Chapter 9
NITROGEN METABOLISM
J. W. MILLBANKAND K. A. KERSHAW
I. Introduction 289 II. Chemical and Biochemical Analyses of Whole Thalli 290
A. Total Nitrogen Content 290 B. Intracellular Nitrogenous Compounds 291
III. Assimilation of Combined Nitrogen 291 IV. Fixation of Elementary Nitrogen 297
A. Release of Fixed Nitrogen by the Phycobiont 298 B. The Rate of Nitrogen Fixation of Lichens 300 C. The Effect of Environmental Factors, Moisture, Light, and
Temperature on Nitrogen Fixation 301 V. Translocation of Nitrogen Compounds between the Symbionts 303
VI. The Relationship of Lichens to other Nitrogen-Fixing Systems 303
References 305
I. Introduction
Metabolic studies of the intact thallus have been largely neglected by biologists probably because of the supposed biological insignificance of lichens as well as the belief that they are difficult to use experimentally and that reliable and consistent results are hard to obtain. Nitrogen metabolism has suffered particularly because of the absence of a suitable radioactive isotope. The combination of slow growth (and, therefore, presumed sluggish metabolism) with the cumbersome and not very sensitive heavy isotope technique was a particularly discouraging situation. It now seems that lichens have, in some cases, surprisingly active metabolic processes and outstanding powers of recovery after periods of severe environmental stress.
Much recent work on nitrogen metabolism has resulted from the availability of the acetylene reduction technique for nitrogenase assay, a much more straightforward procedure. Only those features of nitrogen metabolism peculiar or relevant to the lichen mode of life will be considered in this chapter. This will comprise the absorption and metabolism of nitrogen
289
290 J. W . M I L L B A N K A N D K. A . K E R S H A W
by the intact thallus, together with aspects of the physiology of the isolated symbionts where they are important in an understanding of the events in the thallus.
II. Chemical and Biochemical Analyses of Whole Thalli
A. Total Nitrogen Content
Those reports of thallus total nitrogen which are available are summarized in Table I. Lichens with a blue-green phycobiont have significantly higher
T A B L E I
T H E TOTAL N I T R O G E N CONTENT OF LICHEN THALLUS
Nitrogen content
Lichen (% dry weight) Reference
Lichina confinis0 3.6-3.9 Hitch (1971)
Lobaria laetevirens0 2.2 Goas and Bernard (1967)
Lobaria pulmonariaab 2.7 Goas and Bernard (1967)
Peltigera aphthosa 1.9-3.4 Hitch (1971)
var. leucophlebia00
Peltigera aphthosa 3.0 Millbank and Kershaw (1969)
var. leucophlebiaa,c
Peltigera canina° 3.3 Millbank (1972)
Peltigera polydactyla0 3.6-4.5 Smith (1960a)
Peltigera praetextataa 4.7 Scott (1956)
Placopsis gelida° 0.9-1.3 Hitch (1971)
Sticta sylvatica" 4.0 Goas and Bernard (1967)
Anaptychia fusca 0.90 Hitch (1971)
Candelariella corallizza 4.2-5.0 Masse (1966)
Cladonia foliacea 0.65 Hitch (1971)
Cladonia impexa 0.33 Hitch (1971)
Cornicularia aculeata 0.38 Hitch (1971)
Evernia prunastri 0.84 Hitch (1971)
Lecanora atra 0.69 Hitch (1971)
Lecanora muralis 6.2-9.24 Masse (1966)
Ochrolechia parella 0.61-0.70 Hitch (1971)
Parmelia physodes 0.49 Hitch (1971)
Parmelia sulcata 0.96 Hitch (1971)
Physcia ascendens 1.0-1.3 Hitch (1971)
Physcia tribacia 3.9^1.7 Masse (1966)
Ramalina siliquosa 0.93 Hitch (1971)
Usnea subfloridana 0.58 Hitch (1971)
Xanthoria candelaria 4.2^1.4 Masse (1966)
Xanthoria parietina 1.21-1.71 Hitch (1971)
°Phycobiont a member of the Cyanophyceae.
b Primary phycobiont Myrmecia, Nostoc in cephalodia.
cPrimary phycobiont Coccomyxa, Nostoc in cephalodia.
9. N I T R O G E N M E T A B O L I S M 291 nitrogen contents than lichens with green phycobionts. Exceptions are those individuals that grow on substrata rich in nitrogen, e.g., Candelariella coralliza (Masso, 1966). This is not surprising, but the supposition that these higher nitrogen contents have a direct bearing on the growth rate and thallus size must be viewed with caution. The fact that Peltigera aphthosa benefits greatly from a supply of combined nitrogen (Kershaw and Millbank, 1970) is a case in point. Similarly, the large size of some Parmelias (e.g., Amphigym- nia) and Umbilicarias, which do not have blue-green phycobionts, points to a more complex control of growth rate.
B. Intracellular Nitrogenous Compounds
Goas and Bernard (1967) analyzed the soluble nitrogen fraction of Sticta sylvatica, Lobaria laetevirens, and L. pulmonaria. Glutamic acid was by far the most abundant amino acid found in all cases. The authors later reported the presence of glutamic decarboxylase and three transaminases concerned with the metabolism of glutamic acid (Bernard, 1969; Bernard and Goas, 1969). Glutamic-oxaloacetic transaminase (GOT) was the most active in all cases. Optimal conditions for activity of all the enzymes were established and GOT was assayed in five lichens, i.e., the three mentioned above plus
Sticta fulginosa and S. limbata. Ramakrishnan and Subramanian (1964a,b, 1965, 1966a,b) have made qualitative investigations of the amino acids in the soluble nitrogen fraction and hydrolysates of the following lichens:
Cladonia rangiformis, C. gracilis, Dermatocarpon moulinsii, Lobaria isidiosa, L. subisidiosa, Parmelia nepalensis, P. tinctorum, Peltigera canina, Ramalina sinensis, Roccella montagnei, Umbilicaria pustulata, Usnea flexilis, U. orien- talis, and U. venosa. Fabian-Galan et al (1966) reported the synthesis of several amino acids during photosynthesis in species of Cladonia, Collema, and Parmelia.
