Rigor Mortis in Fish
E. AMLACHER
Institut für Fischerei D. A . L , Berlin-Friedrichshagen, Germany
I. General Comments 385 II. The Process of Rigor Mortis 386
A. Chemical Changes in the Muscle Tissue 386
B. Assessing Rigor Mortis 389 C. The Hydrogen Ion Concentration 397
III. Rigor and Keeping Properties of Fish Flesh 404
References 406
I. General Comments
One of the factors which for a certain lapse of time retards the post
mortem autolytic and bacterial decomposition of the flesh and its protein is the death-stiffening of the muscle tissue—rigor mortis. This process has been subject to many investigations and has been most thoroughly studied in slaughter animals. Rigor mortis in fish generally has a shorter duration than in mammals. It starts 1-7 hr. after death. Its peak in slaughtered fish, kept in ice, lies between 5 and 22 hr. after death. The total duration of the rigor covers 30-120 hr. Suffocated fishes not held in ice show a shorter period of muscle toughening (32-93 hr.). A prolonga
tion of the rigor mortis period, consequently, is of great economic im
portance.
Rigor mortis lasts longer when the fish has exerted less muscular activity prior to death (Amano et al., 1953; Fujimaki and Kojö, 1953;
Noguchi and Yamamoto, 1955b), and is refrigerated immediately (Part- mann, 1954b; Noguchi and Yamamoto, 1955b). Handling prior to and during rigor mortis should be held at a minimum. This implies that fish should be handled carefully during the actual catching. The dragging time in the nets should be made as brief as conditions allow. On board, the fish should be thrown around and bruised as little as possible. Ac
cording to recent concepts, anesthesizing of fish in the net or on board through electroshock is desirable (Kreutzer, 1950; Meyer, 1951). Ac
cording to Ewart (1887), such electric impulses lead to an immediate
"cataleptic" rigor. Flesh from fish that have been caught and handled in 385
a cautious way keeps longer and constitutes a better raw material for the processing industry than flesh from exhausted animals. Similar ob
servations have been made on fresh-water fish (Pohl, 1953).
The general phenomenon of rigor mortis has been reviewed by Bate- Smith (1948) and Whitaker (1959), and the special problems of fish have been discussed in two surveying papers by Reay and Shewan (1949) and Partmann (1954a).
II. The Process of Rigor Mortis
A . CHEMICAL CHANGES IN THE MUSCLE TISSUE
The stiffening effect accompanying rigor mortis is due basically to a contraction of the transverse skeletal muscles. This also takes place in the sinews, thus affecting connective tissue. Biochemical and physiologic processes in the dying muscle are responsible for this phenomenon. There are many theories regarding the character of rigor mortis. Riesser (1925) maintains that rigor mortis constitutes a damage of the colloidal nature of muscle proteins effected by the termination of the normal metabolism of tissue and reflected primarily in an acidification. Flückiger (1937) holds the interruption in the oxidation processes of the cells to be responsible. The formation of excessive lactic acid causes a swelling of the muscle fibrils, which in turn leads to the toughening. Embden (1925) was able to prove that, in addition to lactic acid, phosphoric acid also is formed from "lactacidogen" which was supposed to be a transition product from the glycogen breakdown. This phosphoric acid induces a rapid contraction of the muscles, while the lactic acid chiefly controls the duration of the contracted condition. According to Riesser (1925), additional metabolic disturbances in the dying muscle, besides the post
mortem accumulation of lactic acid, lead to cell disorganization and thus to rigor mortis. As regards the stiffening per se, this is generally attributed to the two major proteins, actin and myosin, combining to form acto
myosin. How this happens is still a mystery, but at the same time the sol
ubility of the protein in sodium chloride solutions is noticeably reduced.
The most comprehensive study on the biochemistry of muscle con
traction, including that in rigor mortis, was carried out by Szent-Györgyi.
A summary of his investigations through many years was published in 1945. Guba (1943) was able to confirm that in all essentials fish pro
tein followed the same pattern. Largely the findings of Szent-Györgyi are still valid, although certain modifications have been made as to
details. Α modern treatise covering this field is a series of lectures given by Dubuisson in 1954. The only new fact of any importance to the specific understanding of fish is the present likelihood that actomyosin might not be the compound responsible for the stiffening in rigor mortis.
Contraction (in rabbit) and Y-protein (in frogs) also appear to be characteristic for the contracted stage.
The present concept of the biochemical processes which lead to rigor mortis can be summarized as follows.
Subsequent to the termination of the heart activities and of the body control by the brain, the tissues are no longer provided with oxygen.
Glycolysis then appears in the muscle tissue. As a consequence of these changes, the pH value falls. Furthermore creatine-phosphate ( C P ) — a reservoir of phosphate-bound energy—normally present in the muscle disappears. A resynthesis is no longer possible. After losing about 6 0 % of the CP, a breakdown of ATP starts, which, in the beginning, is re
versible (Bendall, 1951), for adenylic acid is formed, which reacts with phosphoric acid to re-form ATP. When this process ends, adenylic acid breaks down further to inosinic acid. The phosphoric acid for the afore
mentioned reaction comes from the hydrolysis of CP. Findings on other types of meat than fish, however, point to the clear possibility that no such synthesis takes place. Instead a special enzyme, myokinase, acts as a retarding agent and suppresses the breakdown of ATP. At any rate, the more the ATP content of the muscle declines, the more profound the biochemical changes become. In warm-blooded animals, mechanical firmness of the muscle increases through a contraction and transition into the stage of rigor mortis. The loss of extensibility is indirectly re
lated to the falling ATP level as compared to the resting living muscle.
(Bate-Smith and Bendall, 1947). Consequently, this is not, as was earlier assumed, due to the fall in the pH value through the formation of lactic acid.
The real cause for rigor mortis is the disappearance of ATP, as Erdös (1943) and, independently of him, Bate-Smith (1948) and Bendall (1951) proved through extensive investigations. The decline of ATP in mackerel and red snapper was studied by Noguchi and Yamamoto
(1955c). No difference was found to exist in the rate of this breakdown between these two species—ATP fell to almost zero within 6 to 8 hr. at 25 °C. in carp muscle, the ATPase activity falling off at a much slower rate (Saito and Hidaki, 1956). Breakdown of both ATP and ADP in carp was studied by Saito and Arai (1957) in a wide range of temper-
atures. Asakawa (1957) found, too, that the disappearance of ATP co
incides with the onset of rigor mortis in carp and crucian carp. Further, he describes the stepwise decline in electrical resistance as taking place in six stages, the first three of which correspond to the prerigor, rigor, and immediate postrigor periods respectively. In fish the ATP breakdown is, on the whole, a far more rapid process than in whales and in ter
restrial animals (Partmann, 1954). The softening effect of ATP is gen
erally attributed to this strong binding of calcium ions which otherwise are released and accelerate contraction.
