaktulajdoníthatókedvez®táplálkozás-élet-
tanihatás. A humántáplálkozássorán felvettn3zsírsavakjelent®srésze
(els®sorbanahosszabbszénlánalrendelkez®k)halhúsbólvagyhaltermé-
kekb®l származik. A haltakarmányozásban felhasznált zsírsavak napja-
inkradönt®ennemhal,hanemnövényieredet¶ek,melyekhatékonyságaa
halhúsel®állításbanmégnemteljesmértékbenismert.Vizsgálatunkbana
halolajmellettaszója-éslenolajafrikaiharsalébe(annakteljesszöveti
zsírsavproljába)történ®beépülésiütemét, valamintazeltér®olajok lé
min®ségregyakorolthatásaitelemeztük.
Megállapítottuk, hogy a halolaj részleges kiváltása (partial replae-
ment)növényiolajokkalnembefolyásoltahátrányosanaz afrikaiharsa
növekedési ütemét, míg a relatív magas n3 zsírsavbevitel fokozta a lé
spontán (sepegési veszteség) és indukált (felengedtetési veszteség) víz-
vesztésihajlamát. Azn3zsírsavak metabolizmusábanmégaszokatlanul
magasn3 prekurzorbevitelsem eredményezett fokozott hosszúlánú n3
zsírsavszintetikusaktivitástafrikaiharsában. Alézsírsaveredmények
elemzéséreazún. zsírsavkihígulásimodellt(2.3. fejezet)alkalmaztuk,a
beslésta3.hetiadatokravonatkozóan,a6. hetiadatsormintreferenia
felhasználásávalvégeztük. Mindanövényiolajok,mindpedigakontroll-
kéntalkalmazotthalolajf®bbzsírsavainakléösszlipidjeibetörtén®inkor-
poráiója(melyigen hatékonyvolt)márigenrövidid®-intervallumbanis
relatívjóljellemezhet®nekbizonyultazegyszer¶zsírsavkihígulásimodell
alkalmazásával. Eredményeinkels®sorbana befejez®takarmányozásésa
halhúszsírsavproljánakoptimálismérték¶módosításakapsánlehetnek
hasznosíthatókagyakorlatban.
3.1.4.2. Növényiolajokszövetilipidfrakiókbavalóbeépülésének
elemzéseNílusitilápia (Oreohromis nilotius)szerveiben
A3.1.4.1. fejezetbenbemutatottadatállományhalhúsraésösszlipidekre
vonatkozóan mintegyigazolta,hogyatakarmányváltássorán megváltozó
lipidösszetétel azsírsavak szintjén a szöveti pool kihígulásához vezet.
Azelemzésugyanakkortöbbelhanyagolássalisjárt: nemelemeztekülön
a legf®bb lipidfrakiókat (triglieridek ésfoszfolipidek), valamint a nem
mindenesetbenmagaszsírtartalmúléreirányult. Atovábbiakbanade-
pózsír,amáj,aléésagonádokelemzésétvégeztükel,lipidfrakiónként,
ésafentinéltöbbször mintázott(4mintavételiid®pont)etetési periódus-
ban,tilápián. Atestsúlyahatodikhétvégéreasoportokközöttszigni-
kánsaneltért,akövetkez®sorrendetkövetve: halolaj>lenolaj>szójaolaj
kezelés. Ahalolajetetésmellettalésúlya(6. hét)ismeghaladtaaszó-
jaolajkezelésnélmértértékeket. Atilápiaszövetekeltér®lipidfrakióinak
zsírsavproljábanazttapasztaltuk,hogyatakarmányeredet¶növényiola-
jok zsírsavai nagyon nagyhatékonysággal, éssok esetben az egyszer¶
kihígulásimodellel -jólbesülhet®mértékbenépülnekbeatilápia léés
májtriglieridekbeésfoszfolipidekbe,deakétfrakióbaeltér®arányban.
Eredményeink szerint atilápia képesatakarmánybanbiztosított zsírsav
prekurzort(
α
-linolénsav)DHA-vá konvertálni, a lé triglieridekvonat- kozásában. Igazoltuk továbbá, hogy atakarmány lipidjei atelítetlenségmértékénekmegfelel®enfokozzák amájbeliinvivolipidperoxidáiót.
