REGULAR ARTICLE
The insulin receptor is differentially expressed in somatic and visceral primary sensory neurons
Bence András Lázár1,2&Gábor Jancsó2&István Nagy3&Viktor Horváth4&Péter Sántha2
Received: 5 December 2017 / Accepted: 4 June 2018
#Springer-Verlag GmbH Germany, part of Springer Nature 2018
Abstract
Recent studies demonstrated the expression of the insulin receptor (InsR) and its functional interaction with the transient receptor potential vanilloid type 1 receptor (TRPV1) in primary sensory neurons (PSNs). The present study was undertaken to reveal the target-specific expression of the InsR and its co-localization with the TRPV1 in rat PSNs. We assessed the localization of the InsR and its co-localization with the TRPV1 in PSNs retrogradely labelled with biotin-conjugated wheat germ agglutinin injected into the dorsal hind paw skin, the gastrocnemius muscle, the pancreas and the urinary bladder wall.
The largest proportions of retrogradely labelled InsR-immunoreactive neurons were identified among PSNs serving the pancreas (~ 54%) and the urinary bladder (~ 53%). The proportions of retrogradely labelled InsR-immunoreactive neurons innervating the dorsal hind paw skin and the gastrocnemius muscle amounted to ~ 22 and ~ 21%. TRPV1-immunoreactive neurons amounted to ~ 63, ~ 62, ~ 67 and ~ 65% of retrogradely labelled cutaneous, muscle, pancreatic and urinary bladder PSNs, respectively. Co-localization of the TRPV1 with the InsR was observed in ~ 16, ~ 15, ~ 29 and ~ 30% of retrogradely labelled cutaneous, muscle, pancreatic and urinary bladder PSNs. These quantitative immunohistochemical data demon- strate a preponderance of InsR-immunoreactivity among PSNs, which innervate visceral targets. The present findings suggest that visceral spinal PSNs are more likely to be exposed to the modulatory effects of insulin on sensory functions, including neurotrophic, nociceptive and inflammatory processes.
Keywords Insulin receptor . Transient receptor potential vanilloid type 1 receptor . Primary sensory neurons . Retrograde labelling . Somatic and visceral organs
Introduction
Insulin, apart from being a pivotal regulator of body metabo- lism, is significantly involved in various neuronal processes, such as neuronal survival, initiation of neurite outgrowth and regulation of neuronal activity (Recio-Pinto et al. 1986;
Fernyhough et al.1993; Wan et al.1997; Barber et al.2001;
Stella et al. 2001). Recently, it has also been revealed that neural actions of insulin are mediated by the InsR, which has been demonstrated in the nerve tissue, too (Sugimoto et al.2000; Figlewicz2016). Insulin, at physiological concentra- tions, enhances neurite outgrowth of cultured primary sensory neurons (PSNs) (Fernyhough et al.1993; Singh et al.2012) and exerts trophic effects on a specific subpopulation of PSNs that is sensitive to nerve growth factor (NGF) (Recio-Pinto et al.1984; Recio-Pinto and Ishii1988; Fernyhough et al.1993).
In addition, experiments on cultured rat PSNs demonstrated that insulin enhances the capsaicin-induced cobalt uptake resulting from the activation of the nociceptive ion channel, the transient receptor potential vanilloid type 1 receptor (TRPV1) (Sathianathan et al. 2003). TRPV1-expressing PSNs comprise about 60% of C-fibre ganglion neurons and transmit nerve impulses generated by noxious heat and painful chemical stimuli in somatic and visceral organs (Jancsó et al.
1977; Buck and Burks1986; Holzer1991; Caterina and Julius 2001). In addition to being essential in the transmission of
* Bence András Lázár b.a.lazar.md@gmail.com
1 Department of Psychiatry, University of Szeged, Kálvária sugárút 57, Szeged H-6725, Hungary
2 Department of Physiology, University of Szeged, Szeged, Hungary
3 Department of Surgery and Cancer, Imperial College London, London, UK
4 First Department of Internal Medicine, Semmelweis University, Budapest, Hungary
https://doi.org/10.1007/s00441-018-2868-0
nociceptive information towards the central nervous system, this particular class of PSNs is implicated in the initiation and maintenance of sensory neurogenic vascular and inflammato- ry reactions collectively termed the neurogenic inflammatory response (Jancsó et al.1967,1977; Holzer1991). Further, it has been reported that nociceptive afferent nerves may also promote secretory, contractile and immune functions in the innervated organs (Maggi and Meli 1988; Holzer 1991;
Nagy et al. 2004).