There is a close relationship between the pH value and the electric resistance of the muscle. The latter decreases with the fall of the hydro
gen ion concentration, caused by the higher permeability of the muscle tissue to free ions. From this stage on, the glycolysis slows down until all glycogen is consumed or the glycolytic enzyme system becomes in
activated at pH 5.4 (Bate-Smith, 1948). Glycogen losses accompanying rigor were also established by Amano et al. (1953), Yoshimura et al.
(1953), and Noguchi and Yamamoto (1955a, b ) . In warm-blooded ani
mals, the length of the prerigor period is indirectly related to the mag
nitude of the glycogen reserve. As long as the glycogen supply lasts, anaerobic glycolysis continues—1.5 moles of ATP being formed from ADP for every mole of lactic acid arising from glycogen (Bate-Smith and Bendall, 1949). Largely, this applies to conditions in fish although Sharp
(1934) found that in the fish muscle a limiting hydrogen ion concentra
tion may be attained which puts a stop to both a further production of lactic acid and a breakdown of glycogen. The rigor appearing after the defrosting of frozen fresh fish tissue is basically of another nature. This so-called "thaw-rigor" will be discussed in Chapter 9, Volume II.
The isoelectric point of myosin is also located at pH 5.4, and at this stage the muscle fibrils begin to contract. Simultaneously, the proteins lose their capacity to bind water. These processes gradually become more evident with the increasing acidity developing during the course of rigor. Profound but partly reversible changes are also noticeable in the amount of free SH groups in the flesh (Noguchi and Yamamoto, 1955a, b; Saito and Hidaka, 1956). The number of free SH groups drops as rigor develops, but when the stiffening is resolved, it starts increasing again (Noguchi and Yamamoto, 1955a, b; Saito and Hidaki, 1956). This latter change is attributed to a denaturation of the protein following the termination of rigor (Noguchi and Yamamoto, 1955a) and the onset of spoilage, or both together (Saito and Hidaka, 1956). Certain ions play
a key role in rigor. The myokinase is only active in the presence of magnesium ions, but is inactivated by the related calcium ions. One essential feature in the postmortem changes is the gradual release of calcium ions, normally tightly bound to ATP. Zinc ions exert the same effect as calcium.
The resolution of rigor is seen as connected with the accumulation of lactic acid gradually bringing about the vanishing of all stiffness (Riesser, 1925). Besides, autolytic changes take place—which the layman inter
prets as a tenderizing of the flesh (Steiner, 1939)—after rigor proteolysis sets in. An indication of these changes is, in the final stages, the appear
ance of ammonia (Schwarzfischer, 1949). In selachians, ammonia is readily formed through a bacterial breakdown of urea, but no ammonia appears until rigor has receded (Sebastio and Mercuric, 1956). For analytical figures showing the magnitude of these various changes, ref
erence is made to Partmann (1954b).
It is commonly believed that dark-meat fish spoils more readily than white-meat fish. This has been attributed to differences in the duration of rigor mortis. There is no experimental evidence to substantiate this assertion. On the contrary, those few studies available indicate that no such basic biochemical differences exist (Noguchi and Yamamoto, 1955b). On the other hand, there is the possibility that dark-meat fish may struggle more strongly prior to death and thus deplete their glyco
gen reserves to a greater extent. Contrary to this reasoning is the finding by Braekkan (1956) that red-muscle tissue of tuna showed a higher content of glycogen and less lactic acid than adjacent white-muscle flesh.
Braekkan assumes that, under the anaerobic conditions prevailing during the death struggle, the changes are less profound in the red muscle than in the neighboring ordinary muscles.
An important finding refers to the mobilization of the glycogen. Gen
erally, it has been believed that this took place exclusively through phos
phorylation, but recent Soviet studies report that not only do rabbit and beef muscle contain α-amylases capable of attacking the glycogen, but this is also true of the fish muscle (Andreev, 1958).
B . ASSESSING RIGOR MORTIS
As mentioned above, the formation of lactic acid is, in effect, not the direct cause of rigor mortis. The acid reaction thus created, however, is of great importance for increasing the duration of rigor and for sup
pressing the development of microorganisms causing spoilage.
The external evidence of rigor is a stiffening of the entire body, due primarily to contractions of the transverse, as well as the smooth, muscles and the sinews. The more glycogen the muscle tissue contains at death, the later rigor mortis sets in, and the longer does it last. The metabolic processes in the muscle tissue are in most cases accelerated through intense muscular activity of the fish prior to death, causing a decrease in the content of glycogen reserves. Thereby, the duration of rigor is reduced and it is initiated earlier. This applies particularly to fish caught in dragging nets, through which they become physically exhausted. If, in addition, such fish have a long death struggle on board, the duration of rigor is reduced still more. The mechanical strain on the muscular tissue caused by throwing the fish around, as well as high storage tem
peratures, also influence unfavorably the extension of the toughening.
1. Measuring Toughness
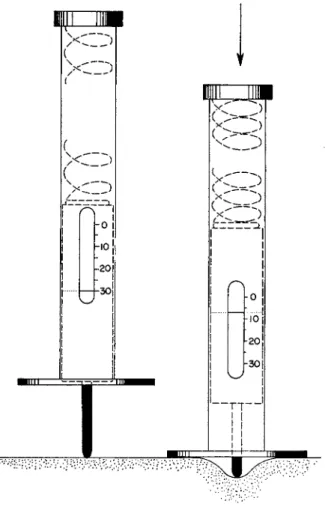
Several methods have been developed for measuring the toughening that takes place during rigor mortis. Most of these have been applied to isolated flesh samples. As the major part of fresh fish arrives on the market as whole animals, only those methods applicable to whole fish deserve attention. Messtorff (1954a) developed an apparatus that made it possible to carry out such evaluations on board without too many accessories. The principles of this method were based on experiments by Mangold (1920). The way this piece of apparatus operates is pre
sented in Fig. 1. A blunt-ended piston is pushed into the flesh by ap
plying a constant pressure transferred via a compression spring. The depth of the imprint is measured through a small, glass-covered opening.
