Plant Diseases Caused toy
Bacteria
Introduction
B A C T E R IA are very small, microscopic plants. About 1600 bacterial species are known. T h e great majority of bacteria are strictly sapro- phytic and as such are beneficial to man b e c a u se they help d e c o m p o se the enormous quantities of organic matter produced yearly by man and his factories as waste products or as a result of the death of plants and animals. Several species cause diseases in man including tuber- culosis, pneumonia, and typhoid fever, and a similar number cause diseases in animals, e.g., brucellosis and anthrax. About 180 species of bacteria have b e e n found to cause d i s e a s es in plants. All pathogenic bacteria are facultative saprophytes and can be grown artificially on nutrient media.
Bacteria are simple microorganisms usually consisting of single cells. T h e y may be rod-shaped, spherical, ellipsoidal, spiral, comma- shaped, or filamentous (threadlike). S o me bacteria can m o ve through liquid m e d ia by means of flagella, while others have no flagella and cannot m o ve themselves. S o me can transform themselves into spores and certain filamentous forms can produce spores, called conidia, at 322
Characteristics of Plant Pathogenic Bacteria 323
the e n d of the filament. Other bacteria, however, do not produce any spores. T h e vegetative stages of most types of bacteria reproduce by simple fission. Bacteria multiply with astonishing rapidity and their significance as pathogens stems primarily from the fact that they can produce tremendous numbers of cells in a short period of time. Bac- terial d i s e a s es of plants occur in every place that is reasonably moist or warm, they affect almost all kinds of plants, and, under favorable environmental conditions, they may b e extremely destructive.
Characteristics of Plant Pathogenic Bacteria Morphology
Almost all plant pathogenic bacteria are rod-shaped, the only excep- tions b e i ng two species of Streptomyces, which are filamentous. T h e rod-shaped bacteria are more or less short and cylindrical, and in young cultures they range from 0.6 to 3.5 μ in length and from 0.3 to 1.0 ì in diameter. In older cultures or at high temperatures, the rods of s o me species are m u ch longer and they may even appear filamentous.
Sometimes deviations from the rod shape in the form of a club, a Y or V shape, and other branched forms occur, and s o me bacteria may occa- sionally occur in pairs or in short chains.
T h e cell walls of bacteria of most species are e n v e l o p ed by a vis- cous, g u m my material which may b e thin (when it is called a " s l i me layer") or may b e thick, forming a relatively large m a ss around the cell (when it is called a " c a p s u l e " ). Most plant pathogenic bacteria are e q u i p p ed with delicate, threadlike flagella which are usually consid- erably longer than the cells by which they are produced. In some bac- terial species each bacterium has only one flagellum, others have a tuft of flagella at one en d of the cell; s o me have a single flagellum or a tuft of flagella at each end, and* still others have flagella distributed over the entire surface of the cell.
In the filamentous Streptomyces species, the cells consist of non- septate branched threads, which usually have a spiral formation and produce conidia in chains on aerial hyphae.
Single bacteria appear hyaline or yellowish-white under the com- p o u nd microscope, and are very difficult to observe in detail. Whe n a single bacterium is allowed to grow (multiply) on the surface or within a solid m e d i u m, its progeny soon produces a visible mass called a col- ony. Colonies of different species may vary in size, shape, form of e d g e s, elevation, color, etc., and are sometimes characteristic of a
given species. Colonies may b e a fraction of a millimeter to several centimeters in diameter, and they are circular, oval, or irregular. Their e d g es may b e smooth, wavy, angular, etc., and their elevation may b e flat, raised, dome-shaped, wrinkled, etc. Colonies of most species are whitish or grayish, but some are yellow, red, or other colors.
Anatomy
Bacterial cells have thin, relatively tough, and somewhat rigid cell walls which s e em to b e quite distinct from the inner cytoplasmic m e m b r a ne but which sometimes appear to intergrade and merge with the outer slime layer or capsule. T h e cell wall contains the cell con- tents and allows the inward p a s s a ge of nutrients and the outward pas- sage of waste matter, digestive enzymes, and other products given off by the bacterial cell.
All the material inside the cell wall constitutes the protoplast. T h e protoplast consists of the cytoplasmic or protoplast m e m b r a n e, which determines the degree of selective permeability of the various sub- stances into and out of the cell; the cytoplasm, which is the complex mixture of proteins, lipids, carbohydrates, many other organic com- pounds, and minerals and water; and the nuclear material, which ap- pears as spherical, ellipsoidal, or d u m b b e l l - s h a p ed bodies within the cytoplasm.
In the cytoplasm there are also certain proteinaceous granules, mi- tochondria, and nuclei. In addition, there are granular structures com- p o s ed of starch, glycogen, fat, volutin, or closely related compounds, all of which are reserve food substances.
Flagella, when present, are long, very fine bundles of complex pro- tein fibrils attached to the cell in various locations and originating from basal granules, just inside the cytoplasmic m e m b r a n e. T h e fla- gella propel the bacteria by spiral waves which pass along the length of each wriggling flagellum. T h e outer surface of a bacterial cell is e n v e l o p ed by a slime layer, or by a microcapsule, each of which is very thin, or by a capsule, which may be up to 10 μ thick. T h e chemi- cal composition of these outer layers is usually very complex, and var- ies widely with the species, but it has specific chemical properties that help us differentiate b e t w e en very similar types of bacteria which could not otherwise be differentiated. T h u s, this distinctive surface constitution of bacteria is useful in the identification of unknown species.
Characteristics of Plant Pathogenic Bacteria 325
Multiplication and Genetics
Rod-shaped phytopathogenic bacteria reproduce by the asexual process known as "binary fission" or "fission." This occurs by the inward growth of the cytoplasmic m e m b r a ne toward the center of the cell forming a transverse m e m b r a n o us partition dividing the cyto- plasm into two approximately equal parts. T wo layers of cell wall ma- terial, continuous with the outer cell wall, are then secreted or syn- thesized b e t w e en the two layers of m e m b r a n e. Whe n the formation of these cell walls is completed, the two layers separate, splitting the two cells apart.
While the cell wall and the cytoplasm are undergoing fission, the nuclear material b e c o m es organized in chromosome-like structures which duplicate themselves and b e c o me distributed equally b e t w e en the two cells formed from the dividing one. Although the internal changes that go on in the bacterial nucleus during cell division are not yet well understood, it s e e ms that they are quite similar to, or the s a me as, those h a p p e n i ng during mitosis in higher forms of life.
Although bacteria multiply by fission, not all the offspring are of the s a me genetic material. In a very small percentage of bacteria, muta- tions occur that cause changes in the genetic material and result in permanent modifications of the characteristics or properties of the bacteria. Also, fission may b e interrupted occasionally in certain spe- cies by a sexual process known as "conjugation." In conjugation two compatible bacteria come into contact side by side and a small portion of the genetic substance (DNA) of the donor cell (male) is transferred to the D NA acceptor cell (female). T h e acceptor cell undergoes fission following conjugation, and its progeny contain a combination of the D NA of two individuals (genetic recombination). Conjugation s e e ms to have a rejuvenating effect on the protoplasm of bacteria enabling them to carry on through fission for many generations.
