GEORGIKON MEZÕGAZDASÁGTUDOMÁNYI KAR NÖVÉNY- ÉS KÖRNYEZETTUDOMÁNYI INTÉZET
KERTÉSZETI TANSZÉK
NÖVÉNYTERMESZTÉSI ÉS KERTÉSZETI TUDOMÁNYOK DOKTORI ISKOLA
Doktori Iskolavezetõ:
DR. GÁBORJÁNYI RICHÁRD egyetemi tanár, az MTA doktora
Témavezetõ:
DR. KOCSIS LÁSZLÓ
egyetemi docens, a mg. tud. kandidátusa
A MAGYARORSZÁGON ELÕFORDULÓ SZÕLÕ-GYÖKÉRTETÛ POPULÁCIÓK VIZSGÁLATÁNAK ÚJABB EREDMÉNYEI
Készítette:
Lipicsné Tóth Hajnalka Ilona PhD hallgató
KESZTHELY 2006
A Magyarországon elõforduló szõlõ-gyökértetû populációk vizsgálatának újabb eredményei
Értekezés doktori (PhD) fokozat elnyerése érdekében Írta:
Lipicsné Tóth Hajnalka Ilona
**Készült a Veszprémi Egyetem Növénytermesztési és Kertészeti Doktori Iskolája keretében
Témavezetõ: Dr. Kocsis László egyetemi docens Elfogadásra javaslom (igen / nem)
(aláírás)**
A jelölt a doktori szigorlaton …... % -ot ért el,
Veszprém/ Keszthely, …...
a Szigorlati Bizottság elnöke Az értekezést bírálóként elfogadásra javaslom:
Bíráló neve:…... igen /nem
……….
(aláírás) Bíráló neve: …... …... igen /nem
……….
(aláírás)
***Bíráló neve: …... …...) igen /nem
……….
(aláírás) A jelölt az értekezés nyilvános vitáján…...% - ot ért el
Veszprém/Keszthely, ……….
a Bíráló Bizottság elnöke A doktori (PhD) oklevél minõsítése…...
………
Az EDT elnöke
Megjegyzés: a * közötti részt az egyéni felkészülõk, a ** közötti részt a szervezett képzésben résztvevõk használják, *** esetleges
oldal
1. Kivonatok 5
1. 1. Magyar nyelvû kivonat 5
1. 2. Angol nyelvû kivonat 7
1. 3. Német nyelvû kivonat 8
2. Bevezetés 9
3. Irodalmi áttekintés 11
3. 1. Szõlõfajok kialakulása és jellemzõi 11
3. 1. 1. Szõlõ-gyökértetû és a szõlõ növény kapcsolata 12 3. 1. 2. A szõlõfajok filoxéra rezisztenciája 13
3. 2. A szõlõ-gyökértetû 16
3. 2. 1. A faj bemutatása 16
3. 2. 2. A kártevõ fejlõdési alakjai 17
3. 2. 3. A filoxéra életmódja 19
3. 2. 4. A szõlõ-gyökértetû változatai 25
3. 2. 5. Filoxéra kártétele 26
3. 3. Filoxéra Magyarországon 30
3. 3. 1. Borászatunk helyzete a kártevõ megjelenése elõtt 30 3. 3. 2. A szõlõ-gyökértetû elterjedése Magyarországon 30
3. 3. 3. Filoxéravész következményei 33
3. 4. Szõlõ-gyökértetû kutatás a világban 35
3. 5. A hazai filoxéra kutatás 47
4. Vizsgálatok anyaga és módszere 53
4. 1. Laboratóriumi gyökérteszt anyaga és módszere 53 4. 1. 1. Laboratóriumi gyökérteszt anyaga 53 4. 1. 2. Laboratóriumi gyökérteszt módszere 56 4. 2. Tenyészedényes kísérlet anyaga és módszere 57 4. 2. 1. Tenyészedényes kísérlet anyaga 57 4. 2. 2. Tenyészedényes kísérlet módszere 58
4. 3. Genetikai vizsgálat anyaga és módszere 60
4. 3. 1. Genetikai vizsgálat anyaga 60
4. 3. 2. Genetikai vizsgálat módszere 60
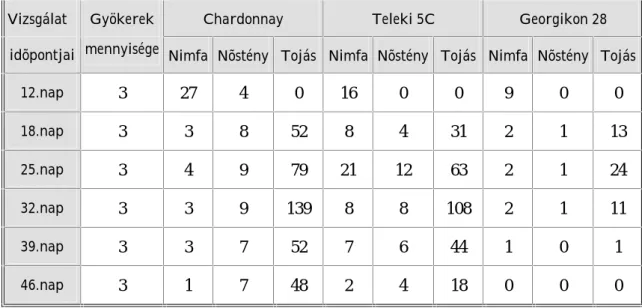
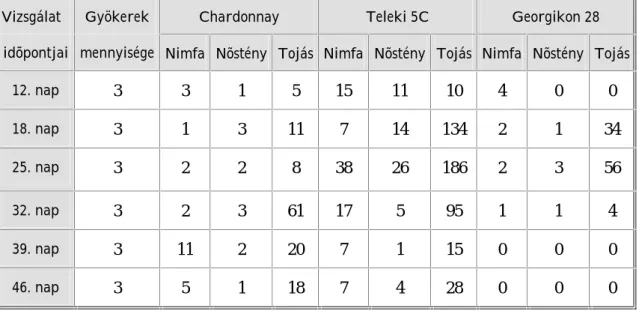
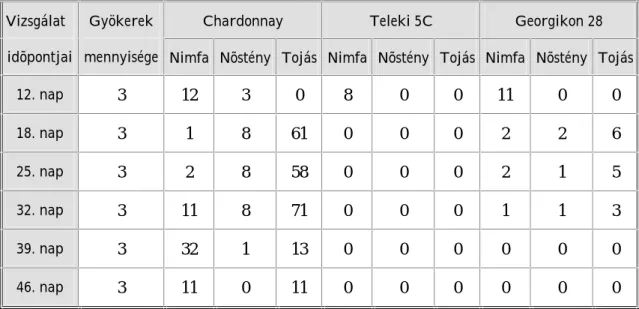
5. Vizsgálatok eredményei és eredményeinek értékelése 63 5. 1. Laboratóriumi gyökérteszt eredményei és eredményeinek értékelése 63 5. 1. 1. Laboratóriumi gyökérteszt eredményei 63 5. 1. 2. Laboratóriumi gyökérteszt eredményeinek értékelése 74 5. 2. Tenyészedényes kísérlet eredményei és eredményeinek értékelése 86 5. 2. 1. Tenyészedényes kísérlet eredményei 86 5. 2. 2. Tenyészedényes kísérlet eredményeinek értékelése 90 5. 3. Genetikai vizsgálat eredményei és eredményeinek értékelése 98 5. 3. 1. Genetikai vizsgálat eredményei 98 5. 3. 2. Genetikai vizsgálat eredményeinek értékelése 101
6. Következtetések, javaslatok 104
7. Összefoglalás 110
8. Köszönetnyilvánítás 112
9. Szakirodalom jegyzéke 113
10. Az értekezés tézisei 127
10. 1. Az értekezés tézisei magyar nyelven 127
10. 2. Az értekezés tézisei angol nyelven 129
1. KIVONATOK
1. 1. MAGYAR NYELVÛ KIVONAT
A Magyarországon elõforduló szõlõ-gyökértetû populációk vizsgálatának újabb eredményei
A szõlõ-gyökértetû (=filoxéra, Daktulosphaira vitifoliae FITCH) az 1800-as évek második felében került Európába és pár évtized alatt kipusztította a szõlõültetvényeket. Az 1960-1970-es években Franciaországban, az 1990-es évek elején Németországban számos alanyon újra észleltek filoxéra fertõzésre visszavezethetõ tõkepusztulást. Ez felveti annak lehetõségét, hogy a szõlõtermesztésben általánossá vált oltványhasználat ellenére a rovar ismét károsít.
A szõlõalanyok tartós használata, a változó termesztéstechnológia és a környezeti adottságok megváltozatása egyaránt elõsegítheti a kártevõ tápnövényhez való adaptálódását. A rovar kultúrnövényhez való alkalmazkodása és gyors szaporodása miatt újra indokolttá vált a kártevõ életmódjának és a szõlõalanyok filoxéra rezisztenciájának alapos vizsgálata. Ezek figyelembe vételével célul tûztük ki a szõlõalanyfajták filoxéra rezisztenciájának vizsgálatát, Magyarország borvidékeirõl begyûjtött szõlõ-gyökértetû populációk különbözõ szõlõfajtákon való fejlõdésének és szaporodásának tanulmányozását, valamint a szõlõ-gyökértetû genetikai vizsgálatával a filoxérák közötti különbségek és azonosságok megállapítását.
Az alanyfajták filoxéra rezisztenciájának értékelésekor megállapításra került, hogy a Georgikon 28 alanyfajta (Vitis vinifera származék) filoxéra ellenálló képessége jelenleg jó, mely az évek során feltehetõen gyengülni fog. A Teleki 5C alanyfajta gyökerén egyes populációk nagyon szaporodtak, nodozitásokat és tuberozitásokat okoztak. Így arra a következtetésre jutottunk, hogy a Teleki 5C és a filoxéra kapcsolatában változás állt be.
A magyarországi szõlõ-gyökértetû populációk szaporodási sajátosságait, populációdinamikai változásait és genetikai felépítését in vitro laboratóriumi (10 populáció), tenyészedényes (20 populáció) és genetikai (23 minta) kísérletekben megfigyelve bizonyosságot nyert, hogy a hazai filoxérák a vizsgált tulajdonságokban eltérõek.
A genetikai vizsgálatban 12 különbözõ DNS sávot tudtunk kimutatni a vizsgált filoxérák mintázata között, így a DNS-szintû elemzés igazolta, hogy a hazai filoxérák között nem csak szaporodásbiológiai, hanem genetikai különbség is van.
Bár a laboratóriumban vagy az üvegházban kapott eredmények, következtetések szabadföldi körülmények között nem minden esetben igazolódnak, az újabb növényegészségügyi gondok felmerülése elõtt javasoljuk aVitis berlandieri x Vitis ripariaalanyok filoxéra rezisztenciájának felülvizsgálatát.