Several studies suggest that interactions among insulin, the InsR and the TRPV1expressed in PSNSs may contribute to physiological and pathophysiological processes including, for example, inflammatory changes of the skin (Gamse et al.
1987; Santicioli et al. 1987), the dura mater (Dux et al.
2007) and the exocrine and endocrine pancreas (Nathan et al.2001; Razavi et al.2006; Gram et al. 2007; Tsui et al.
2007). Immunohistochemical studies provided further support to this notion by showing a substantial co-localization of the TRPV1 and the InsR in rat and mouse PSNs of unidentified target innervation territories (Sugimoto et al.2002; Sántha and Nagy2006; Baiou et al.2007) and in rat pancreatic spinal and vagal PSNs (Lázár et al.2018).
The expression of the InsR in nociceptive sensory nerves of different organs and tissues may bear particular interest as regards the modulatory influence of the ubiquitous metabolic hormone insulin on a variety of tissue processes, in particular on pain and inflammation. Hence, exploring the localization of InsRs in PSNs serving somatic and visceral organs is of critical importance for the further understanding of the role of these particular afferent nerves under physiological and path- ological conditions. Therefore, the aim of the present study is to reveal the expression of InsRs in rat cutaneous, muscle, pancreatic and urinary bladder afferent neurons and its co- localization with the TRPV1.
Materials and methods
All experiments were approved by the Ethics Committee for Animal Care at the University of Szeged and were carried out in full accordance with the Directive 2010/63/EU of the European Parliament and of the Council on the Protection of Animals Used for Scientific Purposes and the guidelines of the Committee for Research. All efforts were made to minimise animal suffering. The number of experimental animals was kept as low as possible.
Retrograde labelling of somatic and visceral spinal primary sensory neurons
Adult male Wistar rats (n= 12), weighing 300–350 g, were anaesthetised with isoflurane (Forene™, AbbVie
Hungary Ltd., Budapest, Hungary). To identify cutaneous, muscle and visceral afferents, biotin-conjugated wheat germ agglutinin (bWGA; Sigma-Aldrich, Gillingham, UK, 1% in distilled water) was injected into the dorsal hind paw skin, the gastrocnemius muscle, the urinary bladder wall and the pancreas using a Hamilton microsyringe. For labelling of dorsal root ganglion (DRG) neurons serving the skin, 2 × 2-μl bWGA solution was injected into the dorsal hind paw skin. The gastroc- nemius muscle was exposed through an incision at the dorsal aspect of the calf and 2 × 2-μl bWGA solution was injected into the muscle. To label DRG neurons in- nervating the pancreas or the urinary bladder, these organs were exposed through a midline abdominal incision and 4 × 2-μl bWGA solution was injected into the parenchyma of the pancreas or the urinary bladder wall. Thereafter, the exposed area was thoroughly rinsed with saline and the wound was closed in layers. After recovery from anaes- thesia, the rats were returned to the animal house. Three days later, the rats were anaesthetised with an overdose of thiopental sodium (Insera Arzneimittel GmbH, Freiburg, Germany) (150 mg/kg i.p.) and perfused transcardially first with 100-ml saline, followed by 4% paraformalde- hyde in 0.1-M phosphate buffer (pH 7.4). Selected DRGs were removed, post-fixed for 2 h and stored in 0.1-M phosphate buffer (pH 7.4). Twenty-four hours be- fore sectioning, the specimens were transferred into a sucrose-buffer solution.
Serial frozen sections, 15μm in thickness, were cut on a cryostat and processed for immunohistochemistry. The L3–L5
(Gamse et al.1982; Swett et al.1991), L4–5(Peyronnard et al.
1986; Swett et al.1991), L3-S1(Jancsó and Maggi1987) and Th10–13(Won et al.1998; Takamido et al.2006; Lázár et al.
2018) DRGs were used to analyse the proportions of InsR- and TRPV1-immunoreactive neurons, which innervate the dorsal hind paw skin, the gastrocnemius muscle, the pancreas and the urinary bladder, respectively.