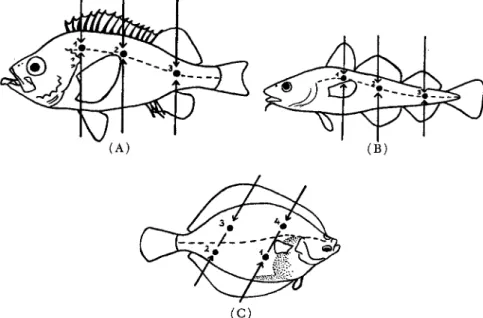
The pressure exerted does not bring about any changes in the mechanical conditions of the muscle of such a nature as to influence negatively the course of rigor. Figure 1 shows the procedure followed in measuring the toughness of fish flesh. The portion selected for measuring is the caudal muscles.
With codfish and redfish, measurements were made on the left side, with flatfish, on the eye side. On roundfish three, and on flatfish four, locations were selected for measurements. On the basis of these data, an average for the total of the caudal muscles was calculated. Figure 2 shows the localization of these points of measurement.
An entirely different approach has been employed by Japanese sci
entists. They measure at each step the remaining capacity of contraction by submitting muscle samples to water perfusion, inducing a contraction,
the so-called "arai phenomenon." The degree of contraction obtained naturally is greatest prerigor, and the ability to contract disappears during rigor, eventually reaching a zero value. These individual percent-
> < r ; )
ho 10 20
3 0 |
I
IEZJ1
<«--=
v )L L
- 0
~-ib
- 2 0 - 3 0
TT"
Fish muscle
FIG. 1. Measuring of the muscle stiffness (after Messtorff, 1954a). First the device is applied vertically to the muscle surface. Then the imprint is measured: the house is pressed down until the hmiting plate at the lower end touches the body surface.
age values are plotted in a time diagram (Noguchi and Yamamoto, 1955a, b ) . The advantage with this procedure is the possibility of obtaining reproducible, accurate values; the difficulty is to relate these figures to
various stages of rigor mortis. Whether the peak of rigor coincides with the complete disappearance of contractibility is still an open question.
2. Initiation, Course, and Termination of Rigor
The procedures described above are based primarily on the methods employed by Messtorff (1954a, b ) and Pohl (1953). Messtorff used for his experiments fish located in the surface areas of the nets and still showing vigorous signs of life. The study began when the nets were
( C )
FIG. 2. Localization of the measuring points in ( A ) redfish, ( B ) cod, ( C ) plaice (after Messtorff, 1954a).
emptied, at the moment when the fish were taken from their natural environment. Pohl chose as a starting point for her studies the occurrence of death, defined as the moment after which independent movements, breathing, and eye reflexes could no longer be seen (so-called partial death). Messtorff (1954a) made his measurements on series of 10 to 15 fish of the same species and of the same size. As a representative value for the degree of imprint and, consequently, for the toughness of the muscles of a single specimen, the average of three to four measurements from different locations on the body was employed.
Because the degree of toughness changes from the initiation of rigor
to its peak (Fig. 3 ) , difficulties exist in comparing the qualitative assess
ments of toughness. Messtorff (1954a, b ) developed the following proce
dure. He computed all values percentagewise, giving the starting value as 100. These relative figures were entered upon a diagram upon which maximum and minimum points, as well as the general course of the rigor, can be traced. Rigor is considered terminated when this curve returns to the point of the initial value (Figs. 4 and 5 ) . This method gave repro
ducible results and allowed statistical evaluation of reliability.
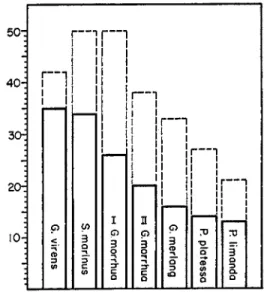
50d
40H
FIG. 3. Differences in level between minimum and maximum values in measure
ments of muscle stiffness during rigor mortis. The bars drawn with solid lines in
dicate the average amplitude of the measurements; the broken lines give the maxi
mum values obtained (Messtorff, 1954b). G. means Gadus; S., Sebastes; P., Pleuro- nectes.
Rigor in fish follows the pattern for mammals appearing in so-called
"descending" order. This means the stiffening starts in the lower jaw and around the gill-cover (operculum), which in many fish, e.g., cod and whiting, is visible through a wide opening of the mouth and of these covers. Further rigor spreads from the head via the chief body muscles to the caudal area. In this final stage the muscles of the fins also stiffen.
In many cases it has been observed that the caudal muscles go into rigor first, which several authors (Messtorff, 1954a, b; Weiss, 1914; Pohl, 1953) attribute to the fact that these muscles are particularly active in the death struggle. The resolution of rigor follows the same general rule,
starting from the head region and spreading to the tail part. This latter sequence was confirmed by Mori (1954) for such Mediterranean fish as mackerel, anchovy, garfish, gray mullet, umbrine, sea bream, and sole when rigor resolves. But in all these cases the onset takes place simul
taneously over the entire body. These differences as to onset and resolu-
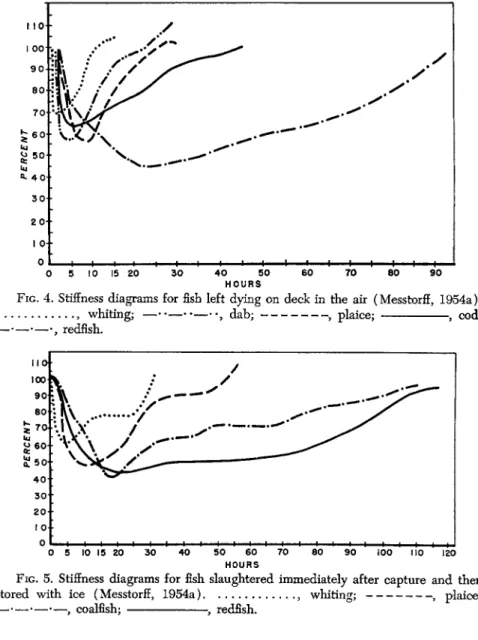
FIG. 4. Stiffness diagrams for fish left dying on deck in the air (Messtorff, 1954a).
, whiting; — · · — · · — · · , dab; , plaice; , cod;
• — · — · , redfish.
0 5 10 15 20
50 60
no
HOURS
FIG. 5. Stiffness diagrams for fish slaughtered immediately after capture and then stored with ice (Messtorff, 1954a) , whiting; , plaice;
— . — . — . —9 coalfish; , redfish.
tion of rigor might explain variations found with regard to pH in dif
ferent parts of the fish body—studied for albacore by Kawabata et al.
(1952).
The rigor diagrams all show a fairly uniform course, even under dif
ferent experimental conditions (Messtorff, 1954a,b). The maximum of the rigor does, however, appear at different times depending upon species and experimental conditions (Figs. 4 and 5 ) . The peak of rigor was reached most rapidly in whiting, i.e., after 1 hr.; the degree of imprint by the measuring device was then only 6 8 % of the initial value. The longest period for the full development of rigor was 22 hr., observed in redfish. In this case the imprint in the muscles amounted to 4 3 % of the initial value. The time until rigor mortis terminated in redfish was 98 hr., in whiting 9 hr. The ending of rigor is, in general, influenced by the external temperature and is, therefore, irregular. There may also be seasonal differences (Kawabata et al., 1952).