Methylamines were found to be abundant in the thallus of Lobaria laeti-
virens (Bernard and Goas, 1968), and Bernard and Larher (1971) demon- strated the important position of glycine in the biosynthesis of mono-, di-, and trimethylamine by this lichen. These authors have proposed a metabolic scheme that summarizes the interrelationships of glycine, the methylamines, sarcosine, and choline and studies are in progress to confirm their proposal.
III. Assimilation of Combined Nitrogen
Most lichens live in barren habitats where their nutrient supply is poor.
Thus, absorption from any solution in contact with the thallus is an im- portant aspect of their physiology and the rates observed are extremely rapid. This is reflected in the high rates of accumulation of radioactive material from atmospheric "fallout" (Gorham, 1959) and equally in that lichens are particularly sensitive indicators of toxic atmospheric pollutants
292 J. W . M I L L B A N K A N D K. A . K E R S H A W
(Hawksworth and Rose, 1970). Ahmadjian (1966) summarized the sparse early work which indicated that due to seasonal low rates of photosynthesis, the organic nutrient requirements of the thallus are quite unable to be met by the phycobiont, and suggested that the deficiency had to be met by direct absorption from the substratum or from liquids in contact with the thallus surface. While this may well be true in many instances, recent work by Lange, Schulze, and Koch (1970) has shown that Ramalina maciformis can achieve a net annual gain in assimilated carbon sufficient for a thallus growth of 5-10%.
In certain instances organic nitrogen derived from the older decaying parts of the thallus may be mineralized by epiphytic microorganisms and the derived ammonium ions absorbed by the remainder of the thallus, giving rise to a circulation of nitrogen (Shields et al., 1957). This concept is supported by the observation that some lichens secrete extracellular enzymes that act upon nitrogenous components of their substrate (Galinou, 1956; Mois- sejeva, 1961). Of the enzymes reported, asparaginase, allantoicase, allan- toinase, "lichenase," and uricase all have nitrogenous substrates. The above authors also reported the secretion of enzymes with polysaccharide or other carbohydrate substrates that could promote the availability of nitro
genous components of decaying plant and animal tissue.
Masse investigated the total nitrogen content (1966) and uricase activity (1969) in several lichens, including species that colonize habitats rich in bird excrement. Uricase was not found in the four lichens with blue-green phyco
bionts and the activity was variable in the others. There was not a clear pattern of activity between "ornithocoprophiles" and "ornithocopro- phobes," but Candelariella, a characteristic coprophile, had a very active uricase. The level of uricase among the coprophobes was generally feeble or zero, but there were exceptions. Ornithocoprophilic lichens with green alga phycobionts had exceptionally high nitrogen contents (Table I), presumably because of the high Ν content of the substrate.
Recent investigations on the uptake and assimilation of combined nitro
gen are reported by Smith (1960a,b) using disks of Peltigera polydactyla. The technique used was to analyze 7-mm disks of thallus for nitrate, nitrite, ammonia, amide, α-amino and total nitrogen. Many disks (up to 450) were used in each experiment and this reduced considerably any metabolic varia
tion that would occur among different thallus specimens. Soluble nitrogen formed about a quarter of the total and of this soluble fraction about 25%
was ammonia, 25% α-amino, 3-5% amide, and the remainder was uncharac- terized. The amounts of nitrate and nitrite nitrogen were negligible.
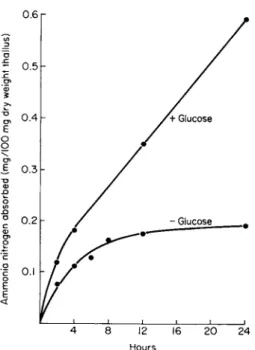
When the disks were shaken in ammonium chloride solution, with and without glucose, some striking effects were observed (Fig. 1). Without glucose, absorption virtually ceased after 12 hours; but in the presence of
9. N I T R O G E N M E T A B O L I S M 293
Hours
FIG . 1. Ammonia absorption by Peltigera polydactyla thallus in 5 mM N H4 CI with and with- out 14 mM glucose. (After Smith, 1960a.)
glucose the enhanced rate of uptake continued throughout the experimental period. The absorbed nitrogen produced large increases in the ammonia and amino nitrogen content of the disks, and in the presence of glucose the increases were about threefold greater. There was no significant change in the amide fraction and amides did not seem to be compounds of metabolic importance. The difference between these results and the effects of ammonia uptake on free-living fungi was particularly notable. Thus, there was almost no shift of nitrogen from the soluble to the insoluble fraction in
Peltigera, even in the presence of glucose, and evidently protein synthesis was not stimulated. About 90% of the ammonia uptake was accounted for in the soluble fractions, even in the presence of glucose. Protein synthesis was apparently limited by factors other than carbohydrate supply.
When disks were incubated in water or buffer without nitrogen for 6 days, the soluble nitrogen fraction of the thallus increased slightly, equivalent to about 5-10% of the total protein nitrogen. Protein breakdown seemed minimal, unlike what is observed in higher plants. The presence of glucose in the medium reduced the level of thallus ammonia nitrogen, indicating a promotion of ammonia utilization. Release of ammonia to the medium was never observed with or without the presence of glucose in the medium.
294 J. W. MILLBANK AND K. A. KERSHAW
The second part of Smith's investigation (1960b) dealt with the uptake and utilization of the acid amides asparagine and glutamine and the corre
sponding amino acids, aspartic and glutamic. This study revealed that as
paragine was absorbed more rapidly and in larger amounts than the other substances, which is difficult to explain since the earlier study found that acid amides apparently were of only slight importance in the nitrogen metabolism of the disks.
The absorption from asparagine solutions by disks of Peltigera over 24 hours is shown in Fig. 2. Absorption was rapid and ranged from 15 to 22.5 μ% N/mg dry weight thallus/24 hours; ammonia was released into the medium, and it may be assumed that asparagine entered the disks as the complete molecule since both amide and amino nitrogen diminished equally.
The evidence for this assumption was a trial in which equimolar mixtures of aspartic acid and ammonia were assimilated much more slowly than an equivalent amount of asparagine. The asparagine appeared to be stored upon absorption, since large increases in amide nitrogen, roughly equiv
alent to the losses from solution, were demonstrated. Analyses after ab
sorption showed that the rate of assimilation was very slow and at least 50%
3 r
Hours
FIG. 2. Changes in the nitrogen fractions of 5 mM asparagine solutions during absorption by disks of Peltigera polydactyla thallus at 20° C. (After Smith, 1960b.)