1. 2. ANGOL NYELVÛ KIVONAT
The further results of researches for grape phylloxera populations existing in Hungary
Hungarian grape phylloxera populations were studied according to their development, reproduction, genetic structure and survival on roots.
The phylloxera feeding on the roots of resistant cultivar (Vitis berlandieri x Vitis riparia Teleki 5C) was described in this study. Our results indicate that a revised study of the relationship of rootstock and grape phylloxera in the field is required.
These experiments were the first concerning with the diversity of the reproduction and development in grape phylloxera populations originating from different Hungarian vinedistricts. The population from Sopron and some population near the Lake Balaton reproduced well on roots and caused some nodosities and tuberosities.
Our experiment was the first to examine grape phylloxera in Hungary by molecular genetic tools.
Differences in Hungarian phylloxera populations were demonstrated with laboratory bioassay, glasshouse examinations and genetic methods.
1. 3. NÉMET NYELVÛ KIVONAT
Die neue Ergebnisse der Untersuchung von der ungarischen Reblauspopulationen
In unserer Arbeit wurden zum ersten Mal die Populationsdynamik, der Anwuchs und die genetische Konstruktion der meisten ungarischen Reblauspopulationen untersucht.
Die unterschiedliche Schadwirkung der Reblaus wurde bei der toleranten Unterlagssorte (Vitis berlandieri x Vitis riparia Teleki 5C) aufgeschrieben. In den kommenden Jahren sollte man diese Erhebung weiterführen und in der Anlage sollte die Veränderung der physiologischen Verbindung von der Unterlagen und der Reblaus erforschen.
Die Population ‚Sopron’ und die Population ‚Balatongegend’ haben sich kurzhin vermehrt und haben viele Nodosität und Tuberosität verursacht.
In diesem Werk wurde die Differenz der genetischen Struktur von den ungarischen Reblauspopulationen bewiesen.
Zwischen den verschiedenen ungarischen Reblauspopulationen wurden die Unterschiede mit dem laboratoriumischen Test in vitro, mit der Arbeit im Gewächshaus und mit genetischer Laborarbeit gefunden.
2. BEVEZETÉS
A szõlõ-gyökértetû (Daktulosphaira vitifoliae FITCH) vagy más néven a filoxéra a szõlõ egy régóta ismert kártevõje, mely a szõlõ növény levelét és gyökerét egyaránt megtámadja. Az 1860-as évek elején került be Európába és ezután a szõlõ legveszedelmesebb kártevõje lett. Az élénkülõ szaporítóanyag-forgalommal világszerte elterjedt, így az 1870-es években Magyarországra is eljutott. A kártevõ megfékezése az elsõ sikeresnek tekinthetõ biológiai védekezési eljárás, az oltványhasználat általános elterjedésével valósult meg. Az új szaporításmód használatáig 1869 és 1900 között a kártevõ 2, 8 millió hektár saját gyökerû szõlõültetvényt pusztított el Európában, s hasonló mértékû kárt okozott a világ többi részein is (BOUBALS, 1966).
Az egyes szõlõfajok és fajták filoxéra rezisztenciája különbözõ. Ennek oka az, hogy 65 millió évvel ezelõtt a földrészek szétválása után a különbözõ éghajlati adottságok és a domborzat miatt a növényzet is eltérõen fejlõdött a földrészeken. Más szõlõfajok alakultak ki Amerikában, mások Eurázsiában és Kelet-Ázsiában.
Az amerikai szõlõfajokkal (pl.Vitis ripariaMichx.,Vitis rupestrisScheele,Vitis aestivalis Michx, Vitis cordifolia Michx) a kártevõ koevolúcióban élt, így ezeknek a filoxérával szemben természetes rezisztenciájuk van. Ez a magyarázata annak, hogy a filoxéra eredeti hazájában, Amerikában eleinte nem okozott észrevehetõ kárt. Az eurázsiai és kelet-ázsiai szõlõfajok, aVitis vinifera L. és aVitis silvestrisviszont nagyon fogékonyak.
Az elmúlt évek kutatási eredményei szerint a tõkék pusztulását a filoxéra a gombák és a baktériumok közremûködésével együttesen okozza. A kutatók kísérlete során bizonyítottá vált, hogy a kórokozók a filoxéra által okozott sérüléseken keresztül jutnak be a gyökérbe. A szõlõ tõkepusztulását számos kórokozó, elsõsorban gombák (Phomopsis viticola, Eutypa lata, Fusarium sp., Phytium sp., Stereum hirsutum, Rosellinia necatrix, Roesleria hypogea) okozhatják (OMER és mtsai, 1995; OMER és mtsai, 1997; OMER és GRANETT, 2000a).
Sokáig keresték a kártevõ elleni védekezés lehetõségét, de ez idáig a leghatékonyabb filoxéra elleni védekezési módszernek a rezisztens szõlõalanyok használata bizonyult. A rezisztenciára való nemesítés eredménye csak akkor tartós, ha sem a gazdanövény, sem a kártevõ örökletes alapja nem változik meg. Elméletileg azonban a gazdanövénynél rügymutáció révén elõfordulhatnak változások.
Másik oldalról gondot jelenhet még a kártevõ megváltozása, melyre több kutató is felhívta a figyelmet. A vizsgálatok során arra a következtetésre jutottak, hogy a filoxéra alkalmazkodik a gazdanövényhez és a környezethez. Erre elsõsorban a populáció nagyságából következtethetünk, hisz az együttélés bizonyos populáció nagyságig nem befolyásolja a faji változatosságot (KOCSIS és mtsai, 1999a).
Európában általánossá vált az oltványhasználat, ennek ellenére a XX. század második felében a szõlõ-gyökértetû újra tõkepusztulást okozott, melyet a filoxéra új biotípusainak kialakulásával és a nemesített fajták gyengébb morfológiai felépítésével magyaráznak. Az 1960-1970-es években Franciaországban Chasselas x Berl. 41 B (BOUBALS, 1966), az 1980-as évek második felében Kaliforniában Aramon x Rupestris Ganzin 1, az 1990-es évek elején NémetországbanV. berlandieri x V. ripariaTeleki 5C alanyon észleltek filoxéra fertõzésre visszavezethetõ tõkepusztulást (PORTEN és mtsai, 2000a). Hazánkban több mint száz éve ugyanazokat a szõlõ-gyökértetû ellenálló szõlõalanyokat használják a szõlõtermesztésben. Ez felveti annak lehetõségét, hogy az oltványhasználat ellenére a rovar ismét károsítóként lép fel.
CÉLKITÛZÉS
Magyarországi felmérések azt mutatják, hogy a szõlõ-gyökértetû minden hazai borvidéken megtalálható. Az alanytelepek és elhanyagolt ültetvények környékén kártétele már gazdasági méreteket is ölt (KOCSIS és mtsai, 1999b). A kártevõ gyors fejlõdését és alkalmazkodó képességét figyelembe véve újra indokolttá vált a rovar szaporodásának és a szõlõalanyok filoxéra rezisztenciájának részletes vizsgálata.
A szõlõ-gyökértetû biológiája, elterjedési területe és a kultúrnövényhez való alkalmazkodása kevéssé kutatott. A kártételi veszélyhelyzet értékeléséhez azonban ilyen irányú vizsgálatokra is szükség van. A fentiek figyelembe vételével munkánk során célul tûztük ki:
• a szõlõalanyfajták filoxéra rezisztenciájának vizsgálatát,
• a borvidékekrõl begyûjtött szõlõ-gyökértetû populációk különbözõ szõlõfajtákon való fejlõdésének és szaporodásának vizsgálatát,
• a szõlõ-gyökértetû DNS-szintû elemzését, a filoxérák között fennálló genetikai különbségek és azonosságok megállapítását.
3. IRODALMI ÁTTEKINTÉS
3. 1. SZÕLÕFAJOK KIALAKULÁSA, JELLEMZÕI
Vitis alnemzetségek, fajtacsoportjaik és fajok kialakulásáról azért kell beszélnünk, mert ennek nagy jelentõsége van a szõlõ növény és a szõlõ-gyökértetû között kialakult kapcsolatban. Vitisalnemzetségek, fajtacsoportok és fajok a természetes elterjedési területeken koevolúcióban éltek a filoxérával. Ezáltal néhány Vitis alnemzetségnél, fajtacsoportnál és fajnál kialakult a szõlõ-gyökértetûvel szembeni természetes rezisztencia, melyet késõbb a filoxéra elleni védekezésnél is felhasználtak (KOZMA, 1964, 1966 és 1991).
A Vitis nemzetségen belül két alnemzetség - Euvitis és a Muscadinia -, több taxonomiailag elkülönített fajcsoport vagy fajsorozat, továbbá földrajzilag elkülönült fõ- és alcsoportok alakultak ki.
Az Euvitis alnemzetség a Föld északi féltekén, túlnyomóan a mérsékelt éghajlatú övezetben, Európában, Ázsiában és Észak-Amerikában s kisebb részben a szubtrópusi, trópusi övezetben, Észak- és Közép-Amerikában, Délkelet-Ázsiában haladt végig a természetes evolúción, s foglalták el jelenlegi fajai (mintegy 59) és alfajai ismert areájukat. A Muscadinia 3 fajának az areája Észak-Amerika déli, meleg éghajlatú tájain (Florida, Mexikó) található (LÕRINCZ, 1999). A Muscadinia alnemzetség fajai immunisak a szõlõ-gyökértetûvel szemben.
A két alnemzetség egymástól tulajdonságaiban jól elkülönült. A különbségek arra engednek következtetni, hogy a két alnemzetség õsi képviselõi régen elváltak a fejlõdés útján.