The duration of rigor depends on environmental temperatures and the type of death of the fish. Low temperatures prolong rigor consider
ably. When fish are slaughtered immediately after being caught, rigor is prolonged, although not to the same extent as through the lowering of the temperature (Messtorff, 1954a, b; Gianelli, 1954; Pavlov, 1956).
Gianelli (1954), however, reports that sole show a reverse reaction in the initiation of rigor, 30 min. at 2 to 3° C. and 3-8 hr. at 23 to 24°C.
Similar findings by Braccio refer to mackerel (1954). Flatfish, as a whole, show a more extensive rigor than roundfish (Reay, 1952; Gianelli, 1954).
In this case rigor develops more slowly but, on the other hand, becomes more intense. This is evident from the imprint values reaching lower readings.
The question of the effect of temperature is still very obscure. The finding that a lowering of the temperature has profound influence on the duration of rigor is in direct conflict with the experimental studies on the apyrase activity of fish muscle tissues. The Q10 values are low for fish (1.2 to 1.7). Four species were studied by Burr Steinbach (1949).
This is also in biochemical accordance with the fact that fish are cold
blooded animals. When Cutting's findings (1938) are analyzed, it is obvious that temperature has less marked influence on the onset of rigor, but some effect on the duration.
The following relationships seem to apply to fish rigor:
1. The intensity of rigor in various fish species becomes greater the longer rigor lasts, ceteris paribus (c.p.)—this expression meaning: given
that the experimental conditions in other respects are identical (Mess
torff, 1954b).
2. In fish of the same species which die in the same way, the dura
tion and intensity of rigor increase with a lowering of the body temper
ature, c.p. (Caracciolo, 1954; Gianelli, 1954; Messtorff, 1954b; Pavlov, 1956).
3. Slaughtered fish show a later onset of rigor than do asphyxiated fish, c.p. Rigor also becomes more intense and lasts longer.
4. For fish belonging to the same species and presumably of the same biotypes (strains) and the same size, the duration of rigor is equally long, c.p.
5. Fish of varying size and age show strong differences in this respect.
In small-sized fish, rigor may last longer than in large-sized specimens, c.p. Messtorff explains these differences as caused by variations in their metabolism due to their different way of life. Thus, mackerel, as ex
tremely active fish with a high metabolism, have an immediate but brief rigor.
Soviet findings indicate that the initiation of rigor is particularly influenced by body size. In Pacific flatfish, small-sized specimens stiffen after 40 to 60 min. while large ones show such symptoms only after 4 to 5 hr. (Meljnikova, 1958). A number of data referring to rigor mortis, its initiation, duration, and other characteristics are summarized in Table I. Messtorff (1954b) gives the following values and their range for slaughtered marine fish put on ice: lag in initiation, 1-7 hr.; peak of rigor, 5-22 hr. after slaughter; total duration of the rigor period: 3 1 - 120 hr.
With regard to fresh-water fish, reference is made to the studies by Pohl (1953). These studies took place under experimental conditions differing from those of Messtorff (1954a, b ) . For this reason, comparisons are justified only in a restricted way. Pohl (1953) chose a different ref
erence point for death; in fish killed by beheading at a water temper
ature of 1 6 ° C , the total duration of rigor mortis was about 21 hr. in lake perch and about
23y
2 hr. in roach. At the same temperature level, rigor mortis lasted 30-32 hr. in fish which were killed by hitting on the head.Asphyxiated fish had a total rigor period of 21 y2 hr. (lake perch) and
25yi
hr. (roach). Fish killed by electroshock had a shorter rigor period than those killed by slaughter or asphyxiation. In extreme cases it was shortened by 21 to 23 hr., as compared to those killed by beheading.Above 20°C. and below 5 ° C , differences in length of rigor became less
influenced by the method of killing. Sardines show a tremendous varia
tion in the duration of rigor, 1.5 to 4 hr. for fish held at 2 5 ° C , up to 50 hr.
for iced fish (Caracciolo, 1954). Pohl (1953), too, reaches the conclusion that the size and age of fish have an obvious influence on the onset and duration of rigor (see Table I under lake perch).
Temperature influences the development of rigor in fresh-water fish in the following way (Pohl, 1953). At 0 ° C , rigor starts about 24 hr. after death in roach, redeye, and perch and lasts for 72 to 80 hr. At 35°C. in the same species, it begins 20-25 min. postmortem and has an average duration of 3 hr. These results were obtained with fish of varying length (14-22 c m ) . Body size has an influence, too, causing certain deviations from the average. Caracciolo and Silveri (1956) found that slaughtered tench specimens had a more extensive rigor than asphyxiated ones. In diseased roaches (attacked by Ligula simplicissima), rigor sets in earlier, develops weakly, and is of very short duration (Pohl, 1953).
The above-mentioned experimental difficulty of establishing a de
fined starting point becomes still more evident when comparing fish with varying capacity for survival in air. Some fresh-water fish, such as carp and tench, may stay alive out of water for hours if kept humid. This presumably explains the fact that observations as to rigor in tench are at great variance (Sebastio and Mercurio, 1956; Caracciolo and Silveri, 1956). Sebastio even maintains that the contraction of the muscles takes place prior to rigor. It was also suggested by these research workers that tench from different lake regions reacted differently.
Sharks and rays seem to exhibit a much shorter rigor than teleosteans (Table I I ) , but so far no studies of the temperature effect are available.
All published figures refer to 26°C. (Sebastio and Mercurio, 1956).
C. T H E HYDROGEN ION CONCENTRATION
Rigor in fish also runs concomitant with an acidification of the muscle tissue, caused by the appearance of lactic acid (Benson, 1928b; Macpher- son, 1930, 1932). This has definite beneficial bactericidal effects. Fish species with a more acid tissue, e.g., halibut (pH 5.6) have good storage quality.