9. N I T R O G E N M E T A B O L I S M 295 of the amide nitrogen originally absorbed still remained after 72 hours.
Synthesis of complex soluble or insoluble compounds during this period was negligible. The only apparent conversion that occurred was deamidation to ammonia; some deamination may also have taken place but this was not supported by experimental evidence. After absorption the asparagine could not be washed out. Smith considered that absorption was an active process since it was inhibited by 25 mM sodium fluoride and accompanied by an increase in the respiration rate approximately equal to that caused by an equivalent number of glucose molecules.
Glucose inhibited the absorption of asparagine, an effect that is opposite to that found in free-living fungi. When free-living fungi were incubated in asparagine media without glucose, deamidation was promoted, i.e., amide nitrogen rapidly diminished, and large amounts of ammonia were released. The amino nitrogen diminished slowly. The addition of glucose to the solution increased the rate of utilization of both amino nitrogen and the released ammonia. Thus, the effect of asparagine plus glucose caused changes in the nitrogen fractions of free-living fungi similar to those brought about in Peltigera by asparagine in the absence of glucose. These enigmatic findings, together with the observations that not only was asparagine ab- sorption depressed in the presence of glucose but glucose absorption was depressed in the presence of asparagine, may indicate two uptake mech- anisms that are controlled by an unknown common limiting factor. Glucose not only retarded the rate of entry of asparagine but promoted deamidation of that which did get in and also promoted the conversion of ammonia to amino nitrogen and thus retarded its loss to the medium by leakage.
Glutamine and glutamic and aspartic acids were absorbed much more slowly than asparagine (Table II). In all cases, ammonia was released into the medium and the disks also showed significant increases in ammonia—and amino nitrogen. It appeared that glutamine, unlike asparagine, was readily deamidated by the thallus tissue and some of this deamidation was extracel- lular. Glutamic acid was absorbed at a similar rate to glutamine and aspartic acid least rapidly.
Although rates of absorption are high relative to the slow rates of growth the utilization of the absorbed material is consistent with the slow growth observed.
Smith has also shown (1961) a seasonal variation in the rate of asparagine absorption by Peltigera polydactyla as well as the total thallus nitrogen content. There was a period of maximum metabolic activity in late winter and early spring when field conditions were moist and cool, and the light intensity was high. The carbohydrate and nitrogen reserves built up during this period were depleted in the summer when conditions for assimilation
296 J. W . M I L L B A N K A N D K. A . K E R S H A W T A B L E I I
C H A N G E S IN THE N I T R O G E N FRACTIONS OF THE INCUBATION MEDIA A N D OF THE THALLUS OF Peltigera polydactyla
AFTER INCUBATION FOR 24 HOURS AT 20° C A N D AT p H 5.7*
Nitrogen Nitrogen source (^g N / 1 0 0 mg dry weight of thallus) fraction
Asparagine Aspartic acid Glutamine Glutamic acid Amide
Medium - 1 0 0 4 0 - 5 2 4 0
Thallus + 669 - 1 0 - 2 8 - 3
Amino
Medium - 1 0 0 4 - 4 9 0 - 8 7 9 - 1 0 4 2 Thallus + 874 + 314 + 492 + 493 Ammonia
Medium + 95 + 74 + 469 + 79
Thallus + 202 + 67 + 322 + 239
flData from Smith (1960b).
were poor. The poor conditions persisted until autumn when a short favour- able period recurred. During the winter the lichens became covered by litter and again drew on their reserves. Asparagine-absorbing activity fol- lowed a similar pattern, as did nitrogen content, but the latter lagged behind the other parameters, the seasonal effect being delayed.
This work by Smith has been described in detail since it was the first and is still the only detailed study of the uptake and utilization of combined nitrogen by lichens. It was also the first physiological study in which a serious attempt was made to overcome variability problems, and in which detailed chemical analysis formed a major part.
Of the findings, the most significant were that the rapid uptake of com- bined nitrogen was not followed by protein synthesis, that amide metabolism is of minor importance, and that protein breakdown in periods of starvation is slight. Whether algal products are the direct cause of the major changes in the physiology of the "lichenized" fungus is a problem that will only be resolved when methods are available for studying the isolated symbionts in an undamaged condition and for the in situ demonstration of metabolites.
At present, cultures of lichen fungi and blue-green algae are necessarily considerably removed from the "symbiotic" condition, having undergone several generations of independent growth.
Nevertheless, Smith's overall findings are clear enough—a biological system has developed in which vigorous powers of nutrient and moisture absorption without special organs have been allied to restrained abilities for synthesis and growth. Thus, every opportunity for absorption of food
9 . N I T R O G E N M E T A B O L I S M 297 material of virtually any kind can be taken, and a steady rate of utilization maintained. This is of great biological advantage in exacting and inhospi- table habitats.
IV. Fixation of Elementary Nitrogen
Nitrogen fixation has been demonstrated only in those lichens with blue- green phycobionts. Of the eight genera of blue-green algae listed by Ahmad- jian (1967) and Duncan (1970) as phycobionts, only lichens with Nostoc and Calothrix have so far been shown to fix nitrogen. The lichens in which fixation has been shown either by means of 1 5N2 or the acetylene reduction technique (Dilworth, 1966; Hardy et aL, 1968) are listed in Table III. The phycobionts, of Peltigera virescens (Watanabe and Kiyohara, 1963) and Collema tenax
(Henriksson, 1951), have been shown to fix nitrogen when isolated and fixation can be assumed to take place in the lichen. The first definitive demonstration of fixation by an intact lichen thallus was by Bond and Scott (1955) who used 1 5N2 and tested Collemagranosum and Leptogium lichenoides.