T wo other processes are known that involve the transfer of genetic material from one bacterium to another. S o m e t i m es genetic material (DNA) is liberated from one bacterium by secretion or by rupture of its cell. S o me of the liberated D NA finds its way to a genetically com- patible bacterium of the same or closely related species and b e c o m es part of the genetic substance of that cell. T h e recipient cell now is genetically different b e c a u se it p o s s e s s es its own characteristics plus the additional characteristics s u p p l i ed by the a d d ed D N A. This pro- cess is called "transformation."
T h e other process is called "transduction" and involves transfer of
genetic material from one bacterium to another by means of bacterio- phages (viruses infecting bacteria). Whe n a bacteriophage infects a bacterium it injects its D NA into the bacterium. As the D NA repli- cates and more bacteriophages are produced, one or several of them may enclose some genetic material of the bacterium within their pro- tein coat. Whe n such a bacteriophage is liberated and infects a ne w bacterial cell, the genetic material from the first cell joins and be - comes part of the genetic material of the second bacterium. This cell is now genetically different and displays ne w or different properties from those it showed before transduction.
Regardless of the genetic p h e n o m e na involved in bacterial repro- duction, bacteria can and do reproduce at an astonishingly rapid rate.
Under favorable conditions bacteria may divide every 20 minutes, one bacterium b e c o m i ng two, two b e c o m i ng four, four b e c o m i ng eight, and so forth. At this rate one bacterium conceivably could produce one million bacteria in 10 hours. But, b e c a u se of the diminution of the food supply, the accumulation of metabolic wastes, and other limiting factors, reproduction slows down and may finally c o me to a stop. Bac- teria do reach tremendous numbers in a short time, however, and cause great chemical changes in their environment. It is these changes c a u s ed by large populations of bacteria that make them of such a great significance in the world of life in general and in the de- velopment of bacterial diseases of plants in particular.
Ecology and Spread
Almost all plant pathogenic bacteria d e v e l op partly in the host plant as parasites and partly in the soil as saprophytes. T h e r e are great dif- ference s among species, however, in the d e g r ee of their d e v e l o p m e nt in the one or the other environment.
S o me bacterial pathogens, such as Erwinia amylovora, which causes fire blight, produce the bulk of their populations and the great- est number of their generations in the plant host, while in the soil p h a se their numbers decline rapidly and usually do not contribute to the propagation of the disease from season to season. T h e se pathogens have d e v e l o p ed sustained plant-to-plant infection cycles, often via insect vectors and, either b e c a u se of the perennial nature of the host or the association of the bacteria with its vegetative propagating or- gans or seed, they have lost the requirements for survival in the soil.
S o me other bacterial pathogens, such as Agrobacterium tumefa- ciens, which causes crown gall, build up their populations within the host plants, but these populations only gradually decline when they
Characteristics of Plant Pathogenic Bacteria 327
are released into the soil. If susceptible hosts are grown in such soil in successive years, sufficiently high numbers of bacteria could be re- l e a s ed to cause a ne t increase of bacterial populations in the soil from season to season. Finally, other bacterial pathogens, such as some of the Erwinia and green-fluorescent Pseudomonas soft rotters, produce their populations largely in the soil. Their relation to plant d i s e a se is erratic and transitory, and it is not essential to the continued existence of these bacterial species.
Whe n in the soil, bacteria live freely or saprophytically or in their natural bacterial ooze, which protects them from various adverse fac- tors. Bacteria may also survive in or on s e e d s, other plant parts, in- sects, etc., found in the soil. On the plants bacteria may survive epi- phytically, in b u d s, on wounds, in their exudate, or inside the various tissues or organs which they infect.
T h e dissemination of plant pathogenic bacteria from one plant to another or to other parts of the s a me plant is carried out primarily by water, insects, other animals, and man. E v en bacteria p o s s e s s i ng fla- gella can m o ve only very short distances on their own power. Rain, by its washing or spattering effect, carries and distributes bacteria from one plant to another, from one plant part to another, and from the soil to the lower parts of plants. Water also separates and carries bacteria on or in the soil to other areas where host plants may b e present. In- sects not only carry bacteria to plants, but they inoculate the plants with the bacteria by introducing them into the particular sites in plants where they can almost surely develop. In some cases bacterial plant pathogens also persist in the insect and d e p e n d on them for their survival and spread. In other cases, insects are important but not es- sential in the dissemination of certain bacterial plant pathogens.
Birds, rabbits, etc., visiting or moving a m o ng plants may also carry bacteria on their bodies. Man helps spread bacteria locally by his handling of plants and by his cultural practices, and over long dis- tances by transportation of infected plants or plant parts to ne w areas or by introduction of such plants from other areas. In cases in which bacteria infect the s e e ds of their host plants, they can be carried in or on them for short or long distances by any of the agencies of s e ed dispersal.
Classification
All bacteria causing plant diseases b e l o ng to the class Schizomy- cetes, orders P s e u d o m o n a d a l e s, Eubacteriales, and Actinomycetales.
Order F a m i ly G e n us Plant p a t h o g e n ic s p e c i es P s e u d o m o n a d a l es P s e u d o m o n a d a e c ae Pseudomonas 90
Xanthomonas 60
E u b a c t e r i a l es R h i z o b i a c e ae Agrobacterium 7 E n t e r o b a c t e r i a c e ae Erwinia 17 C o r y n e b a c t e r i a c e ae Corynebacterium 11 Actinomycetales S t r e p t o m y c e t a c e ae Streptomyces 2
Recently, certain plant-infecting bacteria have b e e n tentatively shown to b e l o ng to other genera, e.g., Aerohacter (Enterobacte- riaceae), Bacillus (Bacillaceae), and others, but their taxonomic position and their importance as plant pathogens are still largely unknown.
Identification
T h e genus Streptomyces can be easily distinguished from the other bacterial gener a b e c a u se of its much-branched, well-developed myce- lium and curled chains of conidia. Identification of bacteria belonging to the rod-shaped genera, however, is a much more complex and diffi- cult process, and it can be m a de by taking into consideration not only visible characteristics such as size, shape, structure, and color, but also such obscure properties as chemical (antigenic) composition, enzymatic action, pathogenicity to plants, susceptibility to certain vi- ruses (bacteriophages).
T h e shape and size of bacteria of a given species in culture can vary with age of the culture, composition and p H of the m e d i u m, tempera- ture, and staining method. Under given conditions, however, the pre- dominating form, size, and arrangement of cells in a pure culture are quite apparent, and they are important and reliable characteristics.
T h e presence, number, and arrangement of flagella on the bacterial cell are also determined, usually after the flagella have b e e n stained with specific stains.
T h e chemical composition of certain substances in bacterial cells can b e easily detected with specific staining techniques. Information about the presence or a b s e n ce of such substances is u s ed for identifi- cation of bacteria. Gram's staining reaction differentiates bacteria into gram positive and gram negative. In this reaction bacteria are treated T h e families and gener a of bacteria that can c a u se d i s e a se in plants are tabulated under each order.
Characteristics of Plant Pathogenic Bacteria 329
with a crystal violet solution for 30 seconds, rinsed gently, treated with iodine solution, and rinsed again with water and then alcohol.