Numerous postmortem measurements of pH in fish muscle have been published through the years (Ewart, 1887; Anderson, 1907; Benson, 1928; Macpherson, 1932; Cutting, 1939; Hjort-Hansen, 1943). Generally, newly killed fish show a slightly alkaline reaction (7.05-7.35), unless exhausted through struggling. The ultimate values reported are, as a
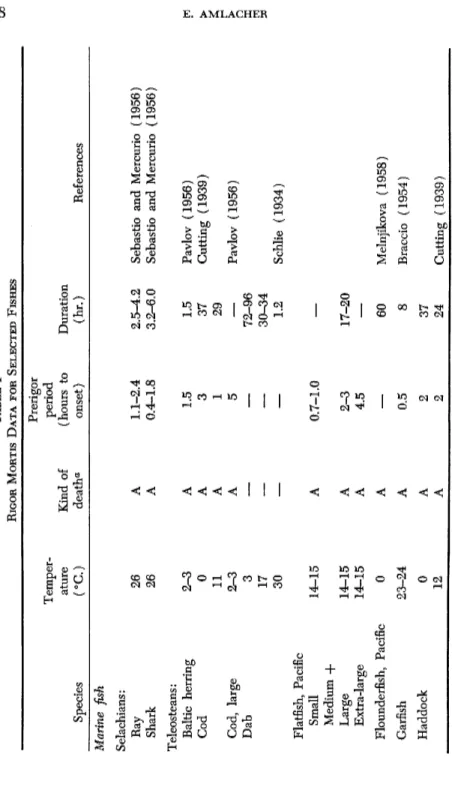
TABLE I RIGOR MORTIS DATA FOR SELECTED FISHES Prerigor Temperperiod ature Kind of (hours to Duration Species (°C) deatha onset) (hr.) References Marine fish Selachians: Ray 26 A 1.1-2.4 2.5-4.2 Sebastio and Mercurio (1956) Shark 26 A 0.4-1.8 3.2-6.0 Sebastio and Mercurio (1956) Teleosteans: Baltic herring 2-3 A 1.5 1.5 Pavlov (1956) Cod 0 A 3 37 Cutting (1939) 11 A 1 29 Cod, large 2-3 A 5
—
Pavlov (1956) Dab 3— —
72-96 17 ——
30-34 30— —
1.2 Schlie (1934) Flatfish, Pacific Small 14-15 A 0.7-1.0 Medium -f Large 14-15 A 2-3 17-20 Extra-large 14-15 A 4.5—
Flounderfish, Pacific 0 A—
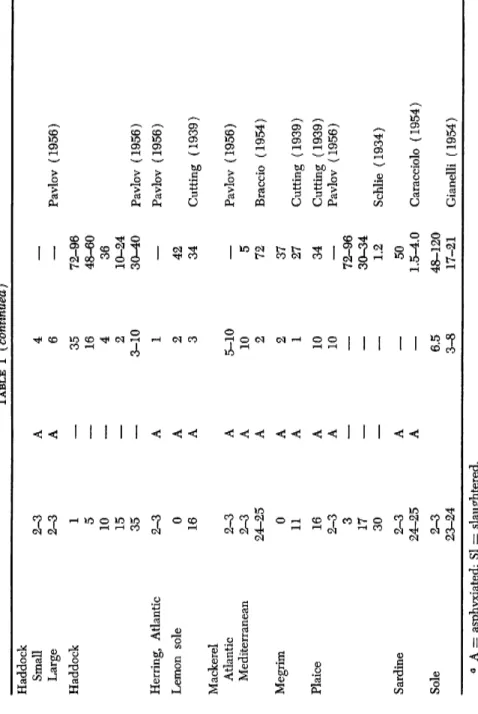
60 Melnjikova (1958) Garfish 23-24 A 0.5 8 Braccio (1954) Haddock 0 A 2 37 12 A 2 24 Cutting (1939)TABLE I (continued) Haddock Small 2-3 A 4 — Large 2-3 A 6 — Pavlov (1956) Haddock 1 — 35 72-96 5 — 16 48-60 10 — 4 36 15 — 2 10-24 35 — 3-10 30-40 Pavlov (1956) Herring, Atlantic 2-3 A 1 — Pavlov (1956) Lemon sole 0 A 2 42 16 A 3 34 Cutting (1939 Mackerel Atlantic 2-3 A 5-10 — Pavlov (1956) Mediterranean 2-3 A 10 5 Pavlov (1956) 24-25 A 2 72 Braccio (1954 Megrim 0 A 2 37 11 A 1 27 Cutting (1939 Plaice 16 A 10 34 Cutting (1939 2-3 A 10 — Pavlov (1956) 3 — — 72-96 17 — — 30-34 30 — — 1.2 Schlie (1934) Sardine 2-3 A 50 24-25 A — 1.5-4.0 Caracciolo (1954 Sole 2-3 6.5 48-120 23-24 3-8 17-21 Gianelli (1954 α A = asphyxiated; Si =: slaughtered.
TABLE I (continued) Species
Temper ature (°C.) Kind of death*
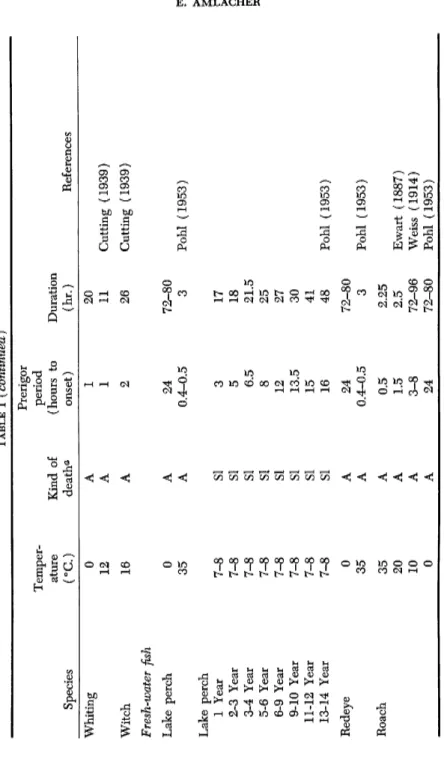
Prerigor period (hours to onset) Duration (hr.) Reference Whiting 0 A 1 20 12 A 1 11 Cutting (1939) Witch 16 A 2 26 Cutting (1939) Fresh-water fish Lake perch 0 A 24 72-80 Lake perch 35 A 0.4-0.5 3 Pohl (1953) Lake perch 1 Year 7-8 SI 3 17 2-3 Year 7-8 SI 5 18 3-4 Year 7-8 SI 6.5 21.5 5-6 Year 7-8 SI 8 25 6-9 Year 7-8 SI 12 27 9-10 Year 7-8 SI 13.5 30 11-12 Year 7-8 SI 15 41 13-14 Year 7-8 SI 16 48 Pohl (1953) Redeye 0 A 24 72-80 35 A 0.4-0.5 3 Pohl (1953) Roach 35 A 0.5 2.25 20 A 1.5 2.5 Ewart (1887) 10 A 3-8 72-96 Weiss (1914) 0 A 24 72-80 Pohl (1953)
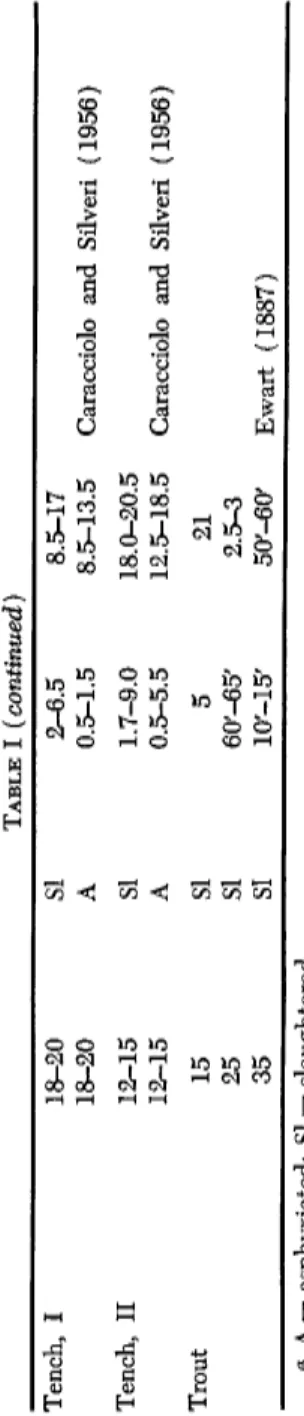
TABLE I (continued) Tench, I 18-20 SI 2-6.5 8.5-17 18-20 A 0.5-1.5 8.5-13.5 Caracciolo and Silveri (1956 Tench, II 12-15 SI 1.7-9.0 18.0-20.5 12-15 A 0.5-5.5 12.5-18.5 Caracciolo and Silveri (1956 Trout 15 SI 5 21 25 SI 60'-65' 2.5-3 35 SI 10'-15' 50'-60' Ewart (1887) a A = asphyxiated; SI = slaughtered.
