A NAC TRANSZKRIPCIÓS FAKTOR SZEREPE A LISZTHARMAT ELLENI VÉDEKEZÉSBEN
T
ÓTHZ
SÓFIA1, K
OVÁCSL
ÁSZLÓ2, K
ISSE
RZSÉBET11
Szent István Egyetem, Genetika és Biotechnológiai Intézet, Gödöllő
2
Missouri State University, Biology Department, Springfield
Az obligát biotróf patogének összetett védekezési reakciót váltanak ki a fogékony gazdanövényben. Korábbi kutatások kimutatták, hogy lisztharmat gomba fertőzés hatására szalicilsav halmozódik fel a szőlő (Vitis vinifera cv. Cabernet Sauvignon) levél szövetei- ben, ami arra enged következtetni, hogy a szalicilsav része a növény védekezési rendszeré- nek. Microarray módszerrel összehasonlítottuk 3000 gén expressziójának lisztharmat és szalicilsav hatására történő változását, és azt tapasztaltuk, hogy a legtöbb gén hasonlóan reagált mindkét kezelésre. Néhány stressz-függő gén expressziója, köztük a NAC transz- kripciós faktort kódoló géné is azonban csak a patogén jelenlétében emelkedett, szalicilsav kezelésre nem reagált. Ezen eredmények alapján feltételezhető, hogy a szalicilsav nem szükséges, illetve szükséges, de önmagában nem elegendő a NAC transzkripciós faktor gén indukálásához. Hogy megvizsgáljuk a NAC működését a gén promóter régióját izoláltuk, amelyet összekapcsoltunk a GUS riporter génnel és a konstrukcióval egy vad és két mutáns Arabidopsis típust transzformáltunk. A mutáns vonalakban a szalicilsav-jelátviteli rendszer gátolva van. Mindhárom típusú transzgénikus vonal GUS alapexpressziót mutatott, viszont Golovinomyces orontii és Oidium neolycopersici fertőzést követően a GUS expressziója szalicilsav szignál-átvitelben blokkolt növényekben is megnövekedett a kontrollhoz ké- pest, ami azt jelenti, hogy a NAC transzkripciós faktor lisztharmat hatására bekövetkező expressziós változása független a szalicilsav-jelátviteli rendszertől.
Kulcsszavak: szalicilsav, patogén, szignál transzdukció, szőlő
ROLE OF NAC TRANSCRIPTION FACTOR IN THE DEFENSE SYSTEM AGAINST POWDERY MILDEW
Z
S. T
ÓTH1, L. K
OVÁ
CS2, E. K
ISS11
Szent Istvan University, Istitute of Genetics and Biotechnology, Godollo
2
Missouri State University, Biology Department, Springfield
The obligate biotroph pathogens trigger a complex defense reaction in susceptible host plants. Earlier studies showed that powdery mildew infection induces salicylic acid accumulation in grape (Vitis vinifera cv. Cabernet Sauvignon) leaf tissue, which suggests that salicylic acid may play a role in the plant defense system. We compared the expres- sion changes of 3000 genes triggered by powdery mildew and salicylic acid using microar- ray, and observed that most of the genes reacted in the same way to both treatments. How- ever the expression of some stress-dependent genes, such as NAC transcription factor gene increased only in the presence of powdery mildew, and not on the effect of salicylic acid treatment. Based on the results we can presume that salicylic acid is not required, or re- quired but insufficient by itself to induce the NAC transcription factor gene expression. To analyze the NAC regulation we isolated the promoter region of the gene, fused to GUS re- porter gene and transferred into a wild and two mutant Arabidopsis types. The mutants are deficient in salicylic acid-mediated signal transduction. All three transgenic lines showed a
basic expression of GUS, but following the Golovinomyces orontii and Oidium neoly- copersici infection a high increase of GUS expression could be observed even in the sali- cylic acid deficient plants compared to the control, which suggests that powdery mildew- mediated regulation of NAC transcription factor is independent of salicylic acid signal transduction.
Keywords: salicylic acid, pathogen, signal transduction, grape
Bevezetés
A lisztharmatgombák obligát biotróf patogének, amelyek a korókozók kö- zött a legnagyobb károkat okozzák a szőlőművelésben. Habár sok kutatási terü- let foglalkozik a növény-patogén közti kapcsolat megismerésével, még mindig keveset tudunk a védekezési mechanizmusról, ami magába foglalhatja gének, jelmolekulák, jelátviteli rendszerek együttes működését.