Immunohistochemistry
Sections were rinsed in phosphate buffered saline (PBS) for 2 × 10 min, incubated in PBS containing 10% horse serum and 0.1% Triton X-100 for 30 min and processed for staining with the indirect immunofluorescence technique using the fol- lowing antibodies: rabbit anti-InsRαsubunit antibody (Santa Cruz Biotechnologies, Dallas, Tex., USA; 1:500), guinea pig anti-TRPV1 antibody (Neuromics, Edina, Minn., USA;
1:1500). The specificity of these antisera were assessed in DRG sections by the lack of staining with the TRPV1 anti- body in specimens obtained from TRPV1−/−mice and by the failure of staining with the InsR antibody after preincubation with the immunising peptide supplied by the manufacturer
(Baiou et al. 2007). Donkey anti-rabbit IgG labelled with DL488 (1:500) and donkey anti-guinea pig IgG labelled with Cy3 (1:500) were used as secondary antibodies (all from Jackson Immunoresearch Laboratories, West Grove, Pa., USA). bWGA lectin was detected by using an extravidine- AMCA conjugate (Jackson Immunoresearch Laboratories).
All antibodies were diluted in phosphate buffered saline con- taining Triton X100 (0.3%). Sections were incubated in the presence of the primary antibodies overnight followed by a 2- h incubation with the secondary antibodies. Specimens were covered with Prolong Gold antifade mounting medium (Invitrogen, Carlsbad, CA, USA).
Analysis of identified somatic and visceral primary sensory neurons
Systemic random serial photomicrographs were taken with a Leica DMLB fluorescence microscope (Wetzlar, Germany) equipped with a Retiga 2000R digital camera (QImaging, Surrey, B.C., Canada). bWGA-positive neurons with visible nuclei were selected and analysis of retrogradely labelled neu- rons was performed by using ImageJ (NIH; Bethesda, Md., USA; IJ1.46r) image analysis software. Values are expressed as mean (standard deviation, SD). Statistical comparison of data was performed with the Fisher’s exact probability test.
Apvalue of≤0.05 was considered significant.
Results
Retrograde labelling of spinal PSNs innervating somatic and visceral organs using bWGA
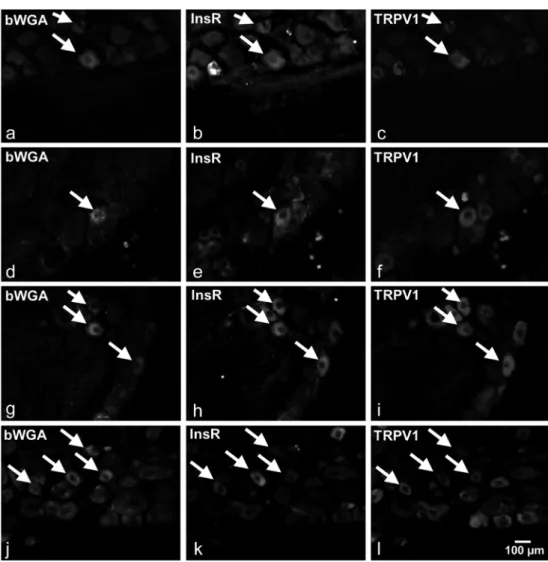
Injections of bWGA into the dorsal hind paw skin, the gas- trocnemius muscle, the pancreas and the urinary bladder wall resulted in the labelling of numerous neurons in the segmental DRGs. We identified and analysed 147 neurons innervating the dorsal hind paw skin (three animals), 138 neurons inner- vating the gastrocnemius muscle (three animals), 174 neurons innervating the pancreas (four animals) and 225 neurons in- nervating the urinary bladder (four animals). The size frequen- cy distribution of the labelled cells shows that bWGA identi- fied small-medium sized neurons. The mean (SD) cross- sectional area of the labelled cutaneous, muscle, pancreatic and urinary bladder spinal PSNs amounted to 311.1 (43.4), 345.8 (55.9), 387.2 (49.4) and 339.2 (54.8)μm2. We analysed the expression of the InsR and the TRPV1 in these retrograde- ly labelled neurons. Figure1illustrates target-identified retro- gradely labelled neurons examined in this study showing InsR- and TRPV1-immunoreactivity.