4. Összefoglalás
Adisszertáióbafoglalteredmények2004 ótaelkészült,alegtöbbeset-
benaz OTKA (D048413, 83150), az NKTH (4/023, 3/023)és az MTA
(BO/108/07 és BO/26/11/4) által támogatott kutatási projektekb®l, a
posztdoktoriésazaztkövet®id®szakbólszármaznakésnégynagytéma-
körbesorolhatók. Tekintettel arra, hogy ezekjelent®sen elkülönülnek,a
f®bberedményeketisnégyestagolásbanfoglalomössze.
4.1. Jelent®sen eltér® energetikai állapotok meta-
bolikushatásainakjellemzésebaromfajokklinikai-
kémiai vizsgálatával
Azáltalunkis alkalmazottklinikai-biokémaimódszertanazállatorvosi
dignosztikábanszéleskörbenelterjedt,kvázirutineljárás. Ennekellenére
saját, els®sorban baromfajokon, többkorsoportban végzettrendszeres
elemzéseinkszintehiánypótlónakszámítottak,els®sorbanavizsgált(spe-
iálisankezeltvagyszelektált)állományokegyirányúszelekiója okán. A
tojótyúkokmesterségesvedletésesoránrészletesenjellemeztükaztameta-
bolikusésenzimatikusadaptáiót,melyeta12napiteljestakarmánymeg-
vonásindukált. Amódszeralkalmasvoltalipidtartalékokkimerülésének
fokozatos nyomonkövetésére, valaminta nagymérték¶fehérje kataboliz-
mus,illetveasökkentzikaiaktivitásjellegzeteshatásainakpontosleírá-
sára. A hústermelésreszelektált brojler éspulykaállományokonvégzett
részletes,afelneveléstöbbid®pontjánkivitelezettelemzésmindasonto-
zat,mindpediganagynövekedésiütem¶vázizomzatfejl®désénekpontos
jellemzésérevoltalkalmas. Kiemelked®fontosságú eredmény,hogymind-
kétfajban már a kelést követ®en el®rejelezhet®nek bizonyult a fokozott
izomhipertróaokoztaszakrolemmasérülés(kreatinkináz). Amódszer-
tannemsakapatoziológiás,hanemanormálélettanifolyamatok(tojás-
szikdepléió, vedlés, lassuló osszikáió)pontoskimutatásátis lehet®vé
tette.
4.2. Az energetikai status hatása a szöveti lipid-
frakiók zsírsavösszetételére madárfajokban
A4.1. pontbanvérbiokémiaianalízisselvizsgáltmadarak(speiálisan
kezeltvagyszelektáltállományok)olyankülönlegesenergetikaisajátossá-
gokkalvagyeltérésekkelrendelkeznek,melyekszervezetszint¶adaptáiót
feltételeznek. Asejtmembránokszintjénfennállólipid-fehérje interakiók
okánamembránlipidekmintazanyagsereütemadóiértelmezhet®k,me-
lyetHulbertésElse(J.Theor. Biol.,1999,257277)átfogóelméletükben
amembránlipidösszetétel,aszövetiszint¶anyagsere,végülpedigaszer-
vezetalapanyagsereintenzitásánakkorrelatívösszefüggésekéntértelmez-
tek.Azállandótesth®méséklet¶állatokszerveirevonatkozóanels®sorban
eml®seredményekvoltak elérhet®k. A madarak, különösenpedig a rö-
vidélettartamú,er®sszövetilipidperoxidáióvaljellemezhet®baromfajok
körében szívizom,vese, máj, tüd® (teljes tüd®ésparenyma) éssurfa-
tantstruktúrálisésszekretáltfoszfolipidekbenigazoltukamembránlipidek
többzsírsavkomponensénekszignikánstestsúlyfüggését. Azáltalunkle-
írtösszefüggésekegybevágnakametabolikusütemdóelmélettel,bizonyos
mértékbenkiterjesztveazt(szekretáltfoszfolipidekre). Abaromfajokkö-
rébenvégzettelemzéssorán mindenemlítettszövettípusbanismétl®d®en
ésmegbízhatóan igazoltuk, hogy a testsúly(az azzal allometrikus kap-
solatbanállóalapanyagsereintenzitásrévén)negatívkapsolatbanálla
membránlipidekdokozahexaénsav részarányával, melypedig szignikáns
korreláiótmutataszövetilipidperoxidáió mértékével.