TABLE II DURATION OF RIGOR MORTIS IN EXPERIMENTS WITH MARINE FISH« Imprint Stiffening Time for Total value in Kind of Storage period resolution rigor % of figure Species death0 conditions (hr.) (hr.) (hr.) at death Plaice A 16-17°C. 9 17 26 56 SI 16-17°C. 14 17-18 31-32 49 A Ice 7 48 55 38 SI Ice 11 43 54 48 Dab A 16-17°C. 4 17 23 50 SI 16-17°C. 6 17 23 50 A Ice 8 24 32 50 Whiting A 16-17°C. 1 9-11 10-12 68 SI Ice 5 26 31 61 Cod II A 16-17°C. 2-3 29-30 32 60 SI 16-17°C. 4 31 35 54 Cod I A 6-7°C. 5 40 45 62 SI Ice 17 63 80 54 Saithe SI 18 92 110 41 Redfish A 6-7°C. 21 72 93 46 SI Ice 22 98 120 43 a Source: Messtorff, 1954b. * A = asphyxiated; SI = slaughtered.
rule, 6.2-6.5. This is in contrast to mammalian meat, which reaches into the range of pH 5.5. Most flatfish seem to constitute an exception among fish, frequently attaining this more acid level (Cutting, 1939; Hjort- Hansen, 1943). Similar low values were reported by Noguchi and Yama
moto for Japanese mackerel (5.50) and red snapper (5.70). Pohl (1953) made detailed measurements of the pH values in the course of rigor in fresh-water fish. These determinations were made on muscle samples from the dorsal and caudal areas of fish killed by hitting on the head.
When death occurs, the pH value is generally around 7.0 but it may fre
quently be higher or lower. Normally, the pH value change in the way indicated in Figs. 6 and 7. There are, however, many deviations from this general pattern in the three species studied. Figure 7 refers to lake perch; at death the reaction was alkaline in the muscle tissue. But fre
quently the pH value does not exceed 7.0 even when rigor terminates.
Roach generally show values of 6.8 to 6.9 at this stage. White flesh of the frigate mackerel constantly shows lower pH values than dark flesh.
Struggling rapidly lowers pH, and values as low as 5.53 are attained with a corresponding drop in glycogen and an increase in lactic acid
(Amano et al., 1953).
When rigor sets in rapidly—it is then mostly of short duration and less intense—the pH values are lower than when rigor develops at a slow rate and becomes stronger. The pH values of roach and redeye are in general 0.1-0.2 pH units further into the acid area than those of lake perch. The lowest pH values measured by Pohl amounted to 6.4 in roach and 6.5 in lake perch. At the onset of rigor, the maximum value was 7.2 for roach and 7.3 for perch. At the end of rigor, 7.0 was obtained for roach and 7.1 for lake perch.
Measurements by Messtorff (1954a, b ) in various ocean fish species gave the pH values shown in the tabulation.
pH value 0-1 hr. after catch 6.5-6.8 During rigor mortis 6.3-6.7 At the end of rigor 6.1-6.9 9 Days after catch 6.8-7.0
The same general pattern was established in Japanese mackerel and red snapper by Noguchi and Yamamoto (1955a, b ) . Data on the drop in glycogen, the increase in lactic acid, and the corresponding pH changes immediately after death were also furnished by Yoshimura et al.
(1953). An early study was that by Ritchie (1926), who not only dem
onstrated the much higher lactic acid values during and after rigor as compared to those before rigor, but also the fact that the more lively haddock became more acid than the less active hake, with cod in be
tween. Benson (1928) claims that the hake muscle remains alkaline even hours after death.
pH
7 3 7 2 7.1 7.0 6.9 6.8 6.7 6.6 6.5 6.4
>
14 16 ( Β )
FIG. 6. ( A and Β ) The pH values during rigor mortis in roach. R marks onset of rigor. The ordinate gives the pH value, the abscissa the time in hours. Dashed line indicates pH prerigor, and solid line pH during rigor (Pohl, 1953).
III. Rigor and Keeping Properties of Fish Flesh
In summarizing, it may be said that rigor is greatly influenced by environmental factors and not exclusively determined by intrinsic factors.
This is most evident from Figs. 4 and 5. The handling of fish on board and the way of preparation exercise some influence. If the fish are handled carefully and stored in ice, a doubling of the rigor mortis period is possible to obtain. Immediate slaughtering of the fish after capture
30
also extends rigor appreciably. Reduced temperatures affected rigor favorably (see Table I ) .