Gram-positive bacteria retain the violet-iodine stain combination b e - cause it forms a complex with certain components of their cell wall and cytoplasm. Gram-negative bacteria have no affinity for the stain combination, which is therefore r e m o v ed by the alcohol rinse and the bacteria remain as nearly invisible as before. Of the rod-shaped phyto- pathogenic bacteria, only the gener a Bacillus and Corynebacterium are gram positive. Agrobacterium, Erwinia, Pseudomonas andXantho- monas are gram-negative. T h e p r e s e n ce or absence of certain food reserve materials is also studied for purposes of identification of phy- topathogenic bacteria. T h e se reserves include primarily volutin, fat, glycogen, and iogen, of which the first two have b e e n found in phyto- pathogens.
T h e enzymatic action of bacterial cells is studied by recording the substances which the bacteria can u se for food and also the substances produced when the bacteria grow on certain media. T h us different bacteria can utilize different combinations of substances containing carbon or nitrogen in various forms, can cause hydrolysis of starch and proteins, or can exhibit lipolytic activity. Whe n bacteria grow on cer- tain substrates they may reduce nitrates to nitrites, may produce hy- drogen sulfide, ammonia, indole, and many other substances.
Determination of minimum, optimum, and maximum temperatures and hydrogen-ion concentrations (pH) for growth of bacteria as well as the thermal death point (that is, the temperature which kills a culture of a given bacterial species in 10 minutes) are also u s ed in their identi- fication.
Whe n an unknown bacterium has b e e n through all these tests, its identification is m a de by comparison of its reactions to those listed in tables available for the known species of bacteria. T h e unknown is considered to b e the same as the species listed that gives exactly the same reactions.
Phytopathogenic bacteria are also tested on various species and va- rieties of host plants for their pathogenicity on them. This test some- times, and for practical purposes, may be sufficient for tentative iden- tification of the bacterium.
Serological methods have b e e n u s ed for quick and very accurate identification of bacteria and have gained popularity in recen t years.
O n ce a phytopathogenic bacterium has b e e n identified by the meth- ods described previously, antiserum is prepared. For this, an animal, e.g., rabbit, is injected with a suspension of the known bacteria. Blood is obtained from the animal several weeks afer injection. After the
clots are removed from the blood, the clear liquid contains the antis- erum. T h e antiserum contains antibodies specific against the bacterial species or strain with which the animal had b e e n injected. If an un- known bacterium is suspected to b e the same as the one that had b e e n u s ed to produce the antiserum, suspensions of the unknown are mixed in test tubes with dilutions of the antiserum for verification. If the two bacteria are of the same species or strain, the bacteria in the test tube with the appropriate antiserum clump together and precipitate to the bottom. If the two bacteria are not the same, no precipitate is formed and the bacteria stay in suspension in the test tube.
In some cases bacterial species and strains can b e identified by the bacteriophages that infect them. Bacterial viruses may b e strain- or species-specific in that they can attack one strain or all the strains within a species of bacteria. Whe n such specific bacteriophages are available and it is suspected that the unknown bacterium is of the strain or species attacked by the virus, the identity of the bacterium is established by bringing the bacteria and the virus together. If the bac- teria are infected by the virus, they are of the species for which the bacteriophage is specific. It must be taken into consideration, howev- er, that specific bacteriophages are not known for all species of phyto- pathogenic bacteria, that some strains may b e insensitive to phages specific for other strains within the same species, and that some phages are polyvirulent or nonspecific (since they can attack bacteria of several species including some which may not even b e plant pathogens). Owing to these limitations the uses of bacteriophages for identification of bacteria remains rather limited.
Symptoms C a u s ed by Bacteria
Plant pathogenic bacteria cause the d e v e l o p m e nt of almost as many kinds of symptoms on the plants they infect as do fungi. T h e y c a u se leaf spots and blights, soft rots of fruit, root, and storage organs, wilts, overgrowths, scabs, cankers, etc. Any given type of symptom can b e c a u s ed by bacterial pathogens in several genera, and each genus con- tains some pathogens capable of causing different types of diseases.
S p e c i es of Agrobacterium, however, can cause only overgrowths or proliferation of organs. On the other hand, overgrowths can also be c a u s ed by certain species of Corynebacterium, Pseudomonas, and Xanthomonas.
How Bacteria Cause Plant Diseases 331 H ow Bacteria C a u se Plant D i s e a s es
Phytopathogenic bacteria penetrate plants through wounds and/or through natural openings. Once inside the plant some bacteria live and reproduce only in the intercellular spaces and cannot penetrate the cells as long as they are'alive. Such bacteria may secrete enzymes that break down the m i d d le lamella and result in maceration of the tis- sues a c c o m p a n i ed by death of the cells. Macerated parenchymatous cells lose their turgor and collapse b e c a u se of loss of water to the in- tercellular spaces and sometimes to the surface of the tissues (where it appears as bacterial ooze). C o l l a p s ed cells die, their death b e i ng sometimes c a u s ed or accelerated by toxins secreted by the pathogen.
T h e soft-rot Erwinias, the fire blight organism (Erwinia amylovora), and the wild fire of tobacco organism (Pseudomonas tabaci) are exam- ples of pathogens acting in this fashion. Sometimes penetration of still living parenchymatous cells may occur, and this s e e ms to hasten their death. Breakdown of cell walls and contents by bacterial enzymes results in the formation of lysigenous cavities. If such cavities form in the cortex, open cankers eventually appear. Whe n the necrotic tissue is on the leaf and is limited in size by the resistance of the host or by the environment, the d i s e a se may appear as a leaf spot. If, however, the host plant cannot stem the advance of the pathogen and the weather is favorable, the bacteria may spread, intercellularly at first, throughout the leaf and into young twigs, where they kill the paren- chymatous tissues in their path and the d i s e a se appears as a blight.
S o me bacterial pathogens affect plants by invading the vessels of the plants where they reproduce, spread, and carry on all their meta- bolic activities. T h e original penetration of the plant may be through wounds in the roots, stem, or foliage or through natural openings.
Sometimes the bacteria c a u se no local symptoms at the point of infec- tion, and at other times they may c a u se leafspots, cankers, or root rots.
T h e main d a m a ge to the plant, however, does not result from such localized injuries but from the invasion of the vessels. Once in the vessels, the bacteria secrete e n z y m es which break down pectic and other substances found in or b e t w e en the vessels and in the neighbor- ing parenchymatous tissues. T h e products of breakdown of these sub- stances may then enter the v e s s e ls where, together with the bacterial cells, the slimy polysaccharides they produce, and the substances produced by the host cells as a reaction to the pathogen, they cause clogging of the vessels. This results in reduction or complete blocking
of translocation of water and nutrients, and the plant b e c o m es stunted, wilts, and dies.