A Vitis nemzetség fejlõdéstörténetében jelentõs folyamat volt a fajok földrajzi csoportjainak kialakulása. Ezt a folyamatot a kontinensek és nagyobb tájegységek eltérõ földrajzi-ökológiai hatásai irányították. A több százezer éves különélés a földrajzi fajcsoportokban jellegzetes morfológiai, különösen biológiai bélyegek és sajátosságok kialakulását eredményezte. A szõlõfajoknak fõleg azok a tulajdonságai alakultak ki, amelyek a környezetük viszonyaihoz való alkalmazkodást tették lehetõvé (pl. a hideg- és szárazságtûrés, kórokozókkal és kártevõkkel szembeni ellenállóság stb.). Viszont a faj fennmaradásával kevésbé kapcsolatos bélyegek (levél, virágtípus, termés) azonosak maradtak. A filoxérával szembeni ellenálló képességük a földrajzi elkülönülés alapján
változó. A legtöbb ellenálló faj Észak–Amerikában maradt fenn. Három nagy Vitis fajcsoport az észak- amerikai, az európai- nyugat- ázsiai (eurázsiai) és a kelet- ázsiai.
3. 1. 1. Szõlõ-gyökértetû és a szõlõ növény kapcsolata
Az amerikai szõlõfajok (pl. Vitis berlandieri, V. rupestris, V. riparia) egyes fajtái a kártevõ bizonyos biotípusaival szemben levél-, illetve gyökér ellenállóságot mutatnak. RAVAZ (1894) és HUSMANN (1930) is vizsgálta a különbözõ fajok, fajták filoxéra rezisztenciáját, melyben 20-as skálán értékelve Vitis rotundifolia, Vitis riparia, Vitis monticola és Vitis cordifolia bizonyultak a legellenállóbbnak. Teljes immunitást csupán a Vitis cinerea esetében tapasztaltak (BÖRNER és HEINZE, 1957). Az európai szõlõk (Vitis vinifera, V. silvestris) gyökérzete erõsen megsínyli a kártevõ gyökérlakó alakjának támadását, de erõsen károsodhatnak egyes amerikai fajok (V. labrusca) is.
A filoxéra rezisztens alanyhibridek létrehozásában hazánkban Teleki Zsigmond jelentõs eredményeket ért el és az amerikai fajok keresztezésével elõállított hibridjeit világszerte használják napjainkban is (BÉNYEI és LÕRINCZ, 1999).
A szõlõfajok ellenálló képessége azonban az F1 és F2 nemzedékben lecsökkenhet (1. táblázat), ezért fontos a fajták filoxéra ellenállóságának folyamatos vizsgálata.
1. táblázat. A filoxéra levéllakó alakjával szembeni ellenálló képesség átöröklõdése a szõlõfajok közötti hibridjeinek F2nemzedékében (NEGRUL, 1936)
3. 1. 2. A szõlõfajok filoxéra rezisztenciája (Az egyes fajok, fajták immunitásának kérdése)
A filoxérával szembeni ellenálló képesség komplex tulajdonság, sokféle magyarázat született rá. Az amerikai fajták ellenálló képességét kémiai tulajdonságaikkal, savak és gyantaszerû anyagok valamint polifenolok jelenlétével magyarázza PETRI (1910a, 1910b, 1911), nem pedig a másodlagos kéregréteg gyors megújulásával.
Más feltevés szerint a szõlõ ellenálló képessége attól függ, hogy a filoxéra nyálával a növénybe jutott ingeranyagok mennyire válnak hatástalanná a növényben (BÖRNER, 1930). Ez az anyag csak az erõs fejlõdésben lévõ levélen vált ki reakciót, ahol igen intenzív a sejtosztódás.
BÖRNER (1933) késõbbi vizsgálatai során is azt igazolta, hogy a szõlõ kémiai védõanyagai teszik hatástalanná a filoxéra gubacsot kiváltó ingeranyagait. A szõlõ- gyökértetû ingeranyaga szerinte a szõlõre nem mérgezõ. Erre a tényre abból következtetett, hogy a gubacsok a kártevõ idõ elõtti elpusztulásakor többé-kevésbé teljesen visszafejlõdnek és valamennyi sejt - a szipókával közvetlenül érintkezõk kivételével - életben maradnak. A filoxéra szúrásának ingermezejében nekrózis következik be, amint a kártevõ gubacsképzõ ingeranyagait a növény védõanyagai közömbösítik, és már csak a szõlõ-gyökértetû erõsen alkálikus hatású nyála hat pusztítólag az érintett sejtekre. A reakció kiváltójának a nyálban található ß-
A magoncok mennyisége (db)
F2hibridek Nem
károsodnak Szúrások Steril
gubacsok Félig norm.
gubacsok Normális gubacsok
37 63 38 589 74
Chasselas rose x Rup. 4401
%- ban 4,6 7,9 4,9 73,5 9,2
22 181 16 6 4
Mourvédre x Rupestris 1202
%-ban 9,6 79 7 2,6 1,7
47 43 8 181 108
Riparia x Gamay 595
%-ban 12,1 11,1 2,1 46,8 28
27 48 17 216 97
Riparia x Gamay 604
%-ban 6,7 11,8 4,2 53,3 23,9
10 8 5 66 30
Riparia x Gamay 605
%-ban 8,4 6,4 4,2 55,5 25,2
indolecetsavat tartották, azonban a mai kutatások szerint bár a ß-indolecetsav okoz hasonló tüneteket, a hatás mégsem azonos a filoxéranyál hatásával (GRANETT, 1990).
Megfigyelhetõ, hogy a gubacsképzésre nem képes V. vinifera fajtákon a tuberozitások nekrózisát korhadás követi, mely az egész gyökérzetre kiterjed.
Azt, hogy a szõlõ védekezési anyagai a filoxérára nem mérgezõek, az bizonyítja, hogy a heteken át nekrózis hatásnak kitett rovarok, védekezési anyagokat nem képzõ szõlõre átkerülve, normálisan fejlõdnek. Mások szerint ennek magyarázata a gazdanövény érzékenységében rejlik (túlérzékenységi reakció). A specializálódás ezek szerint az ingeranyagok kvantitatív különbségein és a szõlõ sejtjeinek eltérõ érzékenységén alapszik. A körülményeknek megfelelõen nekrózis, nekrobiózis vagy ingerhatás fog fellépni. Ha a gazdanövény vagy a sejtje nem érzékeny, akkor a gubacsképzõdés el is maradhat.
A filoxérarezisztencia-nemesítéssel kapcsolatban azt állítják, hogy egy szõlõfajta csak akkor mondható ellenállónak ha a parazitának semmi, vagy legalábbis nem kielégítõ életlehetõségeket nyújt. Ilyen esetben a kártevõ elvándorol vagy fiatal korában elpusztul.
Ez a felfogás az elsõ pillanatban meggyõzõnek látszik, ezért a legtöbb nemesítési módszernek ez volt az alapja. Sajnos ez egyáltalán nem egyezik meg a gyakorlati tapasztalatokkal. HUSFELD és SCHERZ (1934) szerint a szõlõ igen fogékony genotípusai a legerõsebb fertõzés esetén sem mutatnak károsodást. HUSFELD (1952) késõbbi kutatásai során vizsgált más genotípusokat is, amelyek a filoxérának semmi vagy csekély életlehetõséget nyújtanak. Ezért többé-kevésbé rezisztensnek kellett volna lenniük, ennek ellenére az aránylag jelentéktelen szívás esetén is jelentõsen károsodtak.
Újabban éppen azokat a genotípusokat értékelik pozitívnak, amelyek a legjobb életfeltételeket biztosítják a filoxérának, mivel éppen ezeken a szõlõfajtákon észlelhetünk abnormálisan erõs fertõzés esetén is aránylag jelentéktelen károsodást.
A szõlõ gyökércsúcsa a levéllel azonos módon reagál a gubacskiváltó ingerekre (ANDERS, 1957b). Itt is elég nagy mitózis gyakoriságra van szükség az életképes gubacsok fejlõdéséhez. A levélen csak igen rövid ideig vannak meg a gubacsképzõdéshez szükséges elõfeltételek (ANDERS, 1959), a gyökércsúcson viszont közvetlenül a gyökérsüveg mögött minden idõben igen nagy számban fordul elõ sejtmagosztódás és ezért állandóan kedvezõek a feltételek a kárképek elõidézéséhez. A károsítás elengedhetetlen feltétele a fokozott mitózis aktivitás, tehát a levélen idõhöz, a gyökéren helyhez kötött (ANDERS, 1957a).
Az ellenálló amerikai fajtáknál keskeny bélsugarakat találunk, s a gubacsos részeket másodlagos pararéteg választja el az ép gyökérrésztõl. Egyes kutatók így azt gondolták, hogy az ellenálló képesség a sebpara képzõdés gyorsaságától függ (SZPIRIN, 1961). Valószínûsíthetõ, hogy a tömött gyökérszöveti szerkezet az élénk anyagcsere, a gyors regeneráció, a cserzõanyag tartalom, az alacsony keményítõ- és fehérjetartalom együtt eredményezi az ellenálló amerikai fajták rezisztenciáját (SZALAY-MARZSÓ, 1989).
Morfológiai-anatómiai tényezõk is befolyásolják a szõlõ ellenállóságát. A levél és a vitorla szõrözöttsége, szélessége is befolyásolja a szõlõ-gyökértetû megtelepedését a levélen (JOHNSON és LEWIS, 1993). A filoxéra károsításával szemben védelmet jelent a vastag epidermisz és kutikula, a szövetek szoros kapcsolódása.
A rezisztens és nem rezisztens szõlõfajták felépítése teljesen más, melyet az évek során többen is vizsgáltak. A rezisztens szõlõfajták gyökerének felépítése sûrû, szerkezetük apró sejtû, növekedésük és fásodásuk gyors. Szerkezeti részek hirtelen mennek át egymásba. A xilém-gyûrûk jól kialakultak, a kambiális tevékenység erõteljes.