Előzetes kutatások kimutatták, hogy lisztharmat fertőzés hatására szalicil- sav halmozódik fel a fogékony növényben, amelyről már bizonyították, hogy része lehet a védekezési mechanizmusnak (Fung et al., 2008). Egy korábbi expressziós kísérletünkben összehasonlítottuk a lisztharmat és szalicilsav kezelés hatására bekövetkező változásokat a gének működésében. Az eredmények alap- ján azt tapasztaltuk, hogy a legtöbb gén hasonló módon és mértékben reagált mindkét kezelésre, azonosítottunk azonban olyan stressz-függő géneket amelyek expressziója egyedül a lisztharmat fertőzést követően emelkedett. Ebben a cso- portban sztilbén szintáz gének, transzkripciós faktorok génjei szerepelnek, köz- tük a WRKY71 és NAC gének, amelyeknek a kórokozók elleni védekezésben betöltött szerepét már mind bizonyították. A Cabernet Sauvignon szőlőben álta- lunk megfigyelt NAC gén Arabidopsis-ban azonosított ortológjáról (ANAC042) tudjuk, hogy fitoalexinek termeléséért felelős, amelyek közismerten a növény patogének ellen bevetett „vegyszerei”. Az ANAC042 génről már azt is bizonyí- tották, hogy a patogén hasonló expressziót váltott ki a szalicilsav-mentes mutáns növényben, mint a kontrollban (Saga et al. 2012). Az eredményekből arra kö- vetkeztettünk, hogy a szalicilsav nem szükséges, vagy szükséges, de önmagában nem elegendő a NAC transzkripciós faktor indukciójához. A szalicilsavtól való függés, vagy függetlenség meghatározásához a NAC gén promóterét izoláltuk Cabernet Sauvignon fogékony fajtából, összekapcsoltuk GUS riportergénnel, és az így létrehozott vektorral egy vad és két szalicilsav-jelátvitelben gátolt mutáns Arabidopsis-t transzformáltunk.
Anyag és módszer
A NAC gén működését a gén promóterének izolálásához a Grape Genome Browser- Genoscope adatbázis alapján a Vv3989 F 5’-CACCTCAATCACACTCAAAAACCA-3’ és a Vv_24 R 5’-AGTGCTAGTCTTCTCCACCTCCAT-3’ primereket terveztük. Ezzel a primerpárral egy 3981 bp hosszú DNS szakaszt szaporítottunk fel a gén ATG start kodonja előtti régióból. A promóter szekvenciájának elemzését a PLACE adatbázissal végeztük (Higo et al. 1999).
A promóter szakaszt Gateway® technológiával bar szelekciós markert és GUS riportergént tartalmazó pGWB633 (Nakamura et al. 2010) típusú bináris vektorba klónoztuk. A vektort Agrobacterium tumefaciens GV3101 pMP90-es törzsébe juttatva egy Wassilewskija (WS) ökotípusú vad és két szalicilsav-jelátvitelben gátolt (WS-nim1-1 szalicilsav-jelátvitel blokkolt mutáns; WS-nahG szalicilát dehidrogenáz enzimet tartalmazó transzgénikus) (Ryals et al. 1997;
Gaffney et al. 1993) Arabidopsis vonalat transzformáltunk, melyhez a „floral dip” (Clough és Bent 1998) módszert alkalmaztuk. A virágokból fejlődő következő generációhoz a magokat Petri csé- székben csíráztattuk és a növényeket hosszú nappalos hideg fehér fény és 22oC-os hőmérséklet mellett neveltük. A 2 hetes növényeket glüfozinát tartalmú Finale vegyszerrel permeteztük és a túlélőket kiválogattuk. Az egymást követő generációkból kiválogattuk mindhárom típusú növény- ből 3-3 olyan vonalat, melyek egy kópiában tartalmazzák a transzgént. A NAC promóter gomba- fertőzés hatására indukálódó működését 4 hetes egy kópiás növényeken teszteltük. A fertőzéshez Oidium neolycopersici (paradicsomlisztharmat) és Golovinomyces orontii (dohánylisztharmat) gombákat használtunk. A fertőzést követően 11 nappal a GUS expressziót hisztokémiai festéssel (Jefferson et al. 1987) állapítottuk meg. A gombát gyapotkékkel festettük. A mikroszkópus vizsgá- latokhoz az Olympus Leica Leitz DMRB típusú fény- és WILD M3Z sztereomikroszkópokat használtuk.