If the fish species investigated so far are classified according to the onset of rigor and its duration, it becomes evident that these factors are of major importance to the keeping properties. In other words, when rigor is extended in time either through a delay in its onset or by its
pH 7.3 7 2 7.1 7.0 6.9 6 8 67 6 6 6.5 6.4
\
1 0 1 4
(A)
18 20 22 24 26 28 30
pH 7.3 7.2 7.1 7 0 6.9 6.8 6.7 6.6 6.5 6.4
—>>
N T
ft1 0 12 14
( B )
18 20 22 24 26 28 3 0
FIG. 7. (A and Β ) The pH values during rigor mortis in lake perch. R marks onset of rigor. The ordinate gives the pH value, the abscissa the time in hours. Dashed line indicates pH prerigor, and solid line pH during rigor (Pohl, 1953).
duration, this correspondingly defers the attack by the bacteria of spoilage (Lovern, 1952; Messtorff, 1954a). Consequently, rigor in a decisive way predetermines the shelf life of fresh fish. But it also in an intricate way fixes the quality of frozen fish (see Volume I I I , chapter 1 1 ) .
The methods of capture also play an important role via rigor. The more exhausted the fish become before being landed on deck, the less are the glycogen reserves of the body. The more the fish struggle in the
nets, the more they deplete this energy source. In such a case, rigor becomes less accentuated and the time such fish will keep is shortened, even in cold storage. This has been studied experimentally with mackerel, red snapper, and carp. Profound biochemical differences were established as to the rate of ATP breakdown and the disappearance of glycogen.
Exhausted fish, as a whole, showed a rigor period shortened to half the time exhibited by fish beheaded immediately after catch (Noguchi and Yamamoto, 1955b). Similar studies on frigate mackerel were reported by Fujimaki and Kojö (1953) and gave identical results. The ATP break
down measured in the amount of inorganic phosphorus is so much faster in the struggling fish as to render values after 4 hr. twice those of de
capitated fish. Trawl-caught haddock with fatigued muscles consistently showed a more alkaline condition in the tissue as compared to penned fish with rested muscles (Benson, 1928). This was attributed to the lactic acid produced during the struggle. Penned fish with the much higher initial pH values pass into rigor much more slowly (Leim et ah, 1927).
The practical consequences of these findings for the fishing technique are obvious. It is most desirable to curtail within reasonable practical limits the dragging periods. Paralyzing the fish by means of electric shocks immediately after catch is another procedure to be recommended.
REFERENCES
Amano, K., Bito, M., and Kawabata, T. (1953). Handling effect upon biochemical change in the fish muscle immediately after catch. I. Difference of glycolysis in the frigate mackerel killed by various methods. Bull. Japan. Soc. Sei. Fisheries 19(4), 487-498.
Anderson, A. G. (1907). On the decomposition of fish. Fishery Board (Scot.) 26th Ann. Rept. Ill Set. Invest, pp. 13-39.
Andreev, A. K. (1958). Fish muscle amylase. (In Russian.) Biokhimiya 23, 899-903.
Asakawa, S. (1957). A consideration on the post-mortem change of electric re
sistance of fish muscle (preliminary report). /. Bull. Japan. Soc. Sei. Fisheries 22(1), 718-720.
Bate-Smith, E . C. (1948). The physiology and chemistry of rigor mortis, with special reference to the aging of beef. Advances in Food Research 1, 1-38.
Bate-Smith, E. C , and Bendall, J. R. (1947). Rigor mortis and adenosine triphos
phate. /. Physiol. (London) 106, 177-185.
Bate-Smith, E . C , and Bendall, J . R. (1949). Factors determining the time course of rigor mortis. /. Physiol. (London) 110, 47-65.
Bendall, J. R. (1951). The shortening of rabbit muscles during rigor mortis: its relation to the breakdown of adenosine triphosphate and creatine phosphate and to muscular contraction. J. Physiol. (London) 114, 71-88.
Benson, C. C. (1928). Hydrogen ion concentration of fish muscles. J. Biol. Chem.
78, 583-590.
Braccio, G. (1954). Ricerche sul tempo di insorgenza e durata della rigidita cadaverica nel Behne belone a temperatura ambiente di 23-24°C. Atti soc. ital.
sei. vet. 8, 467-469.
Braekkan, O. R. (1956). Function of the red muscle in fish. Nature 178, 747-748.
Burr Steinbach, Η. (1949). Temperature coefficients of muscle apyrase systems.
/. Cellular Comp. Physiol. 33, 123-131.
Caracciolo, S. (1954). Riv. med. vet. e zootec. 6 ( 6 ) , 425-432, quoted in Caracciolo and Silveri (1956).
Caracciolo, S., and Silveri, G. (1956). Ulteriori osservazioni sul 'rigor mortis' della Tinea vulgaris. Atti soc. ital. set. vet. 10, 487-491.
Cutting, C. L. (1939). Immediate post-mortem changes in trawled fish. Ann. Rept.
Food Invest. Board (Gt. BHtain) 1938, 39-40.
Dubuisson, M. (1954). "Muscular Contraction." Charles C Thomas, Springfield, Illinois.
Embden, G. (1925). Chemismus der Muskelkontraktion und Chemie der Musku
latur. In "Handbuch der normalen und pathologischen Physiologie" (A. Bethe, G. von Bergmann, G. Embden, and A. Ellinger, eds.), Vol. 7, pp. 369-400.
Springer, Berlin.
Erdös, T. (1943). Rigor, contracture, and adenosine-triphosphatic acid. (In Hun
garian with German summary.) Studies Inst. Med. Chem. Univ. Szeged 3, 51-56.
Ewart, J. C. (1887). On rigor mortis in fish, and its relation to putrefaction. Proc.
Roy. Soc. 42, 435-450.
Flückiger, G. (1937). Die hauptsächlichsten Veränderungen des Fleisches un
mittelbar nach der Schlachtung. Schweiz. Arch. Tierheilk. 79, 468-475.
Fujimaki, M., and Kojö, K. (1953). Handling effect upon biochemical change in the fish muscle immediately after catch—II. Changes of acid-soluble phosphorus compounds of frigate mackerel muscle. Bull. Japan. Soc. Set. Fisheries 19(4), 499-504.
Gianelli, F . (1954). Ricerche sul tempo di insorgenza e durata della rigiditä cadaverica nella Solea solea in varie condizione di ambiente e temperatura.
Atti soc. ital. set. vet. 8, 464-465.
Guba, F . (1943). Observations on myosin and actomyosin. (In Hungarian with English summary.) Studies Inst. Med. Chem. Univ. Szeged 3, 40-45.
Hjort-Hansen, S. (1943). Studier over den d0de fiskemuskulatur og dens foran- dringer under lagring. II. Fysikalskkjemiske unders0kelser. Rept. Norweg. Fish
eries Research Lab. 1 ( 4 ) .