T A B L E I I I
ESTABLISHED N I T R O G E N - F I X I N G LICHEN SPECIES
Tech- Phycobiont nique
Lichen genus used Reference
Collema coccophorus Nostoc 1 5N2 Rogers et al (1966)
Collema crispum Nostoc Q H2 Hitch (1971)
Collema granosum Nostoc 1 5N2 Bond and Scott (1955) Collema pulposum Nostoc , 5N2 Fogg and Stewart (1968)
Collema tunaeforme Nostoc Henriksson and Simu (1971)
Leptogium lichenoides Nostoc , 5N2 Bond and Scott (1955) Lichina confinis Calothrix 1 5N2 Stewart (1970) Lichina pygmaea Calothrix , 5N2 Stewart (1970)
Lobaria pulmonaria Nostoc (cephalodia) Q H2 Millbank and Kershaw (1970) Myrmecia (thallus)
Peltigera aphthosa Nostoc (cephalodia) 1 5N2 Millbank and Kershaw (1969) var. leucophlebia Coccomyxa (thallus)
Peltigera canina Nostoc Q H2 Millbank and Kershaw (1970) Peltigera polydactyla Nostoc , 5N2 Watanabe and Kiyohara (1963) Peltigera praetextata Nostoc 1 5N2 Scott (1956)
Peltigera pruinosa Nostoc 1 5N2 Watanabe and Kiyohara (1963)
Peltigera rufescens Nostoc Henriksson and Simu (1971)
Placopsis gelida Stigonemal Nostoc CjH2 Hitch (1971)
Stereocaulon sp. Nostoc (cephalodia) , 5N2 Fogg and Stewart (1968) Trebouxia (thallus)
298 J. W. MILLBANK AND K. A. KERSHAW
The existence of nitrogen-fixing bacteria in or on lichens has been referred to frequently; Cengia-Sambo (1923) reported their presence in the cepha
lodia of Peltigera aphthosa and postulated the theory of poly symbiosis. The cephalodia were thought to be analagous to the root nodules of legumes by both Cengia-Sambo and Goebel (1926), although Goebel considered that the Nostoc in the cephalodia were responsible for nitrogen fixation. Henckel and Yuzhakova (1936) reported Azotobacter in epiphytic lichens and these findings were extended in 1938 by Henckel and Iskina, independently, to include other groups of lichens, including Cladonia, which was originally reported as not possessing Azotobacter. Scott (1956) refers to the unpublished findings of Dr. G. Metcalfe, in which Azotobacter was isolated in many British lichens. However, Walenkamp (referred to by Quispel, 1945) and Krasilnikov (1949) did not detect Azotobacter in the 40 species of lichens examined. More recently Panosyan and Nikogosyan (1966) failed to find
Azotobacter in a number of Armenian lichens they examined and considered this to be due to the lack of any respirable substrate. Bond and Scott (1955), while admitting that Azotobacter could have been present epiphytically on the thallus samples they studied, concluded that they made no significant contribution to the fixation observed. They based this conclusion on the lack of the necessary respirable substrate and the absence of anything approaching the number of bacterial cells needed.
In 1956, Scott reported on his studies with Peltigerapraetextata, extending his earlier work with Bond (Bond and Scott, 1955). Nitrogen fixation was taking place, brought about by the Nostoc phycobiont, any contribution by epiphytic Azotobacter being considered insignificant. He also showed that some of the nitrogen fixed by the Nostoc was transferred to the mycobiont.
Results of trials with Cladonia impexa were also reported. This lichen did not fix nitrogen, the phycobiont being a green alga, but as Azotobacter had frequently been associated with it, it was desirable to clear up the question unequivocally. More recent evidence, from electron micrography of Pel
tigera canina, showed the total absence of a third symbiont (Griffiths etal., 1972). Azotobacter can only be a casual epiphyte and the hypothesis of Cengia-Sambo (1923) must be rejected.
A. Release of Fixed Nitrogen by the Phycobiont
Henriksson (1951) showed that between 19 and 28% of the nitrogen fixed by the Nostoc phycobiont of Collema tenax was released into the growth medium. Release of Ν is a normal feature of the metabolism of blue-green algae (see, e.g., Fogg, 1962). She inferred that the same thing is happening within the lichen thallus, and this was confirmed by Scott (1956).
Millbank and Kershaw (1969) working with Peltigera aphthosa and using
1 5N2 showed that fixation in this lichen was confined to the cephalodia, which
9. N I T R O G E N M E T A B O L I S M 299
2 0 0 Γ
FIG. 3. (A) Uptake of elementary nitrogen by Peltigera aphthosa thallus at 1 2 ° C . (B)
1 5N2 enrichment of Peltigera aphthosa cephalodia at 1 2 ° C when exposed to nitrogen gas enriched to 30% 1 5N2. (After Millbank and Kershaw, 1969.)
contained the Nostoc phycobiont. Thallus from which the cephalodia had been removed by dissection showed no evidence of fixation. Experiments carried on for 25 days showed that incorporation of nitrogen from the at
mosphere into the thallus was linear (Fig. 3) but that the level of heavy nitrogen labeling in the cephalodia became constant after 10 days only.
It was concluded that there was a continuous throughput of nitrogen via the
Nostoc and that virtually all the nitrogen fixed by the Nostoc was being released to the mycobiont.
The authors, although considering the mycobiont to be controlling the metabolism of the nitrogen-fixing phycobiont, were not able to supply any evidence as to what form the control took. They tended to favor some form of chemical control of the alga, affecting either nitrogen utilization or permeability. As yet, no substances or extracts from lichen thalli have been demonstrated to influence the rate of nitrogen fixation or nitrogen loss from blue-green algal cells. The possibility that the effect is connected with the oxygen tension in the thallus is at least equally likely, and in view of later work to be described below, is at present favored by the writers.
300 J. W. MILLBANK AND K. A. KERSHAW B. The Rate of Nitrogen Fixation of Lichens
The studies referred to so far have mostly dealt with nitrogen fixation either qualitatively or on a "whole thallus" basis. Millbank and Kershaw (1969) attempted to relate the fixation of nitrogen in lichens to the popula
tion of the nitrogen-fixing symbiont. They found that the total nitrogen content in Peltigera aphthosa was of the order of 3% of the dry weight, and that the algal contribution to the total nitrogen was 7.5% in the thallus and 5.5%* in the cephalodia. Thus, the phycobiont was far less abundant than Scott reported (1956) for P. praetextata. Scott estimated the algal content by visual observation of sections, a simple and straightforward technique, but liable to rather large errors and uncertainties. Millbank and Kershaw further showed that the total nitrogen in the cephalodia represented only about 2-4% of that in the entire thallus, so clearly the Nostoc nitrogen formed a tiny fraction (5.5% of, say, 3%) of the total thallus nitrogen. Thus, although the total amount of nitrogen fixed per unit of thallus was small, the rate of fixation by the Nostoc was rapid and, had it been entirely used for growth, would have permitted a generation time of 18 hours* at 25° C and 24 hours* at 12°C, the latter a representative noon temperature for the natural habitat in spring. This compares with a figure of 19.5 hours for Nostoc muscorum in shake culture at 25° C given by Kratz and Myers (1955).