Eredmények
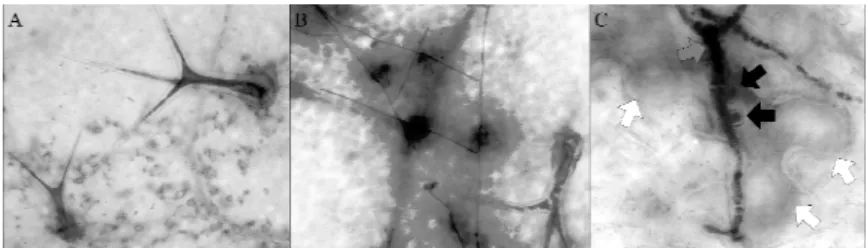
A kiválogatott vonalakat előzetes vizsgálata alapján megállapítottuk, hogy a növények környezeti hatásoktól függetlenül GUS alap-expressziót mutatnak mindhárom típusú növényben. Ez a konstitutív működés leginkább a levélsző- rökben, vaszkuláris szövetekben, merisztémákban és a fejlődő becőkben volt megfigyelhető. Tizenegy napos fertőzést követően azt tapasztaltuk, hogy mind- három típusú növény azokon a területeken növeli meg a GUS expresszióját, ahol a levél felülete érintkezik a gombával. Mikroszkópos vizsgálatok során azt fi- gyeltük meg, hogy azokban a sejtekben, amelyekben a gomba hausztóriumot fejlesztett, nagyobb a GUS kifejeződése, mint az érintetlenekben (1. ábra).
1. ábra Ál-fertőzött (A) és Golovinomyces orontii-val fertőzött (B) transzgénikus növény. (C) A NAC promóter kifejeződése az érintett sejtben
(Oidium neolycopersici fertőzés) Fekete nyíl: a gomba által fejlesztett hausztoriumok; szürke nyíl: gomba hifa; fehér nyíl: Gomba által megtámadott
sejt, amely a β-glükuronidázt expresszálja.
A jelenség mindkét gombafaj fertőzését követően és mindhárom típusú
transzgénikus növény esetében azonosítható volt. A NAC promóter szekvenciáját
elemezve a következő stressz- és hormon-függő motívumokat azonosítottuk:
szalicilsav-, etilén, jazmonsav- és elicitor-érzékeny elemek (1. táblázat), ame- lyek mindegyike felelős lehet a NAC gén transzkripciójának indukálásáért a lisztharmatfertőzésben.
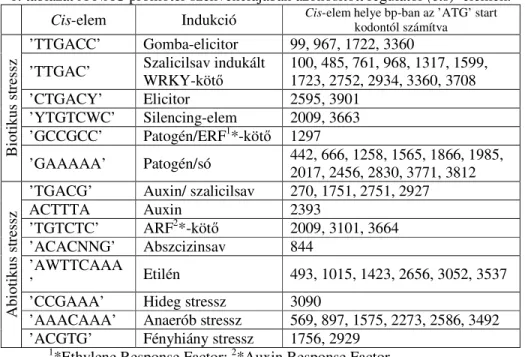
1. táblázat A NAC promóter szekvenciájában azonosított regulátor (cis)–elemek.
Cis-elem Indukció
Cis-elem helye bp-ban az ’ATG’ start kodontól számítva’TTGACC’ Gomba-elicitor 99, 967, 1722, 3360
’TTGAC’ Szalicilsav indukált WRKY-kötő
100, 485, 761, 968, 1317, 1599, 1723, 2752, 2934, 3360, 3708
’CTGACY’ Elicitor 2595, 3901
’YTGTCWC’ Silencing-elem 2009, 3663
’GCCGCC’ Patogén/ERF
1*-kötő 1297
B io ti ku s st re ss z
’GAAAAA’ Patogén/só 442, 666, 1258, 1565, 1866, 1985, 2017, 2456, 2830, 3771, 3812
’TGACG’ Auxin/ szalicilsav 270, 1751, 2751, 2927
ACTTTA Auxin 2393
’TGTCTC’ ARF
2*-kötő 2009, 3101, 3664
’ACACNNG’ Abszcizinsav 844
’AWTTCAAA
’ Etilén 493, 1015, 1423, 2656, 3052, 3537
’CCGAAA’ Hideg stressz 3090
’AAACAAA’ Anaerób stressz 569, 897, 1575, 2273, 2586, 3492
A bi ot ik us s tr es sz
’ACGTG’ Fényhiány stressz 1756, 2929
1
*Ethylene Response Factor;
2*Auxin Response Factor Következtetések
Kutatásunk célja, hogy megismerjük a NAC transzkripciós faktor liszt-
harmat fertőzés során betöltött szerepét, működésének hátterét. A NAC-
promóter::GUS riporter gén konstrukcióval transzformált Arabidopsis növények
vizsgálatakor megfigyeltük, hogy a NAC promóter környezeti hatásoktól függet-
lenül alap-expressziót mutat, ami leginkább a növények levélszőrében,
vaszkuláris szöveteiben, merisztémákban és fejlődő becőkben volt látható. A
jelenség alapján feltételezhető, hogy a NAC traszkripciós faktor lisztharmat elle-
ni védekezésben betöltött szerepe mellett más, talán alap/metabolikus funkciókat
is ellát a növényben. Mivel mindhárom típusú transzgénikus növényben azonosí-
tottuk a GUS alapműködését, ezért valószínűsíthető, hogy e funkciók ellátásához
nem szükséges a szalicilsav jelenléte, sem a szalicilsavas jelátviteli rendszer
működése. A NAC gén lisztharmat elleni védekezésben betöltött szerepét viszont