T h e m o de of action of bacteria in the genus Agrobacterium differs from those of other phytopathogenic bacteria in that, instead of caus- ing reduction of growth or destruction of cellular tissue, the bacteria of this genus stimulate cell division (hyperplasia) and cell enlargement (hypertrophy) in the affected tissues. This can result in the develop- men t of amorphous overgrowths, galls or tumors on roots, stems or other organs (e.g., crown gall, c a u s ed by Agrobacterium tumefaciens) or in proliferation of infected organs (e.g., hairy root, c a u s ed by A. rhi- zogenes). Certain species of Corynebacterium also can stimulate cell division and produce overgrowths on susceptible plants. Regardless of the size of overgrowths produced by bacteria of these two genera, no cell destruction can be found inside them. Any decay develops from the outside inward and is usually the result of secondary inva- sions by other microorganisms. On the other hand, overgrowths pro- d u c ed by bacteria of the genus Pseudomonas and Xanthomonas de- velop large cavities in the center, where the cells are destroyed. T h e walls of the cavities are covered with m a s s es of these bacteria.
Overgrowths of any of the kinds mentioned above affect the plants by diverting nutrients to the overgrowths and away from the normal tissues and processes of the plant and, sometimes, by crushing vascu- lar and other tissues of the plant in their vicinity. In either case, the amount of nutrients available to the plant is reduced, resulting in smaller growth and yields by affected plants.
In general, bacteria more easily infect, and cause much greater d a m a ge to, young seedlings and plants and the young, succulent shoots of older plants than they do to older plants and tissues. This perhaps is d ue to the greater content of water and nutrients of the young, tender tissues and to the a b s e n ce of anatomical or chemical barriers to the spread of bacteria, since many such barriers are formed only in later stages of the plant's life.
Control of Bacterial Diseases of Plants
Bacterial diseases of plants are usually very difficult to control. Fre- quently, a combination of control measures is required to combat a given bacterial disease. Infestation of fields or crops with bacterial pathogens should b e avoided by introducing and planting only healthy s e e ds or plants. Sanitation practices aiming at reducing the inoculum in a field by removing and burning infected plants or branches, and at reducing the spread of bacteria from plant to plant by
Control of Bacterial Diseases of Plants 333
decontaminating tools and hands after handling d i s e a s ed plants, are very important. Adjusting certain cultural practices, such as fertilizing and watering, so that the plants will not b e extremely succulent dur- ing the period of infection may also reduce the incidence of disease.
Crop rotation can b e very effective with disease-producing bacteria that have a limited host range, but is impractical and ineffective with bacteria that can attack many types of crop plants.
T h e u se of crop varieties resistant to certain bacterial d i s e a s es is one of the best ways of avoiding heavy losses. Varying d e g r e es of resist- ance may be available within the varieties of a plant species, and great efforts are m a de at crop breeding stations to increase the resistance of, or introduce n e w types of resistance into, presently popular varieties of plants. Resistant varieties, s u p p l e m e n t ed with proper cultural prac- tices and chemical applications, are the most effective means of con- trolling bacterial diseases, especially when environmental conditions favor the d e v e l o p m e nt of disease.
T h e u se of chemicals to control bacterial diseases has b e e n , general- ly, m u ch less successful than the chemical control of fungal diseases.
Soil infested with phytopathogenic bacteria can b e sterilized with steam or electric heat and with chemicals such as formaldehyde and chloropicrin, but this is practical only in greenhouses and in small b e ds or frames.
S e e d, w h en superficially infected, can b e disinfected with mercuric chloride solutions or by soaking it for several days in a weak solution of acetic acid. Whe n the pathogen is inside the s e ed coat and in the embryo, such treatments are ineffective. Treating s e ed with hot water does not usually control bacterial diseases b e c a u se of the relatively high thermal death point of the bacteria.
Of the chemicals u s ed as foliar sprays, copper c o m p o u n ds have given the best results. However, even they seldom give satisfactory control of the d i s e a se when environmental conditions favor the devel- opment and spread of the pathogen. Bordeaux mixture and fixed cop- pers are most frequently u s ed for the control of bacterial leaf spots and blights. Zineb is also u s ed for the same purpose, especially on young plants which may b e injured by the copper compounds.
Antibiotics have b e e n u s ed in recen t years against certain bacterial diseases, and the results are encouraging. S o me antibiotics are ab- sorbed by the plant and are distributed systemically. T h e y can b e ap- plied as sprays or as dips for transplants. T h e most important antibac- terial antibiotics in agriculture are formulations of streptomycin or of streptomycin and Terramycin. Several others are presently available, but most of them are still u s ed primarily for experimental purposes.
Since bacteriophages kill their host bacteria and since phages spe-
cific against certain phytopathogenic bacteria were found, it was ex- pected that phages would b e very valuable in controlling bacterial plant diseases. In some cases, the incidence and severity of some bac- terial plant diseases were reduced by spraying the plants with specific bacteriophages under experimental conditions. T o date, however, this means of attack against bacterial d i s e a s es has not b e e n d e v e l o p ed sufficiently, and it cannot be u s ed against any bacterial d i s e a se in the field, although work in this area may prove extremely valuable in the near future.
Selected References
B u d d e n h a g e n, I. W. 1965. T h e relation of plant p a t h o g e n ic bacteria to the soil. In E c o l o gy of Soil-borne Plant Pathogens (K. F. B a k er a nd W. C. Snyder, E d s . ), p p.
2 6 9 - 2 7 9 . Univ. of Calif. Press, Berkeley, California.
Carpenter, P. L. 1965. " I m m u n o l o gy a nd S e r o l o g y ," 2 n d ed., 4 5 6 p p. S a u n d e r s, Phila- delphia, Pennsylvania.
D e k k e r, J. 1963. Antibiotics in the control of plant d i s e a s e s. Ann. Rev. Microbiol. 17:
2 4 3 - 2 6 2 .
D o w s o n, W. J. 1957. " P l a nt D i s e a s es D ue to B a c t e r i a ," 2 n d ed., 2 3 2 p p. C a m b r i d ge Univ. Press, L o n d on a nd N ew York.
Elliott, Charlotte. 1951. " M a n u al of Bacterial Plant P a t h o g e n s ," 2n d ed., 186 p p. Chron- ica Botanica, Waltham, M a s s a c h u s e t t s.
Frobisher, M. 1962. " F u n d a m e n t a ls of M i c r o b i o l o g y ," 7th ed., 6 1 0 p p. S a u n d e r s, Phila- delphia, Pennsylvania.
Krueger, W. W., a nd K. R. J o h a n s s o n. 1961. " P r i n c i p l es of M i c r o b i o l o g y ," 2n d ed., 563 p p. S a u n d e r s, Philadelphia, Pennsylvania.
O k a b e, N., and M. Goto. 1963. B a c t e r i o p h a g es of plant pathogens. Ann. Rev. Phyto- pathol. 1: 3 9 7 - 4 1 8 .
Spooner, Å. T. C , a nd B. A. D. Stocker (eds). 1956. " B a c t e r i al A n a t o m y ," 3 6 2 p p. C a m- b r i d ge Univ. Press, L o n d on a nd N e w York.
S t a p p, C. 1961. " B a c t e r i al Plant P a t h o g e n s ," 2 9 2 p p. Oxford Univ. Press, L o n d on a nd N ew York.