Later studies by Millbank (1972) on Peltigera canina, using the acetylene reduction technique for estimating nitrogenase activity, confirmed that fixation was rapid, at least in Peltigera. The Nostoc nitrogen was again found to be a small proportion of the total, about 2.7% (3 χ 106 cells/cm2 thallus).
The cells were found to be very much larger than when grown in the free- living state, having mean dimensions of 8.3 χ 6.7 μπι, compared with 4.0 χ 3.7 μπι. The rate of acetylene reduction was examined in a large number of specimens of thallus, and, as expected, was found to be rather variable. However, the most important finding was that the rate was very rapid. At 10 nmole evolved/minute/mg Nostoc protein the average rate was about three times as fast as was usual in free-living material.
Furthermore, heterocysts, although present, were rather infrequent, repre
senting about 3.3% of the total algal cells (Griffiths et al, 1972). This con
firmed Peat's observation (1968), and incidentally probably accounts for Drew and Smith's (1967) inability to observe any at all when they examined homogenized thallus under the light microscope.
If nitrogen fixation were confined to the heterocysts as has been recently supposed (Fay et al, 1968), their nitrogenase activity becomes equivalent to 300 nmoles Q H4 evolved/minute/mg heterocyst protein. Data on the
* Amended from the figures given in 1969 in the light of later work (Millbank, 1972).
9. N I T R O G E N M E T A B O L I S M 301 specific activity of pure nitrogenase are quoted by Hardy et al. (1971) and, taking a figure of 900 nmole/minute/mg as representative, the conclusion is that about 33% of the total heterocyst protein is nitrogenase. This seems improbable, and fixation in lichens may perhaps be taking place in the vegetative cells. This could well be feasible as a consequence of a possible low internal p02 brought about by fungal activity, and is currently under active investigation. If so, it accords with a general hypothesis concerning the site of nitrogen fixation proposed by Stewart (1971) and supported by the results of van Gorkom and Donze (1971).
Henriksson and Simu (1971) have expressed their results for nitrogenase activity in Collema tunaeforme and Peltigera rufescens on a quantitative basis, but unfortunately relative to the dry weight of the entire thallus. If a number of assumptions are made, the nitrogenase activity per milligram Nostoc protein can be shown to be of the same order as found by Millbank (1972).
It therefore seems that rapid rates of fixation are a characteristic of lichen
Nostoc under good conditions.
Study of the nitrogenase activity over the thallus area shows the most rapid rate of fixation, on both an area and a cell-number basis, to be taking place in the mature part of the thallus, about 3 cm behind the growing edge (Millbank, 1972).
C. The Effect of Environmental Factors, Moisture, Light, and Temperature on Nitrogen Fixation
From their study of ten tropical and nineteen arctic lichens, Scholander et al. (1952) concluded that "the metabolic rate of a lichen is highly variable, depending largely upon its water content and temperature." Their concern was respiration, but nitrogen fixation is as dependent, with light intensity an additional factor.
Stewart (1965) showed that Calothrix sp. and Nostoc sp. were capable of some dark fixation of 1 5N2 over a 24-hour period, but it was considerably less than in the light. He considered that nitrogen fixation was not critically light dependent as long as supplies of carbon skeletons, reducing power, and energy were available.
Studies of temperature effects are very sparse. Millbank and Kershaw (1969) reported that the fixation rate of Peltigera aphthosa under laboratory conditions exhibited a Ql0 of approximately 2 over the temperature range 12°-25°C; and Fogg and Stewart (1968) reported that nitrogen fixation by lichens under antarctic conditions "showed some relation to temperature."
They suggested that "although appreciable fixation occurs in the vicinity of 0° C, the rates increased rapidly with rise in temperature, and it is probable that the bulk of fixation is accomplished in brief periods when the micro- environment reaches temperatures of 10° C or more."
302 J. W . M I L L B A N K A N D K. A . K E R S H A W
Thallus moisture content is very critical, especially as the uptake and loss of water by lichen thalli is largely a physical process. The moisture content and rates of absorption and loss have been quite extensively studied (see Chapter 11). Henriksson and Simu (1971) reported the ability of Collema
tunaeforme and Peltigera rufescens to recover their nitrogenase activity after long periods of desiccation. Storage was for periods of up to 30 weeks in plastic containers in the dark, at 12°C. After storage for periods longer than 4 weeks, the nitrogenase activity upon rewetting was about 50% of that observed with material stored for 1 day only; but these rates were achieved after a recovery period of only 24 hours, and in the case of Collema a re- covery period of 1 hour only was sufficient to restore 50% of the original activity after 22 weeks of storage, and 25% after 30 weeks. These storage conditions were very exacting and show the lichen's remarkable capacity to withstand severe environmental stress.
Hitch (1971) studied the environmental factors affecting nitrogenase activity in lichens, and has reported diurnal and seasonal variation in
Collema, Lichina, and Peltigera. When samples collected from the field in a desiccated state were wetted, nitrogenase activity started after lag periods of 60, 20, and 35 minutes, respectively. The thallus moisture contents, how- ever, changed from the original level of 20% oven-dry weight to 200% within 5 minutes. In these lichens, when the moisture content fell below 80-90% of the oven-dry weight, acetylene reduction stopped. If these results hold true for other species, under field conditions with fast-drying rates of the thallus, little or no fixation may be possible in exposed sites and the significance of nitrogen fixation must be questioned.