4.3. A szöveti triglieridek összetételének natív
alakulása; a faj- és fajta azonosítás lehet®ségei a
TGmolekulaszerkezetrészletesanalízisérealapozva
(regiospeikus analízis)
Els®lépésbenhét faj b®ralatti zsírszövetében,majdpedig két, er®sen
eltér®sertésfajta hétszervénektriglieridtartalmábanvizsgáltukatrigli-
eridmolekulaszerkezetet(azsírsavakhelyzetimegoszlása),azesetlegesen
fennállófaj-,illetvefajta-determinált eltérésekkimutatására, illetve erre
alapozvaafaj-vagyfajtaazonosításlehet®ségeérdekében.Akidolgozott,
részleges hidrolízisen és többszörös kromatográás elválasztáson alapuló
mintael®készítés, majd a 2-monoglierid és triglierid frakiókra vonat-
koztatottgázkromatográás eredményekértelmezéséreúj mutatótvezet-
tünkbe. Azún. helyzeti megoszlási faktora glierint széls® ésközéps®
helyzetbenészterez®zsírsavak hányadosánaklogaritmusakéntgyakorlati-
lagfüggetlenatakarmányeredet¶zsírsavaktól,ugyanisinformáiótartalma
atriglierid molekulafelépítéséreirányul. Ahelyzetimegoszlásifaktorra
alapozvakér®dz®ésmonogasztrikus eml®sésmadárfajokbanminden-
ev®, növényev® és ragadozó táplálkozás mellett meghatároztuk a faji
sajátosságokataregiospeikuszsírsavprolban,faj-elkülönítéstlehet®vé
téve. A módszerasertésfajták megbízhatóelkülönítésérenembizonyult
alkalmasnak, igazoltuk ugyanakkor azt a bioszintetikus szabályosságot,
melyszerintazonosszénlánhosszúságúéstelítetlenség¶zsírsavakközüla
szimmetrikusformaközéps®helyzetiailezésepreferált(aglierinmoleku-
lán).Eredményeinkazsírszövetsajátosságai(alasonyoxidatívérzékeny-
ség)okánakártovábbfeldolgozottélemiszertermékekeredeténekigazolása
soránisalkalmazhatóklehetnek.
4.4. A takarmányeredet¶ zsírsavak szöveti lipi-
dekbeéslipidfrakiókbatörtén®beépülésénekvizs-
gálata, halakban
Azn3zsírsavakhalolajhelyettnövényiolajjaltörtén®bevitelenapjaink
hal-takarmányozásában igen elterjedt módszer, mely alapvet®en kihasz-
nálja a halak fokozott zsírsav-elongáiós képességét. Részleteiben nem
ismertugyanakkor, hogya nagymérték¶n3zsírsav bevitelemilyen hús-
min®ségiváltozásokkaljár,illetvehogyazsírsavakinkorporáiójamilyen
dinamikát mutat, különösen eltér®funkiójúés összetétel¶ lipidfrakiók
esetében. Vizsgálatainksoránigazoltuk,hogyafokozott,növényieredet¶
n3zsírsavbevitelkedvez®tlenülhatahalhúsvíztartóképességére,illetveaz
egyszer¶zsírsavkihígulásimodelleredményeirealapozvaajánlásttettünk
afrikai harsa esetébena befejez® takarmányozás hosszára vonatkozóan.
Tilápia szöveti lipidfrakióiban négyhetes etetési periódust követ®en jól
besülhet® zsírsav beépülésiütemet igazoltunk, mely jólegybevágott az
afrikaiharsaesetébentapasztaltakkal, ugyanakkorer®senfokoztaaszö-
veti lipidperoxidáiót, ezzel negatívanbefolyásolva a hal-eredet¶termék
min®ségét,oxidatívstabilitását.
5. Új tudományos eredmények
5.1. Jelent®sen eltér® energetikai állapotok meta-
bolikushatásainakjellemzésebaromfajokklinikai-
kémiai vizsgálatával
Tojótyúkokmesterségesvedletéseminter®senenergiahiányosállapot
idejénrészletesenjellemeztükamadarakmetabolikusreakióit,vérbioké-
miaiparaméterekrealapozva.