Fire Blight
Occurrence and Importance
Fire blight is the most destructive disease of pear in the eastern half of the United States and also causes d a m a ge to pear and a p p le or- chards in other parts of the United States, in Canada, N ew Zealand,
Fire Blight 335
J a p an and, since 1957, E n g l a n d. It has b e e n reported from Italy, Egypt, Eastern E u r o p e, the Near East, India, Mexico, and other parts of the world, but there is little information on its prevalence and de- structiveness in these areas.
Fire blight is most destructive on pear, making commercial pear growing under certain conditions impossible. Certain apple and quince varieties are very susceptible to the d i s e a se and may be dam- a g ed as severely as pear trees. Many other plant species in the rose family (Rosaceae) and s o me nonrosaceous hosts are affected by fire blight, including several of the stone fruits and many cultivated and wild ornamental species. Although most of these other species can serve as hosts for overwintering of the pathogen and may b e affected to varying degrees, only those in the pome-fruit group are affected seriously.
Fire blight d a m a g es susceptible hosts by killing flowers and twigs, and by girdling of large branches and trunks resulting in the death of the trees. Youn g trees in the nursery or in the orchard may b e killed to the ground by a single infection in one season.
Symptoms
T h e first symptoms of fire blight appear usually on the flowers, which b e c o me water-soaked, then shrivel rapidly, turn brownish to black in color and may fall or remain hanging in the tree (Fig. 56A).
Soon the symptoms spread to the leaves on the s a me spur or on nearby twigs, starting as brown-black blotches along the midrib and main veins or along the margins and b e t w e en the veins. As the blackening progresses, the leaves curl and shrivel, hang downward and usually cling to the curled, blighted twigs.
Terminal twigs and watersprouts ("suckers") are usually infected directly and wilt from the tip downward (Fig. 57A). Their bark turns brownish black and is soft at first but later shrinks and hardens. T h e tip of the twig is hooked, and the leaves turn black and cling to the twig. F r om fruit spurs and terminals the symptoms progress down to the supporting branches, where they form cankers. T h e bark of the branch around the infected twig appears water-soaked at first, later b e c o m i ng darker, sunken, and dry. If the canker enlarges and encir- cles the branch, the part of the branch above the infection dies. If the infection stops short of girdling the branch, it b e c o m es a dormant or inactive canker, with sunken and sometimes cracked margins (Fig. 58).
F i g. 56. (A) B l o s s om blight on p e ar c a u s ed by the fire blight bacterium Erwinia amylo- vora. (B) F i re blight on a p p le fruit. (Photos by courtesy of the D e p a r t m e nt of Plant Pathology, Cornell University.)
Fruit infection usually takes place through the pedicel, but direct infection is not uncommon. T h e fruit b e c o m es water-soaked, then it turns brown, shrivels, mummifies, and finally turns black (Fig. 56B).
D e ad fruit may also cling to the tree for several months after infection.
Under humid conditions, droplets of a milky-colored, sticky ooze may appear on the surface of any recently infected part (Fig. 57B). T h e ooze usually turns brown soon after exposure to the air. T h e droplets may coalesce to form large drops which may run off and form a layer on parts of the plant surface.
The Pathogen: Erwinia amylovora
T h e fire-blight pathogen is a rod-shaped bacterium, 1.0-2.0 x 0.8-1.2 μ in size, although longer forms, sometimes up to 35 ì , have b e e n observed in cultures. T h e bacterium is motile, p o s s e s s i ng fla- gella over its entire surface (Fig. 59). T h e bacteria occur usually sin- gly, but pairs or chains of 3 or 4 bacteria also exist. E a ch bacterial cell is enclosed in a capsule. T h e bacteria are facultative anaerobes. Er
winia amylovora cultures lose their virulence after repeated transfer in artificial media.
Fire Blight 337
F i g. 57. (A) T w ig blight p h a se of fire blight on a p p l e. N o te that the infection b e g an at the tip a nd p r o g r e s s ed d o w n w a r d. (B) D r o p l e ts of bacterial o o ze running d o wn the surface of a p e ar twig affected with fire blight. (Photos by courtesy of the D e- partment of Plant Pathology, Cornell University.)
Development of Disease
T h e bacteria overwinter at the margins of cankers formed during the previous season on pear trees and other susceptible hosts (Fig. 58).
T h e y survive most often in large branches and seldom in twigs less than 1 cm in diameter. In the spring, the bacteria in these " h o l d o v e r"
cankers b e c o me active again, multiply, and spread into the adjoining healthy bark. B e c a u se of the breakdown of tissues and b e c a u se of the
F i g. 58. A fire blight canker that has s p r e ad from the small branch at left to the larger branch or stem. T h e fire blight bacteria over- winter at the margins of such cankers. (Photo by courtesy of the D e p a r t m e nt of Plant Pathology, Cornell University.)
secretions of the bacteria, the osmotic pressure of the liquids contain- ing the bacteria in the infected area increases greatly. During humid or wet weather, water is absorbed by these bacterial m a s s e s, which increase in volume beyond the capacity of the tissues, so that parts of them exude through lenticels and cracks to the surface of the tissue.
This gummy exudation, called bacterial ooze or exudate, consists of plant sap, millions of bacteria, and bacterial byproducts. T h e ooze usually appears first about the time when the pear blossoms are open- ing. Various insects, such as flies, wasps, honey b e e s, are attracted to the sweet, sticky exudate and b e c o me smeared with it. Whe n they visit flowers afterward, they leave some of the bacteria-containing exudate in the nectar of the flower. In some cases bacteria may also b e carried from oozing cankers to flowers by splashing rain (Fig. 59).