InsR expression in retrogradely labelled spinal PSNs
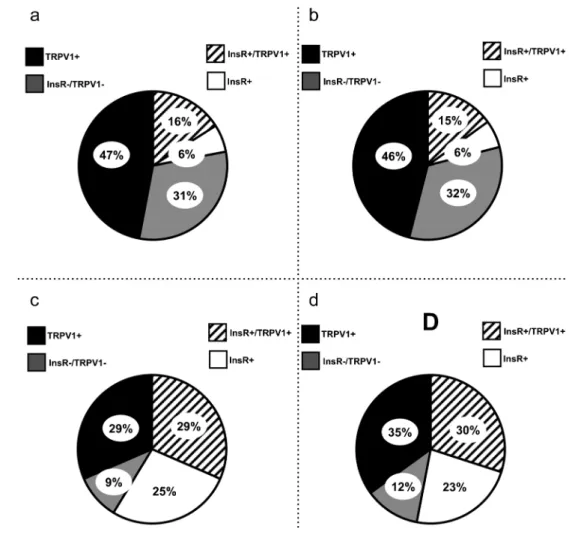
Our quantitative data indicate that 22.4 (2.8) % of cutaneous, 21.8 (1.9) % of muscle, 54.1 (2.9) % of pancreatic and 53.4 (3.1) % of urinary bladder afferent neurons exhibited InsR immunoreactivity (Fig.2). We found that there were no sig- nificant differences in the proportions of the InsR- immunoreactive neurons either between cutaneous and mus- cle afferents or between pancreatic and urinary bladder affer- ents. However, the statistical analysis revealed a highly signif- icant difference between proportions of InsR-immunoreactive somatic and visceral afferent neurons (p< 0.05). We also assessed the proportions of TRPV1-immunoreactiveneurons in the bWGA-labelled InsR-immunoreactive neuron popula- tion. In the DRGs, 72.7 (3.4), 73.3 (2.6), 54.3 (2.9) and 57.1 (3.6) % of the bWGA-labelled InsR-immunoreactive neurons innervating the dorsal hind paw skin, the gastrocnemius mus- cle, the pancreas and the urinary bladder showed TRPV1 im- munoreactivity, respectively (Table1). We found no signifi- cant differences in TRPV1 expression among the four popu- lations of neurons.
TRPV1 labelling of identified spinal PSNs
The large majority of identified PSNs expressed the TRPV1.
We found that 63.1 (3.4) % of cutaneous, 62.5 (2.7) % of muscle, 66.9 (2.5) % of pancreatic and 65.0 (1.8) % of urinary bladder bWGA-labelled afferents displayed TRPV1 immuno- reactivity (Fig.2). There were no significant differences in the TRPV1 immunoreactivity among the subpopulations. We analysed InsR immunoreactivity of the retrogradely labelled TRPV1-positive neurons as well. In the dorsal root ganglia, 25.8 (2.2), 25.5 (2.4), 43.9 (2.3) and 46.6 (1.9) % of the bWGA-labelled TRPV1-immunoreactive neurons innervating the dorsal hind paw skin, the gastrocnemius muscle, the pan- creas and the urinary bladder showed InsR-immunoreactivity, respectively (Table2). We found that there were no significant differences in the proportions of the InsR-immunoreactive neurons either between the cutaneous and the muscle afferents or between the pancreatic and urinary bladder afferents.
However, the differences between the cutaneous and pancre- atic, the cutaneous and bladder, the muscle and pancreatic and muscle and urinary bladder PSNs were significant (p< 0.05).
Co-localization of the InsR with the TRPV1
in retrogradely labelled somatic and visceral spinal PSNs
Through analysing the co-expression of the InsR with the TRPV1 in retrogradely labelled cells, we found that 16.56 (0.6) % of cutaneous, 15.33 (1.1) % of muscle, 29.4 (1.8) % of pancreatic and 30.34 (2.1) % of urinary bladder afferent neurons exhibited both InsR and TRPV1 immunoreactivity,
respectively (Fig.2). There were no significant differences either between the cutaneous and muscle or the pancreatic and urinary bladder neurons. However, there were signifi- cant differences in the proportion of the InsR- and TRPV- immunoreactive neurons between the somatic and visceral PSNs (p< 0.05).
Discussion
Recently, it has been revealed that InsR is expressed in rat and mouse PSNS of unidentified target innervation territories (Sugimoto et al.2002; Sántha and Nagy2006; Baiou et al.
2007) and in rat pancreatic spinal and vagal PSNs (Lázár et al.
2018). Our present quantitative immunohistochemical study shows that up to 20% of the retrogradely labelled spinal PSNs innervating the dorsal hind paw skin or the gastrocnemius muscle express the InsR. The proportions of InsR- immunoreactive neurons were significantly higher, amounting up to 50%, among PSNs innervating visceral organs.
Similarly to previous data, our present observations show that a relatively high proportion of PSNs expresses the InsR. In
addition, we found differential expression of the InsR in PSNs innervating somatic and visceral organs, respectively. In PC12 cells, which share characteristics with nociceptive PSNs, the InsR has recently been shown to form a complex with TrkA, the receptor activated by NGF (Geetha et al.2013). NGF is essential in the development of functional traits and neuro- chemical phenotypes of nociceptive PSNs (Snider and McMahon 1998). Moreover, the expression of the TRPV1 has been shown to be regulated by NGF (Michael and Priestley 1999). It is conceivable to suggest that through interacting with TrkA, the InsR contributes to the develop- ment, differentiation and functional characteristics of nocicep- tive PSNs.