Nagy növekedési erély¶, hústermel®pulykaés brojlersirke állomá-
nyokbanszéleskör¶,sreeningjelleg¶klinikai-kémiaiadatállománytközöl-
tünk,ateljesfelnevelésiperiódusravonatkozóanésfelhívtukagyelmeta
hústermelésreirányuló,egyoldalúszelekiókároshatásaira(kreatinkináz,
K,laktátdehidrogenáz).
5.2. Az energetikai status hatása a szöveti lipid-
frakiók zsírsavösszetételére madárfajokban
Els®ként igazoltuk, hogyamadárszívmembránlipidekzsírsavössze-
tétele,aszövetimalondialdehidtartalomésatestsúlyközöttszoros,allo-
metrikusösszefüggésállfenn.
Els®kéntközöltünkadatokatamadárvese, teljestüd®ésmájösszes
foszfolipidfrakiózsírsavprolja,valamintazinvivolipidperoxidáióésa
testsúlyszignikáns,allometrikusösszefüggésérevonatkozóan.
Els®kéntigazoltuk,hogyateljesmadártüd®benmindaparenhyma,
mindpedigasurfatant foszfolipidekzsírsavprolja(struktúrálisésszek-
retáltfoszfolipidek)szignikáns,allometrikusösszetételiszabályosságokat
mutat, melyben meghatározóa dokozahexaénsav részarányának negatív
allometrikusalakulása.
5.3. A szöveti triglieridek összetételének natív
alakulása; a faj- és fajta azonosítás lehet®ségei a
TGmolekulaszerkezetrészletesanalízisérealapozva
(regiospeikus analízis)
Hatékony módszert fejlesztettünk a szöveti triglieridek fajonkénti
eltéréseinekvizsgálatára,lehet®ségetbiztosítvaafajokmintáinakmegbíz-
hatóelkülönítésére is,egyolyanszövettípusban,melygenetikaivizsgála-
tokrakevésbéalkalmas.
5.4. A takarmányeredet¶ zsírsavak szöveti lipi-
dekbeéslipidfrakiókbatörtén®beépülésénekvizs-
gálata, halakban
Igazoltuk,hogyafrikaiharsa léjébenafokozotttakarmányeredet¶
n3zsírsavbeviteljelent®senésrészbenkedvez®tlenülmódosítjaahúsmin®-
séget,azsírsavakinkorporáiójapedig jólbesülhet®azsírsavkihígulási
modellsegítségével.
Igazoltuk, hogy nemsak a ragadozó, hanem növényev® halakban,
azok több szövetének f®bb lipidfrakióiban jelent®s és jól el®rejelezhet®
arány-összetételiváltozástidéznekel®atakarmányeredet¶n3zsírsavak.
6. A gyakorlatban hasznosítható eredmé-
nyek
6.1. Jelent®sen eltér® energetikai állapotok meta-
bolikushatásainakjellemzésebaromfajokklinikai-
kémiai vizsgálatával
Tojótyúkesetébenakorábbanszéleskörbenalkalmazottésazótabe-
tiltott,teljestakarmánymegvonásonalapulómesterségesvedletésimódszer
kedvez®tlenélettanihatásairavonatkozóadatokatközöltünk.
Intenzívhústermelésreszelektáltbrojleréspulykavonalakbanátfogó
jellemzéstadtunkspeiálisbarom populáiókklinikai-kémiaiparaméte-
reir®l,többkorsoportban,melyatermel®állományokszintjénrefereni-
akéntalkalmazhatónakbizonyult.
6.2. Az energetikai status hatásának elemzése a
szöveti lipidfrakiók zsírsavösszetételére madárfa-
jokban
Apulykamellizomzatimembránlipidekatipikusösszetételi változása
afelnevelés alatt(kapsolódvaavérklinikai-kémiaiparamétereirevonat-
kozóadatokhoz) felhívja a gyelmet e nagy növekedési erély¶madárfaj
szelekiójánakhatározottankáros(ésrészleteibeneddigismeretlen)hatá-
saira.
A testméret, a metabolikusan aktív szervek membránlipid zsírsav-
összetétele és a szöveti lipidperoxidáió mértékének szoros összefüggése
els®sorbanavárhatóélettartamésaperoxidatívkárosodáskapsánbizto-
sítúj,beslésekreisalkalmasadatállományt.