Nitrogenase activity was maximal at light intensities above about 400 ft-c in Peltigera and Lichina. In the dark, there was a progressive loss of nitro- genase activity, the rate of the loss probably reflecting the size of the carbon reserves in the thallus. Under constant light intensity (700 ft-c) and saturated moisture content Lichina confinis showed optimum fixation at 20° C with lower and upper limits of —3° and 35°C, respectively. Peltigera rufescens
had a very similar temperature range, - 3° to 46° C with an optimum of 31 ° C.
The diurnal variation in fixation rate of Peltigera polydactyla and Lichina confinis was correlated directly with light intensity and temperature when the thallus was adequately moist. When long-term seasonal effects were con- sidered, the pattern of activity (measured at midday) was again related to temperature, but completely overridden by moisture content. Thus, in temperate northern latitudes, nitrogenase activity could be greater in February than in a dry May, and lowest of all in June to August.
These results all fit with what is beginning to emerge as the fundamental finding of laboratory and field studies, i.e., that metabolism of lichens is essentially opportunist; when conditions are right, absorption, assimilation, synthesis, or whatever the process may be becomes active for as long as
9. N I T R O G E N M E T A B O L I S M 303 possible and then reverts to a state of quiescence when conditions deteriorate.
Nitrogen fixation is no exception. If it is available to the lichen it is utilized;
but other sources of nitrogen are as avidly accepted and no one process is uniquely significant.
V. Translocation of Nitrogen Compounds between the Symbionts
Isolated lichen algae are known to release a variety of organic substances into their growth media; and it is assumed that this characteristic is continued in the symbiotic condition although there is little direct evidence so far.
Henriksson (1957,1960,1961) has shown that a Nostoc from Collema released about 5% of the nitrogen it fixed into the medium, together with thiamine, biotin, riboflavin, and nicotinic and pantothenic acids. It also released un- known compounds which inhibited the growth of its (cultured) mycobiont.
Bednar (1963) showed that the Coccomyxa phycobiont of Peltigera aphthosa
released biotin and thiamine into the medium. Henriksson again, in 1958, reported that an isolated strain of Nostoc released substances which can sup- port the growth of the mycobiont, though the nature of the substances was not established. Many lichen fungi are deficient in biotin and thiamine and the release of these compounds by the phycobiont could be of vital significance to the maintenance of healthy growth of the combination.
Kershaw and Millbank (1970) investigated the movement of 1 5N2 in the three component lichen Peltigera aphthosa over periods of up to 55 days, and found that the labeled nitrogen, which had been fixed and then released from the Nostoc in the cephalodia, was overwhelmingly present in the thallus mycobiont. After considering the proportions of fungal and green algal cells in the thallus, only about 3% of the expected amount of nitrogen was found in the green algal (Coccomyxa) cells. The lichen, when maintained in a suitable enclosure (Kershaw and Millbank, 1969), benefited greatly by the regular addition of combined nitrogen and it seemed that the cephalodia were of minimal value as a source of nitrogenous nutrient to the green algal component; the greater proportion of blue-green algae per unit of thallus in P. canina may well provide a more adequate supply of nitrogen, but the observation of Stewart (1966, p. 68) that the majority of lichens contain non-nitrogen-fixing Chlorophyceae as phycobionts and do as well on in- hospitable substrates as the nitrogen-fixing ones is nevertheless true.
VI. The Relationship of Lichens to Other Nitrogen-Fixing Symbiotic Systems
Scott (1969) arrived at a definition of symbiosis as a "state of equilibrated physiological interdependence of two or more organisms involving no per-
304 J. W . M I L L B A N K A N D Κ A . K E R S H A W
manent stimulation of defensive reaction mechanisms." This is a synthesis of six criteria, the possession of any four of which he considers adequate to constitute a symbiotic association. These criteria are that the association should (a) be a permanent feature of the organisms' life cycles; (b) involve physical contact between the participants; (c) involve unilateral or bilateral movement of metabolites; (d) ameliorate environmental status, thus giving rise to an extension of ecological range; (e) give rise to morphogenetic effects; and (f) provide opportunity for the production of metabolites not formed by either of the organisms separately.
The other nitrogen-fixing symbiotic systems comprise: the legume-/?A/z0- bium association; the nonleguminous angiosperm-root nodule systems (endophytes unidentified); the Klebsiella-Psychotria leaf nodule association;
and the relationship between blue-green algae with liverworts (Blasia, Cavi- cularia), ferns (Azolla), Gymnosperms (Cycas, Stangeria, Encephalatos, Macrozamia), and Angiosperms (Gunnera).
Since the endophyte(s) in the nonleguminous nodule system have not been identified as yet it is not possible to compare their physiology and attributes to the other systems. Leaving them on one side, therefore, it would seem that the nitrogen-fixing lichens are in one respect, much more closely related to the Blasia, Azolla, Cycas, and Gunnera association than to the strict nitrogen-fixing symbiosis as typified by the legume-Rhizobium system. Thus, although all the associations possess many or all of Scott's criteria, only the legume-Rhizobium association is a strict nitrogen-fixing symbiosis, since in all the other systems the process is carried on by an organism able to fix nitrogen in pure culture or free-living in nature. Fur
thermore, the nitrogen-fixing organism's faculty of release of combined nitrogen is also normal, and is enhanced rather than initiated by the spe
cialized environment. However, the morphological distinctiveness of lichens, in which there is little or no resemblance between the product of the combination and any of the constituent partners, is one of the most out
standing features of the symbiosis, and distinguishes it completely from the green-plant associations. We have evidently an intimate relationship giving rise to profound morphological and biochemical changes but retaining at least some of the fundamental biochemical attributes of the individual partner. Other biochemical features, such as the production of lichen acid and antibiotics and the release of glucose and other carbohydrates by the phycobionts are, however, apparently associated only with the lichenized state and thus qualifies the association under criterion (f) above as well.
Given that mutual advantages are features of a symbiotic relationship, it seems most appropriate to consider that the major benefit to the phyco
bionts) in the lichen association is a considerable extension of ecological range, for which they pay the price of greatly reduced rate of growth and what may be termed a state of controlled parasitism by the mycobiont.
9. N I T R O G E N M E T A B O L I S M 305
References
Ahmadjian, V. (1966). Lichens. In "Symbiosis" (S. M. Henry, ed.), Vol. 1, p. 35-97. Academic Press, N e w York.