a fertőzést követő megemelkedett GUS expresszió bizonyította. A gomba hatásá-
ra bekövetkező indukció mindhárom típusú növényben azonosítható volt, tehát a
NAC gén megnyilvánulásának a lisztharmattól függő megnövekedése is függet-
len a szalicilsavtól, ami összhangban van az Arabidopsis-ban található ANAC042 gén patogén által kiváltott reakciójával szalicilsav-mentes növényben (Saga et al. 2012). Ezek az eredmények azt bizonyítják, hogy nem a szalicilsav a felelős jelmolekula a lisztharmat és NAC gén közötti kapcsolatban, viszont a promóter szekvenciáját elemezve elicitor és etilén-érzékeny elemeket azonosítottunk, amelyek szintén jelmolekulaként működhetnek a biotikus stresszekben. A NAC génnek a fertőzés helyén megfigyelhető specifikus megnyilvánulásából és egy korábbi kutatás eredményéből (Chandran et al. 2010), amely kimutatta a hausztórium körül fokozottan expresszáló, a védekezésben szerepet játszó géne- ket, arra következtethetünk, hogy a NAC transzkripciós faktort a növény termeli védekezés céljából és nem a gomba irányítja működését.
Köszönetnyilvánítás
A kutatásokat az OTKA 77867, a TÁMOP-4.2.2.B-10/1 „A tehetséggondozás és kutató- képzés komplex rendszerének fejlesztése a Szent István Egyetemen” c. pályázat és a Kutató Kari Kiválósági Támogatás-17586-4/2013/TUDPOL támogatta.
Irodalom
Chandran, D., Inada, N., Hather, G., Kleindt, C. K., Wildermuth, M. C. (2010) Laser microdissection of Arabidopsis cells at the powdery mildew infection site reveals site- specific processes and regulators. Proceeding of the National Academy of Sciences 107, 460-465
Clough, S. J., Bent, A. F. (1998) Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. The Plant Journal 16, 735-743.
Fung, R. W. M., Gonzalo, M., Fekete, C., Kovács, L. G., He, Y., Marsh, E., MyIntyre, L. M., Schachzman, D. P., Qiu, W. (2008) Powdery mildew induces defense-oriented reprogramming of the transciptome in a susceptible but not in a resistant grapevine. Plant Physiology 146, 236-249.
Gaffney, T., Friedrich, L., Vernooij, B., Negrotto, D., Nye, G., Uknes, S., Ward, E., Kessmann, H., Ryals, J. (1993) Requirement of salicylic acid for the induction of systemic acquired resistance. Science 261, 754-756.
Higo, K., Ugava, Y., Iwamoto, M., Korenaga, T. (1999) Plant cis-acting regulatory DNA elements (PLACE) database: 1999. Nucleic Acids Research 27, 297-300.
Jefferson, R. A., Kavanagh, T. A., Bevan, M. W. (1987) GUS fusions: beta-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. The EMBO Journal 6, 3901- 3907.
Nakamura, S., Mano, S., Tanaka, Y., Ohnishi, M., Nakamori, C. (2010) Gateway binary vectors with the bialaphos resistance gene, bar, as a selection marker for plant transformation.
Bioscience, Biotechnology, and Biochemistry 74, 100184-100181-100185.
Ryals, J., Weymann, K., Lawton, K., Friedrich, L., Ellis, D., Steiner, H. Y., Johnson, J., Delaney, T. P., Jesse, T., Vos, P., Uknes, S. (1997) The Arabidopsis NIM1 protein shows homology to the mammalian transcription factor inhibitor IκB. The Plant Cell Online 9, 425-439.
Saga, H., Ogawa, T., Kai, K., Suzuki, H., Ogata, Y., Sakurai, N., Shibata, D., Ohta, D. (2012) Identification and characterization of ANAC042, a transcription factor family gene involved in the regulation of camalexin biosynthesis in Arabidopsis. Molecular Plant- Microbe Interactions 25, 684-696.