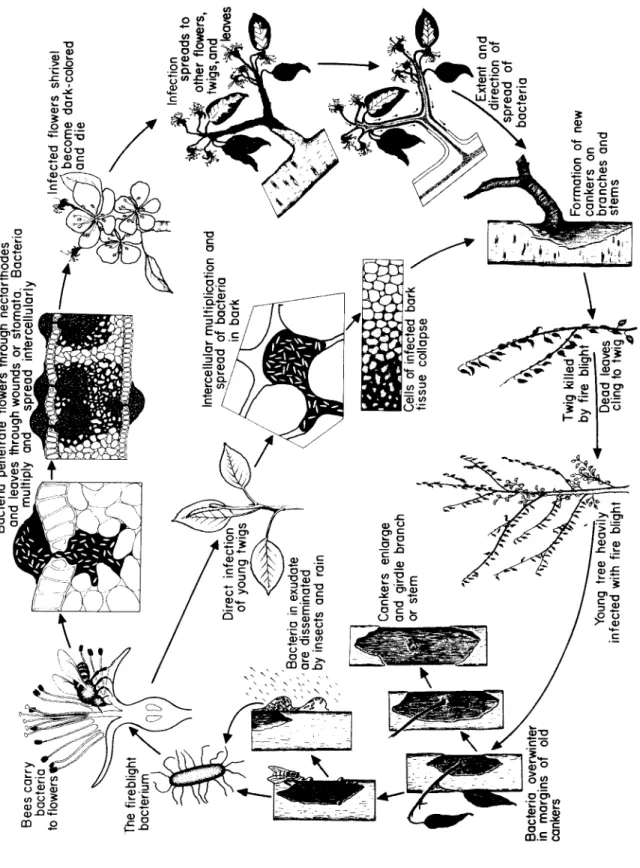
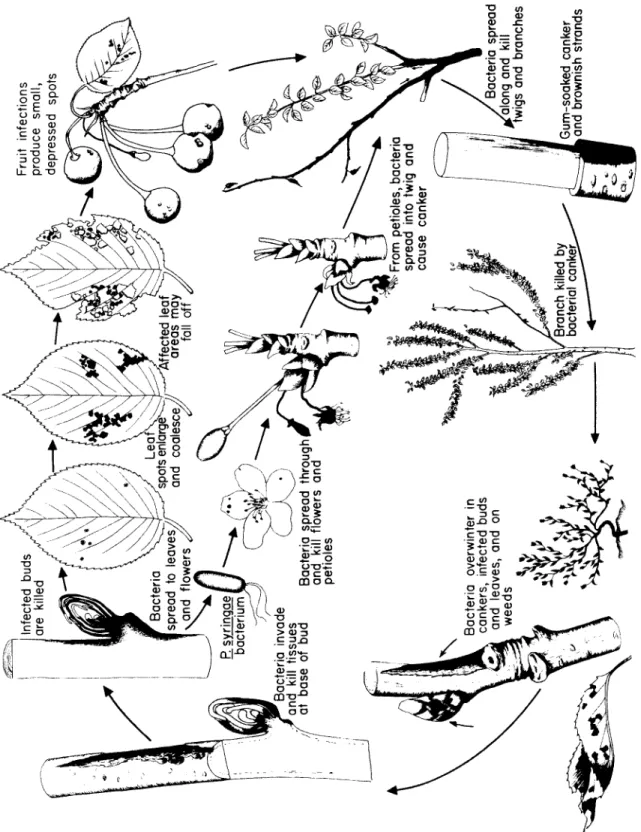
Fig. 59. Disease cycle of fire blight of pear and apple caused by Erwinia amulovora.
Youn g tre e heavil y ^ infecte d wit h fir e bliah t
Bacteri a overwinter
^in margin s o
f oldcanker s Canker s enlarg e an d girdl e branc h or ste m
Bacteri a i
n exudate ar e disseminate d by insect s an
d rain
Th e firebligh t bacteriu m
Bee s carr y bacteri a *
toflower s ^ Direc t infection
^of youn g twig s 1
Intercellula r multiplicatio n an
d ^spread of bacteria / ^^>O
nbar k Cell s o
f infected bar k tissu e collaps e Twi g killed
^by fir e bligh t
9Dea d leave s , clin g t o twi g
iFormatio n o
f newcanker s o
nbranche s an
dstem s
> /Extent and /direction of /spread of • bacteri
a Infectio n spread s t
o ? other flowers ,
1twigs,an d
Infecte d flower s shrive l
f/j\becom e dark-colore d O ;
and dieBacteri a penetrat e flower s throug h nectarthode s
j^andleave s throug h wound s o
r stomata. Bacteri a multipl y an
dsprea d intercellular ^
T h e bacteria multiply rapidly in the nectar, reach the nectarthodes and penetrate through them into the tissues of the flower. B e es visit- ing the nectar of such an infected flower carry bacteria from its nectar to all the succeeding blossoms that they visit. Once inside the flower, the bacteria multiply quickly in the subnectarthodal chamber.
Through enzymes they secrete, they break down the pectic sub- stances in the m i d d le lamella b e t w e en cell walls. T h e bacteria m o ve quickly, primarily through the intercellular spaces but also through the dissolved m i d d le lamella. T h e surrounding parenchymatous cells begin to show marked discoloration, plasmolysis, coagulation, and disintegration of the protoplasts. Sometimes the delicate walls of the flower cells are disrupted, and invasion of the protoplasts follows. Dis- integration of several layers of cells can take place in some cases. This results in fairly large-sized cavities filled with bacteria. From the re- ceptacle the bacteria m o ve intercellularly and through the phloem down the pedicel and into the bark of the fruit spur. Infection of the spur results in the death of all flowers, leaves, and fruit on it, regard- less of whether these have b e e n invaded by the bacteria.
Penetration and invasion of leaves, when it happens, is similar to that of flowers. Although stomata and hydathodes may serve as ports of entry for the bacteria, it s e e ms that most leaf infections take place through wounds m a de by insects, hailstorms, etc. Sucking insects, such as aphids and leafhoppers, s e em to be important not only as vec- tors of bacteria but also as inoculating agents b e c a u se they carry and insert the bacteria into freshly w o u n d ed tissues inside the leaf, where the bacteria are almost certain to develop. T h e bacteria s e em to de- velop better and faster in the spongy mesophyll than in the palisade parenchyma. From the vein parenchyma the bacteria pass into the petiole. T h e r e they invade the phloem, causing breakdown of cells and destruction of the cambium. T h e n they proceed into the xylem parenchyma and sometimes into xylem vessels. T h u s the bacteria may reach the stem through the petiole.
Young, tender twigs may b e infected by bacteria through their lenti- cels, through wounds m a de by various agents, and through sucking insects carrying bacteria and feeding on the twigs. T h e y may also b e infected through flower and leaf infections. In the twig the bacteria travel intercellularly. T h e y soon cause collapse and breakdown of cortical cells, forming large cavities. In young twigs the bacteria may reach the phloem, in which they then are carried upward to the tip of the twig and to the leaves. Invasion of large twigs and branches is re- stricted primarily to the cortex. Progress of the infection d e p e n ds on the succulence of the tissues and on the prevailing temperature and humidity. Under conditions adverse for the d e v e l o p m e nt of the patho-
Fire Blight
gen, the host may form cork layers around the infected area and may limit the expansion of the canker. In susceptible varieties and during warm, humid weather the bacteria may progress from spurs or shoots into the second-year, third-year, and older growth, killing the bark all along the way.
Control
Several m e a s u r es n e e d b e taken for a successful fire blight control program.
D u r i ng the winter all blighted twigs, branches, cankers, and even whole trees, if necessary, should b e cut out about 10 cm below the last point of visible infection and burned. Cutting of blighted twigs, suck- ers, and root sprouts in the summer can reduce the inoculum and pre- vent the production of large cankers on the branches supporting them.
But bacteria are in a very active state in the s u m m er and precautions should b e taken not to spread them to n e w branches or trees. Cutting should b e done about 30 cm below the point of visible infection. T h e tools should b e disinfested after each cut by b e i ng w i p ed with a sponge soaked in 1 0 % Clorox or in a solution of mercuric chloride and mercuric cyanide. T h e latter mixture can also b e u s ed to disinfect large cuts m a de by the removal of branches and cankers.
Since fire blight d e v e l o p m e nt is greatly favored by the p r e s e n ce of young, succulent tissues, certain cultural practices that favor moderate growth of trees are r e c o m m e n d e d. T h e se include: growing trees in sod, balanced fertilization, especially avoiding the overstimulation of growth by heavy nitrogen applications, and limited pruning. Also a good insect control program should b e followed to reduce or eliminate spread of bacteria by insects.
Although no pear or apple varieties are i m m u ne to fire blight when conditions are favorable and the pathogen is abundant, there is a marked difference b e t w e en the susceptibility of the varieties avail- able. In areas where fire blight is destructive, varieties for ne w plant- ings should b e chosen from those most resistant to fire blight.