In the present study, we also examined the TRPV1- immunoreactivity of the identified InsR-expressing PSNs.
Our quantitative data revealed that up to 50% of identified InsR-expressing spinal PSNs exhibit TRPV1 immunoreactiv- ity and in this respect, a significant difference between somatic and visceral neurons was not detected. Additionally, the anal- ysis of the retrogradely labelled TRPV1-immunoreactive neu- rons shows that almost half of the visceral but only a quarter of the somatic TRPV1-immunoreactive PSNs exhibit the InsR.
Fig. 1 Photomicrographs of biotin-conjugated wheat germ agglutinin (bWGA)-labelled primary sensory neurons innervating the dorsal hind paw skin (a,b,c), the gastrocnemius muscle (d,e,f), the pancreas (g,h, i) and the urinary bladder (j,k,l) showing immunoreactivities for the insulin receptor (InsR) and the transient receptor potential vanilloid type 1 receptor (TRPV1). The scale bar indicates 100μm and applies to all photomicrographs
Regarding the cell size and the TRPV1 expression pattern of the identified InsR-immunoreactive PSNs, it is highly likely that these neurons are nociceptive in function.
It has been demonstrated that systemic and local applica- tions of insulin induce vascular changes mediated by the re- lease of calcitonin gene-related peptide (CGRP) from capsaicin-sensitive PSNs (Salem and Dunbar2002). It has also been revealed that InsR by modulating intracellular sig- nalling systems may activate TRPV1-expressing PSNs (Heidenreich et al. 1990; Cesare et al.1999; Chuang et al.
2001; Sathianathan et al.2003). The present observations con- firm the co-localization of the InsR with TRPV1 in PSNs (Sántha and Nagy 2006; Baiou et al. 2007; Lázár et al.
2018) and extend previous findings by showing differential
co-localization of the InsR and the TRPV1 in somatic and visceral afferents. The relatively high co-expression rate of the InsR and the TRPV1 in spinal PSNs suggests a functional interaction between these receptors that may modify local tis- sue reactions effected by chemosensitive PSNs.
The expression of the InsR and the co-expression of the InsR with the TRPV1 may also bear important pathophysi- ological significance under conditions of unbalanced plas- ma insulin level. Insulin, through an action on TRPV1 sig- nalling, may elicit a release of pro-inflammatory neuropep- tides, such as CGRP (Salem and Dunbar2002). This inter- play may be important under hyperinsulinaemic conditions, such as the initial phase of type 2 diabetes mellitus.
Furthermore, insulin-induced activation of TRPV1 may be Fig. 2 Pie charts show the
percentage distribution of populations of retrogradely labelled primary sensory neurons (PSNs) innervating the dorsal hind paw skin (a), the gastrocnemius muscle (b), the pancreas (c) and the urinary bladder (d). Pie charts were constructed by analysing the chemical phenotypes of 147 (a), 128 (b), 174 (c) and 225 (d) retrogradely labelled cutaneous, muscle, pancreatic and urinary bladder PSNs
Table 1 Proportions of retrogradely labelled insulin receptor (InsR)-immunoreactive cutaneous, muscle, urinary bladder and pancreatic primary sensory neurons showing transient receptor potential vanilloid type 1 receptor (TRPV1) immunoreactivity
Target organ Number of InsR-
immunoreactive neurons
TRPV1+ neurons (%)
Dorsal hind paw skin 33 72.7 (3.4)
Gastrocnemius muscle 30 73.3 (2.6)
Pancreas 94 54.3 (3.6)
Urinary bladder 119 57.1 (2.9)
essential in keeping physiological functions of capsaicin- sensitive sensory nerves (Sathianathan et al.2003), which may be affected under hypoinsulinaemic conditions, such as in type 1 diabetes mellitus.
In recent years, many studies have focused on the role of TRPV1 in inflammatory processes of both the exocrine and the endocrine pancreas (Nathan et al.2001; Razavi et al.
2006; Wick et al.2006; Tsui et al. 2007; Iwasaki et al.
2013; Schwartz et al. 2013). More recently, it was also hypothesised that insulin, by modulating TRPV1 activa- tion, has a pivotal role in the initiation of pancreatitis (Razavi et al.2006; Tsui et al.2007; Lázár et al. 2018).