Kawabata, T., Fujimaki, M., Amano, K., and Tomiya, F . (1952). Studies on pH of fish muscle. Variation in pH of fresh albacore muscle on locality examined (Studies on tunny meat I ) . Bull. Japan. Soc. Sei. Fisheries 18(3), 124-132.
Kreutzer, C. O. (1950). Die physiologischen Grundlagen der Elektrofischerei im Meer. Arch. Fischereiwiss. 2, 10-14.
Leim, A. H., Macleod, J . J . R., and Simpson, W. W. (1927). The relationship between rigor mortis and the glycogen, lactic acid, and free phosphorus of fish muscle. Contribs. Can. Biol, and Fisheries 3 ( 2 0 ) , 457-466.
Lovern, J. A. (1952). Chemistry and advances in fish processing. Fishing News 25, 11.
Macleod, J. J. R., and Simpson, N. W. (1927). The immediate post-mortem changes in fish muscle. Contribs. Can. Biol. and Fisheries [N.S.] 3, 439-456.
Macpherson, N. L. (1930). Post-mortem production of lactic acid and glyco
genosis in fish's muscle. Ann. Rept. Food Invest. Board (Gt. Brit.) 1929, 134.
Macpherson, N. L. (1932). Studies on the behaviour of the carbohydrates and lactic acid of the muscle of the haddock (Gadus aeglefinus) after death. Bio
chem. J. 26, 80-87.
Mangold, E. (1920). Muskelhärtemessungen in Toten- und Wärmestarre. Arch, ges. Physiol. Pflügers 182, 205-214.
Melnjikova, Ο. M. (1958). Technological characteristics in the processing of some species of Pacific flounderfishes. (In Russian.) Rybnoe Khozy. 3 4 ( 6 ) , 58-62.
Messtorff, J. (1954a). Untersuchungen über den Totenstarreverlauf bei Seefischen unter Berücksichtigung seiner Bedeutung für die Haltbarkeit und Qualitätser
haltung. Diplomarbeit, Math. Nat. Fak. Univ. Hamburg, Germany.
Messtorff, J. (1954b). Über den Totenstarreverlauf einiger Seefischarten. Kurze Mitt. Inst. Fischereibiol. Univ. Hamburg. 5, 1-21.
Meyer, P. F. (1951). Erfahrungen mit der elektrischen Thunfischangel. Fischerei
welt 3, 176-178.
Mori, L. (1954). Osservazioni sul modo di insorgenza e scomparsa della rigiditä cadaverica in alcune specie di pesci. Atti soc. ital. sei. vet. 8, 459-461.
Noguchi, E., and Yamamoto, J. (1955a). Studies on the "arai" phenomenon (the muscle contraction caused by perfusing water)—III. Change in the amount of ATP, free SH-group in muscle of fish shortly after the death. Bull. Japan. Soc.
Set. Fisheries 20(11), 1020-1022.
Noguchi, E., and Yamamoto, J. (1955b). Studies on the "arai" phenomenon (the muscle contraction caused by perfusing water)—IV. On the rigor-mortis of white meat fish and of red meat fish. Bull. Japan. Soc. Set. Fisheries 20(11), 1023-1026.
Partmann, W. (1954a). To what extent does fish flesh break down through autol
ysis? Proc. Symposium on Cured and Frozen Fish Technol. Swed. Inst. Food Preserv. Research (Göteborg) Publ. No. 100 (1953) Paper No. 17.
Partmann, W. (1954b). The first post-mortem changes in the muscular tissue of fish. Arch. Fischereiwiss. 5, 159-170.
Pavlov, E. G. (1956). "Refrigeration in Vessels for the Fishing Industry." (In Russian.) Pischtepromizdat, Moscow.
Pohl, I. (1953). Untersuchungen über die Totenstarre an Süsswasserfischen. Ζ.
Fischerei Hilfswiss. [N.S.] 2, 421-449.
Reay, G. A. (1949). The spoilage of fresh fish and its control. Chem. ir Ind.
(London) 1949, 35-39.
Reay, G. Α., and Shewan, J. M. (1949). The spoilage of fish and its preservation by chilling. Advances in Food Research 2, 343-398.
Riesser, O. (1925). Totenstarre der glatten Muskulatur. Arch. ges. Physiol. Pflü
gers 209, 395-413.
Ritchie, A. D. (1926). Lactic acid and rigor mortis in fish muscle. /. Physiol.
(London) 61, IV-V.
Saito, K., and Arai, K. (1957). Studies on the organic phosphates in muscle of aquatic animals III. Effects of storing temperature upon the adenosine poly
phosphate content of carp muscle. Bull. Japan. Soc. Set. Fisheries 2 2 ( 9 ) , 569- 573.
Saito, K., and Hidaka, T. (1956). Studies on the biochemical change in the fish muscle V. On the post-mortem changes in myosin fraction N, ATP, free SH- group and ATP-ase activity of carp muscle. Bull. Japan. Soc. Set. Fisheries 21 ( 9 ) , 1025-1028.
Schlie, K. (1934). Über die Totenstarre bei Seefischen und ihren Zusammenhang mit der beginnenden Zersetzung. Kälte-Ind. 31(10), 115-119.
Schwarzfischer, F. (1949). Chemische Vorgänge bei der Lösung der Totenstarre.
Deut. Z. ges. gerichtl. Med. 39, 421-428.
Sebastio, C , and Mercurio, G. (1956). Prime recerche sulla rigidita cadaverica dei selacci. Atti soc. ital. sei. vet. 10, 491-495.
Sharp, J. G. (1934). Post-mortem breakdown of glycogen and accumulation of lactic acid in fish muscle. Proc. Roy. Soc. B114, 506-512.
Steiner, G. (1939). Die postmortalen Veränderungen des Rindermuskels bei ver
schiedenen Temperaturen, gemessen an seinem mechanischen Verhalten. Arch.
Hyg. u. Bakteriol. 121, 193-208.
Szent-Györgyi, A. (1945). Studies on muscle. Acta Physiol. Scand. 9 ( 2 5 ) , Suppl., 115-180.
Weiss, L. (1914). Die Totenstarre bei Süss wasserfischen und ihre marktpolizei
liche Bedeutung. Z. Fleisch- u. Milchhyg. 25, 33-36.
Whitaker, J. R. (1959). Chemical changes associated with aging of meat with emphasis on the proteins. Advances in Food Research 9, 1-60.
Yoshimura, K., Kabo, S., and Ito, Y. (1953). On the biochemical changes of fish muscle after death ( 1 ) . Bull. Fac. Fisheries Hokkaido Univ. 4 ( 2 ) , 157-162.