Satisfactory control of fire blight with chemicals can b e obtained only in combination with the above-mentioned measures. Dormant sprays with copper sulfate (4 pounds to 100 gallons of water) before b ud break, or with Bordeaux mixture (12-12-100) containing 2 % misci- ble-type oil in the d e l a y ed dormant period offer some, but not much, protection from fire blight to a p p le trees. Bordeaux (2-6-100) or strep- tomycin at 50-100 parts per million (ppm) are the only blossom sprays effective. Bordeaux should be a p p l i ed during quick drying conditions to avoid possible russeting of fruit. Streptomycin acts systemically and
341
should be applied either when maximum temperatures are above 18°
or during the night, both conditions favoring absorption of streptomy- cin by the tissues. O ne to four streptomycin applications may b e nec - essary for satisfactory control of b l o s s om blight. Bordeaux, streptomy- cin, or zineb are sometimes u s ed to control twig blight on bearing and nonbearing trees, but none of them gives good control of this phase of the disease.
Selected References
B u s h o n g, J. W., D. Powell, a nd P. D. S h a w. 1964. Influence of c o p p er gradients upon various a p p le leaf a nd twig constituents as related to fire blight i n c i d e n c e. Phyto- pathology 54: 713-717.
Haber, J u l ia M. 1928. T h e relationship b e t w e en Bacillus amylovorus a nd leaf tissues of the a p p l e. Penn. State College Bull. 228: 15 p p.
H i l d e b r a n d, Å. M. 1939. S t u d i es on fire-blight ooze. Phytopathology 29: 142-156.
J o n e s, D. H. 1909. Bacterial blight of a p p l e, pear a nd q u i n ce trees. Ontario Agr. College Bull. 176: 64 p p.
Keil, H. L., B. C. S m a l e, a nd R. A. Wilson. 1966. Role of injury a nd longevity of Erwinia amylovora in the e p i d e m i o l o gy of fire blight of pear. Phytopathology 56: 4 6 4 - 4 6 5 . L e w i s, Sally, a nd R. N. G o o d m a n. 1965. M o de of penetration a nd m o v e m e nt of fire
blight bacteria in a p p le leaf a nd stem tissue. Phytopathology 55 : 7 1 9 - 7 2 3 . L u e p s c h e n, N. S. 1960. F i re blight control with streptomycin, as influenced by tempera-
ture a nd other environmental factors a nd by adjuvants a d d ed to sprays. Cornell Univ. Agr. Expt. Sta. Mem. 375 , 39 p p.
Parker, K. G., N. S. L u e p s c h e n, a nd E . G. F i s h e r. 1961 . T r e e nutrition a nd fire blight d e v e l o p m e n t. Phytopathology 5 1 : 5 5 7 - 5 6 0 .
Powell, D. 1966. F i g h t i ng fire blight in 66. Am. Fruit Grower 86: 3 6 - 3 8 .
R o s e n, H. R. 1929. T h e life history of the fire blight pathogen, Bacillus amylovorus, as related to the m e a ns of overwintering and dissemination. Arkansas Univ.
(Fayetteville) Agr. Expt. Sta. Bull. 2 2 4 , 9 6 p p.
R o s e n, H. R. 1936. M o de of penetration a nd of p r o g r e s s i ve invasion of fire-blight b a c t e- ria into a p p le and pear b l o s s o m s. Arkansas Univ. (Fayetteville) Agr. Expt. Sta. Bull.
3 3 1 : 6 8 p p.
Stewart. 1913. T h e fire blight d i s e a se in nursery stock. Cornell Univ. Agr. Expt. Sta.
Bull. 329: 3 1 3 - 3 7 2 .
Voros, J., a nd R. N. G o o d m a n. 1965. F i l a m e n t o us forms of Erwinia amylovora. Phyto- pathology 55 : 8 7 6 - 8 7 9 .
Crown Gall
Occurrence and Importance
Crown gall has b e e n reported from most countries of the world and is worldwide in distribution. It affects many woody and herbaceous
Crown Gall 343
plants belonging to 140 gener a of more than 60 families. In nature it is found mostly on p o me and stone fruit trees, brambles, and grapes, but several others are sometimes found affected and many more can b e artificially infected.
Crown gall is characterized by the formation of tumors or galls of varying size and form. It is c o m m on on the roots and shoots of various nursery plants which are thus unsalable b e c a u se crown gall is likely to continue on the plants w h en they are removed to orchards and gar- dens. Plants with tumors at their crowns or on their main roots grow poorly and produce r e d u c ed yields. Severely infected plants or vines may die.
Crown gall tumors have certain similarities to human and animal cancers and, therefore, the c a u se and mechanism of their formation have b e e n extensively studied. In spite of the apparent similarities to cancer, however, there are many and basic differences b e t w e en crown gall of plants and malignant tumors of man and animals.
Symptoms
T h e d i s e a se first appears as small overgrowths or excrescences on the stem and roots, particularly near the soil line. In early stages of their d e v e l o p m e nt the tumors are more or less spherical, white or flesh-colored, and quite soft. Since they originate in a wound, at first they cannot b e distinguished from callus. However, they usually de- velop more rapidly than callus. As the tumors enlarge, their surfaces b e c o me more or less convoluted. Later on, the outer tissues b e c o me dark brown or black, d ue to the death and decay of the peripheral cells (Fig. 60). S o m e t i m es there is no distinct line of demarcation b e t w e en the tumor and the plant proper, the tumor appearing as an irregular swelling of the tissues and surrounding the stem or root. Almost as often, however, the tumor lies outside but close to the outer surface of the host, b e i ng connected only by a narrow nec k of tissue. S o me tu- mors are spongy throughout and may crumble or b e c o me detached from the plant. Others b e c o me m u ch more woody and harder, looking knobby or knotty, and reaching sizes up to 30 c m in diameter. S o me tumors rot partially or completely from the surface toward the center in the fall and d e v e l op again in the s a me places during the next grow- ing season; or part of the tumor may rot while n e w tumor centers ap- pear in other parts of the s a me overgrowth.
Tumors are most common on the roots and stem near the soil line but they can also appear on vines u p to 150 cm from the ground, on branches of trees, on petioles, and on leaf veins. Several galls may occur on the same root or stem, continuous or in bunches, along the
F i g. 60. C r o wn gall tumors on stems of rose plants infected with Agrobacterium tume- faciens. (Photo by courtesy of the D e p a r t m e nt of Plant Pathology, Cornell Univer-
sity.)
same or different lines parallel to the length of the plant organs. On some hosts secondary tumors d e v e l op at points distant from the pri- mary tumor and are not connected with it. Secondary tumors are free from bacteria and d e v e l op in their absence.
In addition to forming galls, affected plants may b e c o me stunted, they produce small, chlorotic leaves and in general are more suscepti- ble to adverse environmental conditions, especially to winter injury.