Our morphological findings demonstrating a high degree of co-localization of the InsR with the TRPV1 in neurons innervating the pancreas support this assumption. Similarly, diabetic cystopathy resulting in increased post-voiding re- sidual volumes and enhanced urinary bladder capacity is accompanied by decreased bladder sensation and contrac- tion secondary to damage of visceral afferent fibres in the urinary bladder wall (Kebapci et al.2007). This assumption is supported by observations of increased bladder capacity and disturbances in urinary bladder reflexes and micturition after neonatal capsaicin treatment (Jancsó and Maggi1987;
Maggi et al.1993), which produces a selective loss of noci- ceptive C-fibre PSNs (Jancsó et al.1977; Jancsó and Maggi 1987). Our findings, showing that ~ 50% of TRPV1- immunoreactive neurons co-express the InsRs, suggest a possible modulatory influence of insulin on urinary bladder sensory nerves.
In conclusion, the present study demonstrates that a higher proportion of visceral PSNs express the InsR as compared to somatic ones and that TRPV1-expressing visceral afferent neu- rons show a higher InsR expression rate than somatic PSNs.
Taking these morphological observations into consideration, it could be hypothesised that the TRPV1-dependent functions of visceral PSNs are more sensitive to changes in plasma insulin levels and these neurons may be more susceptible to an inter- play of the InsR and the TRPV1 under pathological conditions.
In addition, regarding the newly found co-localization of the InsR and the TRPV1in identified visceral afferent neurons and the functional interactions between these two receptors, a role of InsR-expressing visceral sensory nerves in the development of visceral inflammatory processes may emerge.
Funding information This work was supported in part by the Hungarian National Research, Development and Innovation Office (GINOP-2.3.2- 15.2016-00034). B.A. Lázár was supported by the UNKP-17-3 New National Excellence Program of the Ministry of Human Capacities and the TÁMOP 4.2.4.A/2-11/1-2012-0001.
Compliance with ethical standards
Conflict of interest The authors declare that they have no conflict of interest.
Ethical approval All procedures performed in studies involving animals were in accordance with the ethical standards of the institution or practice at which studies were conducted (Ethics Committee for Animal Care at the University of Szeged—Directive 2010/63/EU of the European Parliament and of the Council on the Protection of Animals Used for Scientific Purposes and the Guidelines of the Committee for Research.)
References
Baiou D, Sántha P, Avelino A, Charrua A, Bacskai T, Matesz K, Cruz F, Nagy I (2007) Neurochemical characterization of insulin receptor- expressing primary sensory neurons in wild-type and vanilloid type 1 transient receptor potential receptor knockout mice. J Comp Neurol 503:334–347
Barber AJ, Nakamura M, Wolpert EB, Reiter CE, Seigel GM, Antonetti DA, Gardner TW (2001) Insulin rescues retinal neu- rons from apoptosis by a phosphatidylinositol 3-kinase/Akt- mediated mechanism that reduces the activation of caspase-3.
J Biol Chem 276:32814–32821
Buck SH, Burks TF (1986) The neuropharmacology of capsaicin: review of some recent observations. Pharmacol Rev 38:179–226 Caterina MJ, Julius D (2001) The vanilloid receptor: a molecular gateway
to the pain pathway. Annu Rev Neurosci 24:487–517
Cesare P, Dekker LV, Sardini A, Parker PJ, McNaughton PA (1999) Specific involvement of PKC-epsilon in sensitization of the neuro- nal response to painful heat. Neuron 23:617–624
Chuang HH, Prescott ED, Kong H, Shields S, Jordt SE, Basbaum AI, Chao MV, Julius D (2001) Bradykinin and nerve growth factor release the capsaicin receptor from PtdIns(4,5)P2-mediated inhibi- tion. Nature 411:957–962
Dux M, Rosta J, Pintér S, Sántha P, Jancsó G (2007) Loss of capsaicin- induced meningeal neurogenic sensory vasodilatation in diabetic rats. Neuroscience 150:194–201
Fernyhough P, Willars GB, Lindsay RM, Tomlinson DR (1993) Insulin and insulin-like growth factor I enhance regeneration in cultured adult rat sensory neurones. Brain Res 607:117–124
Figlewicz DP (2016) Expression of receptors for insulin and leptin in the ventral tegmental area/substantia nigra (VTA/SN) of the rat: histor- ical perspective. Brain Res 1645:68–70
Table 2 Proportions of retrogradely labelled transient receptor potential vanilloid type 1 receptor (TRPV1)-
immunoreactive cutaneous, muscle, urinary bladder and pancreatic primary sensory neurons showing insulin receptor (InsR) immunoreactivity
Target organ Number of TRPV1-
immunoreactive neurons
InsR+ neurons (%)
Dorsal hind paw skin 93 25.8 (2.2.)