Ahmadjian, V. (1967). A guide to the algae occurring as lichen symbionts, isolation, culture, cultural physiology and identification. Phycologia 6, 127-160.
Bednar, T. W. (1963). Physiological studies on the isolated components of the lichen Peltigera aphthosa. Ph.D. Thesis, University of Wisconsin, Madison.
Bernard, Τ (1969). Contribution a l'etude du metabolisme azote des lichens. Activite de la glutamate decarboxylase de cinq especes de la famille des Stictacees. C. R. Acad. Sci., Ser.
D 269, 823-826.
Bernard, T., and Goas, G. (1968). Contribution a l'etude du metabolisme azote des lichens.
Caracterisation et dosages des methylamines de quelques especes de la famille des Stictacees. C.R. Acad. Sci., Ser. D 267, 622-624.
Bernard, T., and Goas G. (1969). Contribution a l'etude de metabolisme azote des lichens. Mise en evidence de quelques transaminases; activite de la glutamate oxaloacetate transaminase dans cinq especes de la famille des Stictacees. C.R. Acad. Sci., Ser. D 269,1657-1660.
Bernard, T., and Larher, F. (1971). Contribution a l'etude du metabolisme azote des lichens.
Role de la glycine 1 4Q dans la formation des methylamines chez Lobaria laetevirens.
C.R. Acad. Sci., Ser. D 272, 568-571.
Bond, G. and Scott, G . D . (1955). An examination of some symbiotic systems for fixation of nitrogen. Ann. Bot. (London) [N.S.] 19, 6 7 - 7 7 .
Cengia-Sambo, M. (1923). Polysimbiosi nei licheni a cianoficee a significato biologico dei cefalodi. Atti Soc. Ital. Sci. Natur. Mus. Civ. Stor. Natur. Milano 62, 226-238.
Dilworth, M. (1966). Acetylene reduction by nitrogen fixing preparations from Clostridium pasteurianum. Biochim. Biophys. Acta 127, 285-294.
Drew, Ε. Α., and Smith, D . C. (1967). Studies in the physiology of lichens. VII. The physiology of the Nostoc symbiont of Peltigera polydactyla compared with cultured and free-living forms. New Phytol. 66, 379-388.
Duncan, U. (1970). "Introduction to British lichens," T. Buncle and Company, Arbroath.
Fabian-Galan, G., Atanasiu, L., and Salageanu, N. (1969). Organic substances produced by photosynthesis in lichens. Prog. Photosyn. Res. 3, 1553-1558.
Fay, P., Stewart, W. D . P., Walsby, A. E., and Fogg, G. E. (1968). Is the heterocyst the site of nitrogen fixation in blue green algae? Nature (London) 220, 810-812.
Fogg, G. E. (1962). Extracellular Products. In "Physiology and Biochemistry of Algae" (R. A.
Lewin, ed.), p. 475. Academic Press, N e w York.
Fogg, G. E., and Stewart, W. D . P. (1968). In situ determinations of biological nitrogen fixation in Antarctica. Brit. Antarctic Surv. Bull. 15, 39-46.
Galinou, M. A. (1956). Sur la mise en evidence de quelques biocatalyseurs chez les lichens.
Proc. Int. Bot. Congr., 8th, 1954 Sect. XVIII, pp. 2-Λ.
Goas, G., and Bernard, T. (1967). Contribution a l'etude du metabolisme azote des lichens:
Les differentes formes d'azote de quelques especes de la famille des Stictacees. C. R.
Acad. Sci., Ser. D 265, 1187-1190.
Goebel, K. (1926). Ein Beitrag zur Biologie der Flechten. Ann. Jard. Bot. Buitenz. 36, 1.
Gorham, E. (1959). A comparison of lower and higher plants as accumulators of radioactive fallout. Can J. Bot. 37, 327-329.
Griffiths, Η. B., Greenwood, A. D., and Millbank, J. W. (1972). The frequency of heterocysts in the Nostoc phycobiont of the lichen Peltigera canina Willd. New Phytol. 71, 11-13.
Hardy, R. W. F., Holsten, R. D., Jackson, Ε. K., and Burns, R. C. (1968). The acetylene ethylene assay for nitrogen fixation. Laboratory and field evaluation. Plant Physiol. 43, 1185-1207.
306 J. W. MILLBANK AND K. A. KERSHAW Hardy, R. W. F., Burns, R. C. Hebert, R. R., Holsten, R. D., and Jackson, Ε. K. (1971).
Biological nitrogen fixation, a key to world protein. Plant Soil, Spec. Vol. pp. 561-590.
Hawkswoth, D. L., and Rose, F. (1970). Qualitative scale for estimating sulphur dioxide air pollution in England Wales using epiphytic lichens. Nature (London) 227,145-148.
Henckel, P. A. (1938). On the lichen symbiosis. Bull. Soc. Natur. Moscou, Sect. Biol. [N.S.]
47, 13.
Henckel, P. Α., and Yuzhakova, L. A. (1936). On the role of Azotobacter in the lichen symbiosis.
Bull. Inst. Rech. Biol. Perm. 10, 315-325.
Henriksson, E. (1951). Nitrogen fixation by a bacteria-free symbiotic Nostoc strain isolated from Collema. Physiol. Plant. 4, 542-545.
Henriksson, E. (1957). Studies in the physiology of the lichen Collema. 1. The production of extracellular nitrogenous substances by the algal partner under various conditions. Physiol.
Plant. 10, 943-948.
Henriksson, E. (1958). Studies on the physiology of the lichen Co/terna. II. A preliminary report on the isolated fungal partner with special regard to its behaviour when growing together with the symbiotic alga. Sv. Bot. Tidskr. 52, 391-396.
Henriksson, E. (1960). Studies in the physiology of the lichen Collema. III. The occurence of an inhibitory action of the phycobiont on the growth of the mycobiont. Physiol. Plant. 13, 751-754.
Henriksson, E. (1961). Studies in the physiology of the lichen Collema. IV. The occurrence of polysaccharides and some vitamins outside the cells of the phycobiont, Nostoc sp. Physiol.
Plant. 14, 813-817.
Henriksson, E., and Simu, B. (1971). Nitrogen fixation by lichens. Oikos 22, 119-121.