A periderma jól kifejlõdött és rajta vastagodások vannak. A parenchima szövetek fejlõdése gyenge, nagy számú és keskeny bélsugár fejlõdik, amelynek eltérési szöge kicsiny. A prozenchima szövetek fejlõdése erõs. A cseranyagtartalom magas, míg a keményítõ- és a fehérjetartalom alacsony. A sejtfalak ligninesedettek. A szárazanyagtartalom magas, a kalciumoxalát kristályok mennyisége kevés. A daganatképzõdésre ellenálló fajtákon a kártevõ sertéi felszínesen tudnak csak behatolni a tömöttebb szöveti szerkezet miatt. A károsodás mértéke kicsi, a daganatképzõdés és szétesés lassú. A keményítõszintézis szinte zavartalan. Az ellenálló fajták gyökérszövetének védõ reakciója során a seb-periderma tömör rétegû, a szövetek gyorsan forradnak. A háncs ledobása gyûrûsen történik, miközben a cseranyagok az endodermiszben fejhalmozódnak, melynek következményeként a seb-periderma több rétegûvé válik. A gyökerek és a szövetek regenerálódása fokozott (DOBOSNÉ, 1958).
Nem rezisztens szõlõfajták gyökerének felépítése laza, a szerkezete nagysejtû, a szerkezeti elemek fokozatosan mennek át egymásba. A növekedés és fásodás lassan zajlik le. A xilém-gyûrûk gyengén fejlettek. A kambiális tevékenység kevéssé erõteljes. A periderma fejlõdése szintén gyenge, melyben vastagodások ritkán fordulnak elõ. A parenchima szövetek fejlõdése erõs, a bélsugarak szélesek és kis számúak. A parenchima szövetek fejlõdése gyenge, a cseranyagtartalom alacsony, viszont a keményítõ- és
fehérjetartalom magas. A sejtfalak uralkodóan cellulózból állnak. A gyökér szárazanyag tartalma kevés, viszont a kalciumoxalát kristályok mennyisége sok. A gyökerek károsodása nagy, szövetek daganatosodása mélyreható, mivel a kártevõ szúrósertéi mélyre tudnak hatolni benne. A daganatok képzõdése és szétesése gyors, mely során a keményítõszintézis megszûnik. A fogékony fajták gyökérszövetének védõreakciói során seb-periderma ritkán képzõdik. A szövetek forradása során a szállítóedények elzáródása következik be. A háncs ledobása részleges, az endodermiszben cseranyagok nem halmozódnak fel. A seb-periderma általában egyrétegû. Gyökerek szöveteinek regenerálódása lassan indul meg (DOBOSNÉ, 1958).
3. 2. A SZÕLÕ-GYÖKÉRTETÛ
3. 2. 1. A faj bemutatása
• Társnevei: filoxéra, filloxera, fillokszéra, phylloxera
• Latin neve: Daktulosphaira vitifoliae FITCH, Viteus vitifolii FITCH, Dactylosphera vitifolii SHIN, Pemphigius vitifoliiFONCOLOMB, Peritymbia vitisana WESTWOOD, Phylloxera vastatrix FONCOLOMB, Rhizaphis vastatrix PLANCHON, Pemphigus vitifoliae FITCH, Phylloxera pemphigoides DONNADIEU, Filloxera quercusBOYER de FOUSCOLOMBE.
A faj jelenleg érvényes neve: Daktulosphaira vitifoliae FITCH, de az irodalmakban megemlítikViteus vitifoliFITCH néven is.
• Rendszertani helye: Ízeltlábúak törzse (Arthropoda), Rovarok osztálya (Insecta), Növénytetvek rendje (Sternorrhyncha), Levéltetvek alrendje (Aphidina), Filoxéra vagy törpetetû-félék családja (Phylloxeridae) (JENSER és mtsai, 1998).
• Elterjedése: Geopolita kártevõ, az egész világon elterjedt. Õshazája az Amerikai Egyesült Államok, jelenleg Kis-Ázsiában, Kelet-Ázsiában, Indiában, Mexikóban, Közép- és Dél-Amerikában, Ausztrália egyes részein, Új-Zélandon, Észak- és Dél-Afrikában, Madeira szigetén és Európában mindenhol megtalálható.
• Tápnövényei: A filoxérának mintegy 27 faja ismeretes, mely nagy része termesztett növényeken szembetûnõ kárt nem tesz. A Phylloxeridae-hez tartozó kártevõ károsítását pl. a Pecan dió (Carya illinoensis) levelén is igazolták (HEDIN és mtsai, 1985).
A szõlõn élõ faj monofág, melyet a szõlõ legjelentõsebb kártevõjeként tartanak számon. Néhány észak-amerikai fajon (Vitis berlandieri, Vitis rupestris, Vitis riparia) a fejlõdési ciklusa teljes, ezeknek a föld feletti részén is él. Egyes fajoknak, amelyek gyökere ellenálló csak a levelét fertõzi. Az európai fajtákon nem teljes a fejlõdési ciklusa, ezeknek fõleg a gyökerén él (1. ábra), a levelein csak ritkán képes kifejlõdni. Az újabb kutatások viszont azt is igazolták, hogy az európai szõlõfajták levelén is képes fennmaradni (DIÓFÁSI és MÁJER, 1999; GYÖRFFYNÉ és MÁJER, 2001, GYÖRFFYNÉ és mtsai, 2002; GYÖRFFYNÉ és mtsai, 2003).
1. ábra. Szõlõ-gyökértetû kõnyomata
(Horváth László nyomán)
3. 2. 2. A kártevõ fejlõdési alakjai
1.) Gyökérlakó alak (radicola): Kifejlõdve 1,2 - 1,8 mm hosszú, télen sárgásbarna, nyáron sárgászöld színû (SZALAY-MARZSÓ, 1998). Szárnyatlan, teste lapított, ovális (5. ábra). Hátoldalán 70, harántsorokba rendezett, sötétbarna szemölcs látható. Csápja 3 ízbõl áll. 3 pár lába rövid, csak az utolsó ízek látszanak ki a test alól. Nyugalmi állapotban az elsõ lábak között, a hasi oldalon látható szívószája, mely a csõszerû szipókából és a benne elhelyezkedõ 4 szúrósertébõl áll (LEHOCZKY és REICHART, 1968).
A szúróserték és a 3. lábpár lábszárának hossza alapján különíthetõk el az eltérõ életmódot folytató alakjai.
A rövid szipókájú (vitifolii) rassz gubacsban született gubacslakó egyedeire 2,5; a gubacsban született, gyökérlakó egyedeire 2,7 átlagos index jellemzõ, míg a hosszú szipókájú (vastatrix) rassz gubacslakó egyedeire 3,2-es, a gyökérlakó egyedeire a 3,3-as átlagos index jellemzõ (BÖRNER, 1939).
2.) A gyökérlakó alak tojása: Sima, fénytelen felületû, ellipszoid, világossárga (2. ábra), mely a lárva kelése elõtt olajzöldre sötétül. Hossza 0,3 - 0,5 mm, szélessége 0,16 mm.
3.) Gyökérlakó alak lárvája: Színe az anyaállathoz hasonló (3. ábra), de teste csak 0,4 mm hosszú.
4.) Nimfa: Nyúlánk, zöldessárga, kétoldalt barnás szárnykezdeményeket visel. Átalakulásig színe okkersárgává színezõdik és szemölcsössé válik.
5.) Szárnyas alak: Szûznemzéssel szaporodó nõstények. Teste sárga, a szárnya tövénél barna gyûrû van. Szemölcsös teste 1-1, 25 mm hosszú. 2 pár szárnya víztiszta testénél hosszabb (4. ábra).
4. ábra. Filoxéra szárnyas alakja (Fotó: Pintér Csaba) 3. ábra. Filoxéra gyökérlakó alakjának lárvája (Fotó: J. Granett)
2. ábra. Filoxéra fejlõdési alakjai gyökéren (Fotó: J. Granett)
Hossza összecsukott szárnyakkal 2 mm. Csápja 3 ízbõl áll, 3 pár lába hosszúnak látszik.
Csápjai között találhatók nagy összetett szemei, s a környékükön további 6 pontszeme.
6.) Szárnyas alak tojása: Világos- vagy okkersárgák, hatszögletes sejtalakban bordázottak, két típusba sorolhatók. A kisebbek 0, 26 - 0, 27 mm hosszúak és 0, 12 - 0, 14 mm szélesek. Ezekbõl kelnek ki a hímek. A nagyobb tojások 0,44 mm hosszúak és 0,18 - 0,2 mm szélesek, belõlük fejlõdnek ki a nõstények.
7.) Hím: Szárnyatlan, a gyökérlakó alakhoz hasonlít, de csupán 0,27 mm hosszú és 0,13 mm széles, sötétbarna színû (5.ábra). Kifejlõdött ivarszerve van, de tökéletlen szájszerve miatt táplálkozásra képtelen (HEGEDÛS és mtsai, 1966).
8.) Nõstény: Szárnyatlan, zöldessárga. A hímnél jóval nagyobb, 0,48 mm hosszú és 0,24 mm széles. Rendelkezik kifejlett ivarszervvel, de szájszerve fejletlen, így táplálkozásra képtelen.
9.) Téli tojás: Olajzöld, 0,27 - 0,30 mm hosszú és 0,11 - 0,13 mm széles. Felületén hatszögletes sejtbordázat van, egyik végén kampós függeléket visel.
10.) Õsanya (fundatrix): Duzzadt testû, sárgászöld és szárnyatlan (6. ábra). Hátoldalról sem csápjai, sem lábai nem láthatók. Testén nincsenek szemölcsök. Hossza 1,5 - 2 mm (PRINCZ, 1962).
11.) Õsanya tojása: Fényes világos zöldek (6. ábra), 0,25 - 0,28 mm hosszú és 0,11 - 0,14 mm széles.
12.) Õsanya lárvája:Alakra és méretre is hasonló a gyökérlakó alak lárváihoz (6. ábra).
13.) Gubacslakó alak (gallicola): Sötét okkersárga (7. ábra, 8. ábra), 1, 6 - 1, 8 mm hosszú, és 1 - 1, 2 mm széles.
14.) Gubacslakó alak tojásai és lárvái: Hasonlók az õsanya tojásaihoz, illetve a gyökérlakó és õsanya-lárvákhoz (7. ábra, 8. ábra).