6.3. A szöveti triglieridek összetételének natív
alakulásána leírásak a faj- és fajta azonosítás le-
het®ségeinek felmérése érdekében
Aszövetitriglieridekfajonkéntieltéréseinekfeltárásafriss,tároltés
továbbfeldolgozott (esetlegtermékbekevert)zsírokesetébenis alkalmaz-
hatómódszertan,melyatermékekeredetazonosításábanjelent®stámpont
lehet,els®sorbanazélelmiszeriaparban(deakárigazságügyivizsgálatokis
felmerülhetnek). Fontoskiegészítés,hogyakidolgozott módszerakárol-
vasztással nyert állati zsiradékokon is alkalmazható, egy olyan, jelent®s
mennyiségbenel®állóállatiterméken,melyesetébenagenetikaialapúfaj-
meghatározásnemlehetséges.
6.4. A takarmányeredet¶ zsírsavak szöveti lipi-
dekbeéslipidfrakiókbatörtén®beépülésénekvizs-
gálata, halakban
A takarmányeredet¶n3zsírsavak fokozottésid®zített, vagymegha-
tározottid®tartamúbevitele,illetveavárhatószövetizsírsavprolpontos
besülhet®ségenagybansegíti a befejez®takarmányozásoptimalizálását.
A kihígulási modell alkalmazása a bevont paraméterekre alapozva költ-
séges analitikai eljárást, agázkromatográás zsírsavprol elemzést segít
kiváltaniegyszer¶súlyméréssel.
7. A tézisek fejezeteinek megfeleltethet®
saját publikáiók listája
7.1.1. Eltér®energetikaiállapotokmetabolikushatásainakjellem-
zésebaromfajok klinikai-kémiai vizsgálatával
7.1.1.1. Tojótyúk mesterséges vedletése,a koplalás vérparamé-
terekkeltörtén® nyomonkövetése
Romvári,R.,Szabó,A.,Andrássy-Baka,G., Süt®,Z. Molnár,T., Bázár,
Gy.,Horn,P.(2005): Trakingforedmoultbyomputertomographyand
serumbiohemialparametersinlayinghens. Arh.Geügelk. 69(6):245-
251.
7.1.1.2. Hústermelésre szelektált pulykavérbiokémiai paramé-
tereineknyomonkövetése,akelést®la vágásig
Szabó, A.,Mézes, M., Horn, P.,Süt®, Z., Romvári,R.(2005): Develop-
mental dynamisof some blood biohemial parameters inthe growing
turkey(Meleagrisgallopavo). AtaVet. Hung. 53(4):397-409.
7.1.1.3. Abrojlersirke kliniko-kémiaivizsgálata(follow-up) a
teljesfelnevelésalatt
Szabó,A.,Milisits,G.(2007): Cliniohemialfollow-upofbroilerrearing
-ave-weekstudy. AtaVet. Hung. 55(4):451-462.
7.1.2. Az energetikai status hatása a szöveti lipidfrakiók zsír-
savösszetételére madárfajokban
7.1.2.1. Tojótyúkokmesterségesvedletése,adrasztikusenergia-
megvonáshatásaamájésszíveltér®lipidfrakióinak(triglierid
ésfoszfolipid)összetételére
Szabó,A.,Fébel,H.,Mézes,M.,Horn,P.,Balogh,K.,Romvári,R.(2005):
Dierentialutilizationofhepatiandmyoardialfattyaidsduringfored
moultoflayinghens. PoultrySi. 84(1):106-112.
7.1.2.2. Pulykamellizommembránlipidekontogenezisalattivál-
tozásainakleírása
Szabó,A.,Fébel,H.,Horn,P.,Bázár,Gy.,Romvári,R.(2006): Ontogeni
developmentofthefattyaylhainompositionoftheturkey(Meleagris
gallopavo) petoralis musle membranes(an allometriapproah). Ata
Biol. Hung.57(2):165-180.
7.1.2.3. Eltér® kifejlett kori testsúlyú madárfajok szívizmának
membránlipidjei,allometrikusanalízis
Szabó,A.,Fébel,H.,Mézes,M.,Balogh,K.,Horn,P.,Romvári,R.(2006):
Bodysize relatedadaptationsoftheavianmyoardialphospholipidfatty
aylhainomposition.Comp. Biohem. Physiol. B.144(4):496-502.