Gastrocnemius muscle 86 25.5 (2.4)
Pancreas 116 43.9 (1.9)
Urinary bladder 146 46.6 (2.3)
Gamse R, Petsche U, Lembeck F, Jancsó G (1982) Capsaicin applied to peripheral nerve inhibits axoplasmic transport of substance P and somatostatin. Brain Res 239:447–462
Gamse R, Posch M, Saria A, Jancsó G (1987) Several mediators appear to interact in neurogenic inflammation. Acta Physiol Hung 69:343– 354
Geetha T, Rege SD, Mathews SE, Meakin SO, White MF, Babu JR (2013) Nerve growth factor receptor TrkA, a new receptor in insulin signaling pathway in PC12 cells. J Biol Chem 288:23807–23813 Gram DX, Ahrén B, Nagy I, Olsen UB, Brand CL, Sundler F, Tabanera
R, Svendsen O, Carr RD, Sántha P, Wierup N, Hansen AJ (2007) Capsaicin-sensitive sensory fibers in the islets of Langerhans con- tribute to defective insulin secretion in Zucker diabetic rat, an animal model for some aspects of human type 2 diabetes. Eur J Neurosci 25:
213–223
Heidenreich KA, Toledo SP, Brunton LL, Watson MJ, Daniel-Issakani S, Strulovici B (1990) Insulin stimulates the activity of a novel protein kinase C, PKC-epsilon, in cultured fetal chick neurons. J Biol Chem 265:15076–15082
Holzer P (1991) Capsaicin: cellular targets, mechanisms of action, and selectivity for thin sensory neurons. Pharmacol Rev 43:143–201 Iwasaki Y, Shimomura K, Kohno D, Dezaki K, Ayush EA, Nakabayashi
H, Kubota N, Kadowaki T, Kakei M, Nakata M, Yada T (2013) Insulin activates vagal afferent neurons including those innervating pancreas via insulin cascade and Ca2+influx: its dysfunction in IRS2-KO mice with hyperphagic obesity. PLoS One 8:e67198 Jancsó G, Maggi CA (1987) Distribution of capsaicin-sensitive urinary
bladder afferents in the rat spinal cord. Brain Res 418:371–376 Jancsó N, Jancsó-Gábor A, Szolcsányi J (1967) Direct evidence for neu-
rogenic inflammation and its prevention by denervation and by pre- treatment with capsaicin. Br J Pharmacol Chemother 31:138–151 Jancsó G, Kiraly E, Jancsó-Gábor A (1977) Pharmacologically induced
selective degeneration of chemosensitive primary sensory neurones.
Nature 270:741–743
Kebapci N, Yenilmez A, Efe B, Entok E, Demirustu C (2007) Bladder dysfunction in type 2 diabetic patients. Neurourol Urodyn 26:
814–819
Lázár BA, Jancsó G, Oszlács O, Nagy I, Sántha P (2018) The insulin receptor is colocalized with the TRPV1 nociceptive ion channel and neuropeptides in pancreatic spinal and vagal primary sensory neu- rons. Pancreas 47:110–115
Maggi CA, Meli A (1988) The sensory-efferent function of capsaicin- sensitive sensory neurons. Gen Pharmacol 19:1–43
Maggi CA, Lecci A, Santicioli P, Del Bianco E, Giuliani S (1993) Cyclophosphamide-induced cystitis in rats: involvement of capsaicin-sensitive primary afferents. Agents Actions 38 Spec No:
C28–C30
Michael GJ, Priestley JV (1999) Differential expression of the mRNA for the vanilloid receptor subtype 1 in cells of the adult rat dorsal root and nodose ganglia and its downregulation by axotomy. J Neurosci 19:1844–1854
Nagy I, Sántha P, Jancsó G, Urbán L (2004) The role of the vanilloid (capsaicin) receptor (TRPV1) in physiology and pathology. Eur J Pharmacol 500:351–369
Nathan JD, AA P, McVey DC, Thomas JE, Prpic V, Vigna SR, RA L (2001) Capsaicin vanilloid receptor-1 mediates substance P release in experimental pancreatitis. Am J Physiol Gastrointest Liver Physiol 281:G1322–G1328
Peyronnard JM, Charron LF, Lavoie J, Messier JP (1986) Motor, sympa- thetic and sensory innervation of rat skeletal muscles. Brain Res 373:288–302
Razavi R, Chan Y, Afifiyan FN, Liu XJ, Wan X, Yantha J, Tsui H, Tang L, Tsai S, Santamaria P, Driver JP, Serreze D, Salter MW, Dosch HM (2006) TRPV1+sensory neurons controlβcell stress and islet in- flammation in autoimmune diabetes. Cell 127:1123–1135