5. ábra. Filoxéra hím
6. ábra. Filoxéra õsanya, tojásai (Fotó: J. Granett) és lárvái (Fotó: J. Granett)
7. ábra Filoxéra gubacslakó alakja és tojásai (Fotó: L. Tóth) 3. 2. 3. A filoxéra életmódja
A szõlõ-gyökértetû életmódját számos tudós vizsgálta. Az elsõk között volt APPENDIX (1880), aki már az 1870-es években elkezdte vizsgálni a rovar fejlõdését, majd tapasztalatait a Sonoma-i konferencián mutatta be. Úgy gondolta, hogy a filoxéra minden földrészen téli tojásokkal telel át. Szerinte az esõs idõ beköszöntése után egyre több téli alak alakul ki és ez a fejlõdési alak többé-kevésbé már az alvó forma. Kortársai Planchon, Lichtenstein és Balbiani is azt állítják, hogy a filoxéra hibernált (alvó) tojásból kel ki. Ezt a feltevést kiegészítve hozzá kell tennünk, hogy a hazai megfigyelések szerint a talajban történõ telelését nem hibernált tojás, hanem hibernált fejlõdési alak (lárva vagy nimfa) formájában tölti, viszont a föld feletti telelése elõfordulhat tojás alakban is (HORVÁTH és mtsai, 2000). A kifejlõdött egyed a levélre felmászva gubacsot hoz létre és megtermékenyítés nélkül több száz tojást rak le. Planchon a hibernált alak utáni elsõ generációnál közel 800 tojást számolt. Ha ez igaz, akkor egy vegetáció alatt a nyolc generáció közel 56 billió egyedet hoz létre. Hazai körülmények között az elsõ generáció fajtától függõen átlagosan 50-400 tojást hoz létre, így egy tojásból évente körülbelül 25 millió egyed fejlõdhet ki. Az év vége felé aztán a kártevõ öltözetet vált, szárnyas alakok fejlõdnek. Ezek pedig a levegõ áramlásának segítségével messzire juthatnak. A szárnyas alakok néhány tojást (2-5) hoznak létre, mely közül a kisebb a hím, a nagyobb a nõstény.
A kártevõ csak a levélgubacsokban él, ha az idõjárás megfelelõ. Ha viszont az idõjárás kedvezõtlenné válik, akkor a levéllakó alak gyökéren élõ alakká változik. Planchon
11. ábra Filoxéra fejlõdési alakjai a gubacsban (www.ipm.uiuc.edu) 8. ábra Filoxéra fejlõdési alakjai a
gubacsban (Fotó: J. Granett)
kutatásai is felhívták a figyelmet a két alak közötti hasonlóságra. Említésre méltó, hogy már ebben az idõben is megkülönböztették a szõlõ-gyökértetû nõstény és hím alakját.
Fatio az 1800-as évek végén a gyökéren élõ alakokat vizsgálva a kortársaihoz hasonlóan a gyökéren lévõ nodozitás okozói és a levélgubacsok lakói között azonosságot állapított meg. A filoxéra föld alatti és a föld feletti alakja annyira eltérõ, hogy korábban ezeket külön fajnak vélték és így a hasonlóság feltárása nagy lépést jelentett a filoxéra kutatás terén (PONGRÁCZ, 1978).
VIALA (1893) négy fejlõdési alakot különített el: a szexuális formát, a levéllakó alakot, a gyökérlakó alakot és a szárnyas nõstényt. Szaporodásbiológiai vizsgálatai során alapvetõ megállapításokat tett, amelyel hozzájárult a filoxéra kutatás fejlõdéséhez. A vizsgálataiban megfigyelte, hogy a szexuális alak a szárnyas nõstény lerakott tojásából kel ki. A levéllakó alak tavasszal jelenik meg és levélgubacsokat hoz létre, melynek nyílása a levél színén van. A gyökérlakó alak pedig nodozitást és tuberozitást okoz. WINKLER (1962) kutatásai alapján azt tapasztalta, hogy a szexuális alak nem fejlõdik Kaliforniában, így a föld feletti ciklus itt nem zajlik le.
BRAUN és RIEM (1950) fejlõdési ciklusa kiváló ábrázolása a kártevõ életmódjának, mely segít a rovar bonyolult szaporodásának megértésében. A vizsgálat eredményeként megkülönböztettek egy föld feletti és egy föld alatti ciklust, amely egymásba fonódik és körfolyamatot alkot. Megállapították, hogy a hibernált gyökérlakó alak szûznemzõ nimfává alakul és tojásokat rak. A tojásokból kikelõ lárvák újra nimfákká fejlõdnek, majd több átalakuláson szárnyas alakká formálódnak és a földfelszín fölé másznak. A szárnyas alak aztán ivaros tojásokat rak (hím és nõstény tojás), melyekbõl kikelõ egyedek egymással szaporodva ivaros tojást hoznak létre. Az ebbõl kikelõ egyedek felmásznak a levélre és a fejlõdési folyamatok végeredményeként nimfává alakulva több száz tojást raknak le a gubacsokban. A hideg beköszöntésével a fejlõdési alakok a talaj felé igyekeznek és ott hibernált lárva alakban telelnek át (9. ábra). A Braun és Riem-féle fejlõdési ciklus nagyon jónak bizonyult, ezért módosítások nélkül is sokáig ezt használták.
A filoxéra életmódját sokan kutatták, melyek közül a Braun és Riem-féle (9.
ábra), a Lewin-féle (10. ábra) és a Granett-féle (11. ábra) életciklus a legismertebb.
Fejlõdése földrészenként, elterjedési területenként változik, így az életmódját a helyi sajátosságokat figyelembe véve kell kutatni. Biztosat ma sem állíthatunk, de
legvalószínûbbnek az tûnik, hogy hazánkban a következõkben leírt módon zajlik a szõlõ- gyökértetû fejlõdése (12. ábra).
A gyökérlakó alak nõstényekbõl áll, melyek lárvaként (egyes leírások szerint nimfa) telelnek a vékony gyökereken. Tavasszal kifejlõdve a nõstény 3-4 vedlés után ivaréretté válik és szûznemzéssel 4-6 hét alatt lerakja 20-120 tojását, melyekbõl 5-10 nap után kelnek ki a lárvák, amik a gyökereket kezdik szívogatni. Körülbelül 4 hét és 3 vedlés után ivaréretté válnak, s szintén szûznemzéssel továbbszaporodnak. Hazai viszonyok közt 6-7 nemzedéke is lehet évente (PONGRÁCZ, 1978). Más irodalmak szerint csupán 4 nemzedéke fejlõdik ki egy évben (BOGNÁR és HUZIÁN, 1979). Az embrionális fejlõdés üteme a hõmérséklettõl függ: 13°C-on 57 napig, 25°C-on 13 napig tart. A nemzedékszám is hõmérsékletfüggõ: Németországra a 4, Szicíliára a 10-12 nemzedék éves kifejlõdése a jellemzõ (SZALAY-MARZSÓ, 1989).
Nyár végén és õsszel a gyökérlakó alak egy része eltérõen fejlõdik az eddig leírtaktól. A fejlõdési alak 3 vedléssel szárnykezdeménnyel rendelkezõ nimfává alakul, mely a talajfelszín felé mozog. A felszín közelében újra átalakul, ezúttal szárnyas szexupar nõsténnyé, mely nyár végén, õsszel rajzik ki. Egyes kutatások szerint a szárnyas szexupar nõstény a szárnyai és a szél segítségével nagy távolságot tesz meg. Leszállva 7- 8 tojást rak le a szõlõtõkék fás részeire vagy a levelek fonákára. A kisebb tojásokból ivarérett hímek, a nagyobbakból nõstények fejlõdnek ki. A táplálkozásra képtelen ivaros alakok a kikelés után párosodnak. A megtermékenyült nõstény egyetlen tojást rak le a 2- 3 éves vesszõk repedéseibe -ez az ún. "téli tojás"-, majd a nõstény is elpusztul.
Az áttelelõ téli tojásból ún. õsanyává fejlõdõ lárvák (nimfa) a felszín fölé mászva a föld feletti életmódú nemzedéksor kiinduló tagjai lesznek. Az ún. õsanyák május közepétõl-végétõl (JENSER, 1960) a fejlõdõ levelek színére vándorolnak, s ott megtelepedve szívogatni kezdik a növényi nedvet. Nyáluk hatására a levél fejlõdése sérül. Az õsanya alatti sejtek besüllyednek, a mellette fekvõk kiemelkednek, majd egy kis nyílás kivételével - melyet növényszõrök zárnak le - össze is zárulnak felette; május végére létrehozzák a jellegzetes filoxéra-gubacsot. Ez a körülbelül borsó nagyságú, a levélfonák felé dudorodó képzõdmény csak az amerikai szõlõfajtákra és hibridjeikre jellemzõ, de már az európai szõlõfajtákon is elõfordul (BOGNÁR és KANYÓ, 1984). Az újabb kutatási eredmények igazolják, hogy a filoxéra gubacsa nem homológ a korábban vélt indol-ecetsav hatására keletkezõ gubaccsal (GRANETT, 1990).
A nõstény a gubacsában 4-600 tojást rak, melyek a gubacs nyílásán kijutva a szomszéd leveleken hasonló gubacsok képzõdését indukálják. Tapasztalataink szerint a gubacsban létrejövõ tojások mennyisége nagy mértékben függ a fajtától, az ültetvény korától is. Az elsõ fejlõdési fokú lárvák két napig mozognak, táplálkoznak a leveleken, majd vedlésük után tovább fejlõdnek. A második fejlõdési fokú lárva szívogatásának hatására indul meg a gubacsképzõdés, mely a 6-7. napon fejezõdik be a gubacs záródásával (JENSER, 1960). Ezekben a fiatal lárvák kifejlõdnek, s szûznemzéssel 4, egymásba folyó nemzedéket nevelnek (BOGNÁR és HUZIÁN, 1979). A nyár elõrehaladtával a 4 nemzedék tagjainak egyre nagyobb hányada gyökérlakó lárvaként kel ki. Ezek levonulnak a gyökérre, ott kifejlõdve szûznemzéssel gyökérlakó nemzedéket nevelnek. Év végén a gubacsból származó teljes lárvanemzedék gyökérlakó alakká válik a szõlõ gyökerein, lezárva ezzel a 2 éves teljes fejlõdési ciklust (a levélgubacsban nem képes áttelelni). A teljes ciklus az európai szõlõfajokon és fajtákon csak elvétve megy végbe, rajtuk csak a gyökérlakó alak él meg.