The Pathogen: Agrobacterium tumefaciens
T h e crown gall pathogen is a rod-shaped bacterium about 1-3 ì long by 0.4-0.8 ì in diameter. It occurs singly or in short chains and has 2-4 flagella at the s a me pole. It forms a capsule. On certain m e d ia A. tumefaciens forms star-shaped arrangements which have b e e n con- sidered as stages in the sexual processes of these bacteria by which
Crown Gall 345
recombination and segregation of nuclear material takes place. Var- ious strains of this bacterium are known, and they exhibit different degrees of virulence. T h e virulence of each strain, however, s e e ms to be quite stable and is not r e d u c ed appreciably by repeated transfers of the bacteria in culture media. T h e pathogen is sensitive to sunlight and drying, but in soils with sufficient moisture it remains viable and virulent for many years.
T h e most characteristic property of this bacterium is its ability to transform normal plant cells to tumor cells in short periods of time.
Once the transformation to tumor cells has b e e n completed, these cells b e c o me i n d e p e n d e nt of the bacterium and continue to grow and divide abnormally even in the a b s e n ce of the bacteria.
Development of Disease
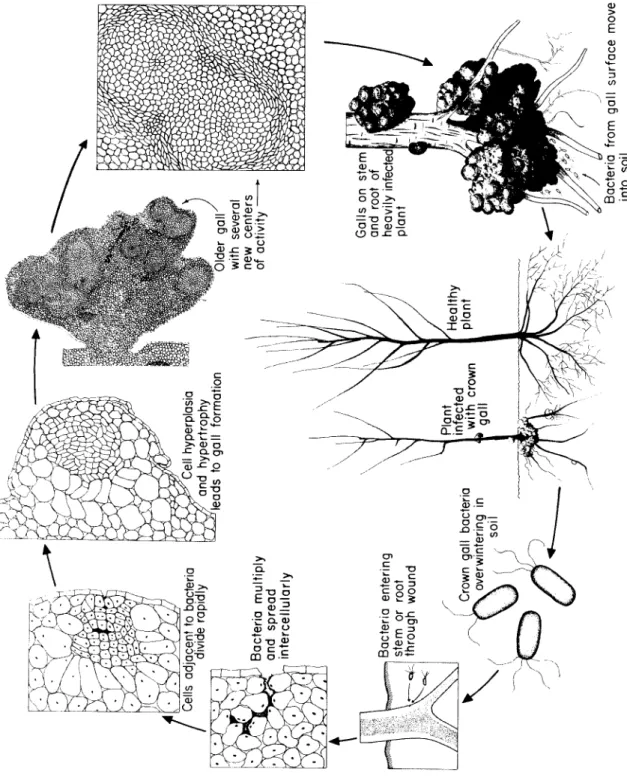
T h e bacterium overwinters in infested soils, where it can live as a saprophyte for several years. Whe n plants of any of the many suscepti- ble hosts are growing in such infested soils, the bacterium enters the roots or stems near the ground through fairly recen t wounds m a de by cultural practices, grafting, insects, etc. Once inside the tissue the bac- teria occur primarily intercellularly and stimulate the surrounding cells to divide (Fig. 61). O ne or more groups or whorls of hyperplastic cells appear in the cortex or in the cambial layer d e p e n d i ng on the depth of the wound. T h e se cells may contain one to several nuclei.
T h e y divide at a very fast rate, producing cells that show no differen- tiation or orientation. T h e cells continue to divide and to elongate, so that 10-14 days after inoculation a small swelling can b e seen with the naked eye. As the irregular division and enlargement of the cells con- tinue unchecked, the swelling enlarges, d e v e l o p i ng into a young tumor. Bacteria are absent from the center of the tumors but can be found intercellularly in their periphery. By this time certain cells have differentiated into vessels or tracheids which, however, are unorga- nized. T h e y occur singly or in strands a m o ng hyperplastic cells, unor- ganized, without regular orientation and with little or no connection with the vascular system of the host plant. As the tumor cells increase in n u m b er and size they exert pressure on the surrounding and under- lying normal tissues which may b e c o me distorted or crushed. Crush- ing of xylem vessels by tumors sometimes reduces the amount of wa- ter reaching the upper parts of a plant to as little as 2 0 % of the normal.
T h e smooth and soft young tumors are not protected by epidermis or other hard tissues. Therefore, they are easily injured and attacked by insects and saprophytic microorganisms. T h e se secondary invaders
Fig. 61. Disease cycle of crown gall caused by Agrobacterium tumefaciens.
Bacteria from gall surface mov into soil Galls on stem and root of J heavily infected! plant f Healthy plant Plant infected with crown 1 gall Crown gall bacteria \ overwintering in \ / , soil
Bacteria entering stem or root through wound Bacteria multiply and spread intercellularly
3ells adjacent to bacteria divide rapidly A^Xell hyperplasia j/and hypertrophy leads to gall formation Older gair with several new centers of activity
Crown Gall 347
cause decay and discoloration of the peripheral cell layers of the tu- mors, which turn brown to black. Breakdown of the peripheral tumor tissues releases crown gall bacteria into the soil, where they can b e carried in the water and infect n e w plants.
As the tumors enlarge they sometimes b e c o me woody and hard.
Finally, these consist chiefly of twisted, dislocated, and lignified vas- cular b u n d l es and woody fibers m i n g l ed with more or less paren- chyma. At other times, however, the tumors consist mostly of rapidly proliferating, round or spindle-shaped parenchymatous cells and rela- tively few, abnormally thin-walled, more or less lignified, disoriented vascular bundles. T h e reduction in volume of the normal xylem ves- sels limits the amount of nutrients and water that can reach the tumor tissues. T h e incomplete and disarranged vascular b u n d l es that may be present in the tumor itself are ineffective. Whe n tumors are unable to obtain as much water and nourishment as is required to carry them beyond a certain point in growth, their enlargement stops, decay sets in, and the necrotic tissues are sloughed off. In s o me cases the tumor regresses and no ne w one appears. More often, however, some portion of the tumor remains alive and forms additional tumor tissue during the s a me or the following season.
Whe n very young and expanding tissues are infected, in addition to the primary tumor which d e v e l o ps at the point of infection, secondary tumors appear. T h e se usually form below, but often above, the pri- mary tumor and at varying distances from it. Sometimes the secondary tumors d e v e l op at the scars of fallen leaves or at wounds m a de by var- ious agents. At other times secondary tumors d e v e l op on apparently u n w o u n d ed parts of the stem, on the petiole, and even on leaf midribs or larger veins several internodes a b o ve the primary tumor. Their starting point s e e ms to b e in the xylem of the vascular b u n d l e s. In sev- eral cases secondary tumors s e em to b e connected with the primary tumors by "tumor strands,, , that is by whorls or cells seemingly in a hyperplastic condition. In other cases they can b e explained as the result either of contamination of w o u n d ed tissues with bacteria through the surface or of spread of bacteria inside through the xylem vessels. S o me secondary tumors, however, are m u ch too far from the primary tumor for the two to have b e e n connected with tumor strands.
T h e se form on u n w o u n d ed tissue and, most important of all, they are free from bacteria, since no bacteria can b e isolated when these tu- mors are plated on nutrient media. Whe n fragments of such bacteria- free tumors are grafted on healthy plants, they d e v e l op into large tu- mors similar in appearance and structure to the primary tumors but remain completely devoid of bacteria. This indicates that the bacteria