Recio-Pinto E, Ishii DN (1988) Insulin and related growth factors: effects on the nervous system and mechanism for neurite growth and re- generation. Neurochem Int 12:397–414
Recio-Pinto E, Lang FF, Ishii DN (1984) Insulin and insulin-like growth factor II permit nerve growth factor binding and the neurite forma- tion response in cultured human neuroblastoma cells. Proc Natl Acad Sci U S A 81:2562–2566
Recio-Pinto E, Rechler MM, Ishii DN (1986) Effects of insulin, insulin- like growth factor-II, and nerve growth factor on neurite formation and survival in cultured sympathetic and sensory neurons. J Neurosci 6:1211–1219
Salem N, Dunbar JC (2002) The insulin-mediated vascular and blood pressure responses are suppressed in CGRP-deficient normal and diabetic rats. Diabetes Metab Res Rev 18:238–244
Sántha P, Nagy I (2006) Insulin and insulin-like growth factor-I induce thermo-sensitive ionic currents in capsaicin-sensitive primary sen- sory neuron. FENS Forum Abstr 3
Santicioli P, Gamse R, Maggi CA, Meli A (1987) Cystometric changes in the early phase of streptozotocin-induced diabetes in rats: evidence for sensory changes not correlated to diabetic neuropathy. Naunyn Schmiedeberg’s Arch Pharmacol 335:580–587
Sathianathan V, Avelino A, Charrua A, Sántha P, Matesz K, Cruz F, Nagy I (2003) Insulin induces cobalt uptake in a subpopulation of rat cultured primary sensory neurons. Eur J Neurosci 18:2477–2486 Schwartz ES, La J-H, Scheff NN, Davis BM, Albers KM, Gebhart GF
(2013) TRPV1 and TRPA1 antagonists prevent the transition of acute to chronic inflammation and pain in chronic pancreatitis. J Neurosci 33:5603–5611
Singh B, Xu Y, McLaughlin T, Singh V, Martinez JA, Krishnan A, Zochodne DW (2012) Resistance to trophic neurite outgrowth of sensory neurons exposed to insulin. J Neurochem 121:263–276 Snider WD, McMahon SB (1998) Tackling pain at the source: new ideas
about nociceptors. Neuron 20:629–632
Stella SLJ, Bryson EJ, Thoreson WB (2001) Insulin inhibits voltage- dependent calcium influx into rod photoreceptors. Neuroreport 12:947–951
Sugimoto K, Murakawa Y, Zhang W, Xu G, Sima AA (2000) Insulin receptor in rat peripheral nerve: its localization and alternatively spliced isoforms. Diabetes Metab Res Rev 16:354–363
Sugimoto K, Murakawa Y, Sima AAF (2002) Expression and localization of insulin receptor in rat dorsal root ganglion and spinal cord. J Peripher Nerv Syst 7:44–53
Swett JE, Torigoe Y, Elie VR, Bourassa CM, Miller PG (1991) Sensory neurons of the rat sciatic nerve. Exp Neurol 114:82–103
Takamido S, Kataoka Y, Tanano A, Cui Y, Ikeura T, Shimatani M, Kubota Y, Okazaki K, Yamada H (2006) Intrapancreatic axonal hyperbranching of dorsal root ganglia neurons in chronic pancre- atitis model rats and its relation to pancreatic pain. Pancreas 33:
268–279
Tsui H, Razavi R, Chan Y, Yantha J, Dosch HM (2007)BSensing^auto- immunity in type 1 diabetes. Trends Mol Med 13:405–413 Wan Q, Xiong ZG, Man HY, Ackerley CA, Braunton J, Lu WY, Becker
LE, MacDonald JF, Wang YT (1997) Recruitment of functional GABA(A) receptors to postsynaptic domains by insulin. Nature 388:686–690
Wick EC, Hoge SG, Grahn SW, Kim E, Divino LA, Grady EF, Bunnett NW, Kirkwood KS (2006) Transient receptor potential vanilloid 1, calcitonin gene-related peptide, and substance P mediate nociception in acute pancreatitis. Am J Physiol Gastrointest Liver Physiol 290:G959–G969
Won MH, Park HS, Jeong YG, Park HJ (1998) Afferent innervation of the rat pancreas: retrograde tracing and immunohistochemistry in the dorsal root ganglia. Pancreas 16:80–87