A fejlõdési ciklus teljessége elterjedési területenként, szõlõfajonként és fajtánként is eltérõ. Az Észak-Kelet Amerikában õshonos szõlõfajokon fõleg a levéllakó alak figyelhetõ meg - ritkább a gyökérlakó alak. Az ivaros szakaszt szárnyas nimfák kezdik meg. Ugyanitt a V. vinifera-n csak a gyökérlakó alak él meg és az ivaros szaporodás is föld alatt zajlik (GRANETT és mtsai, 2001). A tapasztalat alapján a Dél- Nyugat Amerikában õshonos szõlõfajokon csak levéllakó alak él, sem szárnyas, sem ivaros alakot nem találtak a gubacsokban. Kaliforniában aV. vinifera-n és hibridjein csak gyökérlakó alak él. Az amerikai kutatások szerint a szárnyas alakok kifejlõdnek ezeken, de többségük tojásrakásra nem képes (GRANETT és TIMPER, 1987).
Ahol az oltvánnyal telepített ültetvényeken élõ szõlõ-gyökértetû populáció mérete igen kicsiny, ott a levélgubacsok csak elvétve fordulnak elõ - évtizedenként 1-2 évjáratban. Amerikai kutatók a V. californica-n csak néhány gyökérlakó alakot találtak, viszont Oregon, Washington és Brit KolumbiaV. viniferaültetvényein csak a gyökérlakó változat él meg. Az ültetvényekben megfigyelték a szárnyas alak kialakulását is, de nem tudják, hogy ez képes-e termékeny tojások rakására. Elõfordulnak olyan venezuelai õshonos szõlõfajok, melyeken csak a levéllakó alak él meg. A perui V. vinifera ültetvényeken levél és gyökérlakó alak is elõfordul. Ausztráliában a V. vinifera-n csak gyökérlakó alakot találtak, oltványokon gyökér- és levéllakók egyaránt elõfordulnak (GRANETT szóbeli közlése alapján).
A szõlõ-gyökértetû fejlõdésmenetei közül a legismertebbek:
9. ábra. Braun and Riem-féle életciklus (1950)
10. ábra. Lewin-féle életciklus (1993)
11. ábra. Granett-féle javított fejlõdési ciklus (GRANETT és mtsai, 2001)
12. ábra. Filoxéra hazai fejlõdésmenete, életmódja (HORVÁTH és mtsai, 2000)
3. 2. 4. A szõlõ-gyökértetû változatai
A szõlõ-gyökértetûnek két eltérõ típusa régóta ismert: a "Phylloxera vastatrix"
(KRIESCH, 1872) és a "Phylloxera vitifolii", melyeket szipókájuk hossza alapján különböztetünk meg (SCHILDER, 1947). Régebben még két külön fajnak vélték a két típust, melyek egymással keresztezõdhetnek és így a típusokon belül több fiziológiai rassz is létrejön. Az 1950-es és az 1960-as években JENSER (1959a) vizsgálta a filoxéra rövidszipókájú rasszának elõfordulását hazánkban.
Nemesítési szempontból 8 rassz jelentõs (BÖRNER, 1933; CSIZMAZIA, 1958), melyek alaptípusai közül az R436 hosszúszipókájú (vastatrix) és a 378 rövidszipókájú (vitifolii). A 208-as és a 438-as rassz hibrid, az 521-es fiziológiai rassz öntermékenyülésbõl keletkezett. További rasszok: 20-as, 140-es, 943-as. A rasszok az indukált tüneteiben és morfológiailag is eltérést mutatnak (DOBOSNÉ, 1958).
A filoxérának nemcsak rasszai vannak, számos tudós beszámol a különbözõ biotípusok létezésérõl is. Elsõként Kanadából gyûjtött filoxéra törzseket vizsgálva STEVENSON (1970, 2000) állapította meg, hogy legalább kétféle biotípus él Ontarioban.
Amerikában a leromló szõlõket vizsgálva 1983-ban Goheen (DE BENEDICTIS, 1996) a szõlõ gyökerén filoxérát talált. A körülmények a filoxéra okozta tõkepusztulásra utaltak, mely hatására újabb kutatási áramlat indult el világszerte.
1989-tõl egy új "B-típus" károsított Kaliforniában, mely 40-szeres szaporulatával egy milliárd USA dollár kárt okozott az Aramon x Rupestris alanyú szõlõültetvényekben, és a felmérések szerint további adaptálódott típusok megjelenésére és károsítására is lehet számítani (HERPAY, 1994). Az amerikai kutatásokban a "B- típus"-ú filoxéra a tojásrakás gyakoriságában, szaporodási rátájában, élethosszában, tojásszámban és fertõzõképességben is meghaladta a többi Napa-völgyi filoxéra populációt. A vizsgálat szerint megél, sõt szaporodik az "A-típusra" rezisztens St.
George alanyon is, viszont a V. vinifera õs nélküli alanyokon nem képes fennmaradni (GRANETT és mtsai, 1985).
Valószínûsíthetõen a filoxéra populációk alkalmazkodnak az élõhelyükhöz és több szõlõfajhoz kötõdõ biotípusuk jön létre (SONG és GRANETT, 1990; KOCSIS és mtsai, 1997). Erre utal, hogy a világ különbözõ tájairól és eltérõ szõlõfajairól,
szõlõfajtáiról gyûjtött filoxéra-típusok eltérõ mértékû kártételt mutattak gyökértenyészeteken és szabadföldi kísérletekben.
A filoxéra osztályozására a rassz, a biotípus és a törzs szavakat használják, melynek angol megfelelõi a ‚race’, a ‚biotype’ és a ‚strain’ szó. Sokszor a genetikai jelentésük nélkül használják a kifejezéseket (GRANETT és mtsai, 2001), ezért ezek tisztázása nélkülözhetetlen (PAP, 2004):
• A rasszok az egymástól elsõsorban morfológiai, részben biológiai tulajdonságaik alapján is elkülöníthetõ csoportok (BALÁS és SÁRINGER, 1982).
• A biotípusok valamilyen biológiai tulajdonság, elsõsorban az egyes tápnövényeken való viselkedés alapján egymástól különbözõ populációk. Ez azt jelenti, hogy az egyes biotípusok a szõlõfajokat, fajtákat és hibrideket különbözõ mértékben képesek fertõzni, illetve egyeseken egyáltalán nem tudnak megtelepedni, szaporodni (BALÁS és SÁRINGER, 1982).
• A törzs egyetlen tojásból származó, ivartalanul szaporodó populáció, melynek egyedei genetikai állományukban teljesen megegyeznek (GRANETT és mtsai, 1996).
3. 2. 5. Filoxéra kártétele
A filoxéra kártétele levélen és gyökéren egyaránt jelentkezhet. A levéllakó (v.
gubacslakó) alak szívogatásának hatására a levél fonákán vagy a hajtáson apró borsószem nagyságú, rücskös
felületû gubacsok jönnek létre (13. ábra). A gubacsok nyílása mindig a levél színén található, melyeket sûrû serték zárnak le (BOZAI és ÁNGYÁN, 1989;
BREIDER, 1939). Nagy levélfertõzés régebben csak az amerikai szõlõfajokon és hibridjeiken fordult elõ.
13. ábra. A filoxéra levélgubacsai (Fotó: L. Tóth H.)
Az amerikai szõlõfajokon és hibridjeiken lévõ rövidszipókájú rassz fertõzése olyan mértékû is lehet, hogy a gubacstömegtõl a levél csõvé sodródik (BÖRNER, 1914; BÖRNER és SCHILDER, 1933). A képzõdõ gubacs alakja a szõlõfajtától és a vegetációs állapottól függ: lehet nyeles vagy ülõ, gömbölyded vagy hosszúkás, sima vagy rovátkolt, szürkészöldes, szürkés barnás, vöröses (14. ábra), barnás vagy sárgás (BÖRNER, 1939).
Az 1950-es évek végén a filoxéra a hazai szõlõalany-telepeken 10 %-os vesszõhozam csökkenést okozott a V. berlandieri x V. ripariahibrideken és a V. riparia portalis-on (JENSER, 1960). A kártevõ terjedése növényi részekkel, szerszámokkal, karókkal és oltványokkal fokozottabb mértékû a természetes terjedési módnál.
Repedésekben, elváló kéregrészek alatt nagyobb csoportokban él. A gyökérlakó alak szívása nyomán, a korábbi kutatások szerint a nyálban talált béta-indolecetsav hatására (NYSTERAKIS, 1948; MILES, 1968) a vékony szõlõgyökereken gémfejre emlékeztetõ vastagodások (nodozitások) jönnek létre (15. ábra, 16. ábra).
15. ábra. Filoxéra kártétele szõlõgyökéren 16. ábra. Filoxéra gyökéren
(Fotó: J. Granett) (Fotó: J. Granett)
14. ábra. Vörös gubacsos fiatal levél (Fotó: J. Granett)
Szívogatásuk hatására a vastagabb gyökereken 1,5-2 mm széles és 0,5 mm magas dudorok (tuberozitások) keletkeznek. Vastagabb gyökereken a torzulás akár ökölnyi is lehet.
A dudorok idõvel elhalnak, leválásuk után alattuk kerek, barna folt marad (OMER és mtsai, 1999b). Õsszel a gyökérgubacsok a baktériumok, a sebzéseken át fertõzõ gombák - fõként FusariumésAspergillus fajok - közremûködésével szétesnek, elrothadnak és az egész gyökérrész tönkremegy (SZPIRIN, 1961).
Egyes kutatók szerint a tõkék pusztulását a filoxéra a gombák és a baktériumok közremûködésével együttesen okozza. Amerikai kutatók kísérlete során bizonyítottá vált, hogy a filoxéra által okozott sérüléseken keresztül jutnak be a gyökérbe a kórokozók (OMER és mtsai, 1995).
GRANETT (1998a) más kutatók munkájával egybehangzóan arra a következtetésre jutott, hogy a filoxéra vektorként a gombákat az egészséges növényre szállítja, így a filoxérának másodlagos kártételérõl is beszélhetünk. A gomba feltehetõen a filoxéra által okozott sebeken keresztül jut be és befolyásolja a filoxéra fejlõdését és szaporodását is.