7.1.2.4. Eltér® kifejlett kori testsúlyú madárfajok tüd®, vese,
májésagyfoszfolipid zsírsavproljának allometrikus elemzése
Szabó,A.,Mézes, M., Romvári,R.,Fébel,H.(2010): Allometrisaling
offattyayl hainsinfowlliver,lungand kidneybutnotinbrainphos-
pholipids. Comp. Biohem. Physiol. B.155(3):301-308.
7.1.2.5. Atüd®parenhymaliséssurfatantfoszfolipidjeinekal-
lometrikuselemzése baromfajokban
Szabó,A.,Mézes,M.,Balogh,K.,Romvári,R.,Horn,P.,Fébel,H.(2012):
Negativeallometry ofdoosahexaenoiaid inthefowllungand pulmo-
narysurfatantphospholipids. AtaBiol. Hung. 63(2):202-217.
7.1.3. A szöveti triglieridek összetételének natív alakulása; a
faj-ésfajtaazonosításlehet®ségeiaTGmolekulaszerkezetrész-
letesanalízisére alapozva (regiospeikus analízis)
7.1.3.1. Afajspeikustulajdonságokelemzésetriglieridekmo-
lekulaszerkezetivizsgálatára alapozva
Szabó,A.,Fébel,H.,Sugár,L., Romvári,R.(2007): Fattyaid regiodis-
tributionanalysisofdivergentanimaltriaylglyerolsamplesapossible
approahforspeiesdierentiation.J.FoodLipids. 14(1):62-77.
7.1.3.2. Afajtaszint¶eltérésekvizsgálatasertésgenotípusoktri-
glieridjeinekmolekulaszerkezetivizsgálatával
Szabó,A.,Horn,P.,Romvári,R.,Házas,Z.,Fébel,H.(2010):Comparison
ofMangaliaandHungarianLargeWhitepigsatidentialbodyweightII.
Fattyaid regiodistributionanalysisof thetriaylglyerols. Arhivfür
Tierzuht. 53(2):147-161.
7.1.4. A takarmányeredet¶ zsírsavak szövetilipidekbe éslipid-
frakiókba történ® beépülésének (inkorporáió) vizsgálata, ha-
lakban
7.1.4.1. Afrikaiharsalézsírsavösszetételeéshúsmin®sége,nö-
vényiolajok etetésétkövet®en
Szabó,A.,Romvári,R.,Szathmári,L.,Molnár,T.,Losmándi,L.,Bázár,
Gy.,Molnár,E.,Horn,P.,Hanz,Cs. (2009): Eetsofdietaryvegetable
oilsupplementationonlletquality traits,hemialand fatty aid om-
positionofAfrianatsh(Clariasgariepinus). ArhivfürTierzuht. 52:
321-333.
7.1.4.2. Növényi olajok szöveti lipidfrakiókba való beépülésé-
nekelemzése Nílusi tilápia (Oreohromis nilotius)szerveiben
Szabó,A.,Mézes,M.,Hanz,Cs.,Molnár,T.,Romvári,R.,Varga,D.,Fé-
bel,H.(2011): Inorporationdynamisofdietaryvegetableoilfattyaids
intothetriaylglyerolsandphospholipidsoftilapia(Oreohromis niloti-
us)tissues(llet,liver,viseralfatandgonads). AquaultureNutrition.
(17):132-147.
8. Köszönetnyilvánítás
HálásköszönetemetszeretnémkifejezniHornPéterakadémikusésRom-
váriRóbertprofesszor uraknakazértaz1998 ótanyújtottönzetlenszak-
maiéserkölsitámogatásért,melynélkülsemabemutatottmunka,sema
dolgozatnemkészültvolnael. Köszönömmentoraimnak,KovásMelinda
akadémikus asszonynak és Sarudi Imre professzor úrnak a szemléletfor-
málótanítást! Ezútonszeretném ®szinteköszönetemetkifejezni Szendr®
Zsolt professzor úrnak támogatásáért, Dr. Fébel Hedvignekaz analiti-
kaimunkáért,MézesMiklósakadémikusúrnakalipidperoxidáióvalkap-
solatos segítségéért. Az egyetemi források mellett OTKA Iroda és az
Akadémia Bolyai János Kutatási Ösztöndíj Kuratóriuma a bemutatott
eredmények megteremtését öt, illetve hat éves id®szakokban támogatta,
amiértehelyenisköszönetetmondok.
KülönköszönetteléshálávaltartozomCsaládomnakésSzüleimnek,tü-
relmükértéstoleraniájukért.