Hitch, C. J. B. (1971). A study of some environmental factors affecting nitrogenase activity in lichens. M. Sc. Thesis, University of Dundee.
Iskina, R. Y. (1938). On nitrogen fixing bacteria in lichens. Bull. Inst. Rech. Biol. Perm. 11,133.
Kershaw, Κ. Α., and Millbank, J. W. (1969). A controlled environment lichen growth chamber.
Lichenologist 4, 83-87.
Kershaw, Κ. Α., and Millbank, J. W. (1970). Nitrogen metabolism in lichens. II. The partition of cephalodial-fixed nitrogen between the mycobionts and phycobiont of Peltigera aphthosa.
New Phytol. 69, 75-79.
Krasilnikov, N. A. (1949). Is Azobacter present in lichens? Mikrobiologiya 18, 3.
Kratz, W. Α., and Myers, J. (1955). Nutrition and growth of several blue-green algae. Amer.
J. Bot. 42, 282-287.
Lange, O. L., Schulze, E. D., and Koch, W. (1970). Experimentell-okologische Untersuchun
gen an Flechten der Negev-Wuste II. C 02 gaswechsel und Wasserhaushalt von Ramalina maciformis (Del.) Bory am naturlichen Standort wahrend der sommerlichen Trocken- periode. Flora 159, 38-62.
Masse, L. C. (1966). Etude comparee des teneurs en azote des lichens et de leurs substrate: Les especes ornithocoprophiles. CR. Acad. Sci., Ser. D 262, 1721-1724.
Masse, L. C. (1969). Quelques aspects de l'uricolyse enzymatique chez les lichens. CR. Acad.
Sci., Ser. D. 268, 2896-2898.
Millbank, J. W. (1972). Nitrogen metabolism in lichens. IV. The nitrogenase activity of the Nostoc phycobiont in Peltigera canina. New Phytol. 71, 1-10.
Millbank, J. W., and Kershaw, K. A. (1969). Nitrogen metabolism in lichens. I. Nitrogen fixation in the cephalodia of Peltigera aphthosa. New Phytol. 68, 721-729.
Millbank, J. W., and Kershaw, K. A. (1970). Nitrogen metabolism in lichens. III. Nitrogen fixation by internal cephalodia of Lobaria pulmonaria. New Phytol. 69, 595-597.
Moissejeva, Ε. N . (1961). "Biochemical Properties of Lichens and Their Practical Importance,"
p. 82. Izd-Akad. Nauk U.S.S.R., Moscow (in Russian with English summary).
9. N I T R O G E N M E T A B O L I S M 307
Panosyan, A. K., and Nikogosyan, K. (1966). The presence of nitrogen fixers in lichens. Biol.
Zh. Arm. 19, 3 - 1 1 .
Peat, A. (1968). Fine structure of the vegetative thallus of the lichen Peltigera polydactyla. Arch.
Mikrobiol. 61, 212-222.
Quispel, A. (1945). The mutual relations between algae and fungi in lichens. Rec. Trav.Bot.
Neer. 4 0 , 4 1 3 - 5 4 1 .
Ramakrishnan, S., and Subramanian, S. S. (1964a). Amino acids of Roccella montagnei and Palmelia tinctorum. Indian J. Chem. 2, 467.
Ramakrishnan, S., and Subramanian, S. S. (1964b). Amino acids of Peltigera canina. Curr. Sci.
33, 522.
Ramakrishnan, S., and Subramanian, S S. (1965). Amino acids composition of Cladonia rangiferina, Cladonia gracilis and Lobaria isidiosa. Curr. Sci. 34, 345-347.
Ramakrishnan, S., and Subramanian, S. S. (1966a). Amino acids of Lobaria subisidiosa, Umbilicaria pustulata, Parmelia nepalensis and Ramalina sinensis. Curr. Sci. 35, 124.
Ramakrishnan, S., Subramanian, S. S. (1966b). Amino acids of Dermatocarpon moulinsii. Curr.
Sci. 35, 284.
Rogers, R. W., Lange, R. T., and Nicholas, D. J. D . (1966). Nitrogen fixation by lichens of arid soil crusts. Nature (London) 209, 96-97.
Scholander, P. F., Flagg, W., Walters, V., and Irving, L. (1952). Respiration in some arctic and tropical lichens in relation to temperature. Amer. J. Bot. 39, 707-713.
Scott, G. D . (1956). Further investigations of some lichens for fixation of nitrogen. New Phytol.
55, 111-116.
Scott, G. D. (1969). "Plant Symbiosis." Arnold, London.
Shields, L. M., Mitchell, C , and Drovet, F. (1957). Alga- and lichen-stabilised crusts as soil nitrogen sources. Amer. J. Bot. 44, 489^498.
Smith, D . C. (1960a). Studies in the physiology of lichens. I. The effects of starvation and of ammonia absorption upon the nitrogen content of Peltigera polydactyla. Ann. Bot. (London) [N.S.] 24, 52-62.
Smith, D. C. (1960b). Studies in the physiology of lichens. 2. Absorption and utilisation of some simple organic nitrogen compounds by Peltigera polydactyla. Ann. Bot. (London) [N.S.]
24, 172-185.
Smith, D . C. (1961). The physiology of Peltigera polydactyla (Neck.) Hoffm. Lichenologist 1, 209-226.
Stewart, W. D. P. (1965). Nitrogen turnover in marine and brackish habitats. 1. Nitrogen fixation. Ann. Bot. (London) [N.S.] 29, 229-239.
Stewart, W. D. P. (1966). "Nitrogen Fixation in Plants". Athlone Press, London.
Stewart, W. D. P. (1970). Algal fixation of atmospheric nitrogen. Plant Soil 32, 555-588.
Stewart, W. D. P. (1971). Physiological studies on nitrogen fixation by algae. Plant Soil, Spec.
Vol. pp. 145-160.
van Gorkom, H. J., and Donze, M. (1971). Localization of nitrogen fixation in Anabaena. Nature (London) 234, 231-232.
Watanabe, Α., and Kiyohara, T. (1963). Symbiotic blue-green algae of lichens, liverworts and cycads. In "Studies on Microalgae and Photosynthetic Bacteria." Japanese Society of Plant Physiologists, pp. 189-196. Univ. of Tokyo Press, Tokyo.