A késõbbi kutatások azt igazolták, hogy az elhalást Fusarium solani, Fusarium oxysporum és Pythium ultimum fajok okozták. Összefüggés figyelhetõ meg az egyes szõlõfajok filoxéra ellenálló képessége, és a F. oxysporum érzékenysége között. A fertõzés következtében bekövetkezõ gyökérrothadás visszaveti a rovar túlélési és szaporodási mutatóit, ezért a szõlõ-gyökértetû elvándorol a rothadó gyökérrészekrõl, és új helyen fertõz (OMER és GRANETT, 2000a). Feltételezik azt is, hogy a korhasztó gombákon kívül a szõlõ-gyökértetû a szõlõ sárga mozaik vírusát (GFLV) is terjeszti (HORVÁTH, 1972; L. TÓTH és mtsai, 2004b).
A szõlõ pusztulásának vizsgálatakor OMER (2000b) összehasonlította a talajban élõ fitopatogének valamint a szõlõ-gyökértetû szerepét. A kísérletben a gomba aktivitása 18-26 oC között megnövekedett és kártételének jelentõsége felért a filoxéráéval. A szõlõ-gyökértetû károsításának következtében a gyökérzet pusztulni kezd, majd tönkremegy. Ennek hatására a föld feletti része gyengébben fejlõdik. A rothadás jele a felszíni részek gyengülõ fejlõdése - a rövidebb, vékonyabb vesszõképzõdés. Az ilyen vesszõk ízközei rövidebbek, levelei kisebbek és halványabbak, a termés elmarad, vagy törpe fürtök, töpörödött, lehulló szemek jellemzik.
A szõlõ-gyökértetû okozta kártétel mértékére a hosszanti gyökérnövekedésbõl is következtethetünk, de ehhez tudnunk kell a különféle szõlõfajokon és fajtákon a nodozitástól független hosszanti gyökérnövekedés %-os arányát.
A pusztulóban lévõ tõkérõl a gyökértetvek az egészségesre vándorolnak, így a fertõzött gócok egyre szélesednek. A fertõzés körkörösen szokott terjedni.
A tõkék pusztulása összefügg a kondíciójukkal, így silányabb talajon 3-4, jó termõtalajon 6-7 év alatt következik be (2. táblázat).
2. táblázat. A nodozitásképzés és a továbbnövekedési képesség 4 szõlõfajtán 4 különbözõ talajban (n=10 növény) (Hoffmann, 1957 cit. DOBOSNÉ, 1958)
Fajta Talajnem Az összes
nodozitások mennyisége
A túlnõtt nodozitások
mennyisége
A túlnõtt nodozitások
%-a
Kerti föld 158 17 10,7
Meszesmárga 102 9 8,8
Homokos vályog 88 5 5,7
Rizling 90
Agyag 70 3 4,3
Kerti föld 153 53 34,7
Meszesmárga 145 77 53,1
Homokos vályog 78 21 26,9
T.K.5BB
Agyag 108 33 30,6
Kerti föld 130 66 50,9
Meszesmárga 159 109 39,5
Homokos vályog 156 83 52,2
F.S.4- 201-39
Agyag 277 31 19,9
Kerti föld 80 33 41,3
Meszesmárga 152 32 21,1
Homokos vályog 146 83 56,8
F.S.4- 195-39
Agyag 53 21 39,6
3. 3. FILOXÉRA MAGYARORSZÁGON
3. 3. 1. Borászatunk helyzete a kártevõ megjelenése elõtt
A reformkorban megindult fejlõdésnek további lendületet adtak az 1867-es kiegyezés teremtette kedvezõ gazdasági feltételek. Elkezdõdött a szakemberképzés, melyre már nagy szükség volt. A Vincellér- és Kertészképzõ Gyakorlati Tanintézet ezidõtájt nyitotta meg kapuit.
Ekkor az ültetvényeinkre jellemzõ volt a szabálytalan telepítés, a nagy tõkeszám - 10-17 ezer tõke/hektár -, a bak- és fejmûvelés valamint a rövidcsapos metszés.
Területeink jelentõs részén semmilyen támaszrendszert nem alkalmaztak, máshol is csak karót használtak. Sokféle fajtát termesztettek hazánkban, de fõleg a pontusiakat részesítették elõnyben. Csemegeszõlõ-termelésünk elenyészõ volt, a termelt bor 58,4 %- ban fehér, 41,6 %-ban vörös és rozébor volt (CSELÕTEI és mtsai, 1978).
Mindezek ellenére az 1890-es évekig Magyarország a 4,5 millió hektoliternyi éves átlagtermelésével a világ negyedik bortermelõje volt és a minõség terén boraink a francia borokkal vetekedtek az elsõ helyért.
3. 3. 2. A szõlõ-gyökértetû elterjedése Magyarországon
A Nyugat-Európai ültetvények fokozódó mértékû pusztulását a szaklapok - Borászati Füzetek, Természettudományi Közlöny – leközölték ugyan, de a késõbb vaklármának bizonyuló 1872-es pozsonyi szõlõ-gyökértetû elõfordulása ösztönözte csak lépésre a Földmûvelésügyi Minisztériumot. A Minisztérium ezután körlevelekben szólította fel a gazdasági és borászati egyleteket a kártevõ felbukkanásának azonnali jelentésére, és figyelmeztette a termelõket a francia és olasz szõlõvesszõ-behozatal veszélyességére. A kártevõ felbukkanásának hatására 1873-ban azonnal megtiltották a gyökeres vesszõk behozatalát (HERMAN, 1879).
Ekkor még nem tudhatták, hogy elkéstek ezekkel az intézkedésekkel (BECK, 2000). A filoxéra hazánkba történõ behurcolásában valószínûleg a legnagyobb szerepet a Klosterneuburg kísérleti szõlõültetvényeinek késõn felismert (1874) fertõzõdése játszotta, mivel a szaporítóanyag innen az ezt megelõzõ idõszakban Magyarország legkülönbözõbb részeire került (SZALAY-MARZSÓ, 1989).
Egy felderítõ vizsgálat során egyesek szerint Gerger Ede távirdaellenõr 1873- ban vagy 1875-ben, más írások alapján Deininger Imre 1876 szeptemberében fedezte fel a kártevõt Pancsova (Torontál vármegye, ma Szerbia) határában (BOGNÁR, 1976). A kormányrendelet, melyben a terület kiirtását határozták el, az elsõ szõlõmûveléssel foglalkozó jogszabály volt Magyarországon (FEYÉR, 1970). A fertõzés 1876-ra 70-rõl 140 holdra terjedt ki. Az irtást a nagy költségek miatt abbahagyták, a kártevõ terjedését a pancsovai szõlõterületek zárlat alá helyezésével oldották meg. Az importált vesszõvel történõ telepítések felülvizsgálatát ezután is folyamatosan végezték. További hat helyen fedezték fel a kártevõt 1878-ban, a pancsovai góc már 700 holdra nõtt. Ennek hatására a kormány országosan betiltotta a szõlõvesszõ-forgalmat, megkezdõdött a szénkéneges védekezés az Érmelléki borvidéken, majd 1879-ben a kártevõ zalai felbukkanásának hírére a kormány további szénkéneg-mennyiséget rendelt Németországból. Ebben az évben hozták létre a Borászati Kormánybizottságot Miklós Gyula vezetésével (SZABÓ, 1985). Törvényerõre emelték 1880-ban a Berni egyezmény elõírásait (FEYÉR, 1970), s megalakult az Országos Filoxéra Bizottság.
Az egy holdon aluli területeken irtást, a nagyobb területeken pedig a szénkéneges gyérítést rendelt el a Bizottság. Pancsován felállították az Országos Phylloxera Kísérleti Állomást, ahol francia tapasztalatok nyomán ellenálló fajtákkal végeztek kísérleteket. Egyaránt vizsgálták a szénkéneg hatását és a kártevõ biológiáját is.
Nem sokkal a Bizottság megalakulása után filoxérát találtak Versecen, Tahitótfalun, Székesfehérváron, Beregszász és Szendrõ határában is. Ennek hatására szeptember elsejétõl megkezdték a szõlõk országos átvizsgálását.
Az elsõ ellenálló szõlõ szállítmány 1881-ben érkezett meg a farkasdi kísérleti állomásra Riparia, Elvira, York Madeira és Kék Izabella fajtákkal. Szinte azonnal megkezdõdtek a kísérletek a különféle oltási módokkal. Hiába volt ismert és elterjedt a gyümölcsfák oltása, nagy volt a bizalmatlanság a szõlõ oltásával szemben, pedig a külföldön is jól ismert szõlõ zöld oltása magyar oltási mód.
Az állam is a szénkéneges gyérítést támogatta a meglévõ ültetvények védelme érdekében. A francia tanulmányutak és a saját tapasztalatok is azt igazolták, hogy az oltvány egy évtized után is életképes, a kezdeti nagy pusztulás oka a nem megfelelõ párosítási mód és alany- nemes választás, illetve a talajviszonyok voltak.
Még 1880-ban rendelettel írták elõ azt, hogy a községek június 10. és 25.
között összes szõlõiket vizsgáltassák át és jelentsék az eredményt. Ennek felügyeletével a
járási filoxérabiztost bízták meg (SZABÓ, 1985). Egyre pontosabb és ijesztõbb eredmények születtek a kártevõ terjedésérõl. Míg 1879-ben 7 góc volt fertõzött, 1880- ban már 38, 1881-ben 50, 1882-ben pedig 79 községben igazolták a kártevõ elõfordulását (HERMAN, 1880).
Az 1883-as XVII. tc. összegezte az eddigi tapasztalatokat. A törvény tiltotta, illetve korlátozta a szõlõkben használt eszközök forgalmát. Az állam az immunis homokterületeken és az árasztásos rendszerben telepített szõlõkre adókedvezményt nyújtott, illetve díjtalanul biztosította a szénkéneget. Rohamosan terjedt a homoki telepítés. Futóhomokon 1896-ig éves átlagban 2.462 hektár új szõlõt telepítettek. A homoki szõlõk részaránya 1897-re 25 %-ról 36 %-ra emelkedett (FÜR, 1976). 1890-ben 51.765 hektár, 1915-ben pedig már 132.353 hektár homoki ültetvény állt (ZILAI, 1983).
Mindez nem gátolta a kártevõ terjedését. 1883-ban 130 községben, 1884-ben 246, 1885-ben 396 község határában fedezték fel jelenlétét. Ugyanebben évben 50 ezer, 1886-ban már 85 ezer hold pusztulásával számoltak. Már a kormány is módosította a korábbi rendeleteket. A zárlati csoport tagjai közt és az immunis homokon történõ telepítésekhez is engedélyezte a vesszõforgalmazást, mivel a vesszõforgalom teljes tilalma gátolta az újratelepítéseket. 1887-ben 810 község 132.352 holdja volt fertõzött.
Alig terjedt a szénkéneges gyérítés, mert ezt az állam már nem szolgáltatta ingyenesen. Az 1890-es évek végéig a szert csak 2-3 ezer hektáron használták, a fertõzött szõlõk 2-4 %-án (FÜR, 1976). Állami támogatást kaptak az ellenálló fajtákat szaporító szõlõtelepek. Használatukat azonban gátolta az is, hogy az új fajtákkal szemben elég nagy volt a bizalmatlanság. 1888-ra 1.249 község, 1889-re 1.566 község volt fertõzött, s a filoxéra eljutott Erdélybe is. Megindultak a szénkénegezõ tanfolyamok.
1890-ben 2.061 fertõzött községet tartottak nyilván.
Új módon szabályozták a szõlõvesszõ-forgalmat. A zárolt területekre és az immunis homoktalajú ültetvényekre bármilyen vesszõt be lehetett vinni, a zár alatt nem lévõ területekre csak homokról kizárólag sima vesszõt vihettek be. Eleinte szállítási bizonyítvánnyal vihettek csak vesszõt, késõbb nélküle is.
A fokozódó borhamisítás megszüntetése érdekében 1893-ban megszületett az elsõ bortörvény. 1896-ban érvénybe lépett a szõlõ-felújítási törvény is. A filoxéravész következményeinek felszámolásában a törvény fontos szerepet játszott, mely Darányi Ignácz magyar királyi földmûvelésügyi miniszter nevéhez fûzõdik (DARÁNYI, 1898).
Törvénybe foglalták a borvidék-beosztást és a javasolt-engedélyezett fajták listáját.
A kormány fokozta az elpusztult szõlõk felújításának támogatását, biztosította az oltványellátást, a tanfolyamok és pinceegyletek szervezését valamint kedvezményes hitelt nyújtott. Támogatta az oltvány-elõállító magántelepeket és elõírta az éves munkafolyamatokat a mûvelési tervben.
Az alanytelepek kezdetben minden amerikai szõlõfajt kipróbáltak, amihez csak hozzájutottak. A 90-es évek közepére már csak a bevált Riparia portalis, Vitis solonis, Rupestris metallica maradt termesztésben. Az 1890-es és 1897-es idõszak között 30 állami telep 538 kataszteri holdon 50 millió amerikai vesszõt termelt. Gondot jelentett azonban a hazai termesztésbe vonás után, hogy a Riparia portalis nem fejlõdik megfelelõen meszes talajú borvidékeinken, a Rupestris metallica pedig hõigénye miatt csak a déli országrészeken vált be. A mésztûrõ V. berlandierinehezen szaporítható, így hazai körülmények között igazán csak a V. berlandieri x V. riparia hibridek váltak be, amelyek Teleki Zsigmond 1896-tól induló keresztezéseinek eredményei. A megfelelõ eszközökkel 1897-ben sikerült megfékezni a vészt, de addig is 225.200 hektár szõlõültetvény pusztult el (CSELÕTEI és mtsai, 1978) - más adatok szerint 230.185 hektár (BOGNÁR és KANYÓ, 1984).
3. 3. 3. A "filoxéravész" hatása
A filoxéravész új termesztéstörténeti korszakot nyitott és a hátrányok mellett pozitív változásokat is hozott.
• kötött talajokon általánossá vált a filoxéra-ellenálló alanyú oltványok telepítése;
• az "immunis" homoktalajokon nagymértékben kiterjedt a saját gyökerû ültetvények területe;
• az alanyvesszõ-termesztéssel és a hajtatott oltványtermesztéssel bõvültek a szõlõszaporítási eljárások;
• általánossá vált a szabályos sor- és tõketávolságra ültetés, amely elõmozdította a természetes gépesítést (KOZMA, 1991);
• elindította a fajták választékának a racionalizálását, állami szabályozását;
• megkezdõdött a filoxéra elleni vegyszeres és technikai védelem;
• elindította a kétezer éves termesztési eljárások (támrendszer, tõkemûvelés, metszés, trágyázás stb.) korszerûsítését (KOZMA, 1991);
• serkentette a szelekciós és keresztezéses szõlõnemesítést, fajtahonosítást, a módszeres kutatások megindulását és a kutatóhálózat kifejlesztését (KOZMA, 1991);
• élénkítette a szõlõtermesztõ országok nemzetközi kapcsolatainak a kiépítését.
A szõlõ-gyökértetû két évtizedes pusztítása során közel 300 ezer kataszteri hold területû és fõleg elavult-elhanyagolt hazai szõlõket semmisített meg. A felújítással nem csak technológiánk, de a szõlõültetvények területi szerkezete is megváltozott (FEYÉR, 1970).
Az oltványhasználat, a forgatásos mûvelés megdrágította a termelést, lassította a termõre fordulást. Csak a megfelelõ hozamot hozó, termékeny területeket telepítették újra oltványokkal, a nehezen mûvelhetõ magashegyi szõlõket nem (FEYÉR, 1970). Az immunis alföldi homokos talajokat betelepítették.
Megváltozott a fajtaösszetétel is. A gyengén termõ tájfajták helyét a bekerülõ francia fajták vették át. Elterjedt az Olasz rizling, Oportó, Kékfrankos, Cabernet, Merlot, Sauvignon, Szürkebarát és a csemegeszõlõk. Sajnos a "direkttermõ" fajták aránya 20-25
%-ra nõtt. Megnõtt az egy tulajdonosra jutó ültetvényméret (CSIZMADIA, 1998).
Több helyen a szõlõk helyét más kultúra foglalta el. Makón a hagyma, Békésben a búza, Buda környékén az õszibarack, Dél-Dunántúlon a szarvasmarha tenyésztés vette át a szõlõmûvelés helyét.
A szõlõk pusztulása fokozott lendületet adott a gazdasági válság miatt jelentkezõ kivándorlásnak. 1881-1898 között 372 ezer ember vándorolt ki az országból.
Mûveléstechnológiai újítás a szõlõk rendszeres trágyázása, néhol mûtrágyázása valamint a táblás, soros, négyzetes és fajtatiszta telepítés. Elterjedt a karóhasználat, a talajforgatás, a hosszúcsapos és szálvesszõs metszés. Rendszeressé vált a növényvédelem.
Közvetlenül a kártevõnek köszönhetõ a szõlészetet és borászatot szabályzó törvények megszületése, valamint a növényvédelmi és növény-egészségügyi intézményrendszer kialakulása.
3. 4. SZÕLÕ-GYÖKÉRTETÛ KUTATÁS A VILÁGBAN
A filoxéra kutatással kapcsolatos nemzetközi szakirodalmat földrészenként, országonként taglaljuk, mert a kártevõ okozta gondok és megoldásuk földrajzi egységenként változnak (Ausztrália-karantén rendszer, Európa-oltványhasználat stb.).
Amerikai Egyesült Államok
Az 1860-as években hurcolták be a filoxérát Európába, majd pár évtized múltán több országban és földrészen is jelentkezett a kártétele. A rovar károsítását elõször az európai behurcolással egyidõben észlelték Kaliforniában. A kártevõ elleni védekezés alapját itt is az Európában nemesített alanyfajták adták. 1905-tõl folyamatosan végeztek kísérleteket, hogy megállapíthassák a Kaliforniai körülmények között melyik alanyfajták alkalmasak leginkább a termesztésre. Az Egyesült Államokban több mint 50 év kutatás alatt összegyûlt adatok alapján az A×R#1-nek nevezett (V. vinifera Aramon× V.
rupestris Ganzin 1) alanyfajtát ajánlották az éghajlathoz, helyi talajhoz, nemes fajtákhoz legjobban alkalmazkodó alanyként. A kutatások megalapozottak voltak, így az ajánlások alapján 1960-tól „telepítési láz” tört ki és közel 20 év alatt az ültetvények 60-70%-át ezzel az alannyal telepítették (GRANETT és mtsai, 1996).
Az amerikai kutatások során a legjobbnak bizonyult alanyfajtát 1879-ben Ganzin állította elõ Vitis vinifera Aramon és a Vitis rupestris keresztezésével (CSEPREGI és ZILAI, 1976). A keresztezés elõnye aV. rupestrisfiloxéra ellenállósága és aV. vinifera- tól örökölt szárazság- és mésztûrés volt. Most már tudjuk, hogy a Ganzin 1 alanyfajta filoxéra rezisztenciája idõvel gyengül. Európa és Dél-Afrika egyes helyein már pár évtizedes használat során azt tapasztalták, hogy az alanyfajta elvesztette a filoxérával szembeni rezisztenciáját. Az 1980-as években Kaliforniában is megfigyelték a filoxéra károsítását a Ganzin 1 alanyfajta gyökerén.
A filoxéra rezisztenciával kapcsolatos kutatások során megindultak a rovar életmódjával, alaktanával és szaporodásával kapcsolatos kutatások is. STEVENSON (1970) Kanada különbözõ részeirõl gyûjtött filoxéra törzsekkel különbözõ szõlõfajtákat és alanyfajtákat fertõzött meg. A filoxéra törzsek szaporodását és károsítását megfigyelve az egyes alanyfajták levelén tapasztalt tulajdonságokból (gubacs okozása és levélszáradás kiváltásának képessége) megállapította, hogy legalább kétféle filoxéra (két biotípus) él Kanadában.